
Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
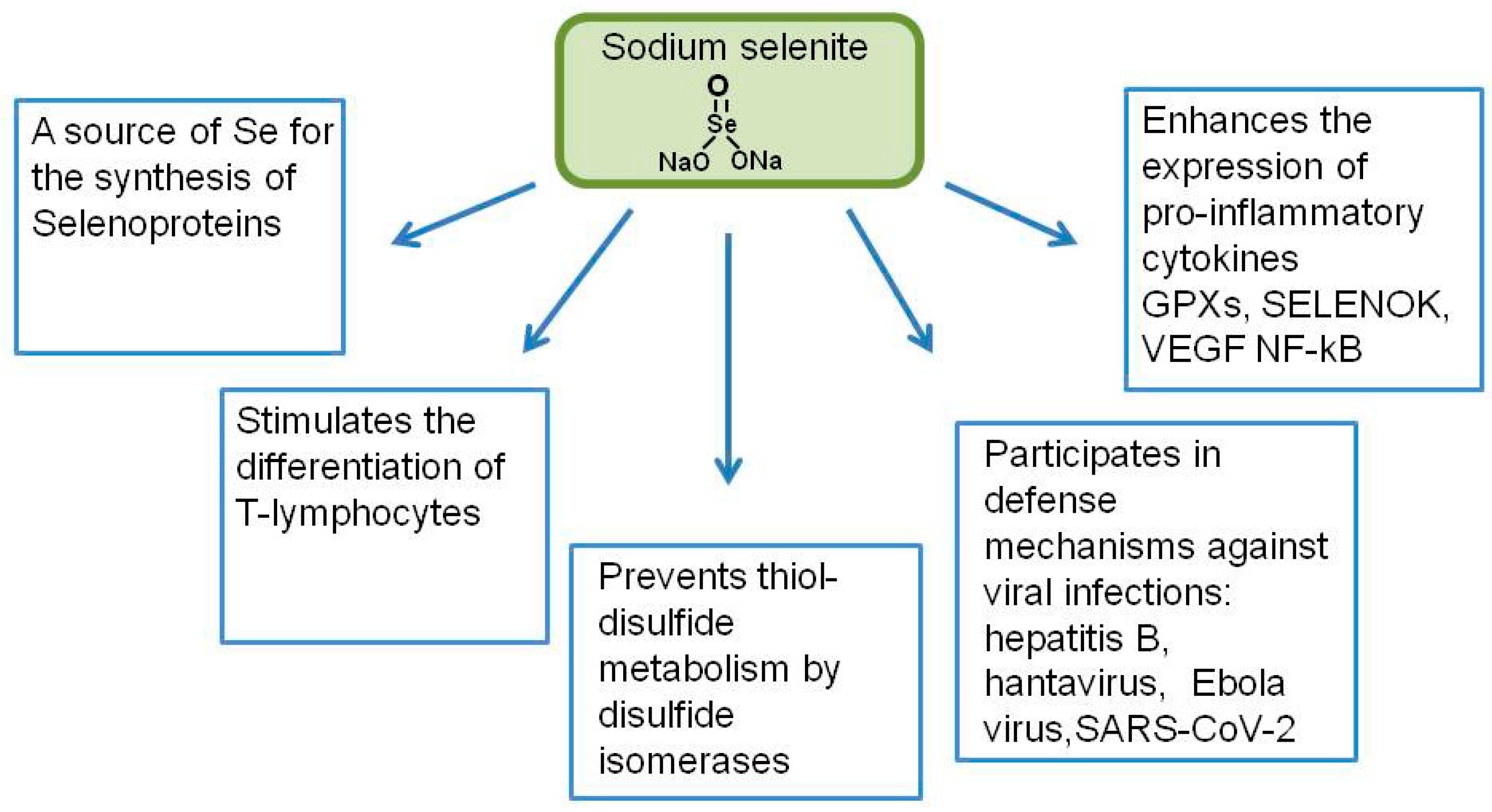
2. Participation of Sodium Selenite in the Regulation of Diseases of the Immune System of Various Etiologies: Possible Role of Sodium Selenite in the Treatment of COVID-19
3. Role of Organic Selenium Compounds in the Immune System and in Protection against COVID-19
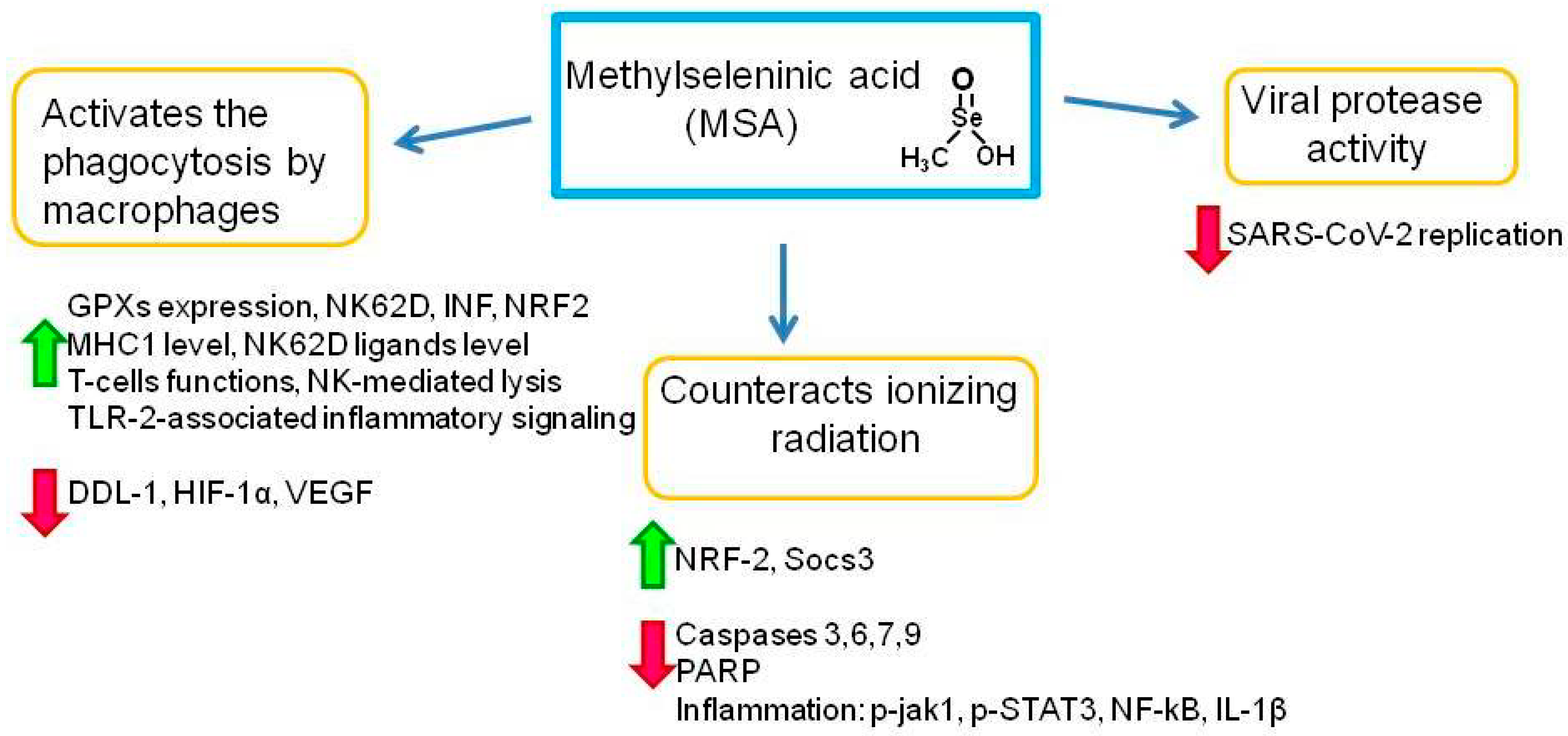
3.1. Role of Methylseleninic Acid in the Immune System and in Protection against COVID-19
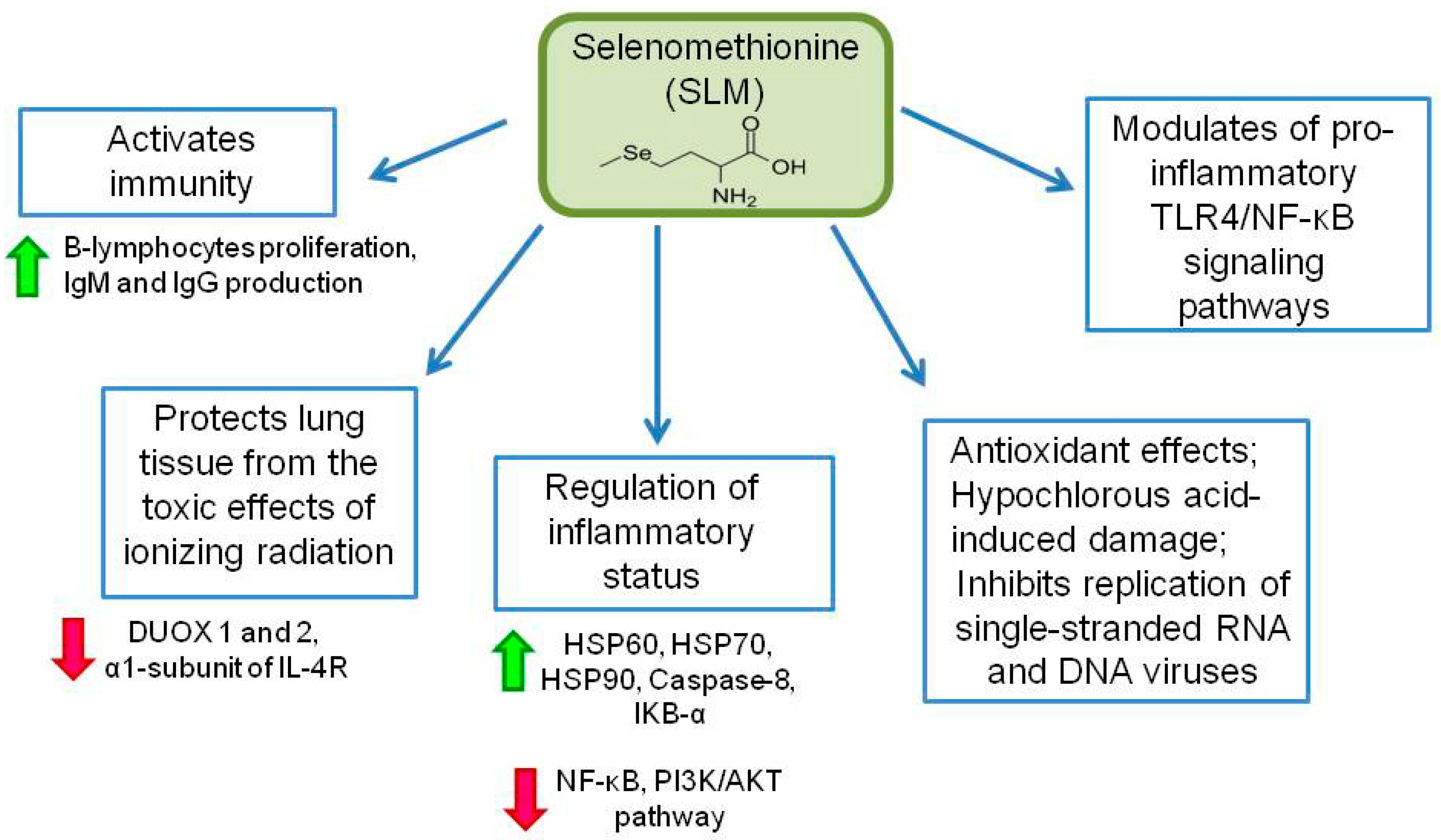
3.2. Role of Selenomethionine in the Immune System and in Protection against COVID-19
3.3. Role of Se-Methylselenocysteine in the Immune System and in Protection against COVID-19
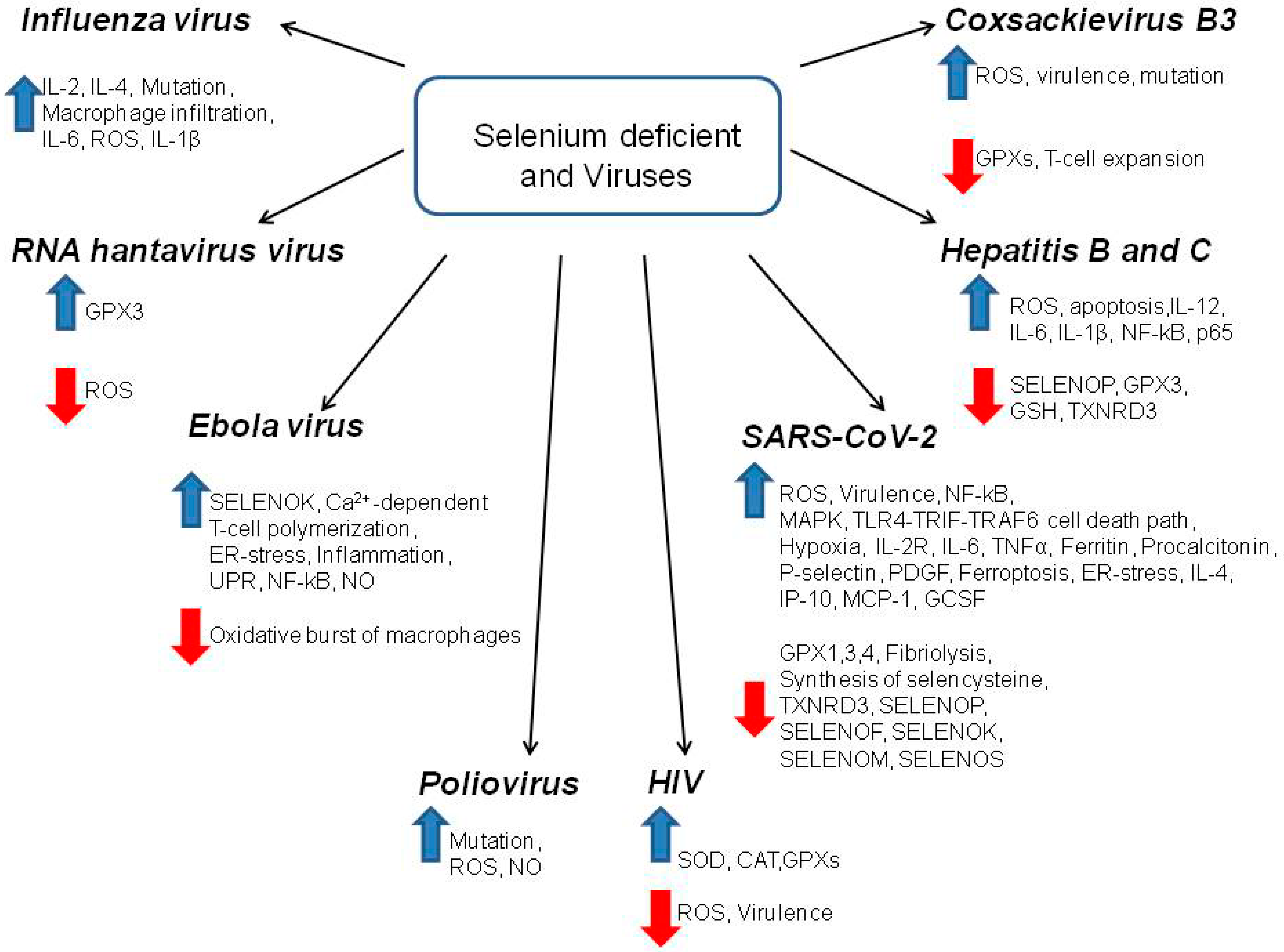
4. The Mechanism of Antiviral Action of Selenium Mediated through Selenoproteins and Selenium Nanoparticles
5. Pharmacological and Toxicological Properties of the Selenium Compounds
6. Discussion
7. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACE2 | Angiotensin converting enzyme 2 |
| CD4+T cells | Cluster of differentiation 4 T helper cells |
| CXCL 9, 10, 11 | Chemokines 9, 10, 11 |
| DFT | Density functional theory |
| FADD | Fas-associated protein with death domain |
| GPX4 | Glutathione peroxidase 4 |
| HSP 60, 70,90 | Heat shock protein 60, 70,90 |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| IRF | Interferon |
| JAK1 | Janus Kinase 1 |
| LAP | LC3-associated phagocytosis |
| MHCI | major histocompatibility complex |
| MLKL | Mixed lineage kinase domain like pseudokinase |
| MMP-9 | Matrix metallopeptidase 9 |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK-cells | Natural killer cells |
| NO | Nitric oxide |
| NRF2 | Nuclear factor erythroid-derived 2-like 2 |
| P53 | Tumor protein P53 |
| PARP | Poly (ADP-ribose) polymerase |
| PI3K/AKT/mTOR pathway | Phosphatidylinositol 3-kinases/lpha serine/threonine-protein kinase/mammalian target of rapamycin |
| ROS | Reactive oxygen species |
| Se | Selenium |
| Sec | Selenocysteine |
| SELENO | Selenoprotein |
| STAT1 | Signal transducer and activator of transcription 1 |
| TLR2,4 | Toll-like receptor 2,4 |
| TNFα | Tumor necrosis factor α |
| TXNRD1,2,3 | Thioredoxin reductase 1,2,3 |
References
- Jeong, D.-W.; Yoo, M.-H.; Kim, T.S.; Kim, J.-H.; Kim, I.Y. Protection of Mice from Allergen-induced Asthma by Selenite. J. Biol. Chem. 2002, 277, 17871–17876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, B.A.; Yoo, M.H.; Shrimali, R.K.; Irons, R.; Gladyshev, V.N.; Hatfield, D.L.; Park, J.M. Role of selenium-containing proteins in T-cell and macrophage function. Proc. Nutr. Soc. 2010, 69, 300–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varlamova, E.G.; Turovsky, E.A.; Babenko, V.A.; Plotnikov, E.Y. The Mechanisms Underlying the Protective Action of Selenium Nanoparticles against Ischemia/Reoxygenation Are Mediated by the Activation of the Ca2+ Signaling System of Astrocytes and Reactive Astrogliosis. Int. J. Mol. Sci. 2021, 22, 12825. [Google Scholar] [CrossRef] [PubMed]
- Turovsky, E.A.; Varlamova, E.G. Mechanism of Ca2+-Dependent Pro-Apoptotic Action of Selenium Nanoparticles, Mediated by Activation of Cx43 Hemichannels. Biology 2021, 10, 743. [Google Scholar] [CrossRef]
- Liu, Y.; Jiao, X.; Teng, X.; Gu, X.; Teng, X. Antagonistic effect of selenium on lead-induced inflammatory injury through inhibiting the nuclear factor-κB signaling pathway and stimulating selenoproteins in chicken hearts. RSC Adv. 2017, 7, 24878–24884. [Google Scholar] [CrossRef] [Green Version]
- Varlamova, E.G.; Turovsky, E.A.; Blinova, E.V. Therapeutic Potential and Main Methods of Obtaining Selenium Nanoparticles. Int. J. Mol. Sci. 2021, 22, 10808. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Goltyaev, M.V.; Mal’tseva, V.N.; Turovsky, E.A.; Sarimov, R.M.; Simakin, A.V.; Gudkov, S.V. Mechanisms of the Cytotoxic Effect of Selenium Nanoparticles in Different Human Cancer Cell Lines. Int. J. Mol. Sci. 2021, 22, 7798. [Google Scholar] [CrossRef]
- Stoffaneller, R.; Morse, N.L. A Review of Dietary Selenium Intake and Selenium Status in Europe and the Middle East. Nutrients 2015, 7, 1494–1537. [Google Scholar] [CrossRef]
- Tutel’ian, V.A. Norms of physiological requirements in energy and nutrients in various groups of population in Russian Federation. Vopr. Pitan. 2009, 78, 4–15. (In Russian) [Google Scholar]
- Sunde, R.A. Selenium. In Present Knowledge in Nutrition, 9th ed.; Bowman, B., Russell, R., Eds.; International Life Sciences Institute: Washington, DC, USA, 2006; pp. 480–497. [Google Scholar]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes: Vitamin C, Vitamin E, Selenium, and Carotenoids; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Jirong, Y.; Huiyun, P.; Zhongzhe, Y.; Birong, D.; Weimin, L.; Ming, Y.; Yi, S. Sodium selenite for treatment of Kashin-Beck disease in children: A systematic review of randomised controlled trials. Osteoarthr. Cartil. 2012, 20, 605–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, M.; Wiebe, N.; Hemmelgarn, B.; Klarenbach, S.; Field, C.; Manns, B.; Thadhani, R.; Gill, J. Trace elements in hemodialysis patients: A systematic review and meta-analysis. BMC Med. 2009, 7, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, C.A.; Kawai, K.; Kupka, R.; Fawzi, W.W. Role of selenium in HIV infection. Nutr. Rev. 2010, 68, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.B.; Schomburg, L.; Kohrle, J.; Pedersen, I.B.; Hollenbach, B.; Hog, A.; Ovesen, L.; Perrild, H.; Laurberg, P. Selenium status, thyroid volume, and multiple nodule formation in an area with mild iodine deficiency. Eur. J. Endocrinol. 2011, 164, 585–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, N.E.; Appleby, P.N.; Roddam, A.W.; Tjonneland, A.; Johnsen, N.F.; Overvad, K.; Boeing, H.; Weikert, S.; Kaaks, R.; Linseisen, J.; et al. Plasma selenium concentration and prostate cancer risk: Results from the European Prospective Investigation into Cancer and Nutrition (EPIC). Am. J. Clin. Nutr. 2008, 88, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. The effect of selenium and vitamin E on risk of prostate cancer and other cancers: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2009, 301, 39–51. [Google Scholar] [CrossRef] [Green Version]
- U.S. Food and Drug Administration. Qualified Health Claims: Letters of Enforcement Discretion. Available online: https://www.fda.gov/food/food-labeling-nutrition/qualified-health-claims-letters-enforcement-discretion (accessed on 5 January 2022).
- Hercberg, S.; Kesse-Guyot, E.; Druesne-Pecollo, N.; Touvier, M.; Favier, A.; Latino-Martel, P.; Briançon, S.; Galan, P. Incidence of cancers, ischemic cardiovascular diseases and mortality during 5-year follow-up after stopping antioxidant vitamins and minerals supplements: A postintervention follow-up in the SU.VI.MAX Study. Int. J. Cancer 2010, 127, 1875–1881. [Google Scholar] [CrossRef]
- Stranges, S.; Marshall, J.R.; Trevisan, M.; Natarajan, R.; Donahue, R.P.; Combs, G.F.; Farinaro, E.; Clark, L.C.; Reid, M.E. Effects of selenium supplementation on cardiovascular disease incidence and mortality: Secondary analyses in a randomized clinical trial. Am. J. Epidemiol. 2006, 163, 694–699. [Google Scholar] [CrossRef] [Green Version]
- MacFarquhar, J.K.; Broussard, D.L.; Melstrom, P.; Hutchinson, R.; Wolkin, A.; Martin, C.; Burk, R.F.; Dunn, J.R.; Green, A.L.; Hammond, R.; et al. Acute selenium toxicity associated with a dietary supplement. Arch. Intern. Med. 2010, 170, 256–261. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Taylor, E.W.; Bennett, K.; Saad, R.; Rayman, M.P. Association between regional selenium status and reported outcome of COVID-19 cases in China. Am. J. Clin. Nutr. 2020, 111, 1297–1299. [Google Scholar] [CrossRef]
- Martinez, S.S.; Huang, Y.; Acuna, L.; Laverde, E.; Trujillo, D.; Barbieri, M.A.; Tamargo, J.; Campa, A.; Baum, M.K. Role of Selenium in Viral Infections with a Major Focus on SARS-CoV-2. Int. J. Mol. Sci. 2021, 23, 280. [Google Scholar] [CrossRef] [PubMed]
- Im, J.H.; Je, Y.S.; Baek, J.; Chung, M.H.; Chung, M.-H.; Kwon, H.Y.; Lee, J.-S. Nutritional status of patients with COVID-19. Int. J. Infect. Dis. 2020, 100, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.; Nagabhushanam, K.; Gowda, S.; Mundkur, L. An Exploratory Study of Selenium Status in Healthy Individuals and in Patients with COVID-19 in a South Indian Population: The Case for Adequate Selenium Status. Nutrition 2021, 82, 111053. [Google Scholar] [CrossRef]
- Younesian, O.; Khodabakhshi, B.; Abdolahi, N.; Norouzi, A.; Behnampour, N.; Hosseinzadeh, S.; Alarzi, S.S.H.; Joshaghani, H. Decreased Serum Selenium Levels of COVID-19 Patients in Comparison with Healthy Individuals. Biol. Trace Elem. Res. 2021, 200, 1562–1567. [Google Scholar] [CrossRef] [PubMed]
- Skalny, A.V.; Timashev, P.S.; Aschner, M.; Aaseth, J.; Chernova, L.N.; Belyaev, V.E.; Grabeklis, A.R.; Notova, S.V.; Lobinski, R.; Tsatsakis, A.; et al. Serum Zinc, Copper, and Other Biometals Are Associated with COVID-19 Severity Markers. Metabolites 2021, 11, 244. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.; Heller, R.A.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Selenium Deficiency Is Associated with Mortality Risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef]
- Pincemail, J.; Cavalier, E.; Charlier, C.; Cheramy–Bien, J.-P.; Brevers, E.; Courtois, A.; Fadeur, M.; Meziane, S.; Goff, C.L.; Misset, B.; et al. Oxidative Stress Status in COVID-19 Patients Hospitalized in Intensive Care Unit for Severe Pneumonia. A Pilot Study. Antioxidants 2021, 10, 257. [Google Scholar] [CrossRef]
- Khomich, O.A.; Kochetkov, S.N.; Bartosch, B.; Ivanov, A.V. Redox Biology of Respiratory Viral Infections. Viruses 2018, 10, 392. [Google Scholar] [CrossRef] [Green Version]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Forceville, X.; Laviolle, B.; Annane, D.; Vitoux, D.; Bleichner, G.; Korach, J.-M.; Cantais, E.; Georges, H.; Soubirou, J.L.; Combes, A.; et al. Effects of high doses of selenium, as sodium selenite, in septic shock. A placebo-controlled, randomized, double-blind, phase II study. Crit Care 2007, 11, R73. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Hoffmann, F.W.; Kumar, M.; Huang, Z.; Roe, K.; Nguyen-Wu, E.; Hashimoto, A.S.; Hoffmann, P.R. Selenoprotein K knockout mice exhibit deficient calcium flux in immune cells and impaired immune responses. J. Immunol. 2011, 186, 2127–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, M.A.; Nelson, H.K.; Shi, Q.; Van Dael, P.; Schiffrin, E.J.; Blum, S.; Barclay, D.; Levander, O.A. Selenium deficiency increases the pathology of an influenza virus infection. FASEB J. 2001, 15, 1481–1483. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Z.; Guo, M.; Xia, Y.; Zhao, M.; Wang, C.; Xu, T.; Chen, T.; Zhu, B. Inhibitory Activity of Selenium Nanoparticles Functionalized with Oseltamivir on H1N1 Influenza Virus. Int. J. Nanomed. 2017, 12, 5733–5743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, M.A.; Esworthy, R.S.; Ho, Y.-S.; Chu, F.-F. Glutathione peroxidase protects mice from viral-induced myocarditis. FASEB J. 1998, 12, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Carmean, C.M.; Landeche, M.; Chellan, B.; Sargis, R.M. Selenomethionine Modulates Insulin Secretion in the MIN6-K8 Mouse Insulinoma Cell Line. FEBS Lett. 2021, 595, 3042–3055. [Google Scholar]
- Loscalzo, J. Keshan disease, selenium deficiency, and the selenoproteome. N. Engl. J. Med. 2014, 370, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- Cermelli, C.; Vinceti, M.; Scaltriti, E.; Bazzani, E.; Beretti, F.; Vivoli, G.; Portolani, M. Selenite inhibition of Coxsackie virus B5replication: Implications on the etiology of Keshan disease. J. Trace Elem. Med. Biol. 2002, 16, 41–46. [Google Scholar] [CrossRef]
- Barrett, C.W.; Reddy, V.K.; Short, S.P.; Motley, A.K.; Lintel, M.K.; Bradley, A.M.; Freeman, T.; Vallance, J.; Ning, W.; Parang, B. Selenoprotein P influences colitis-induced tumorigenesis by mediating stemness and oxidative damage. J. Clin. Investig. 2015, 125, 2646–2660. [Google Scholar] [CrossRef] [Green Version]
- Hamid, M.; Abdulrahim, Y.; Liu, D.; Qian, G.; Khan, A.; Huang, K. The Hepatoprotective Effect of Selenium-Enriched Yeast and Gum Arabic Combination on Carbon Tetrachloride-Induced Chronic Liver Injury in Rats. J. Food Sci. 2018, 83, 525–534. [Google Scholar] [CrossRef]
