
Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications




Abstract
:1. Introduction
2. Carbohydrases and Phenolic Compounds in Plants
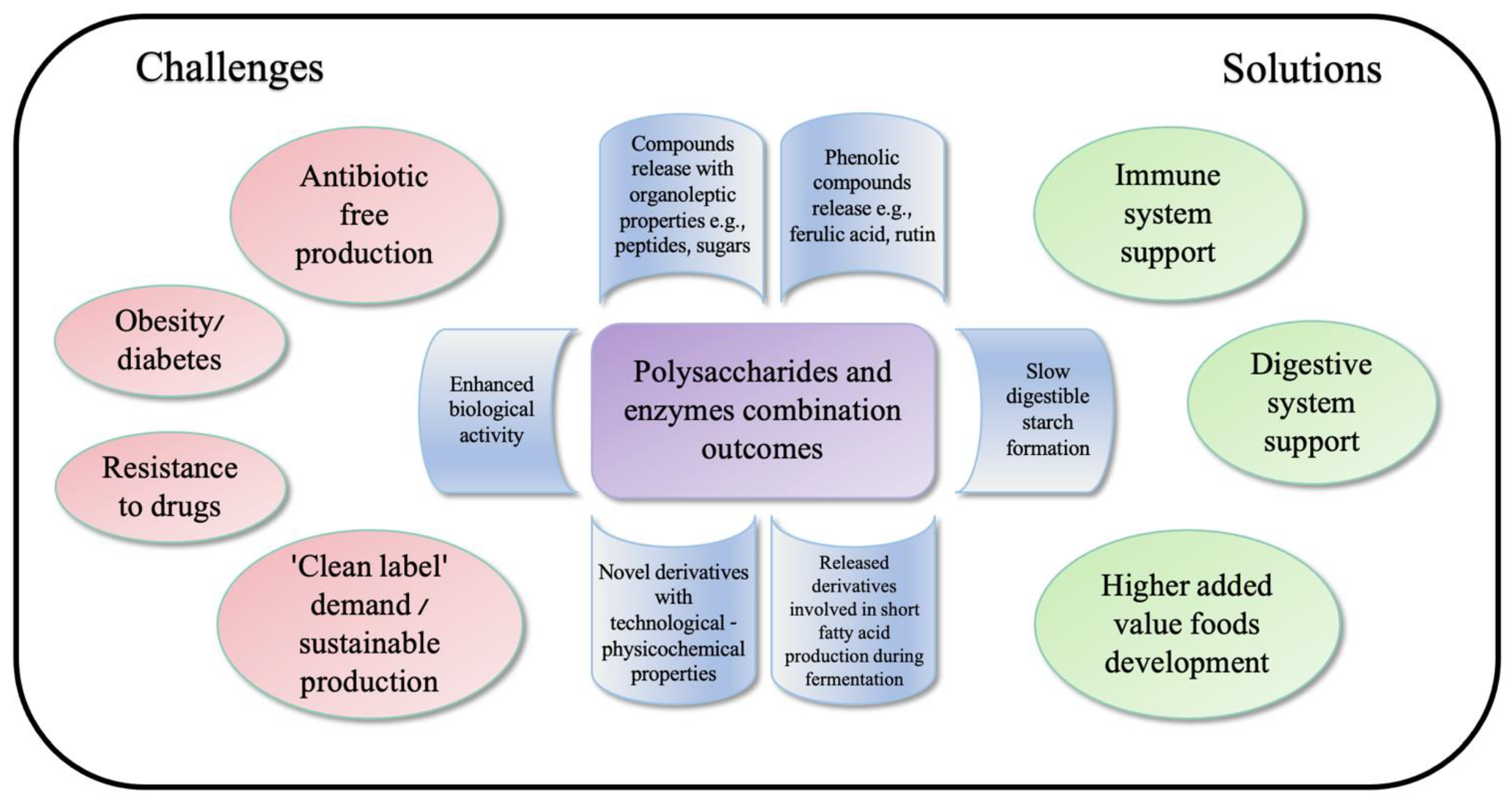
3. Enzymatically Treated Polysaccharides as Possible Functional Components
4. Enzymes-Assisted Processes for Plant Materials
4.1. Bioactives Extraction from By-Products
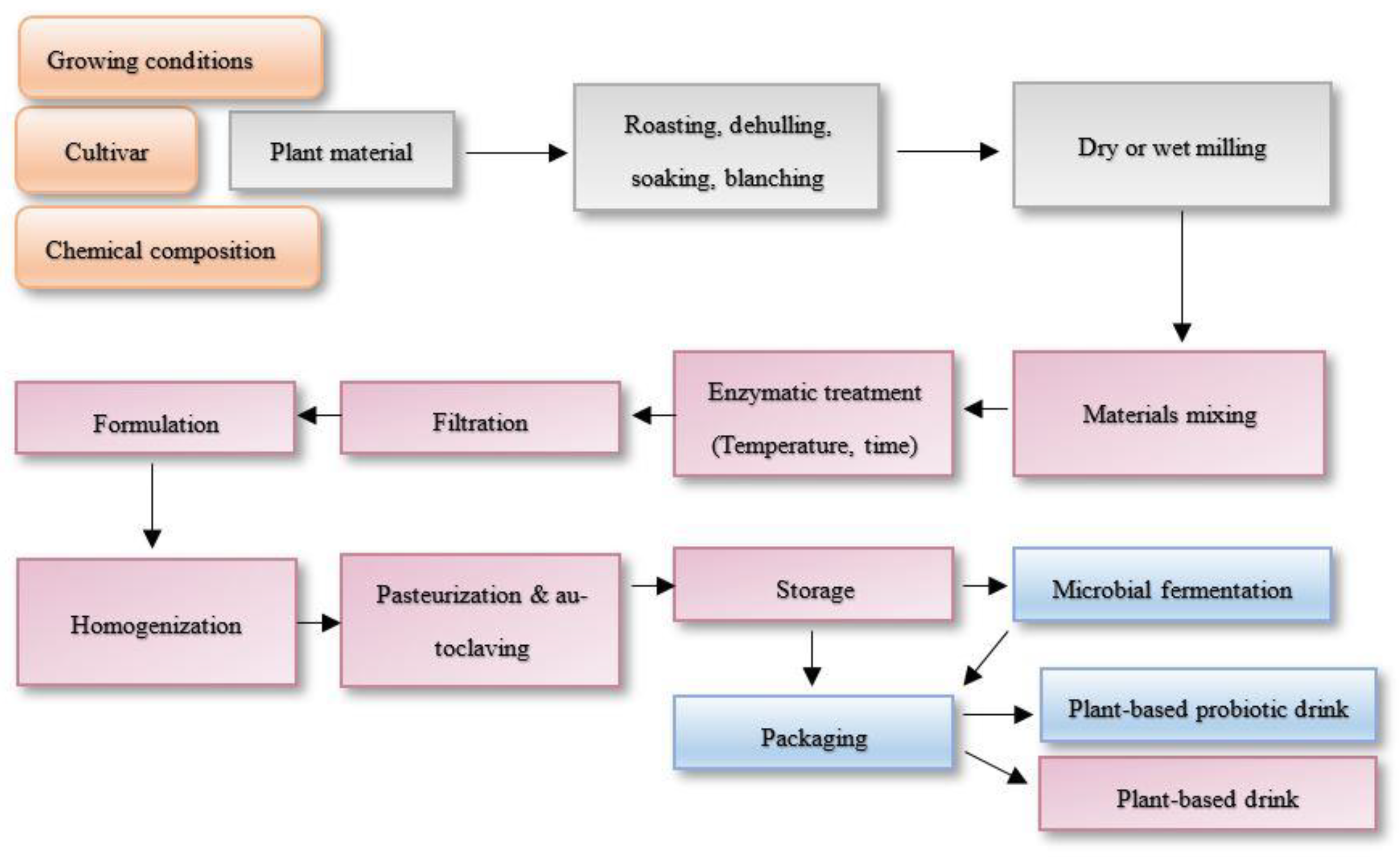
4.2. Plant-Based Drinks from Grains and Fermented Drinks Production
4.3. Nanocrystals, Nanofibers, and Nanocellulose
5. Existing Rules on the Use of Enzymes and Their Products
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rose, N.; Reynolds, T.; Kolodinsky, J. P90 Consumer Use of Food Labels Increases as “Clean Label” Trend Continues. J. Nutr. Educ. Behav. 2020, 52, S58–S59. [Google Scholar] [CrossRef]
- Chawla, G.K.; Garg, K.; Kaur, K.; Chopra, V.; Suri, R. Pattern of drug resistance among patients of pulmonary tuberculosis. Indian J. Tuberc. 2021. [Google Scholar] [CrossRef]
- Grunert, K.G.; De Bauw, M.; Dean, M.; Lähteenmäki, L.; Maison, D.; Pennanen, K.; Sandell, M.A.; Stasiuk, K.; Stickel, L.; Tarrega, A.; et al. No lockdown in the kitchen: How the COVID-19 pandemic has affected food-related behaviours. Food Res. Int. 2021, 150, 110752. [Google Scholar] [CrossRef] [PubMed]
- Gligor, O.; Mocan, A.; Moldovan, C.; Locatelli, M.; Crișan, G.; Ferreira, I.C. Enzyme-assisted extractions of polyphenols—A comprehensive review. Trends Food Sci. Technol. 2019, 88, 302–315. [Google Scholar] [CrossRef]
- Noda-Garcia, L.; Liebermeister, W.; Tawfik, D.S. Metabolite–Enzyme Coevolution: From Single Enzymes to Metabolic Pathways and Networks. Annu. Rev. Biochem. 2018, 87, 187–216. [Google Scholar] [CrossRef] [PubMed]
- BlueWeave Consulting and Research Pvt Ltd. Global industrial enzymes market projected to reach worth $9.2 bn by 2027. Focus Catal. 2021, 2021, 2. [Google Scholar] [CrossRef]
- Mackėla, I.; Andriekus, T.; Venskutonis, P.R. Biorefining of buckwheat (Fagopyrum esculentum) hulls by using supercritical fluid, Soxhlet, pressurized liquid and enzyme-assisted extraction methods. J. Food Eng. 2017, 213, 38–46. [Google Scholar] [CrossRef]
- Socas-Rodríguez, B.; Álvarez-Rivera, G.; Valdés, A.; Ibáñez, E.; Cifuentes, A. Food by-products and food wastes: Are they safe enough for their valorization? Trends Food Sci. Technol. 2021, 114, 133–147. [Google Scholar] [CrossRef]
- Lao, Y.; Zhang, M.; Li, Z.; Bhandari, B. A novel combination of enzymatic hydrolysis and fermentation: Effects on the flavor and nutritional quality of fermented Cordyceps militaris beverage. LWT 2019, 120, 108934. [Google Scholar] [CrossRef]
- Cunningham, M.; Vinderola, G.; Charalampopoulos, D.; Lebeer, S.; Sanders, M.E.; Grimaldi, R. Applying probiotics and prebiotics in new delivery formats—Is the clinical evidence transferable? Trends Food Sci. Technol. 2021, 112, 495–506. [Google Scholar] [CrossRef]
- Włodarczyk, M.; Śliżewska, K. Obesity as the 21st Century’s major disease: The role of probiotics and prebiotics in prevention and treatment. Food Biosci. 2021, 42, 101115. [Google Scholar] [CrossRef]
- Alou, M.T.; Lagier, J.-C.; Raoult, D. Diet influence on the gut microbiota and dysbiosis related to nutritional disorders. Hum. Microbiome J. 2016, 1, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Janssen, A.W.F.; Kersten, S. The role of the gut microbiota in metabolic health. FASEB J. 2015, 29, 3111–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, G.T.C.; Tamashiro, W.M.D.S.C. Role of prebiotics in regulation of microbiota and prevention of obesity. Food Res. Int. 2018, 113, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Daïen, C.I.; Tan, J.; Audo, R.; Mielle, J.; Quek, L.-E.; Krycer, J.R.; Angelatos, A.S.; Duraes, M.; Pinget, G.V.; Ni, D.; et al. Gut-derived acetate promotes B10 cells with antiinflammatory effects. JCI Insight 2021, 6, 144156. [Google Scholar] [CrossRef]
- Mäkinen, O.E.; Wanhalinna, V.; Zannini, E.; Arendt, E.K. Foods for Special Dietary Needs: Non-dairy Plant-based Milk Substitutes and Fermented Dairy-type Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 339–349. [Google Scholar] [CrossRef]
- Penha, C.B.; Santos, V.D.P.; Speranza, P.; Kurozawa, L.E. Plant-based beverages: Ecofriendly technologies in the production process. Innov. Food Sci. Emerg. Technol. 2021, 72, 102760. [Google Scholar] [CrossRef]
- Cantu-Jungles, T.M.; Zhang, X.; Kazem, A.E.; Iacomini, M.; Hamaker, B.R.; Cordeiro, L.M. Microwave treatment enhances human gut microbiota fermentability of isolated insoluble dietary fibers. Food Res. Int. 2021, 143, 110293. [Google Scholar] [CrossRef]
- Zhang, Y.-G.; Kan, H.; Chen, S.-X.; Thakur, K.; Wang, S.; Zhang, J.-G.; Shang, Y.-F.; Wei, Z.-J. Comparison of phenolic compounds extracted from Diaphragma juglandis fructus, walnut pellicle, and flowers of Juglans regia using methanol, ultrasonic wave, and enzyme assisted-extraction. Food Chem. 2020, 321, 126672. [Google Scholar] [CrossRef]
- Puri, M.; Sharma, D.; Barrow, C.J. Enzyme-assisted extraction of bioactives from plants. Trends Biotechnol. 2012, 30, 37–44. [Google Scholar] [CrossRef]
- Islam, S.M.; Ju, L.-K. Enzymatic soybean flour processing: Modeling for insights into optimal carbohydrases composition and carbohydrate monomerization from complex biomass. Catal. Commun. 2021, 149, 106244. [Google Scholar] [CrossRef]
- Jeske, S.; Zannini, E.; Cronin, M.F.; Arendt, E.K. Impact of protease and amylase treatment on proteins and the product quality of a quinoa-based milk substitute. Food Funct. 2018, 9, 3500–3508. [Google Scholar] [CrossRef] [PubMed]
- Dhasarathan, P.; AlSalhi, M.S.; Devanesan, S.; Subbiah, J.; Ranjitsingh, A.; Binsalah, M.; Alfuraydi, A.A. Drug resistance in Candida albicans isolates and related changes in the structural domain of Mdr1 protein. J. Infect. Public Health 2021, 14, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Balciunaitiene, A.; Viskelis, P.; Viskelis, J.; Streimikyte, P.; Liaudanskas, M.; Bartkiene, E.; Zavistanaviciute, P.; Zokaityte, E.; Starkute, V.; Ruzauskas, M.; et al. Green Synthesis of Silver Nanoparticles Using Extract of Artemisia absinthium L., Humulus lupulus L. and Thymus vulgaris L., Physico-Chemical Characterization, Antimicrobial and Antioxidant Activity. Process 2021, 9, 1304. [Google Scholar] [CrossRef]
- Chandhirasekar, K.; Thendralmanikandan, A.; Thangavelu, P.; Nguyen, B.-S.; Nguyen, T.-A.; Sivashanmugan, K.; Nareshkumar, A.; Nguyen, V.-H. Plant-extract-assisted green synthesis and its larvicidal activities of silver nanoparticles using leaf extract of Citrus medica, Tagetes lemmonii, and Tarenna asiatica. Mater. Lett. 2021, 287, 129265. [Google Scholar] [CrossRef]
- Jalab, J.; Abdelwahed, W.; Kitaz, A.; Al-Kayali, R. Green synthesis of silver nanoparticles using aqueous extract of Acacia cyanophylla and its antibacterial activity. Heliyon 2021, 7, 08033. [Google Scholar] [CrossRef]
- Naidu, D.; Hlangothi, S.P.; John, M.J. Bio-based products from xylan: A review. Carbohydr. Polym. 2018, 179, 28–41. [Google Scholar] [CrossRef]
- De Bhowmick, G.; Sarmah, A.K.; Sen, R. Lignocellulosic biorefinery as a model for sustainable development of biofuels and value added products. Bioresour. Technol. 2018, 247, 1144–1154. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Carpita, N.C.; McCann, M.C. Redesigning plant cell walls for the biomass-based bioeconomy. J. Biol. Chem. 2020, 295, 15144–15157. [Google Scholar] [CrossRef]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Jiang, B.; Jin, Z.; BeMiller, J.N. Microbial Starch-Converting Enzymes: Recent Insights and Perspectives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1238–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchart, V.; Šuchová, K.; Biely, P. Xylanases of glycoside hydrolase family 30—An overview. Biotechnol. Adv. 2021, 47, 107704. [Google Scholar] [CrossRef] [PubMed]
- Danalache, F.; Mata, P.; Alves, V.D.; Moldão-Martins, M. Chapter 10—Enzyme-Assisted Extraction of Fruit Juices. In Fruit Juices; Rajauria, G., Brijesh, K., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 183–200. ISBN 978-0-12-802230-6. [Google Scholar]
- Kawamori, M.; Morikawa, Y.; Takasawa, S. Induction and production of cellulases by l-sorbose in Trichoderma reesei. Appl. Microbiol. Biotechnol. 1986, 24, 449–453. [Google Scholar] [CrossRef]
- Juturu, V.; Wu, J.C. Microbial cellulases: Engineering, production and applications. Renew. Sustain. Energy Rev. 2014, 33, 188–203. [Google Scholar] [CrossRef]
- Pedersen, N.R.; Ravn, J.L.; Pettersson, D. A multienzyme NSP product solubilises and degrades NSP structures in canola and mediates protein solubilisation and degradation in vitro. Anim. Feed Sci. Technol. 2017, 234, 244–252. [Google Scholar] [CrossRef]
- Ire, F.S.; Chima, I.J.; Ezebuiro, V. Enhanced xylanase production from UV-mutated Aspergillus niger grown on corn cob and sawdust. Biocatal. Agric. Biotechnol. 2021, 31, 101869. [Google Scholar] [CrossRef]
- Tan, H.; Nie, S. Deciphering diet-gut microbiota-host interplay: Investigations of pectin. Trends Food Sci. Technol. 2020, 106, 171–181. [Google Scholar] [CrossRef]
- Muley, A.B.; Thorat, A.S.; Singhal, R.S.; Babu, K.H. A tri-enzyme co-immobilized magnetic complex: Process details, kinetics, thermodynamics and applications. Int. J. Biol. Macromol. 2018, 118, 1781–1795. [Google Scholar] [CrossRef]
- Ladole, M.R.; Pokale, P.B.; Varude, V.R.; Belokar, P.G.; Pandit, A.B. One pot clarification and debittering of grapefruit juice using co-immobilized enzymes@chitosanMNPs. Int. J. Biol. Macromol. 2021, 167, 1297–1307. [Google Scholar] [CrossRef]
- Biesalski, H.-K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive compounds: Definition and assessment of activity. Nutrition 2009, 25, 1202–1205. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Chapter 8—Environmental Stress and Secondary Metabolites in Plants: An Overview. In Plant Metabolites and Regulation under Environmental Stress; Ahmad, P., Ahanger, M.A., Singh, V.P., Tripathi, D.K., Alam, P., Nasser, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. ISBN 978-0-12-812689-9. [Google Scholar]
- Shamsi, A.; Shahwan, M.; Khan, M.S.; Husain, F.M.; Alhumaydhi, F.A.; Aljohani, A.S.M.; Rehman, T.; Hassan, I.; Islam, A. Elucidating the Interaction of Human Ferritin with Quercetin and Naringenin: Implication of Natural Products in Neurodegenerative Diseases: Molecular Docking and Dynamics Simulation Insight. ACS Omega 2021, 6, 7922–7930. [Google Scholar] [CrossRef] [PubMed]
- Paz, A.; Outeiriño, D.; Guerra, N.P.; Domínguez, J.M. Enzymatic hydrolysis of brewer’s spent grain to obtain fermentable sugars. Bioresour. Technol. 2019, 275, 402–409. [Google Scholar] [CrossRef]
- Uddin, S.; Kabir, T.; Niaz, K.; Jeandet, P.; Clément, C.; Mathew, B.; Rauf, A.; Rengasamy, K.R.; Sobarzo-Sánchez, E.; Ashraf, G.M.; et al. Molecular Insight into the Therapeutic Promise of Flavonoids against Alzheimer’s Disease. Molecules 2020, 25, 1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.P.E. Flavonoids: Modulators of brain function? Br. J. Nutr. 2008, 99, ES60–ES77. [Google Scholar] [CrossRef] [Green Version]
- Gulpinar, A.R.; Orhan, I.E.; Kan, A.; Senol, F.S.; Celik, S.A.; Kartal, M. Estimation of in vitro neuroprotective properties and quantification of rutin and fatty acids in buckwheat (Fagopyrum esculentum Moench) cultivated in Turkey. Food Res. Int. 2012, 46, 536–543. [Google Scholar] [CrossRef]
- Urbanavičiūtė, I.; Liaudanskas, M.; Bobinas, Č.; Šarkinas, A.; Rezgienė, A.; Viskelis, P. Japanese Quince (Chaenomeles japonica) as a Potential Source of Phenols: Optimization of the Extraction Parameters and Assessment of Antiradical and Antimicrobial Activities. Foods 2020, 9, 1132. [Google Scholar] [CrossRef]
- Raudonė, L.; Liaudanskas, M.; Vilkickytė, G.; Kviklys, D.; Žvikas, V.; Viškelis, J.; Viškelis, P. Phenolic Profiles, Antioxidant Activity and Phenotypic Characterization of Lonicera caerulea L. Berries, Cultivated in Lithuania. Antioxidants 2021, 10, 115. [Google Scholar] [CrossRef]
- Yang, B.; Jiang, Y.; Shi, J.; Chen, F.; Ashraf, M. Extraction and pharmacological properties of bioactive compounds from longan (Dimocarpus longan Lour.) fruit—A review. Food Res. Int. 2011, 44, 1837–1842. [Google Scholar] [CrossRef]
- Teo, C.C.; Tan, S.N.; Yong, J.W.H.; Hew, C.S.; Ong, E.S. Pressurized hot water extraction (PHWE). J. Chromatogr. A 2010, 1217, 2484–2494. [Google Scholar] [CrossRef]
- Gardossi, L.; Poulsen, P.B.; Ballesteros, A.; Hult, K.; Švedas, V.K.; Vasić-Rački, Đ.; Carrea, G.; Magnusson, A.; Schmid, A.; Wohlgemuth, R.; et al. Guidelines for reporting of biocatalytic reactions. Trends Biotechnol. 2010, 28, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhou, X.; Zhang, J. Optimization of enzyme assisted extraction of polysaccharides from Astragalus membranaceus. Carbohydr. Polym. 2014, 111, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Zuorro, A.; Lavecchia, R.; González-Delgado, Á.D.; García-Martinez, J.B.; L’Abbate, P. Optimization of Enzyme-Assisted Extraction of Flavonoids from Corn Husks. Process 2019, 7, 804. [Google Scholar] [CrossRef] [Green Version]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2021, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Bulmer, G.S.; de Andrade, P.; Field, R.A.; van Munster, J.M. Recent advances in enzymatic synthesis of β-glucan and cellulose. Carbohydr. Res. 2021, 508, 108411. [Google Scholar] [CrossRef]
- Woo, S.-Y.; Yang, J.Y.; Lee, H.; Ahn, H.J.; Lee, Y.B.; Do, S.H.; Kim, J.Y.; Seo, W.D. Changes in metabolites with harvest times of seedlings of various Korean oat (Avena sativa L.) cultivars and their neuraminidase inhibitory effects. Food Chem. 2021, 373, 131429. [Google Scholar] [CrossRef]
- Cole, M.R.; Eggleston, G.; Gaines, D.K.; Heckemeyer, M. Development of an enzyme cocktail to bioconvert untapped starch in sweet sorghum processing by-products: Part I. Ind. Crop. Prod. 2019, 133, 142–150. [Google Scholar] [CrossRef]
- González, M.J.A.; Carrera, C.; Barbero, G.F.; Palma, M. A comparison study between ultrasound–assisted and enzyme–assisted extraction of anthocyanins from blackcurrant (Ribes nigrum L.). Food Chem. X 2021, 13, 100192. [Google Scholar] [CrossRef]
- Kongthitilerd, P.; Thilavech, T.; Marnpae, M.; Rong, W.; Yao, S.; Adisakwattana, S.; Cheng, H.; Suantawee, T. Cyanidin-3-rutinoside stimulated insulin secretion through activation of L-type voltage-dependent Ca2+ channels and the PLC-IP3 pathway in pancreatic β-cells. Biomed. Pharmacother. 2021, 146, 112494. [Google Scholar] [CrossRef]
- Song, L.-W.; Qi, J.-R.; Liao, J.-S.; Yang, X.-Q. Enzymatic and enzyme-physical modification of citrus fiber by xylanase and planetary ball milling treatment. Food Hydrocoll. 2021, 121, 107015. [Google Scholar] [CrossRef]
- Belmiro, R.H.; Oliveira, L.D.C.; Geraldi, M.V.; Junior, M.R.M.; Cristianini, M. Modification of coffee coproducts by-products by dynamic high pressure, acetylation and hydrolysis by cellulase: A potential functional and sustainable food ingredient. Innov. Food Sci. Emerg. Technol. 2021, 68, 102608. [Google Scholar] [CrossRef]
- Phirom-On, K.; Apiraksakorn, J. Development of cellulose-based prebiotic fiber from banana peel by enzymatic hydrolysis. Food Biosci. 2021, 41, 101083. [Google Scholar] [CrossRef]
- Ninga, K.A.; Desobgo, Z.S.C.; De, S.; Nso, E.J. Pectinase hydrolysis of guava pulp: Effect on the physicochemical characteristics of its juice. Heliyon 2021, 7, 08141. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, Y.; Zeng, J.; Tian, X.; Bei, Q.; Wu, Z. Enhancing three phenolic fractions of oats (Avena sativa L.) and their antioxidant activities by solid-state fermentation with Monascus anka and Bacillus subtilis. J. Cereal Sci. 2020, 93, 102940. [Google Scholar] [CrossRef]
- Pinelo, M.; Arnous, A.; Meyer, A.S. Upgrading of grape skins: Significance of plant cell-wall structural components and extraction techniques for phenol release. Trends Food Sci. Technol. 2006, 17, 579–590. [Google Scholar] [CrossRef]
- Zhou, Z.; Ju, X.; Chen, J.; Wang, R.; Zhong, Y.; Li, L. Charge-oriented strategies of tunable substrate affinity based on cellulase and biomass for improving in situ saccharification: A review. Bioresour. Technol. 2021, 319, 124159. [Google Scholar] [CrossRef]
- HozoVá, B.; Kuniak, L.; Moravcikova, P.; Gajdosova, A. Determination of Water-Insoluble Beta-D-Glucan in the Whole-Grain Cereals and Pseudocereals. Czech J. Food Sci. 2007, 25, 316. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M. Cereal beta-glucans: An underutilized health endorsing food ingredient. Crit. Rev. Food Sci. Nutr. 2020, 1–20. [Google Scholar] [CrossRef]
- Nishantha, M.D.L.C.; Zhao, X.; Jeewani, D.C.; Bian, J.; Nie, X.; Weining, S. Direct comparison of β-glucan content in wild and cultivated barley. Int. J. Food Prop. 2018, 21, 2218–2228. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A Concise Review on the Molecular Structure and Function Relationship of β-Glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Masoodi, F.; Gani, A.; Ashwar, B. Dual enzyme modified oat starch: Structural characterisation, rheological properties, and digestibility in simulated GI tract. Int. J. Biol. Macromol. 2018, 106, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Herburger, K.; Kirkensgaard, J.J.K.; Khakimov, B.; Hansen, A.R.; Blennow, A. Sequential maltogenic α-amylase and branching enzyme treatment to modify granular corn starch. Food Hydrocoll. 2021, 120, 106904. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, P.; Zhao, J. Ferulic acid mediates prebiotic responses of cereal-derived arabinoxylans on host health. Anim. Nutr. 2021. [Google Scholar] [CrossRef]
- Pereira, G.V.; Abdel-Hamid, A.M.; Dutta, S.; D’Alessandro-Gabazza, C.N.; Wefers, D.; Farris, J.A.; Bajaj, S.; Wawrzak, Z.; Atomi, H.; Mackie, R.I.; et al. Degradation of complex arabinoxylans by human colonic Bacteroidetes. Nat. Commun. 2021, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wastyk, H.C.; Fragiadakis, G.K.; Perelman, D.; Dahan, D.; Merrill, B.D.; Yu, F.B.; Topf, M.; Gonzalez, C.G.; Van Treuren, W.; Han, S.; et al. Gut-microbiota-targeted diets modulate human immune status. Cell 2021, 184, 4137–4153.e14. [Google Scholar] [CrossRef]
- Vangay, P.; Johnson, A.; Ward, T.L.; Al-Ghalith, G.A.; Shields-Cutler, R.R.; Hillmann, B.M.; Lucas, S.K.; Beura, L.K.; Thompson, E.A.; Till, L.M.; et al. US Immigration Westernizes the Human Gut Microbiome. Cell 2018, 175, 962–972.e10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Abdo, A.A.A.; Kaddour, B.; Wu, Q.; Xin, L.; Li, X.; Fan, G.; Teng, C. Xylan-oligosaccharides ameliorate high fat diet induced obesity and glucose intolerance and modulate plasma lipid profile and gut microbiota in mice. J. Funct. Foods 2020, 64, 103622. [Google Scholar] [CrossRef]
- Yang, F.; Wei, J.-D.; Lu, Y.-F.; Sun, Y.-L.; Wang, Q.; Zhang, R.-L. Galacto-oligosaccharides modulate gut microbiota dysbiosis and intestinal permeability in rats with alcohol withdrawal syndrome. J. Funct. Foods 2019, 60, 103423. [Google Scholar] [CrossRef]
- Li, E.; Long, X.; Liao, S.; Pang, D.; Li, Q.; Zou, Y. Effect of mulberry galacto-oligosaccharide isolated from mulberry on glucose metabolism and gut microbiota in a type 2 diabetic mice. J. Funct. Foods 2021, 87, 104836. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Widmer, W.W.; Grohmann, K.; Cameron, R. Hydrolysis of grapefruit peel waste with cellulase and pectinase enzymes. Bioresour. Technol. 2007, 98, 1596–1601. [Google Scholar] [CrossRef]
- Qi, W.J.; Sheng, W.S.; Peng, C.; Xiaodong, M.; Yao, T.Z. Investigating into anti-cancer potential of lycopene: Molecular targets. Biomed. Pharmacother. 2021, 138, 111546. [Google Scholar] [CrossRef] [PubMed]
- Catalkaya, G.; Kahveci, D. Optimization of enzyme assisted extraction of lycopene from industrial tomato waste. Sep. Purif. Technol. 2019, 219, 55–63. [Google Scholar] [CrossRef]
- Urbonaviciene, D.; Viskelis, P. The cis-lycopene isomers composition in supercritical CO2 extracted tomato by-products. LWT 2017, 85, 517–523. [Google Scholar] [CrossRef]
- Kundu, D.; Karmakar, S.; Banerjee, R. Simultaneous debittering and clarification of enzyme mediated mixed citrus juice production. Appl. Food Res. 2021, 2, 100031. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, Y.; Tian, H. Effects of carboxymethylation, acidic treatment, hydroxypropylation and heating combined with enzymatic hydrolysis on structural and physicochemical properties of palm kernel expeller dietary fiber. LWT 2020, 133, 109909. [Google Scholar] [CrossRef]
- Marić, M.; Grassino, A.N.; Zhu, Z.; Barba, F.J.; Brnčić, M.; Brnčić, S.R. An overview of the traditional and innovative approaches for pectin extraction from plant food wastes and by-products: Ultrasound-, microwaves-, and enzyme-assisted extraction. Trends Food Sci. Technol. 2018, 76, 28–37. [Google Scholar] [CrossRef]
- Yuliarti, O.; Goh, K.K.; Matia-Merino, L.; Mawson, J.; Brennan, C. Extraction and characterisation of pomace pectin from gold kiwifruit (Actinidia chinensis). Food Chem. 2015, 187, 290–296. [Google Scholar] [CrossRef]
- Rodríguez De Luna, S.L.; Ramírez-Garza, R.E.; Serna-Saldívar, S.O. Environmentally friendly methods for flavonoid extraction from plant material: Impact of their operating conditions on yield and antioxidant properties. Sci. World J. 2020, 2020, 6792069. [Google Scholar] [CrossRef]
- Yazdi, A.P.G.; Barzegar, M.; Sahari, M.A.; Gavlighi, H.A. Optimization of the enzyme-assisted aqueous extraction of phenolic compounds from pistachio green hull. Food Sci. Nutr. 2018, 7, 356–366. [Google Scholar] [CrossRef]
- Costa, R.D.S.; de Almeida, S.S.; Cavalcanti, E.D.C.; Freire, D.M.G.; Moura-Nunes, N.; Monteiro, M.; Perrone, D. Enzymes produced by solid state fermentation of agro-industrial by-products release ferulic acid in bioprocessed whole-wheat breads. Food Res. Int. 2021, 140, 109843. [Google Scholar] [CrossRef]
- Yin, Z.; Wu, W.; Sun, C.; Lei, Z.; Chen, H.; Liu, H.; Chen, W.; Ma, J.; Min, T.; Zhang, M.; et al. Comparison of releasing bound phenolic acids from wheat bran by fermentation of three Aspergillus species. Int. J. Food Sci. Technol. 2018, 53, 1120–1130. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rempel, A.; Machado, T.; Treichel, H.; Colla, E.; Margarites, A.C.; Colla, L. Saccharification of Spirulina platensis biomass using free and immobilized amylolytic enzymes. Bioresour. Technol. 2018, 263, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.A.; Uekane, T.M.; de Miranda, J.F.; Ruiz, L.F.; da Motta, J.C.B.; Silva, C.B.; Pitangui, N.D.S.; Gonzalez, A.G.M.; Fernandes, F.F.; Lima, A.R. Kombucha beverage from non-conventional edible plant infusion and green tea: Characterization, toxicity, antioxidant activities and antimicrobial properties. Biocatal. Agric. Biotechnol. 2021, 34, 102032. [Google Scholar] [CrossRef]
