
Thidiazuron Promotes Leaf Abscission by Regulating the Crosstalk Complexities between Ethylene, Auxin, and Cytokinin in Cotton

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Thidiazuron Induces Transcriptome Change in Leaf, Petiole, and AZ of Cotton
2.2. Thidiazuron Regulates the Expression of Plant Hormone Related Genes in Leaf, Petiole, and AZ
2.3. Thidiazuron Regulates the Gene Expression of Ethylene Metabolism and Signaling
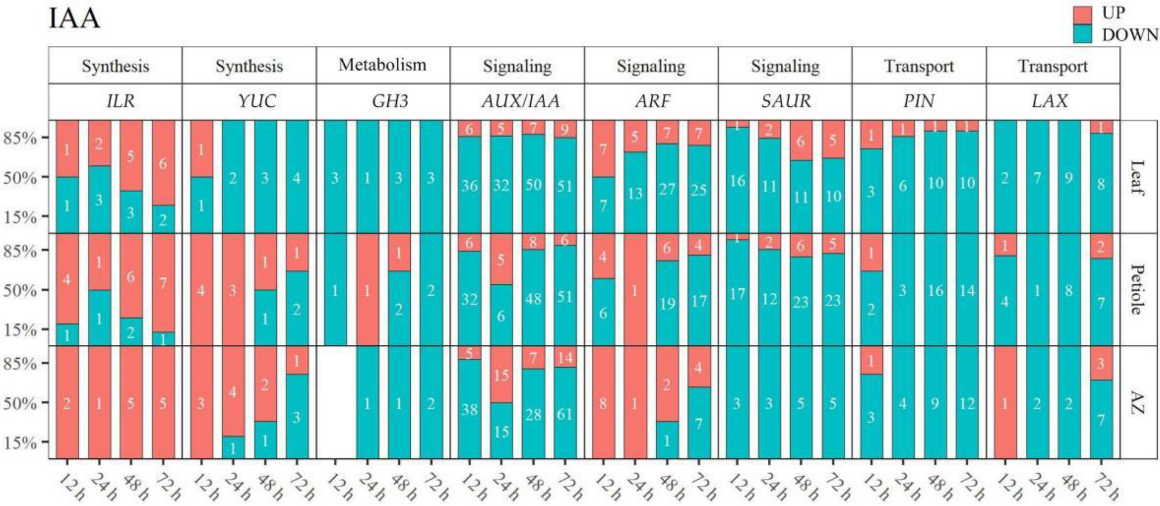
2.4. Thidiazuron Regulates the Gene Expression of IAA Metabolism, Transport, and Signal
2.5. Thidiazuron Regulates the Gene Expression Related to CTK and ABA
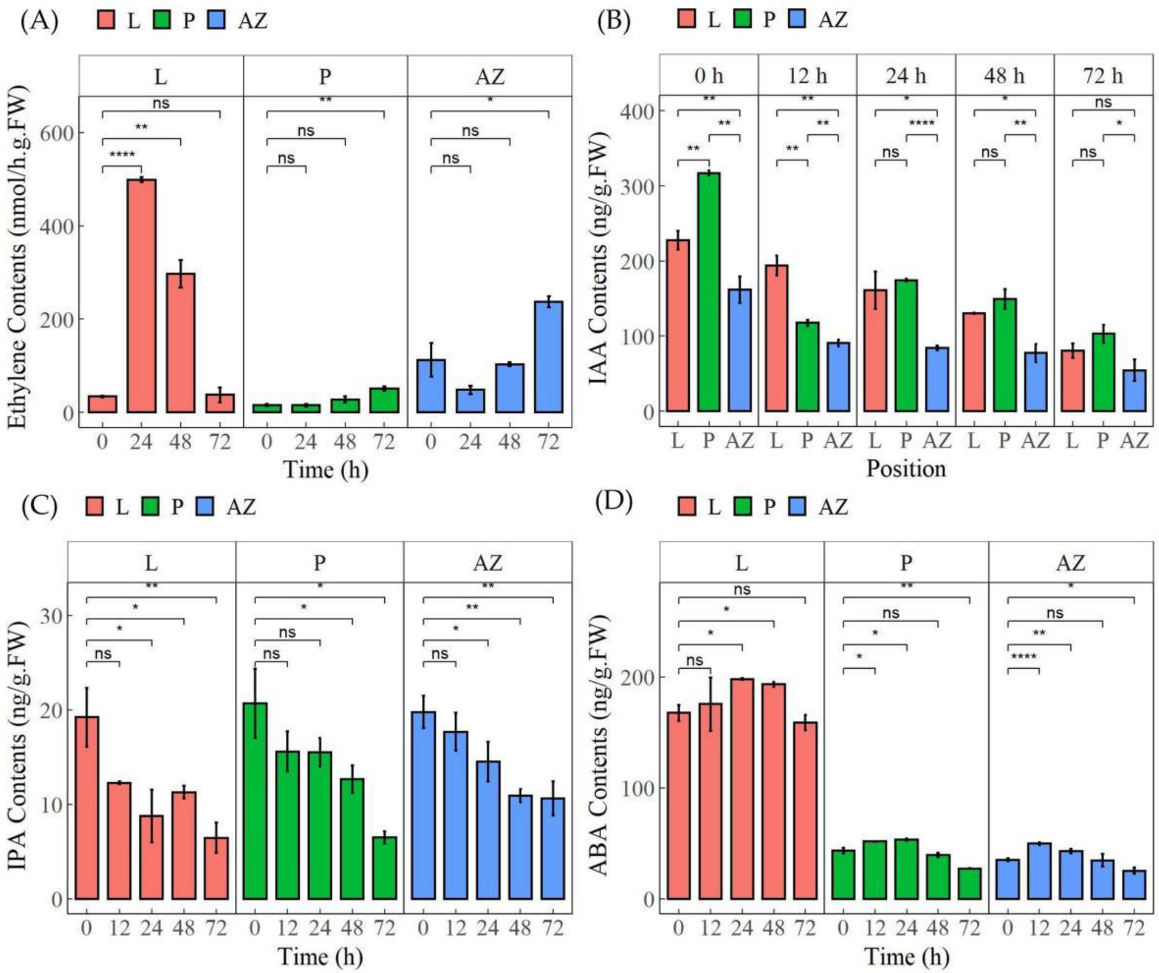
2.6. Thidiazuron Modulates the Content of Multiple Plant Hormones
2.7. Thidiazuron Regulates the Gene Expression of Cell Cycle and Cell Wall in Abscission Zone
2.8. Thidiazuron Regulates the Gene Expression of Transcript Factors in Abscission Zone
3. Discussion
3.1. Thidiazuron Promotes Cotton Leaf Abscission by Activating Gene Expression of Ethylene Synthesis and Signaling
3.2. Thidiazuron Promotes Cotton Leaf Abscission by Regulating the Gene Expression of Auxin Synthesis, Transport, and Signaling
3.3. Thidiazuron Regulates Genes Expression of CTK Synthesis as Well as ABA Synthesis and Signaling
3.4. The Balance and Synergism of Ethylene, IAA, and Cytokinin Regulates the Cotton Leaf Abscission
3.5. Differential Expression of Transcription Factors in Different Stages of Abscission Process
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Experimental Design
4.3. RNA Extraction, cDNA Library Preparation and Sequencing for RNA-Seq
4.4. Statistical Analysis of RNA-Seq
4.5. Quantitative Reverse Transcription PCR (RT-qPCR) Analysis
4.6. Analysis of Differential Gene Expression and KEGG Enrichment
4.7. Measurement of Endogenous Hormonal Contents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gulfishan, M.; Jahan, A.; Bhat, T.A.; Sahab, D. Chapter 16—Plant Senescence and Organ Abscission. In Senescence Signalling and Control in Plants; Sarwat, M., Tuteja, N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 255–272. ISBN 978-0-12-813187-9. [Google Scholar]
- Patharkar, O.R.; Walker, J.C. Connections between Abscission, Dehiscence, Pathogen Defense, Drought Tolerance, and Senescence. Plant Sci. 2019, 284, 25–29. [Google Scholar] [CrossRef]
- Patharkar, O.R.; Walker, J.C. Advances in Abscission Signaling. J. Exp. Bot. 2018, 69, 733–740. [Google Scholar] [CrossRef] [PubMed]
- McKim, S.M.; Stenvik, G.E.; Butenko, M.A.; Kristiansen, W.; Cho, S.K.; Hepworth, S.R.; Aalen, R.B.; Haughn, G.W. The BLADE-ON-PETIOLE Genes Are Essential for Abscission Zone Formation in Arabidopsis. Development 2008, 135, 1537–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinyopich, A.; Ditta, G.S.; Savidge, B.; Liljegren, S.J.; Baumann, E.; Wisman, E.; Yanofsky, M.F. Assessing the Redundancy of MADS-Box Genes during Carpel and Ovule Development. Nature 2003, 424, 85–88. [Google Scholar] [CrossRef]
- Liu, D.; Wang, D.; Qin, Z.; Zhang, D.; Yin, L.; Wu, L.; Colasanti, J.; Li, A.; Mao, L. The SEPALLATA MADS-Box Protein SLMBP21 Forms Protein Complexes with JOINTLESS and MACROCALYX as a Transcription Activator for Development of the Tomato Flower Abscission Zone. Plant J. 2014, 77, 284–296. [Google Scholar] [CrossRef]
- Adamczyk, B.J.; Lehti-Shiu, M.D.; Fernandez, D.E. The MADS Domain Factors AGL15 and AGL18 Act Redundantly as Repressors of the Floral Transition in Arabidopsis. Plant J. 2007, 50, 1007–1019. [Google Scholar] [CrossRef]
- Shi, C.L.; Stenvik, G.E.; Vie, A.K.; Bones, A.M.; Pautot, V.; Proveniers, M.; Aalen, R.B.; Butenko, M.A. Arabidopsis Class I KNOTTED-Like Homeobox Proteins Act Downstream in the IDA-HAE/HSL2 Floral Abscission Signaling Pathway. Plant Cell 2011, 23, 2553–2567. [Google Scholar] [CrossRef] [Green Version]
- Tucker, M.L.; Yang, R. IDA-like Gene Expression in Soybean and Tomato Leaf Abscission and Requirement for a Diffusible Stelar Abscission Signal. AoB Plants 2012, 2012, pls035. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Sundaresan, S.; Philosoph-Hadas, S.; Yang, R.; