
Biodiversity and Metabolic Potential of Bacteria in Bulk Soil from the Peri-Root Zone of Black Alder (Alnus glutinosa), Silver Birch (Betula pendula) and Scots Pine (Pinus sylvestris)

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
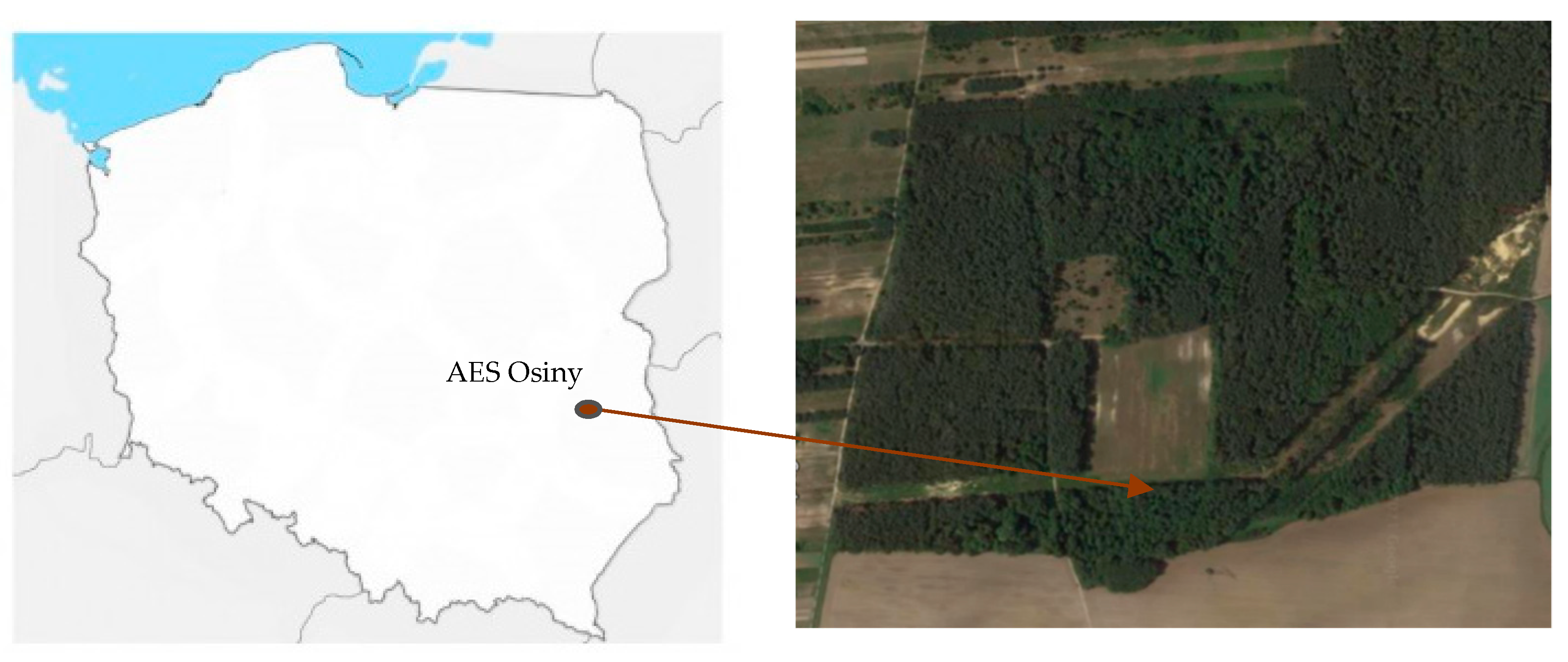
2.1. Field Site and Soil Chemical Analyses
2.2. Enzymatic Activities
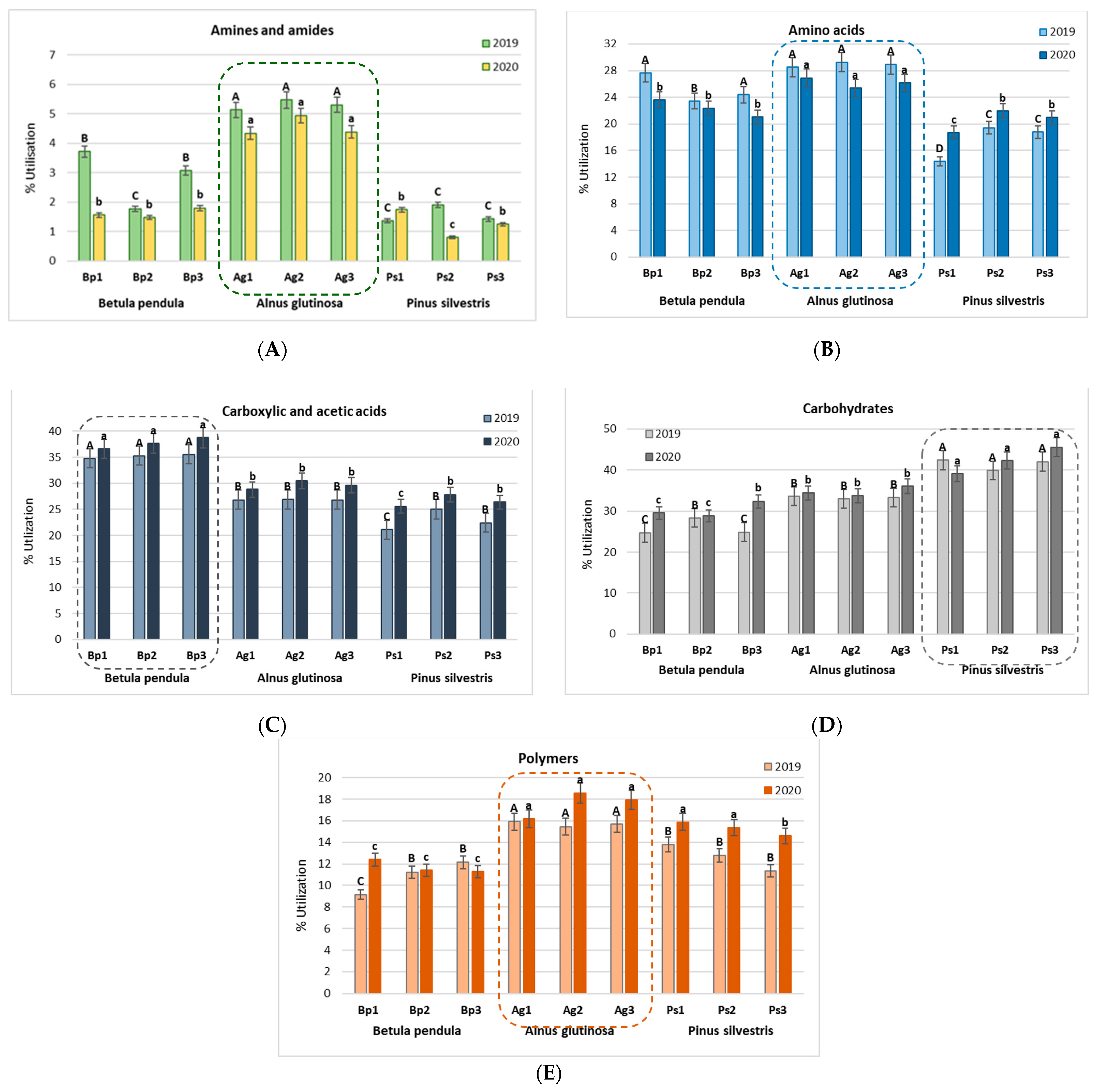
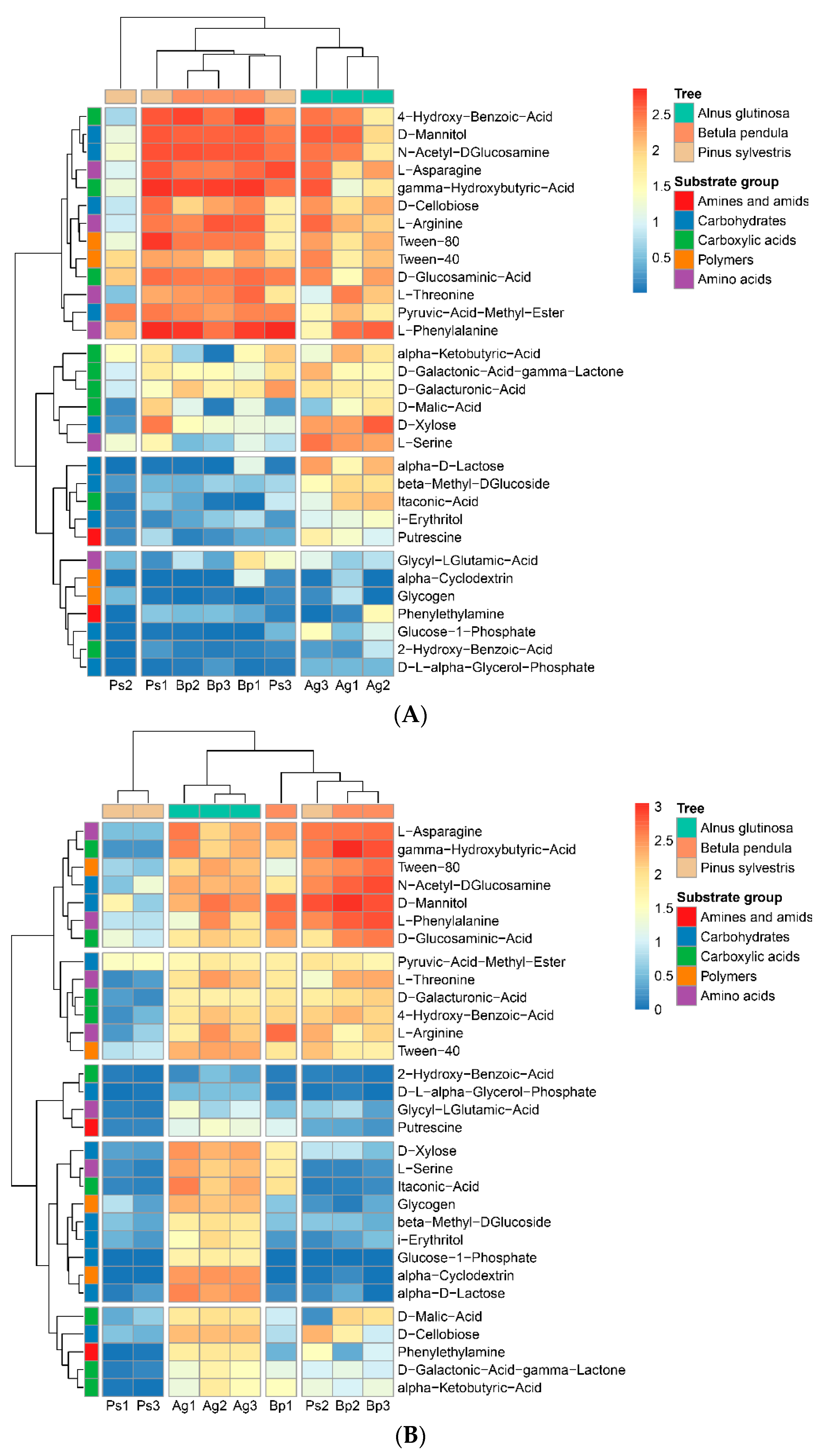
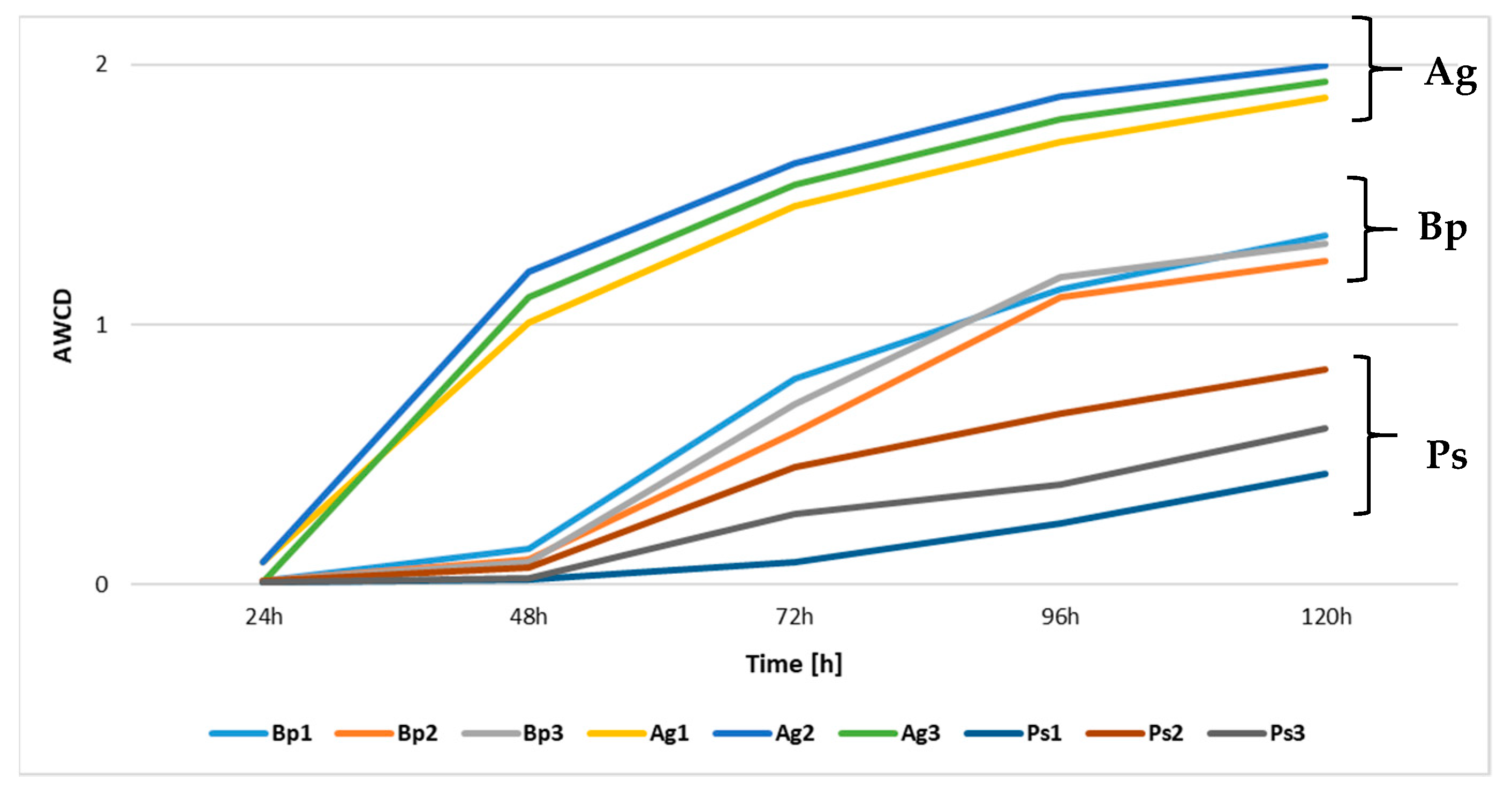
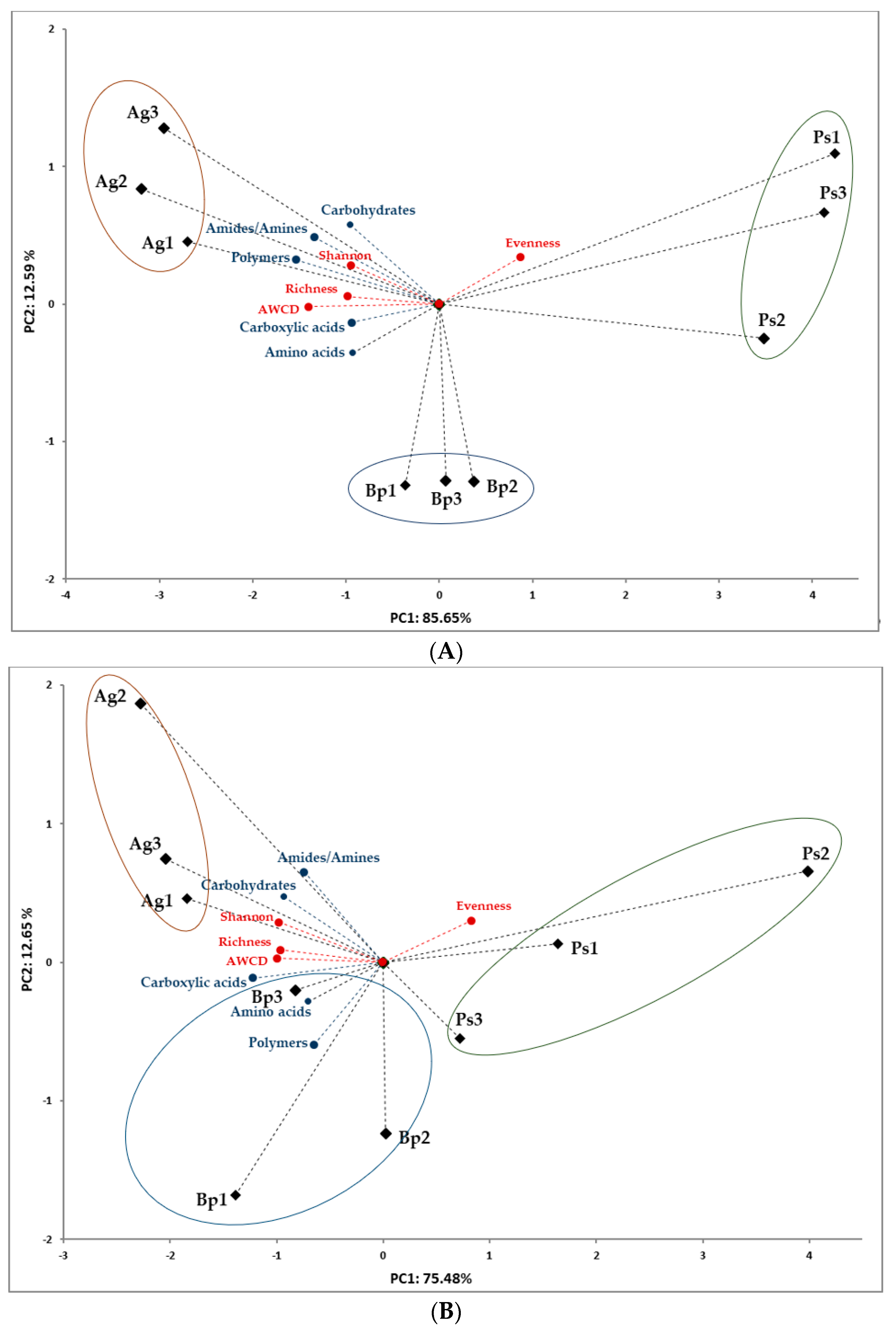
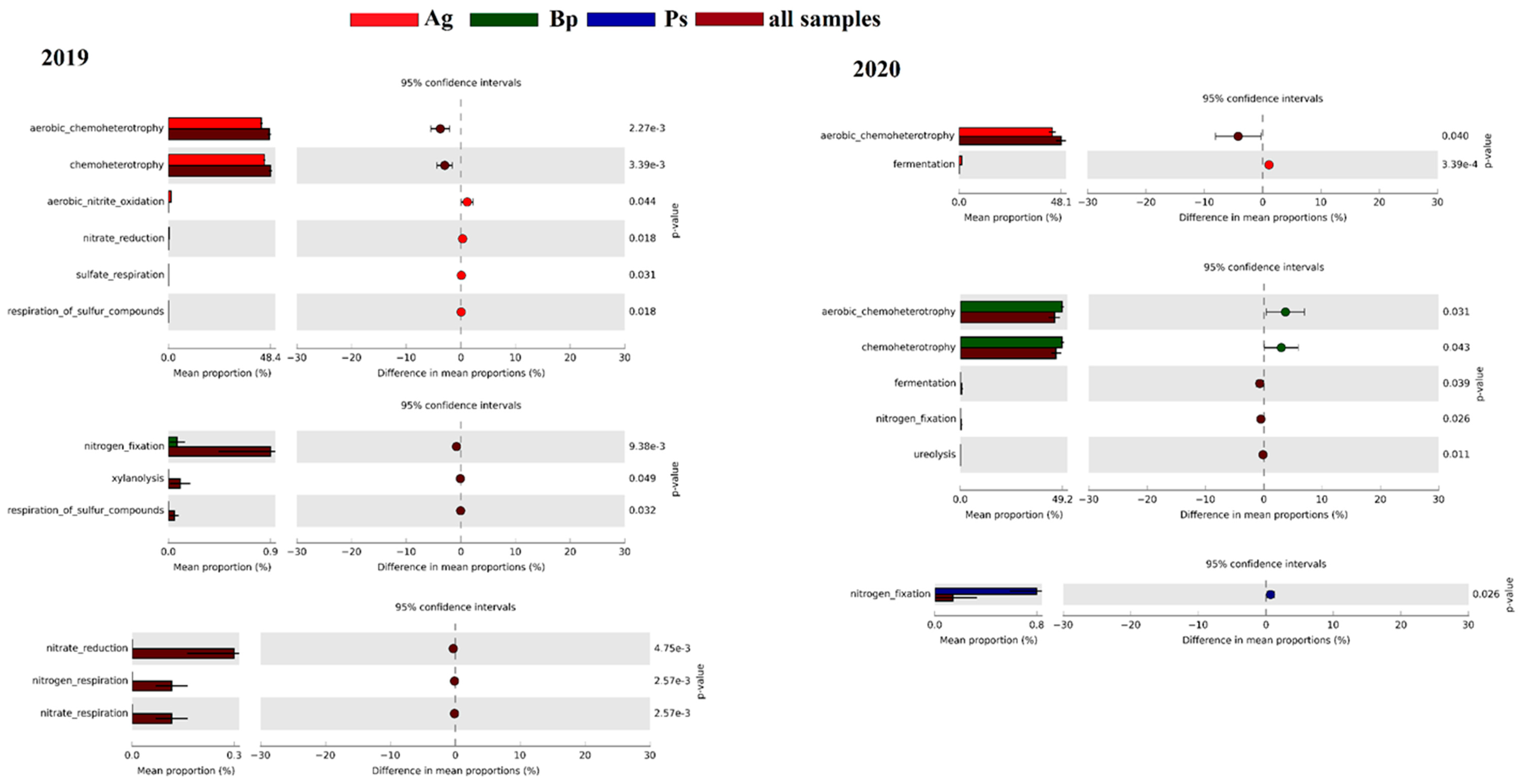
2.3. Metabolic Profiles of Bacterial Community Based on Biolog EcoPlates
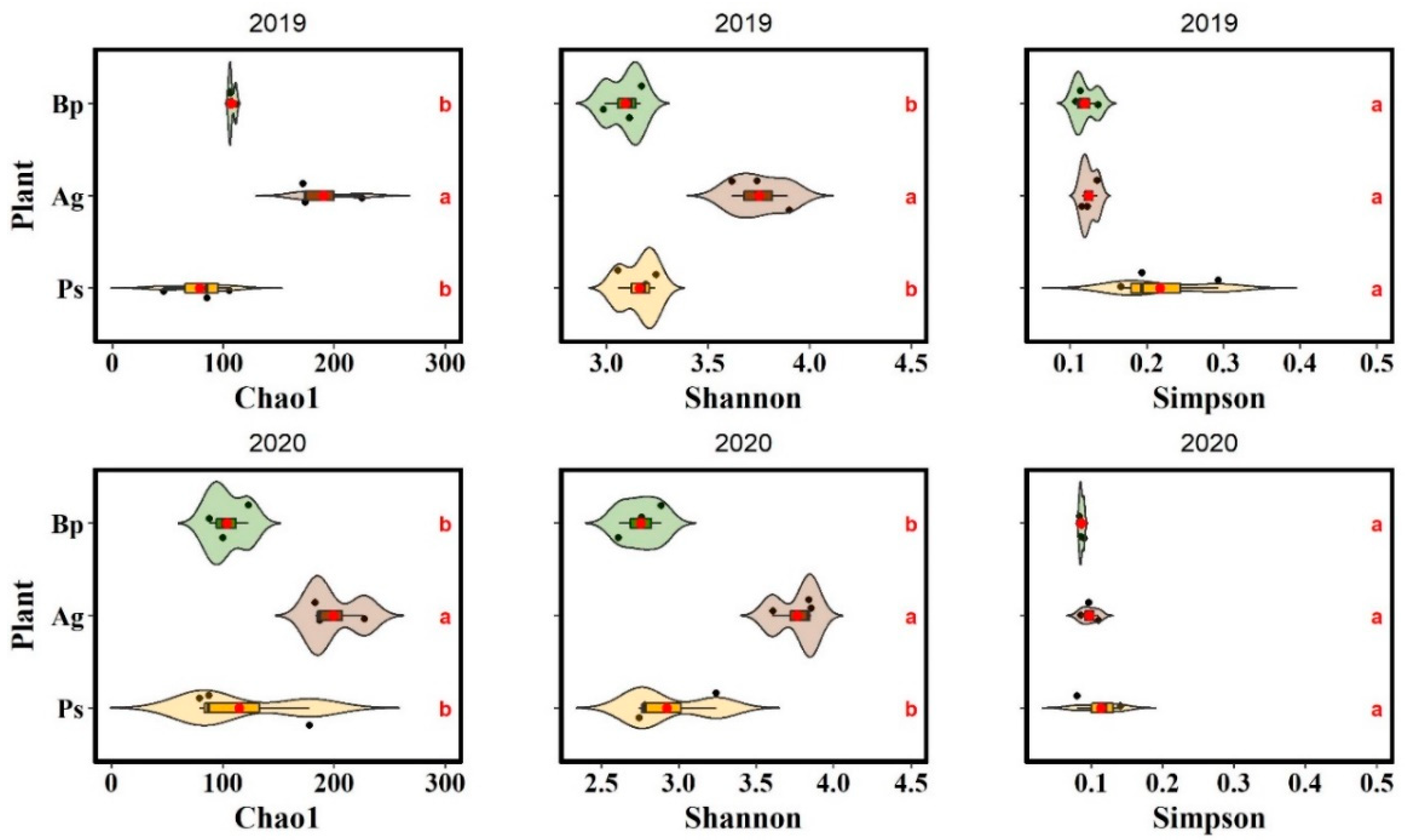
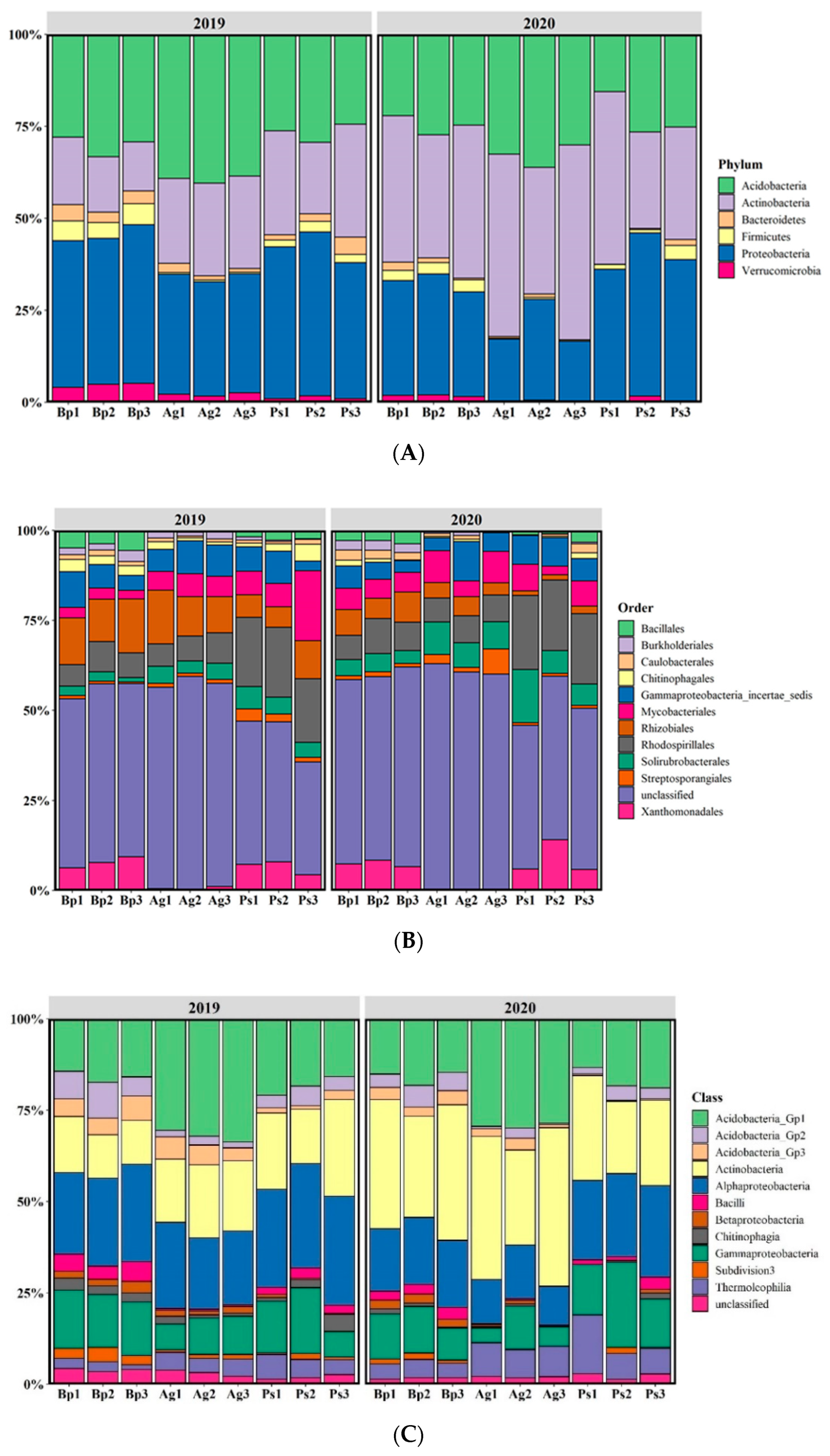
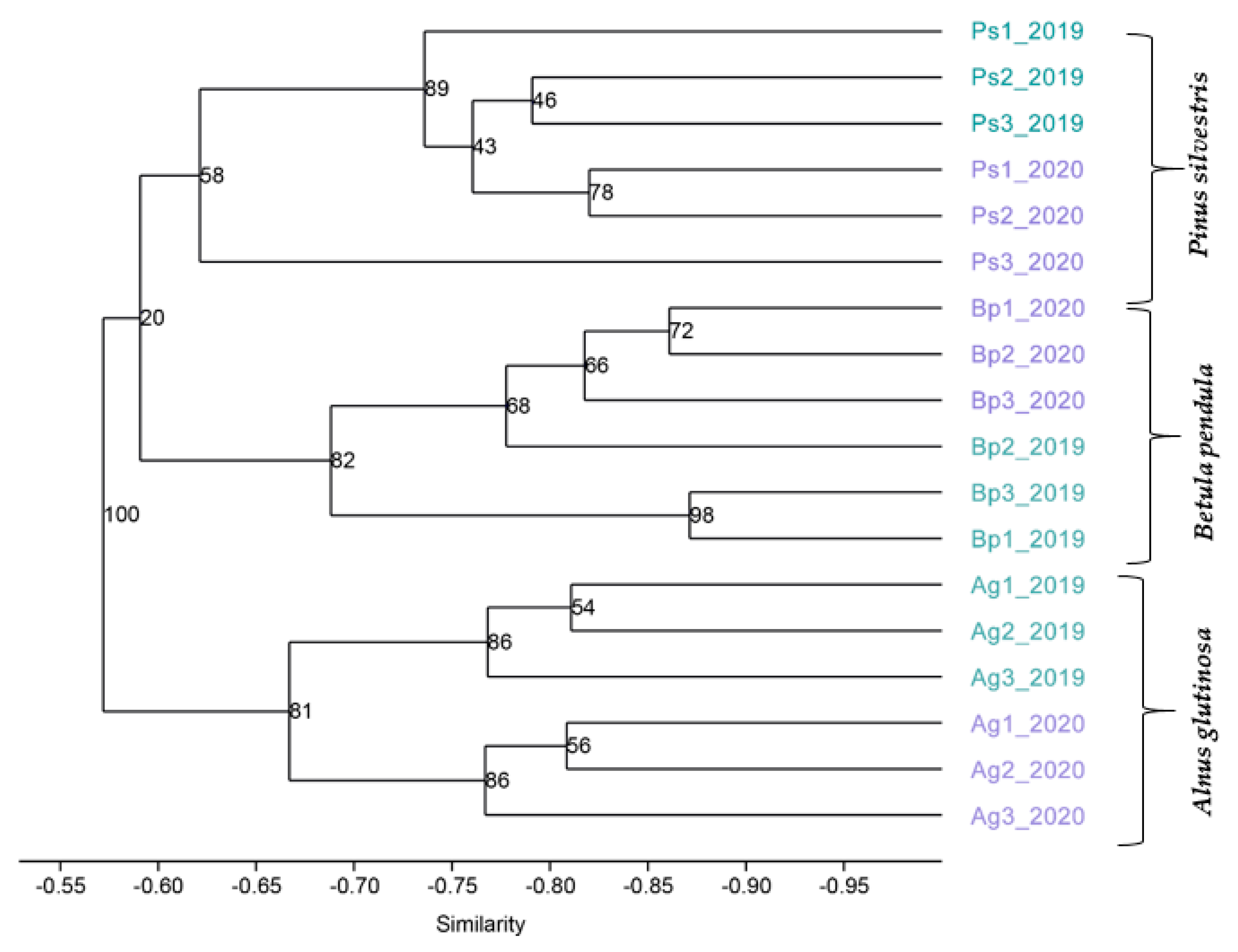
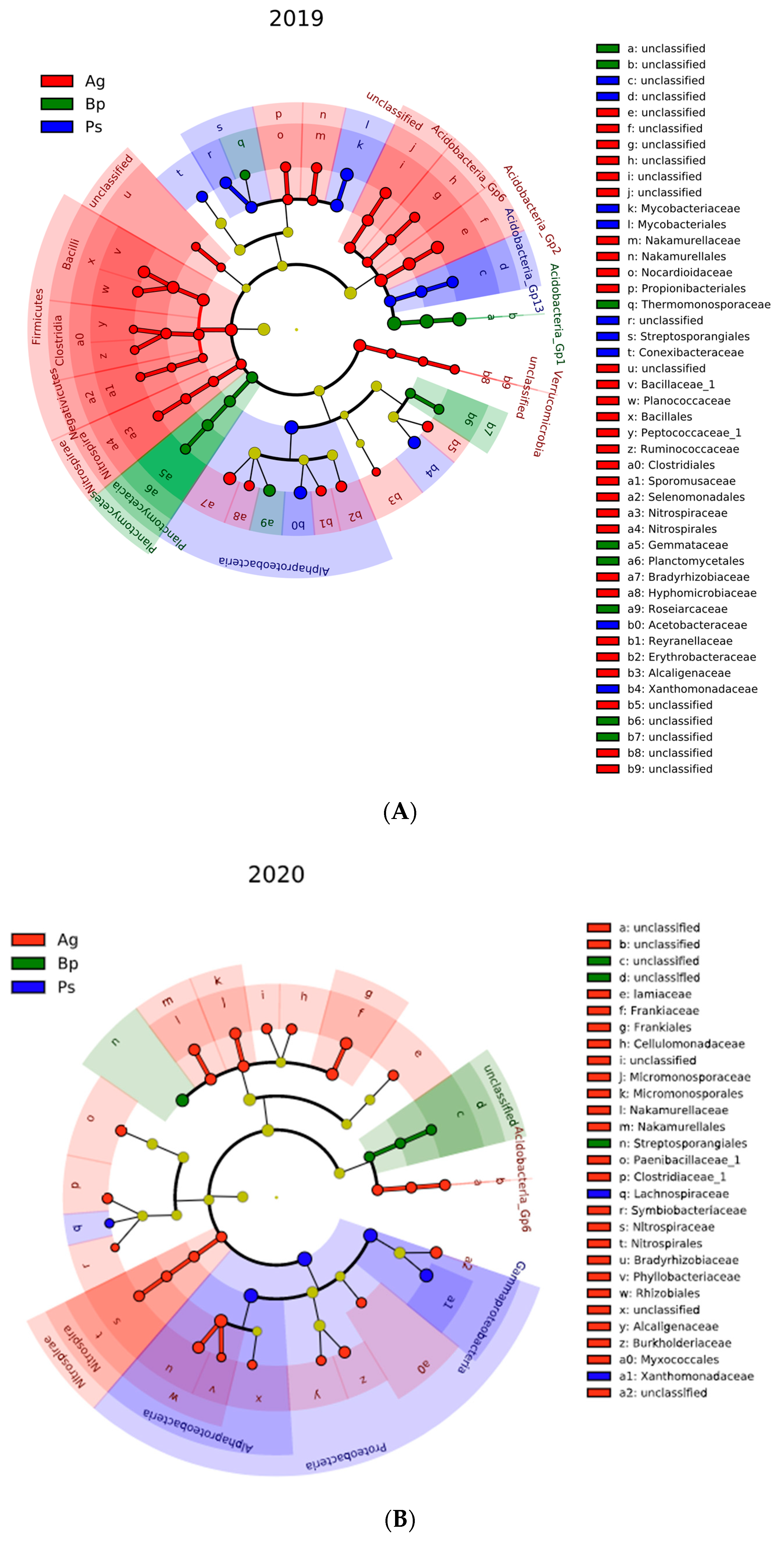
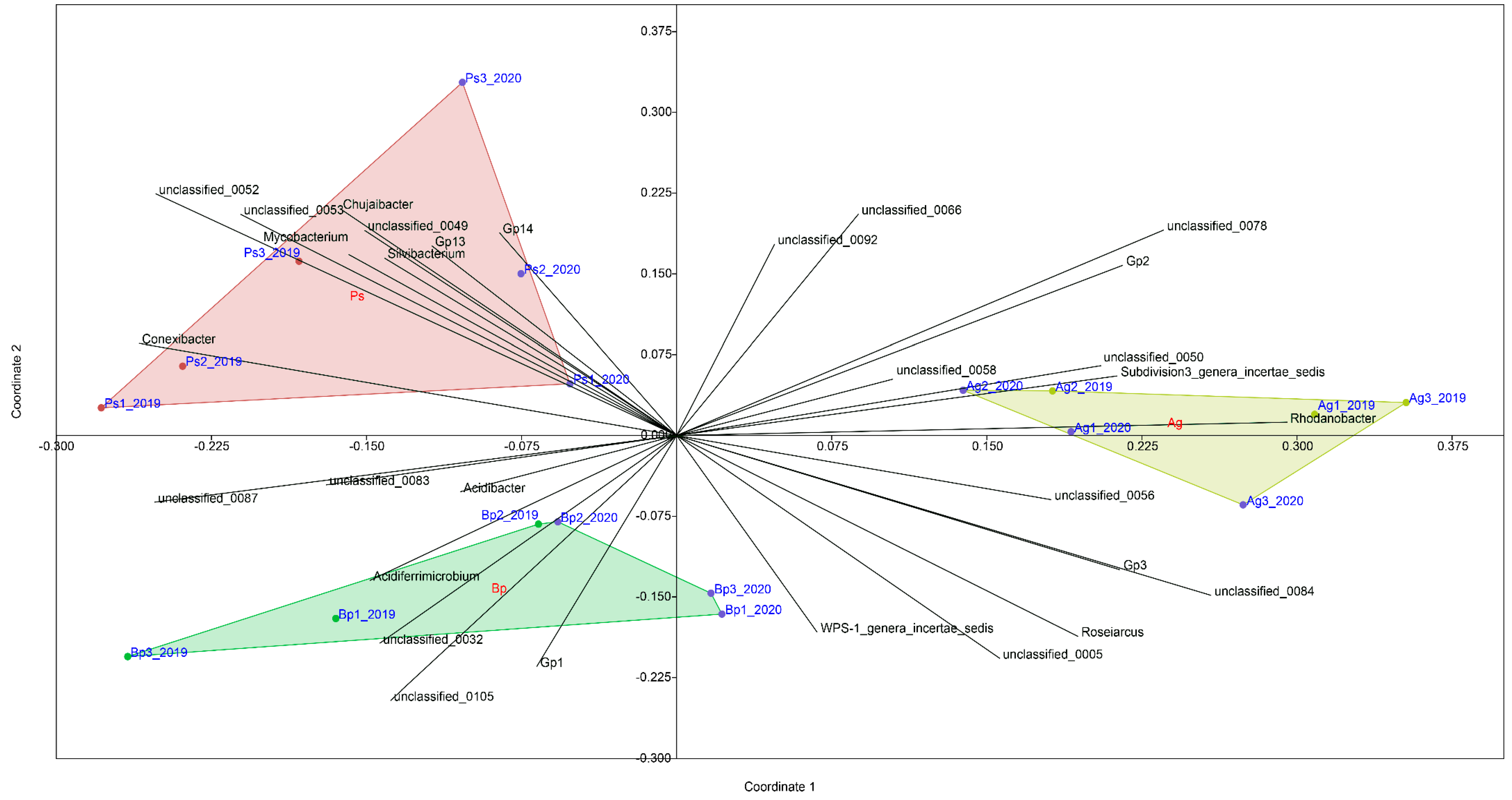
2.4. 16S rRNA Sequencing
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Soil Physico-Chemical Analysis
4.3. Enzymatic Activities
4.4. Biolog EcoPlates
4.5. DNA Extraction, Amplification, and Next-Generation Sequencing