- Barrett, C.W.; Short, S.P.; Williams, C.S. Selenoproteins and oxidative stress-induced inflammatory tumorigenesis in the gut. Cell. Mol. Life Sci. 2017, 74, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Nettleford, S.K.; Prabhu, K.S. Selenium and Selenoproteins in Gut Inflammation-A Review. Antioxidants 2018, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Gao, X.; Cao, Y.; Jiang, H.; Wang, T.; Song, X.; Guo, M.; Zhang, N. Selenium Deficiency Facilitates Inflammation Through the Regulation of TLR4 and TLR4-Related Signaling Pathways in the Mice Uterus. Inflammation 2015, 38, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhang, Z.; Li, Y.; Shen, P.; Hu, X.; Cao, Y.; Zhang, N. Selenium Deficiency Facilitates Inflammation Following S. aureus Infection by Regulating TLR2-Related Pathways in the Mouse Mammary Gland. Biol. Trace Elem. Res. 2016, 172, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, P.R.; Jourdan-Le Saux, C.; Hoffmann, F.W.; Chang, P.S.; Bollt, O.; He, Q.; Tam, E.K.; Berry, M.J. A role for dietary selenium and selenoproteins in allergic airway inflammation. J. Immunol. 2007, 179, 3258–3267. [Google Scholar] [CrossRef]
- Lipinski, B. Can Selenite be an Ultimate Inhibitor of Ebola and Other Viral Infections? BJMMR 2015, 6, 319–324. [Google Scholar] [CrossRef]
- Sappey, C.; Legrand-Poelsl, S.; Best-Belpomme, M.; Favier, A.; Rentier, B.; Piette, J. Stimulation of glutathione peroxidase activity decreases HIV type 1 activation after oxidative stress. AIDS Res. Hum. Retroviruses 1994, 10, 1451–1461. [Google Scholar] [CrossRef]
- Fang, L.Q.; Goeijenbier, M.; Zuo, S.Q.; Wang, L.-P.; Liang, S.; Klein, S.L.; Li, X.-L.; Liu, K.; Liang, L.; Gong, P.; et al. The association between hantavirus infection and selenium deficiency in mainland China. Viruses 2015, 7, 333–351. [Google Scholar] [CrossRef] [Green Version]
- Broome, C.S.; McArdle, F.; Kyle, J.A.; Andrews, F.; Lowe, N.M.; Hart, C.A.; Arthur, J.R.; Jackson, M.J. An increase in selenium intake improves immune function and poliovirus handling in adults with marginal selenium status. Am. J. Clin. Nutr. 2004, 80, 154–162. [Google Scholar] [CrossRef]
- Huang, Z.; Rose, A.H.; Hoffmann, P.R. The role of selenium in inflammation and immunity: From molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal. 2012, 16, 705–743. [Google Scholar] [CrossRef] [Green Version]
- Tomo, S.; Saikiran, G.; Banerjee, M.; Paul, S. Selenium to selenoproteins-role in COVID-19. EXCLI J. 2021, 20, 781–791. [Google Scholar] [PubMed]
- Ganther, H.E. Selenotrisulfides. Formation by the reaction of thiols with selenious acid. Biochemistry 1968, 7, 2898–2905. [Google Scholar] [CrossRef] [PubMed]
- Ganther, H.E. Reduction of the selenotrisulfide derivative of glutathione to a persulfide analog by gluthathione reductase. Biochemistry 1971, 10, 4089–4098. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Bjornstedt, M.; Holmgren, A. Selenite is a substrate for calf thymus thioredoxin reductase and thioredoxin and elicits a large non-stoichiometric oxidation of NADPH in the presence of oxygen. Eur. J. Biochem. 1992, 207, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, H.S.; Ganther, H.E. Acid-volatile selenium formation catalyzed by glutathione reductase. Biochemistry 2002, 14, 1632–1636. [Google Scholar] [CrossRef]
- Byard, J.L. Trimethyl selenide. A urinary metabolite of selenite. Arch. Biochem. Biophys. 1969, 130, 556–560. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S.; Bzducha-Wróbel, A.; Kurcz, A. Effects of Selenium on Morphological Changes in Candida utilis ATCC 9950 Yeast Cells. Biol. Trace Elem. Res. 2016, 169, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Paolicchi, E.; Gemignani, F.; Krstic-Demonacos, M.; Dedhar, S.; Mutti, L.; Landi, S. Targeting hypoxic response for cancer therapy. Oncotarget 2016, 7, 13464–13478. [Google Scholar] [CrossRef] [Green Version]
- Gailer, J. Arsenic–selenium and mercury–selenium bonds in biology. Coord. Chem. Rev. 2007, 251, 234–254. [Google Scholar] [CrossRef]
- Angstwurm, M.W.A.; Schottdorf, J.; Schopohl, J.; Gaertner, R. Selenium replacement in patients with severe systemic inflammatory response syndrome improves clinical outcome. Crit. Care Med. 1999, 27, 1807–1813. [Google Scholar] [CrossRef]
- Rusolo, F.; Pucci, B.; Colonna, G.; Capone, F.; Guerriero, E.; Milone, M.; Nazzaro, M.; Volpe, M.G.; Bernardo, G.D.; Castello, G.; et al. Evaluation of Selenite Effects on Selenoproteins and Cytokinome in Human Hepatoma Cell Lines. Molecules 2013, 18, 2549–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paskett, E.D.; Dean, J.A.; Oliveri, J.M.; Harrop, J.P. Cancer-Related Lymphedema Risk Factors, Diagnosis, Treatment, and Impact. JCO 2012, 30, 3726–3733. [Google Scholar] [CrossRef] [PubMed]
- Kiremidjian-Schumacher, L.; Roy, M.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Supplementation with selenium and human immune cell functions. Biol. Trace Elem. Res. 1994, 41, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Yu Yu, S.; Zhu, Y.J.; Li, W.G. Protective role of selenium against hepatitis B virus and primary liver cancer in Qidong. Biol. Trace Elem. Res. 1997, 56, 117–124. [Google Scholar] [CrossRef]
- Hou, J.-C. Inhibitory effect of selenite and other antioxidants on complement-mediated tissue injury in patients with epidemic hemorrhagic fever. Biol. Trace Elem. Res. 1997, 56, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Lipinski, B. Selenium supplementation in the prevention of coronavirus infections (COVID-19). Med. Hypotheses 2020, 143, 109878. [Google Scholar] [CrossRef]
- Schomburg, L. A large-bolus injection, but not continuous infusion of sodium selenite improves outcome in peritonitis. Shock 2010, 33, 554–555. [Google Scholar] [CrossRef]
- Lima, L.W.; Nardi, S.; Santoro, V.; Schiavon, M. The Relevance of Plant-Derived Se Compounds to Human Health in the SARS-CoV-2 (COVID-19) Pandemic Era. Antioxidants 2021, 10, 1031. [Google Scholar] [CrossRef]
- Nogueira, C.W.; Barbosa, N.V.; Rocha, J.B.T. Toxicology and pharmacology of synthetic organoselenium compounds: An update. Arch. Toxicol. 2021, 95, 1179–1226. [Google Scholar] [CrossRef]
- Dalia, A.M.; Loh, T.C.; Sazili, A.Q.; Samsudin, A.A. Influence of bacterial organic selenium on blood parameters, immune response, selenium retention and intestinal morphology of broiler chickens. BMC Vet. Res. 2020, 16, 365. [Google Scholar] [CrossRef]