- Bai, J.; Li, Y.; Zhang, W.; Fan, M.; Qian, H.; Zhang, H.; Qi, X.; Wang, L. Source of gut microbiota determines oat β-glucan degradation and short chain fatty acid-producing pathway. Food Biosci. 2021, 41, 101010. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, L.; Zhang, Z.; Ding, L.; Hang, S. Combination of fiber-degrading enzymatic hydrolysis and lactobacilli fermentation enhances utilization of fiber and protein in rapeseed meal as revealed in simulated pig digestion and fermentation in vitro. Anim. Feed Sci. Technol. 2021, 278, 115001. [Google Scholar] [CrossRef]
- Nissen, L.; di Carlo, E.; Gianotti, A. Prebiotic potential of hemp blended drinks fermented by probiotics. Food Res. Int. 2020, 131, 109029. [Google Scholar] [CrossRef]
- Huebner, J.; Wehling, R.; Parkhurst, A.; Hutkins, R. Effect of processing conditions on the prebiotic activity of commercial prebiotics. Int. Dairy J. 2008, 18, 287–293. [Google Scholar] [CrossRef]
- Du, J.; Pan, R.; Obadi, M.; Li, H.; Shao, F.; Sun, J.; Wang, Y.; Qi, Y.; Xu, B. In vitro starch digestibility of buckwheat cultivars in comparison to wheat: The key role of starch molecular structure. Food Chem. 2021, 368, 130806. [Google Scholar] [CrossRef]
- Shori, A.B.; Aljohani, G.S.; Al-Zahrani, A.J.; Al-Sulbi, O.S.; Baba, A.S. Viability of probiotics and antioxidant activity of cashew milk-based yogurt fermented with selected strains of probiotic Lactobacillus spp. LWT 2021, 153, 112482. [Google Scholar] [CrossRef]
- Panghal, A.; Janghu, S.; Virkar, K.; Gat, Y.; Kumar, V.; Chhikara, N. Potential non-dairy probiotic products—A healthy approach. Food Biosci. 2018, 21, 80–89. [Google Scholar] [CrossRef]
- Guimarães, J.T.; Balthazar, C.F.; Silva, R.; Rocha, R.S.; Graça, J.S.; A Esmerino, E.; Silva, M.C.; Sant’Ana, A.S.; Duarte, M.C.K.H.; Freitas, M.Q.; et al. Impact of probiotics and prebiotics on food texture. Curr. Opin. Food Sci. 2020, 33, 38–44. [Google Scholar] [CrossRef]
- Shori, A.B.; AL Zahrani, A.J. Non-dairy plant-based milk products as alternatives to conventional dairy products for delivering probiotics. Food Sci. Technol. 2021. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Wei, Z.; Yin, B.; Man, C.; Jiang, Y. Enhancement of functional characteristics of blueberry juice fermented by Lactobacillus plantarum. LWT 2021, 139, 110590. [Google Scholar] [CrossRef]
- Gao, B.; Wang, J.; Wang, Y.; Xu, Z.; Li, B.; Meng, X.; Sun, X.; Zhu, J. Influence of fermentation by lactic acid bacteria and in vitro digestion on the biotransformations of blueberry juice phenolics. Food Control 2021, 133, 108603. [Google Scholar] [CrossRef]
- Ahmed, J.; Gultekinoglu, M.; Edirisinghe, M. Bacterial cellulose micro-nano fibres for wound healing applications. Biotechnol. Adv. 2020, 41, 107549. [Google Scholar] [CrossRef]
- Rossi, B.R.; Pellegrini, V.O.; Cortez, A.A.; Chiromito, E.M.; Carvalho, A.J.; Pinto, L.O.; Rezende, C.A.; Mastelaro, V.R.; Polikarpov, I. Cellulose nanofibers production using a set of recombinant enzymes. Carbohydr. Polym. 2021, 256, 117510. [Google Scholar] [CrossRef]
- Meftahi, A.; Samyn, P.; Geravand, S.A.; Khajavi, R.; Alibkhshi, S.; Bechelany, M.; Barhoum, A. Nanocelluloses as skin biocompatible materials for skincare, cosmetics, and healthcare: Formulations, regulations, and emerging applications. Carbohydr. Polym. 2021, 278, 118956. [Google Scholar] [CrossRef]
- Ceylan, R.; Demirbas, A.; Ocsoy, I.; Aktumsek, A. Green synthesis of silver nanoparticles using aqueous extracts of three Sideritis species from Turkey and evaluations bioactivity potentials. Sustain. Chem. Pharm. 2021, 21, 100426. [Google Scholar] [CrossRef]
- Liu, S.; Qamar, S.A.; Qamar, M.; Basharat, K.; Bilal, M. Engineered nanocellulose-based hydrogels for smart drug delivery applications. Int. J. Biol. Macromol. 2021, 181, 275–290. [Google Scholar] [CrossRef]
- Yarbrough, J.M.; Zhang, R.; Mittal, A.; Wall, T.V.; Bomble, Y.J.; Decker, S.R.; Himmel, M.E.; Ciesielski, P.N. Multifunctional Cellulolytic Enzymes Outperform Processive Fungal Cellulases for Coproduction of Nanocellulose and Biofuels. ACS Nano 2017, 11, 3101–3109. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Chae, M.; Beyene, D.; Danumah, C.; Tosto, F.; Bressler, D.C. Co-Production of Cellulose Nanocrystals and Fermentable Sugars Assisted by Endoglucanase Treatment of Wood Pulp. Materials 2018, 11, 1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladics, G.S.; Fan, L.; Sewalt, V.J.; Spök, A. Chapter Eight—Safety Assessment and Regulation of Food Enzymes. In Enzymes; Kermasha, S., Michael, N.A., Eds.; Academic Press: San Diego, CA, USA, 2021; pp. 203–258. ISBN 978-0-12-800217-9. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Starch Polysaccharides | |||
|---|---|---|---|
| Hemicellulose | Cellulose | Pectin | |
| Consist of | xyloglucans | cellulose nanofibrils: | homogalacturonan |
| glucuronoarabinoxylan | (a) xylan | rhamnogalacturonan I and II | |
| β–glucan | (b) mannan | xylogalacturonan | |
| glucomannan | |||
| Enzymes used in processes | Xylanases: | Cellulases: | Pectinases: |
| exoxylanases | endo–(1,4)–β–d–glucanase (EC 3.2.1.4) | polygalacturonases | |
| β–xylosidases, | exo–(1,4)–β–d–glucanase (EC 3.2.1.91) | pectin esterases | |
| xylan–1,4–β-xylosidase | β–glucosidases (EC 3.3.1.21) | pectate lyase | |
| endoxylanases | β–glucosidases (EC 3.3.1.21) | ||
| Starches | |||
| Consist of | amylose | ||
| amylopectin | |||
| Enzymes used in processes | α–Amylases (EC 3.2.1.1) | ||
| β–amylase (EC 3.2.1.2) | |||
| glucoamylase (EC 3.2.1.3) | |||
| α-glucosidase (EC 3.2.1.20) | |||
| pullulanase or amylopullulanase (EC 3.2.1.41) | |||
| cyclodextrin glycosyltransferase (EC 2.4.1.19) | |||
| Enzymes | Producent | Substrate | Enzymes Quantity | Liquid to Substrate Ratio | pH | Temp. °C | Time | Ref. |
|---|---|---|---|---|---|---|---|---|
| Xylanase cocktail | A. niger | Citrus fiber | 0.45% | 1:20 | 4.5−6.5 | 50 | 120 min | Song et al. [62] |
| Cellulase | A. niger | Coffee by-products | 5−15 U | 1:25 | 5.0−6.0 | 50 | 30−20 min | Belmiro et al. [63] |
| Cellulase from Celluclast 1.5 L | T. reesei | Banana peel | 5 FPU/ml | 1:20 | 6.0−7.0 | 50 | 120 h | Phirom-on et al. [64] |
| Pectinase | A. niger | Guava pulp | 0.10% | 2:5 | 2.97−3.97 | 45 | 3–90 min | Ninga et al. [65] |
| Pectinase | A. niger | Blackcurrant | 108 U/g | 0.1:15 and 0.2:15 | 5−6 | 60 | 10−90 min | Gonzalez et al. [60] |
| Heat stable alpha-amylase | Bacillus sp. | Oat flours | 0.01% | 1:5 | 5.0−9.0 | 100 | 15−75 min | Chen et al. [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Streimikyte, P.; Viskelis, P.; Viskelis, J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. Int. J. Mol. Sci. 2022, 23, 2359. https://doi.org/10.3390/ijms23042359
Streimikyte P, Viskelis P, Viskelis J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. International Journal of Molecular Sciences. 2022; 23(4):2359. https://doi.org/10.3390/ijms23042359
Chicago/Turabian StyleStreimikyte, Paulina, Pranas Viskelis, and Jonas Viskelis. 2022. "Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications" International Journal of Molecular Sciences 23, no. 4: 2359. https://doi.org/10.3390/ijms23042359
APA StyleStreimikyte, P., Viskelis, P., & Viskelis, J. (2022). Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. International Journal of Molecular Sciences, 23(4), 2359. https://doi.org/10.3390/ijms23042359