4.6. Bioinformatic and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, Ł.; Buma, B.; Šamonil, P.; Kvaček, J.; Gałązka, A.; Kohout, P.; Malik, I. Impact of trees and forests on the Devonian landscape and weathering processes with implications to the global Earth’s system properties—A critical review. Earth-Sci. Rev. 2021, 205, 103200. [Google Scholar] [CrossRef]
- Brożek, S.; Zwydak, M.; Lasota, J.; Różański, W. Założenia metodyczne badań związków między glebą a zespołami roślinnymi w lasach. Rocz. Glebozn. 2011, 62, 16–38. (In Polish) [Google Scholar]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Abramczyk, K.; Gałązka, A. Różnorodność mikroorganizmów glebowych obszarów chronionych i o znaczących walorach przyrodniczych. Sylwan 2017, 161, 496–503. (In Polish) [Google Scholar]
- Buée, M.; de Boer, W.; Martin, F.; van Overbeek, L.; Jurkevitch, E. The rhizosphere zoo: An overview of plant-associated communities of microorganisms, including phages, bacteria, archaea, and fungi, and of some of their structuring factors. Plant Soil 2009, 321, 189–212. [Google Scholar] [CrossRef]
- Chemidlin, A.; Prevost-Boure, N.; Maron, P.-A.; Ranjard, L.; Nowak, V.; Dufrene, E.; Damesin, C.; Soudani, K.; Lata, J.-C. Seasonal dynamics of the bacterial community in forest soils under different quantities of leaf litter. Appl. Soil Ecol. 2011, 47, 14–23. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Singh, E.; Dennis, P.G.; Pieterse, C.M.J.; Schenk, P.M. Inner plant values: Diversity, colonization and benefits from endophytic bacteria. Front. Microbiol. 2017, 19, 2552. [Google Scholar] [CrossRef]
- Baldrian, P. Forest microbiome: Diversity, complexity and dynamics. nFEMS Microbiol. Rev. 2017, 41, 109–130. [Google Scholar] [CrossRef] [Green Version]
- Abdul Rahman, N.S.N.; Abdul Hamid, N.W.; Nadarajah, K. Effects of Abiotic Stress on Soil Microbiome. Int. J. Mol. Sci. 2021, 22, 9036. [Google Scholar] [CrossRef]
- Bergkemper, F.; Scholer, A.; Engel, M.; Lang, F.; Kruger, J.; Schloter, M.; Schulz, S. Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 2016, 18, 1988–2000. [Google Scholar] [CrossRef]
- Habiyaremye, J.D.; Goldmann, K.; Reitz, T.; Herrmann, S.; Buscot, F. Tree Root Zone Microbiome: Exploring the Magnitude of Environmental Conditions and Host Tree Impact. Front. Microbiol. 2020, 11, 749. [Google Scholar] [CrossRef]
- Kim, M.; Kim, W.S.; Tripathi, B.M.; Adams, J. Distinct bacterial communities dominate tropical and temperate zone leaf litter. Microb. Ecol. 2014, 67, 837–848. [Google Scholar] [CrossRef]