- Wang, Y.; Zhan, X.; Yuan, D.; Zhang, X.; Wu, R. Influence of dietary selenomethionine supplementation on performance and selenium status of broiler breeders and their subsequent progeny. Biol. Trace Elem. Res. 2011, 143, 1497–1507. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium intake, status, and health: A complex relationship. Hormones 2020, 19, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méplan, C.; Hughes, D.J. The Role of Selenium in Health and Disease: Emerging and Recurring Trends. Nutrients 2020, 12, 1049. [Google Scholar] [CrossRef] [PubMed]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, Selenoproteins and Viral Infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santi, C.; Bagnoli, L. Celebrating Two Centuries of Research in Selenium Chemistry: State of the Art and New Prospective. Molecules 2017, 22, 2124. [Google Scholar] [CrossRef] [Green Version]
- Bermano, G.; Méplan, C.; Mercer, D.K.; Hesketh, J.E. Se and viral infection: Are there lessons for COVID-19? Br. J. Nutr. 2021, 125, 618–627. [Google Scholar] [CrossRef]
- Ip, C.; Thompson, H.J.; Zhu, Z.; Ganther, H.E. In vitro and in vivo studies of methylseleninic acid: Evidence that a monomethylated selenium metabolite is critical for cancer chemoprevention. Cancer Res. 2000, 60, 2882–2886. [Google Scholar]
- Zhu, Z.; Jiang, W.; Ganther, H.E.; Thompson, H.J. Mechanisms of cell cycle arrest by methylseleninic acid. Cancer Res. 2002, 62, 156–164. [Google Scholar]
- Babaer, D.; Zheng, M.; Ivy, M.T.; Zent, R.; Tiriveedhi, V. Methylselenol producing selenocompounds enhance the efficiency of mammaglobin-A peptide vaccination against breast cancer cells. Oncol. Lett. 2019, 18, 6891–6898. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Wu, M. The Inhibitory Efficacy of Methylseleninic Acid Against Colon Cancer Xenografts in C57BL/6 Mice. Nutr. Cancer 2015, 67, 831–838. [Google Scholar] [CrossRef]
- Zhang, J.; Basher, F.; Wu, J.D. NKG2D Ligands in Tumor Immunity: Two Sides of a Coin. Front. Immunol. 2015, 6, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlenbrock, F.; Hagemann-Jensen, M.; Kehlet, S.; Andresen, L.; Pastorekova, S.; Skov, S. The NKG2D ligand ULBP2 is specifically regulated through an invariant chain-dependent endosomal pathway. J. Immunol. 2014, 193, 1654–1665. [Google Scholar] [CrossRef] [PubMed]
- Hagemann-Jensen, M.; Uhlenbrock, F.; Kehlet, S.; Andresen, L.; Gabel-Jensen, C.; Ellgaard, L.; Gammelgaard, B.; Skov, S. The selenium metabolite methylselenol regulates the expression of ligands that trigger immune activation through the lymphocyte receptor NKG2D. J. Biol. Chem. 2014, 289, 31576–31590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seliger, B. The link between MHC class I abnormalities of tumors, oncogenes, tumor suppressor genes, and transcription factors. J. Immunotoxicol. 2014, 11, 308–310. [Google Scholar] [CrossRef] [PubMed]
- Lennicke, C.; Rahn, J.; Bukur, J.; Hochgräfe, F.; Wessjohann, L.A.; Lichtenfels, R.; Seliger, B. Modulation of MHC class I surface expression in B16F10 melanoma cells by methylseleninic acid. Oncoimmunology 2016, 6, e1259049. [Google Scholar] [CrossRef] [Green Version]
- Nair, D.; Rådestad, E.; Khalkar, P.; Diaz-Argelich, N.; Schröder, A.; Klynning, C.; Ungerstedt, J.; Uhlin, M.; Fernandes, A.P. Methylseleninic Acid Sensitizes Ovarian Cancer Cells to T-Cell Mediated Killing by Decreasing PDL1 and VEGF Levels. Front. Oncol. 2018, 8 28, 407. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Qiu, C.; Li, W.; Mu, W.; Li, C.; Guo, M. Selenium Plays a Protective Role in Staphylococcus aureus-Induced Endometritis in the Uterine Tissue of Rats. Biol. Trace Elem. Res. 2016, 173, 345–353. [Google Scholar] [CrossRef]
- Chen, Y.C.; Sosnoski, D.M.; Gandhi, U.H.; Novinger, L.J.; Prabhu, K.S.; Mastro, A.M. Selenium modifies the osteoblast inflammatory stress response to bone metastatic breast cancer. Carcinogenesis 2009, 30, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Gawish, R.A.; Fahmy, H.A.; Abd, E.L.; Fattah, A.I.; Nada, A.S. The potential effect of methylseleninic acid (MSA) against gamma-irradiation induced testicular damage in rats: Impact on JAK/STAT pathway. Arch. Biochem. Biophys. 2020, 679, 108205. [Google Scholar] [CrossRef]
- Jayawardena, R.; Sooriyaarachchi, P.; Chourdakis, M.; Jeewandara, C.; Ranasinghe, P. Enhancing immunity in viral infections, with special emphasis on COVID-19: A review. Diabetes Metab. Syndr. 2020, 14, 367–382. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Turovsky, E.A. The main cytotoxic effects of methylseleninic acid on various cancer cells. Int. J. Mol. Sci. 2021, 22, 6614. [Google Scholar] [CrossRef]
- Cai, Z.; Dong, L.; Song, C.; Zhang, Y.; Zhu, C.; Zhang, Y.; Ling, Q.; Hoffmann, P.R.; Li, J.; Huang, Z.; et al. Methylseleninic Acid Provided at Nutritional Selenium Levels Inhibits Angiogenesis by Down-regulating Integrin beta 3 Signaling. Sci. Rep. 2017, 25, 9445. [Google Scholar] [CrossRef] [Green Version]
- Dhanjal, N.I.K.; Sharma, S.; Prabhu, K.S.; Prakash, N.T. Selenium supplementation through Se-rich dietary matrices can upregulate the anti-inflammatory responses in lipopolysaccharide-stimulated murine macrophages. Food Agric. Immunol. 2017, 28, 1374–1392. [Google Scholar] [CrossRef] [Green Version]
- Sanmartín, C.; Plano, D.; Palop, J.A. Selenium compounds and apoptotic modulation: A new perspective in cancer therapy. Mini-Rev. Med. Chem. 2008, 8, 1020–1031. [Google Scholar] [CrossRef]
- Hirano, T.; Murakami, M. COVID-19: A new virus, but a familiar receptor and cytokine release syndrome. Immunity 2020, 52, 731–733. [Google Scholar] [CrossRef]
- Hiffler, L.; Rakotoambinina, B. Selenium and RNA Virus Interactions: Potential Implications for SARS-CoV-2 Infection (COVID-19). Front. Nutr. 2020, 7, 164. [Google Scholar] [CrossRef]
- Chen, W.; Liu, Z.; Zheng, Y.; Wei, B.; Shi, J.; Shao, B.; Wang, D. Selenium donor restricts the intracellular growth of Mycobacterium tuberculosis through the induction of c-Jun-mediated both canonical autophagy and LC3-associated phagocytosis of alveolar macrophages. Microb Pathog. 2021, 161 Pt B, 105269. [Google Scholar] [CrossRef]
- Nasim, M.J.; Zuraik, M.M.; Abdin, A.Y.; Ney, Y.; Jacob, C. Selenomethionine: A Pink Trojan Redox Horse with Implications in Aging and Various Age-Related Diseases. Antioxidants 2021, 10, 882. [Google Scholar] [CrossRef]
- Bakhshalinejad, R.; Akbari Moghaddam Kakhki, R.; Zoidis, E. Effects of different dietary sources and levels of selenium supplements on growth performance, antioxidant status and immune parameters in Ross 308 broiler chickens. Br. Poult. Sci. 2018, 59, 81–91. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Malevu, T.D.; Sochor, J.; Baron, M.; Melcova, M.; Zidkova, J.; et al. A Summary of New Findings on the Biological Effects of Selenium in Selected Animal Species—A Critical Review. Int. J. Mol. Sci. 2017, 18, 2209. [Google Scholar] [CrossRef]
- Shi, X.; Wang, W.; Zheng, S.; Zhang, Q.; Xu, S. Selenomethionine relieves inflammation in the chicken trachea caused by LPS though inhibiting the NF-kappaB pathway. Biol. Trace Elem. Res. 2020, 194, 525–535. [Google Scholar] [CrossRef]
- Duntas, L.H. Selenium and inflammation: Underlying anti-inflammatory mechanisms. Horm. Metab. Res. 2009, 41, 443–447. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, Y.; Wang, J.; Xie, H.; Sun, X.; Zhu, X.; Wei, L.; Liu, Y. Protective Effect of Selenomethionine on T-2 Toxin-Induced Rabbit Immunotoxicity. Biol. Trace Elem. Res. 2022, 200, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Qu, Y.; Yin, K.; Zhang, X.; Lin, H. Selenomethionine ameliorates LPS-induced intestinal immune dysfunction in chicken jejunum. Metallomics 2021, 13, mfab003. [Google Scholar] [CrossRef]
- Zhuang, C.; Liu, G.; Barkema, H.W.; Zhou, M.; Xu, S.; Ur Rahman, S.; Liu, Y.; Kastelic, J.P.; Gao, J.; Han, B. Selenomethionine Suppressed TLR4/NF-kappaB Pathway by Activating Selenoprotein S to Alleviate ESBL Escherichia coli-Induced Inflammation in Bovine Mammary Epithelial Cells and Macrophages. Front. Microbiol. 2020, 8, 1461. [Google Scholar] [CrossRef]
- Wichman, J.; Winther, K.H.; Bonnema, S.J.; Hegedüs, L. Selenium Supplementation Significantly Reduces Thyroid Autoantibody Levels in Patients with Chronic Autoimmune Thyroiditis: A Systematic Review and Meta-Analysis. Thyroid 2016, 26, 1681–1692. [Google Scholar] [CrossRef]
- Pirola, I.; Rotondi, M.; Cristiano, A.; Maffezzoni, F.; Pasquali, D.; Marini, F.; Coperchini, F.; Paganelli, M.; Apostoli, P.; Chiovato, L.; et al. Selenium supplementation in patients with subclinical hypothyroidism affected by autoimmune thyroiditis: Results of the SETI study. Endocrinol. Diabetes Nutr. (Engl. Ed.) 2020, 67, 28–35. [Google Scholar]
- Flouda, K.; Gammelgaard, B.; Davies, M.J.; Hawkins, C.L. Modulation of hypochlorous acid (HOCl) induced damage to vascular smooth muscle cells by thiocyanate and selenium analogues. Redox Biol. 2021, 41, 101873. [Google Scholar] [CrossRef]
- Ivory, K.; Prieto, E.; Spinks, C.; Armah, C.N.; Goldson, A.J.; Dainty, J.R.; Nicolettiet, C. Selenium supplementation has beneficial and detrimental effects on immunity to influenza vaccine in older adults. Clin. Nutr. 2017, 36, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Dound, Y.A.; Sehgal, R. Preclinical Efficacy and Safety Studies of Formulation SSV-003, a Potent Anti-Viral Herbal Formulation. J. Exp. Pharmacol. 2021, 13, 913–921. [Google Scholar] [CrossRef]
- Sagaama, A.; Brandan, S.A.; Ben Issa, T.; Issaoui, N. Searching potential antiviral candidates for the treatment of the 2019 novel coronavirus based on DFT calculations and molecular docking. Heliyon 2020, 6, e04640. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.D.; Morrissey, R.L.; Kapetanovic, I.; Crowell, J.A.; McCormick, D.L. Subchronic toxicity studies of Se-methylselenocysteine, an organoselenium compound for breast cancer prevention. Food Chem. Toxicol. 2008, 46, 1068–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.; Qi, C.; Li, J.; Huang, C.; Zhang, J.; Zhang, Y.; Li, Y.; Fan, B. Se-Methylselenocysteine Alleviates Liver Injury in Diethylnitrosamine (DEN)-Induced Hepatocellular Carcinoma Rat Model by Reducing Liver Enzymes, Inhibiting Angiogenesis, and Suppressing Nitric Oxide (NO)/Nitric Oxide Synthase (NOS) Signaling Pathway. Med. Sci. Monit. 2021, 27, e929255. [Google Scholar] [CrossRef] [PubMed]
- Lendvai, G.; Szekerczés, T.; Kontsek, E.; Selvam, A.; Szakos, A.; Schaff, Z.; Björnstedt, M.; Kiss, A. The Effect of Methylselenocysteine and Sodium Selenite Treatment on microRNA Expression in Liver Cancer Cell Lines. Pathol. Oncol. Res. 2020, 26, 2669–2681. [Google Scholar] [CrossRef]
- Selvam, A.K.; Björnstedt, M.A. Novel Assay Method to Determine the beta-Elimination of Se-Methylselenocysteine to Monomethylselenol by Kynurenine Aminotransferase 1. Antioxidants 2020, 9, 139. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Xia, X.; Wang, W.; Xia, J.; Li, M. Protective Effect of Se-Methylselenocysteine on Elaidic Acid-Induced Inflammation in Human Arterial Endothelial Cells. J. Nutr. Sci. Vitaminol. 2020, 66, 577–582. [Google Scholar] [CrossRef]
- Wang, G.; Zhan, Q.; Wu, H. Suppression of lipopolysaccharide-induced activation of RAW 264.7 macrophages by Se-methylseleno-l-cysteine. Int. Immunopharmacol. 2020, 89 Pt A, 107040. [Google Scholar] [CrossRef]
- Hasnain, S.Z.; Lourie, R.; Das, I.; Chen, A.C.-H.; McGuckin, M.A. The interplay between endoplasmic reticulum stress and inflammation. Immunol. Cell. Biol. 2012, 90, 260–270. [Google Scholar] [CrossRef]
- Hendrayani, S.F.; Al-Harbi, B.; Al-Ansari, M.M.; Silva, S.; Abdelilah, A. The inflammatory/cancer-related IL-6/STAT3/NF-kappaB positive feedback loop includes AUF1 and maintains the active state of breast myofibroblasts. Oncotarget 2016, 7, 41974–41985. [Google Scholar] [CrossRef] [Green Version]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [Green Version]
- Ramanathan, C.S.; Taylor, E.W. Computational genomic analysis of hemorrhagic fever viruses: Viral selenopro-teins as a potential factor in pathogenesis. Biol. Trace Elem. Res. 1997, 56, 93–106. [Google Scholar] [CrossRef]
- Taylor, E.W.; Ramanathan, C.S.; Jalluri, R.K.; Nadimpalli, R.G. A basis for new approaches to the chemotherapy of AIDS: Novel genes in HIV-1 potentially encode selenoproteins expressed by ribosomal frameshifting and termination suppression. J. Med. Chem. 1994, 37, 2637–2654. [Google Scholar] [CrossRef]
- Zhang, W.; Ramanathan, C.S.; Nadimpalli, R.G.; Bhat, A.A.; Cox, A.G.; Taylor, E.W. Selenium-dependent glutathione peroxidase modules encoded by RNA viruses. Biol. Trace Elem. Res. 1999, 70, 97–116. [Google Scholar] [CrossRef]
- Heller, R.A.; Sun, Q.; Hackler, J.; Seelig, J.; Seibert, L.; Cherkezov, A.; Minich, W.B.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox Biol. 2020, 38, 101764. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.; Sun, Y.; Stubbs, D.; He, J.; Li, W.; Wang, F.; Liu, Z.; Ruzicka, J.A.; Taylor, E.W.; et al. SARS-CoV-2 suppresses mRNA expression of selenoproteins associated with ferroptosis, endoplasmic reticulum stress and DNA synthesis. Food Chem. Toxicol. 2021, 153, 112286. [Google Scholar] [CrossRef]