- Kuźniar, A.; Włodarczyk, K.; Grządziel, J.; Goraj, W.; Gałązka, A.; Wolińska, A. Culture-independent analysis of an endophytic core microbiome in two species of wheat: Triticum aestivum L. (cv. ‘Hondia’) and the first report of microbiota in Triticum spelta L. (cv. ‘Rokosz’). Syst. Appl. Microbiol. 2019, 43, 126025. [Google Scholar] [CrossRef]
- Uroz, S.; Courty, P.E.; Pierrat, J.C.; Peter, M.; Buéé, M.; Turpault, M.P.; Garbaye, J.; Frey-Klett, P. Functional profiling and distribution of the forest soil bacterial communities along the soil mycorrhizosphere continuum. Microb. Ecol. 2013, 66, 404–415. [Google Scholar] [CrossRef]
- Štursová, M.; Bárta, J.; Šantrùcková, H.; Baldrian, P. Small-scale spatial heterogeneity of ecosystem properties, microbial community composition and microbial activities in a temperate mountain forest soil. FEMS Microbiol. Ecol. 2016, 92, fiw185. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [Green Version]
- Furtak, K.; Grządziel, J.; Gałązka, A.; Niedźwiecki, J. Analysis of Soil Properties, Bacterial Community Composition, and Metabolic Diversity in Fluvisols of a Floodplain Area. Sustainability 2019, 11, 3929. [Google Scholar] [CrossRef] [Green Version]
- Gałązka, A.; Grzęda, E.; Jończyk, K. Changes of Microbial Diversity in Rhizosphere Soils of New Quality Varieties of Winter Wheat Cultivation in Organic Farming. Sustainability 2019, 11, 4057. [Google Scholar] [CrossRef] [Green Version]
- Lasota, J.; Błońska, E.; Babiak, T.; Piaszczyk, W.; Stępniewska, H.; Jankowiak, R.; Boroń, P.; Lenart-Boroń, A. Effect of Charcoal on the Properties, Enzyme Activities and Microbial Diversity of Temperate Pine Forest Soils. Forests 2021, 12, 1488. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Cong, J.; Yang, Y.; Liu, X.; Lu, H.; Liu, X.; Zhou, J.; Li, D.; Yin, H.; Ding, J.; Zhang, Y. Analyses of soil microbial community compositions and functional genes reveal potential consequences of natural forest succession. Sci. Rep. 2015, 5, 10007. [Google Scholar] [CrossRef] [Green Version]
- Xu, J. Metagenomics and Ecosystems Biology: Conceptual Frameworks, Tools and Methods (w) Metagenomics. Theory, Methods and Applications, Red; Marco, D., Ed.; Caister Academic Press: Norfolk, UK, 2010; pp. 1–14. [Google Scholar]
- Lahti, L.; Shetty, S. Tools for Microbiome Analysis in R. 2017. Available online: http://microbiome.github.com/microbiome (accessed on 19 July 2021).
- Lebeis, S.L. Greater than the sum of their parts: Characterizing plant microbiomes at the community-level. Curr. Opin. Plant Biol. 2015, 24, 82–86. [Google Scholar] [CrossRef]
- Levy-Booth, D.J.; Prescott, C.E.; Grayston, S.J. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 2014, 75, 11–25. [Google Scholar] [CrossRef]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.; Gilbert, J.; Meyer, F. Metagenomics—A guide from sampling to data analysis. BMC Microb. 2012, Inf. Exp. 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Torres-Cortés, G.; Millán, V.; Ramírez-Saad, H.C.; Nisa-Martínez, R.; Toro, N.; Martínez-Abarca, F. Characterization of novel antibiotic resistance genes identified by functional metagenomics on soil samples. Environ. Microbiol. 2011, 13, 1101–1114. [Google Scholar] [CrossRef] [PubMed]
- PN-ISO 10381-6; Quality of Soil Collected Samples—Principles of Collected and Kept of Soil Samples to the Microbiological Research in Laboratory Conditions. Polish Standard. ISO: Geneva, Switzerland, 1998.