- Codo, A.C.; Davanzo, G.G.; de Brito Monteiro, L.; de Souza, G.F.; Muraro, S.P.; Virgilio-da-Silva, J.V.; Prodonoff, J.S.; Carregari, V.C.; de Biagi Junior, C.A.O.; Crunfli, F.; et al. Elevated glucose levels favor SARS-CoV-2 infection and monocyte response through a HIF-1alpha/Glycolysis-Dependent Axis. Cell Metabol. 2020, 32, 437–446 e435. [Google Scholar] [CrossRef]
- Cuadrado, A.; Pajares, M.; Benito, C.; Jiménez-Villegas, J.; Escoll, M.; Fernández-Ginés, R.; Yagüe, A.J.G.; Lastra, D.; Manda, G.; Rojo, A.I.; et al. Can activation of NRF2 Be a strategy against COVID-19? Trends Pharmacol. Sci. 2020, 41, 598–610. [Google Scholar] [CrossRef]
- Taylor, E.W.; Radding, W. Understanding selenium and glutathione as antiviral factors in COVID-19: Does the viral M(pro) protease target host selenoproteins and glutathione synthesis? Front. Nutr. 2020, 7, 143. [Google Scholar] [CrossRef]
- Lee, B.C.; Peterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs regulate actin assembly and macrophage function via reversible stereoselective methionine oxidation. Mol. Cell. 2013, 51, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.C.; Lee, S.G.; Choo, M.K.; Kim, J.H.; Lee, H.M.; Kim, S.; Fomenko, D.E.; Kim, H.Y.; Park, J.M.; Gladyshev, V.N. Selenoprotein MsrB1 promotes anti-inflammatory cytokine gene expression in macrophages and controls immune response in vivo. Sci. Rep. 2017, 7, 5119. [Google Scholar] [CrossRef]
- Lee, S.C.; Lee, N.H.; Patel, K.D.; Jun, S.K.; Park, J.H.; Knowles, J.C.; Kim, H.W.; Lee, H.H.; Lee, J.H. A Study on Myogenesis by Regulation of Reactive Oxygen Species and Cytotoxic Activity by Selenium Nanoparticles. Antioxidants 2021, 10, 1727. [Google Scholar] [CrossRef]
- Han, H.W.; Patel, K.D.; Kwak, J.H.; Jun, S.K.; Jang, T.S.; Lee, S.H.; Knowles, J.C.; Kim, H.W.; Lee, H.H.; Lee, J.H. Selenium Nanoparticles as Candidates for Antibacterial Substitutes and Supplements against Multidrug-Resistant Bacteria. Biomolecules 2021, 11, 1028. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Gong, G.; Xia, Y.; Wang, C.; Chen, Y.; Hua, L.; Zhong, J.; Tang, Y.; Liu, X.; et al. Restriction of H1N1 Influenza Virus Infection by Selenium Nanoparticles Loaded with Ribavirin via Resisting Caspase-3 Apoptotic Pathway. Int. J. Nanomed. 2018, 13, 5787–5797. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Li, Y.; Xu, T.; Guo, M.; Wang, C.; Zhao, M.; Chen, H.; Kuang, J.; Li, W.; Zhang, Y.; et al. Inhibition of Enterovirus 71 by Selenium Nanoparticles Loaded with siRNA through Bax Signaling Pathways. ACS Omega 2020, 5, 12495–12500. [Google Scholar] [CrossRef]
- Mahdavi, M.; Mavandadnejad, F.; Yazdi, M.H.; Faghfuri, E.; Hashemi, H.; Homayouni-Oreh, S.; Farhoudi, R.; Shahverdi, A.R. Oral Administration of Synthetic Selenium Nanoparticles Induced Robust Th1 Cytokine Pattern after HBs Antigen Vaccination in Mouse Model. J. Infect. Public Health 2017, 10, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Morss, S.G.; Olcott, H.S. Absence of effect of tocopherol on acute oral toxicity of sodium selenite in the rat. Proc. Soc. Exp. Biol. Med. 1967, 124, 483–485. [Google Scholar] [CrossRef]
- Cummins, L.M.; Kimura, E.T. Safety evaluation of selenium sulfide antidandruff shampoos. Toxicol. Appl. Pharmacol. 1971, 20, 89–96. [Google Scholar] [CrossRef]
- Pletnikova, I.P. Biological action and the non-injuriousness level of selenium when it enters the organism together with drinking water. Gig. Sanit. 1970, 35, 14–19. [Google Scholar]
- Franke, K.W.; Moxon, A.L. A comparison of the minimum fatal doses of selenium, tellurium, arsenic and vanadium. J. Pharmacol. Exp. Ther. 1936, 58, 454–459. [Google Scholar]
- Smith, M.I.; Westfall, B.B.; Stohlman, E.F. The elimination of selenium and its distribution in the tissues. Public Health Rep. 1937, 52, 1171–1177. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, J.; Xu, J.F.; Pi, J. The Advancing of Selenium Nanoparticles against Infectious Diseases. Front. Pharmacol. 2021, 12, 682284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.S.; Gao, X.Y.; Zhang, L.D.; Bao, Y.P. Biological effects of a nano red elemental selenium. Biofactors 2001, 15, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Shakibaie, M.; Shahverdi, A.R.; Faramarzi, M.A.; Hassanzadeh, G.R.; Rahimi, H.R.; Sabzevari, O. Acute and subacute toxicity of novel biogenic selenium nanoparticles in mice. Pharm Biol. 2013, 51, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Amini, P.; Kolivand, S.; Saffar, H.; Rezapoor, S.; Motevaseli, E.; Najafi, M.; Nouruzi, F.; Shabeeb, D.; Musa, A.E. Protective effect of selenium-L-methionine on radiation-induced acute pneumonitis and lung fibrosis in Rat. Curr. Clin. Pharmacol. 2019, 14, 157–164. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 6, 667–695. [Google Scholar] [CrossRef]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Kassam, S.; Goenaga-Infante, H.; Maharaj, L.; Hiley, C.T.; Juliger, S.; Joel, S.P. Methylseleninic acid inhibits HDAC activity in diffuse large B-cell lymphoma cell lines. Cancer Chemother. Pharmacol. 2011, 68, 815–821. [Google Scholar] [CrossRef] [Green Version]
- Bera, S.; De Rosa, V.; Rachidi, W.; Diamond, A.M. Does a role for selenium in DNA damage repair explain apparent controversies in its use in chemoprevention? Mutagenesis 2013, 28, 127–134. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mal’tseva, V.N.; Goltyaev, M.V.; Turovsky, E.A.; Varlamova, E.G. Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. Int. J. Mol. Sci. 2022, 23, 2360. https://doi.org/10.3390/ijms23042360
Mal’tseva VN, Goltyaev MV, Turovsky EA, Varlamova EG. Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. International Journal of Molecular Sciences. 2022; 23(4):2360. https://doi.org/10.3390/ijms23042360
Chicago/Turabian StyleMal’tseva, Valentina N., Michael V. Goltyaev, Egor A. Turovsky, and Elena G. Varlamova. 2022. "Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19" International Journal of Molecular Sciences 23, no. 4: 2360. https://doi.org/10.3390/ijms23042360
APA StyleMal’tseva, V. N., Goltyaev, M. V., Turovsky, E. A., & Varlamova, E. G. (2022). Immunomodulatory and Anti-Inflammatory Properties of Selenium-Containing Agents: Their Role in the Regulation of Defense Mechanisms against COVID-19. International Journal of Molecular Sciences, 23(4), 2360. https://doi.org/10.3390/ijms23042360