- Kononowa, M. Substancje Organiczne Gleby ich Budowa, Właściwości i Metody Badań; Wydawnictwo Rolnicze i Leśne Warszawa: Warsaw, Poland, 1968. (In Polish) [Google Scholar]
- Kędziora, A. Przyrodnicze podstawy ochrony ekosystemów rolniczych. Frag. Agronom. 2007, 3, 213–223. (In Polish) [Google Scholar]
- Yadav, A.N.; Verma, P.; Singh, B.; Chauhan, V.S.; Suman, A.; Saxena, A.K. Plant growth promoting bacteria: Biodiversity and multifunctional attributes for sustainable agriculture. Adv. Biotechnol. Microbiol. 2017, 5, 1–16. [Google Scholar] [CrossRef]
- Baldrian, P.; Kolarˇík, M.; Štursová, M.; Kopecký, J.; Valaskova, V.; Větrovský, T.; Žifčáková, L.; Šnajdr, J.; Rídl, J.; Vlček, C.; et al. Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 2012, 6, 248–258. [Google Scholar] [CrossRef] [Green Version]
- Brzostek, E.R.; Greco, A.; Drake, J.E.; Finzi, A.C. Root carbon inputs to the rhizosphere stimulate extracellular enzyme activity and increase nitrogen availability in temperate forest soil. Biogeochemistry 2012, 115, 65–76. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A. The rhizosphere microbiome and its beneficial effects on plants. Postępy Mikrobiol. 2019, 58, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; Baldrian, P. Community-level physiological profiling analyses show potential to identify the copiotrophic bacteria present in soil environments. PLoS ONE 2017, 12, e0171638. [Google Scholar] [CrossRef]
- Grządziel, J.; Furtak, K.; Gałązka, A. Community-level physiological profiles of microorganisms from different types of soil characteristic to Poland—A long-term microplot experiment. Sustainability 2019, 11, 56. [Google Scholar] [CrossRef] [Green Version]
- Wolińska, A.; Kuźniar, A.; Gałązka, A. Biodiversity in the Rhizosphere of Selected Winter Wheat (Triticum aestivum L.) Cultivars—Genetic and Catabolic Fingerprinting. Agronomy 2020, 10, 953. [Google Scholar] [CrossRef]
- Woźniak, M.; Gałązka, A.; Tyśkiewicz, R.; Jaroszuk-Ściseł, J. Endophytic Bacteria Potentially Promote Plant Growth by Synthesizing Different Metabolites and their Phenotypic/Physiological Profiles in the Biolog GEN III MicroPlateTM Test. Int. J. Mol. Sci. 2019, 20, 5283. [Google Scholar] [CrossRef] [Green Version]
- Si, P.; Shao, W.; Yu, H.; Yang, X.; Gao, D.; Qiao, X.; Wang, Z.; Wu, G. Rhizosphere Microenvironments of Eight Common Deciduous Fruit Trees Were Shaped by Microbes in Northern China. Front. Microbiol. 2018, 9, 3147. [Google Scholar] [CrossRef]
- Thiem, D.; Gołębiewski, M.; Hulisz, P.; Piernik, A.; Hrynkiewicz, K. How Does Salinity Shape Bacterial and Fungal Microbiomes of Alnus glutinosa Roots? Front. Microbiol. 2018, 9, 651. [Google Scholar] [CrossRef]
- Rosier, C.L.; Polson, S.W.; D’Amico, V., III; Kan, J.; Trammell, T.L.E. Urbanization pressures alter tree rhizosphere microbiomes. Sci. Rep. 2021, 11, 9447. [Google Scholar] [CrossRef]
- Gałazka, A.; Niedźwiecki, J.; Grządziel, J.; Gawryjołek, K. Evaluation of Changes in Glomalin-Related Soil Proteins (GRSP) Content, Microbial Diversity and Physical Properties Depending on the Type of Soil as the Important Biotic Determinants of Soil Quality. Agronomy 2020, 10, 1279. [Google Scholar] [CrossRef]
- Warren, D.A.; Cheng, L.; Wang, P. Soil acid and alkaline phosphatase activity as pH adjustment indicators. Soil Biol. Biochem. 2020, 32, 1915–1919. [Google Scholar] [CrossRef]
- Margalef, O.; Sardans, J.; Fernández-Martínez, M.; Molowny-Horas, R.; Janssens, I.A.; Ciais, P.; Goll, D.; Richter, A.; Obersteiner, M.; Asensio, D.; et al. Global patterns of phosphatase activity in natural soils. Sci. Rep. 2017, 7, 1337. [Google Scholar] [CrossRef] [Green Version]
- Lemanowicz, A.; Bartkowiak, A. Changes in the activity of phosphatase and the content of phosphorus in salt-affected soils grassland habitat Natura 2000. Pol. J. Soil Sci. 2016, 49, 149. [Google Scholar] [CrossRef] [Green Version]
- Kamutando, C.N.; Vikram, S.; Kamgan-Nkuekam, G.; Makhalanyane, T.P.; Greve, M.; Le Roux, J.J.; Richardson, D.M.; Cowan, D.A.; Valverde, A. The Functional Potential of the Rhizospheric Microbiome of an Invasive Tree Species, Acacia dealbata. Microb. Ecol. 2019, 77, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Marupakula, S.; Mahmood, S.; Finlay, R.D. Analysis of single root tip microbiomes suggests that distinctive bacterial communities are selected by Pinus sylvestris roots colonized by different ectomycorrhizal fungi. Environ. Microbiol. 2016, 18, 1470–1483. [Google Scholar] [CrossRef]
- Izumi, H.; Anderson, I.C.; Alexander, I.J.; Killham, K.; Moore, E.R. Endobacteria in some ectomycorrhiza of Scots pine (Pinus sylvestris). FEMS Microbiol. Ecol. 2006, 56, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Pinho, D.; Barroso, C.; Froufe, H.; Brown, N.; Vanguelova, E.; Egas, C.; Denman, S. Linking Tree Health, Rhizosphere Physicochemical Properties, and Microbiome in Acute Oak Decline. Forests 2020, 11, 1153. [Google Scholar] [CrossRef]
- Casida, L.E.J.; Klein, D.A.; Santoro, T. Soil dehydrogenases activity. Soil Sei. 1964, 98, 371–376. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphate activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Gałązka, A.; Gawryjołek, K.; Perzyński, A.; Gałązka, R.; Księżak, J. Changes of enzymatic activities and microbial communities in soil under long-term maize monoculture and crop rotation. Pol. J. Environ. 2017, 26, 39–46. [Google Scholar] [CrossRef]
- Grządziel, J.; Gałązka, A. Microplot long-term experiment reveals strong soil type influence on bacteria composition and its functional diversity. Appl. Soil Ecol. 2017. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Wright, E.S. RDP v16 Modified Training Set for 16S rRNA Classification, 2019, [WWW Document]. (Udostępniono 2.1.19). Available online: http://www2.decipher.codes/Classification/TrainingSets/RDP_v16-mod_March2018.RData (accessed on 19 July 2021).
- Murali, A.; Bhargava, A.; Wright, E.S. IDTAXA: A novel approach for accurate taxonomic classification of microbiome sequences. Microbiome 2018, 6, 140. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree | Symbol | Soil Texture Class (USDA) * | Particle Size Distribution [%] | Corg (%) | Humus (%) | Ctot (%) | Ntot (%) | C/N | P | K | Mg | pH | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sand (2.0–0.05 mm) | Silt (0.05–0.002 mm) | Clay (<0.002 mm) | ||||||||||||
| Betula pendula | Bp1 | sand | 96.62 | 3.36 | 0.02 | 1.830 | 3.155 | 2.121 | 0.173 | 12.260 | 6.979 | 6.354 | 1.640 | 3.235 |
| Bp2 | sand | 95.68 | 4.12 | 0.19 | 1.376 | 2.372 | 1.587 | 0.127 | 12.492 | 5.532 | 4.907 | 0.890 | 3.225 | |
| Bp3 | sand | 95.73 | 4.17 | 0.09 | 1.357 | 2.339 | 1.536 | 0.131 | 11.770 | 10.192 | 5.692 | 1.330 | 3.190 | |
| Alnus glutinosa | Ag1 | sand | 91.85 | 8.00 | 0.16 | 0.988 | 1.703 | 1.133 | 0.065 | 17.502 | 3.183 | 1.290 | 0.380 | 3.500 |
| Ag2 | sand | 93.52 | 6.45 | 0.02 | 1.093 | 1.884 | 1.236 | 0.071 | 17.473 | 6.170 | 1.014 | 0.300 | 3.190 | |
| Ag3 | sand | 92.98 | 7.00 | 0.02 | 0.640 | 1.103 | 0.716 | 0.039 | 18.346 | 5.478 | 1.086 | 0.350 | 3.710 | |
| Pinus sylvestris | Ps1 | sand | 95.58 | 4.40 | 0.02 | 1.157 | 1.994 | 1.284 | 0.080 | 16.044 | 1.115 | 1.553 | 0.810 | 3.080 |
| Ps2 | sand | 92.94 | 6.54 | 0.52 | 0.999 | 1.723 | 1.117 | 0.068 | 16.419 | 1.458 | 1.280 | 0.400 | 3.020 | |
| Ps3 | sand | 93.64 | 5.86 | 0.50 | 1.251 | 2.157 | 1.422 | 0.088 | 16.251 | 1.352 | 1.643 | 0.280 | 2.985 | |
| Sample | DHA | AlP | AcP |
|---|---|---|---|
| 2019 | |||
| Bp1 | 6.283 b ± 0.705 | 9.535 b ± 0.298 | 41.633 b ± 2.033 |
| Bp2 | 8.044 b ± 0.806 | 9.337 b ± 0.346 | 41.442 b ± 3.828 |
| Bp3 | 5.262 b ± 0.305 | 8.611 b ± 0.704 | 35.007 b ± 0.388 |
| Ag1 | 51.504 a ± 2.618 | 16.113 a ± 0.396 | 96.383 a ± 2.454 |
| Ag2 | 35.676 a ± 2.835 | 14.667 a ± 0.409 | 79.701 a ± 2.715 |
| Ag3 | 26.270 a ± 1.504 | 13.359 a ± 0.280 | 82.302 a ± 6.598 |
| Ps1 | 0.931 c ± 0.115 | 7.874 c ± 0.856 | 16.934 c ± 1.614 |
| Ps2 | 1.151 c ± 0.119 | 6.591 c ± 0.357 | 18.471 c ± 1.016 |
| Ps3 | 0.607 c ± 0.005 | 6.670 c ± 0.417 | 13.778 c ± 1.120 |
| 2020 | |||
| Bp1 | 12.064 b ± 0.649 | 11.513 b ± 1.089 | 38.436 b ± 2.565 |
| Bp2 | 12.027 b ± 0.617 | 10.585 b ± 1.300 | 44.292 b ± 1.347 |
| Bp3 | 9.473 b ± 0.725 | 10.142 b ± 0.269 | 37.711 b ± 2.937 |
| Ag1 | 25.067 c ± 0.427 | 15.077 a ± 0.697 | 81.241 a ± 5.076 |
| Ag2 | 19.952 c ± 0.320 | 14.232 a ± 0.136 | 80.230 a ± 1.743 |
| Ag3 | 24.525 c ± 0.206 | 13.109 a ± 0.443 | 74.925 a ± 1.951 |
| Ps1 | 1.875 c ± 0.118 | 7.085 c ± 0.567 | 16.252 c ± 0.695 |
| Ps2 | 1.145 c ± 0.118 | 7.740 c ± 0.256 | 15.009 c ± 1.023 |
| Ps3 | 1.377 c ± 0.061 | 6.704 c ± 0.211 | 14.571 c ± 1.567 |
| Sample ID | H’ | R | E | AWCD590 |
|---|---|---|---|---|
| 2019 | ||||
| Bp1 | 3.148 b ± 0.016 | 26.000 b ± 0.577 | 0.986 b ± 0.005 | 1.342 b ± 0.057 |
| Bp2 | 3.052 b ± 0.031 | 23.333 b ± 0.377 | 0.979 b ± 0.006 | 1.345 b ± 0.133 |
| Bp3 | 3.122 b ± 0.028 | 23.000 b ± 1.732 | 0.975 b ± 0.018 | 1.310 b ± 0.069 |
| Ag1 | 3.355 a ± 0.021 | 30.000 a ± 0.377 | 0.957 c ± 0.006 | 1.873 a ± 0.048 |
| Ag2 | 3.374 a ± 0.013 | 30.333 a ± 0.577 | 0.959 c ± 0.002 | 1.997 a ± 0.080 |
| Ag3 | 3.379 a ± 0.013 | 30.333 a ± 0.577 | 0.960 c ± 0.002 | 1.935 a ± 0.060 |
| Ps1 | 2.609 c ± 0.033 | 16.333 c ± 1.082 | 1.045 a ± 0.039 | 0.424 c ± 0.030 |
| Ps2 | 2.735 c ± 0.003 | 18.333 c ± 0.577 | 0.992 a ± 0.009 | 0.763 c ± 0.036 |
| Ps3 | 2.921 b ± 1.128 | 17.000 c ± 1.000 | 1.032 a ± 0.049 | 0.403 c ± 0.064 |
| 2020 | ||||
| Bp1 | 3.144 b ± 0.025 | 25.333 b ± 1.155 | 0.973 b ± 0.010 | 1.338 b ± 0.061 |
| Bp2 | 3.138 b ± 0.042 | 22.000 b ± 1.732 | 0.984 b ± 0.012 | 1.363 b ± 0.084 |
| Bp3 | 2.993 b ± 0.010 | 23.667 b ± 0.577 | 0.988 b ± 0.006 | 1.278 b ± 0.072 |
| Ag1 | 3.361 a ± 0.019 | 29.667 a ± 0.577 | 0.962 c ± 0.006 | 1.792 a ± 0.173 |
| Ag2 | 3.292 a ± 0.006 | 28.333 a ± 0.577 | 0.954 c ± 0.006 | 1.752 a ± 0.021 |
| Ag3 | 3.344 a ± 0.019 | 29.667 a ± 0.577 | 0.977 c ± 0.001 | 1.832 a ± 0.075 |
| Ps1 | 2.607 c ± 0.026 | 16.667 c ± 1.528 | 0.992 a ± 0.012 | 0.539 c ± 0.055 |
| Ps2 | 2.740 c ± 0.115 | 15.667 c ± 0.528 | 0.998 a ± 0.066 | 0.776 c ± 0.076 |
| Ps3 | 3.005 b ± 0.031 | 18.333 c ± 1.528 | 0.964 a ± 0.012 | 0.660 c ± 0.039 |
| Sample ID | ASVs | Classified ASVs (%) | Sequence Read Archive (SRA) NCBI ID |
|---|---|---|---|
| 2019 | |||
| Bp1 | 100 | 59 | SRR16926273 |
| Bp2 | 123 | 62 | SRR16926272 |
| Bp3 | 88 | 59 | SRR16926271 |
| Ag1 | 227 | 66 | SRR16926270 |
| Ag2 | 183 | 64 | SRR16926269 |
| Ag3 | 187 | 64 | SRR16926267 |
| Ps1 | 87 | 62 | SRR16926266 |
| Ps2 | 79 | 61 | SRR16926265 |
| Ps3 | 178 | 65 | SRR16926264 |
| 2020 | |||
| Bp1 | 106 | 55 | SPR16962104 |
| Bp2 | 106 | 57 | SPR16962103 |
| Bp3 | 111 | 57 | SPR16962102 |
| Ag1 | 172 | 65 | SPR16962101 |
| Ag2 | 174 | 61 | SPR16962100 |
| Ag3 | 225 | 65 | SPR16962098 |
| Ps1 | 85 | 60 | SPR16962097 |
| Ps2 | 105 | 64 | SPR16962096 |
| Ps3 | 46 | 52 | SPR16962095 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gałązka, A.; Marzec-Grządziel, A.; Varsadiya, M.; Niedźwiecki, J.; Gawryjołek, K.; Furtak, K.; Przybyś, M.; Grządziel, J. Biodiversity and Metabolic Potential of Bacteria in Bulk Soil from the Peri-Root Zone of Black Alder (Alnus glutinosa), Silver Birch (Betula pendula) and Scots Pine (Pinus sylvestris). Int. J. Mol. Sci. 2022, 23, 2633. https://doi.org/10.3390/ijms23052633
Gałązka A, Marzec-Grządziel A, Varsadiya M, Niedźwiecki J, Gawryjołek K, Furtak K, Przybyś M, Grządziel J. Biodiversity and Metabolic Potential of Bacteria in Bulk Soil from the Peri-Root Zone of Black Alder (Alnus glutinosa), Silver Birch (Betula pendula) and Scots Pine (Pinus sylvestris). International Journal of Molecular Sciences. 2022; 23(5):2633. https://doi.org/10.3390/ijms23052633
Chicago/Turabian StyleGałązka, Anna, Anna Marzec-Grządziel, Milan Varsadiya, Jacek Niedźwiecki, Karolina Gawryjołek, Karolina Furtak, Marcin Przybyś, and Jarosław Grządziel. 2022. "Biodiversity and Metabolic Potential of Bacteria in Bulk Soil from the Peri-Root Zone of Black Alder (Alnus glutinosa), Silver Birch (Betula pendula) and Scots Pine (Pinus sylvestris)" International Journal of Molecular Sciences 23, no. 5: 2633. https://doi.org/10.3390/ijms23052633
APA StyleGałązka, A., Marzec-Grządziel, A., Varsadiya, M., Niedźwiecki, J., Gawryjołek, K., Furtak, K., Przybyś, M., & Grządziel, J. (2022). Biodiversity and Metabolic Potential of Bacteria in Bulk Soil from the Peri-Root Zone of Black Alder (Alnus glutinosa), Silver Birch (Betula pendula) and Scots Pine (Pinus sylvestris). International Journal of Molecular Sciences, 23(5), 2633. https://doi.org/10.3390/ijms23052633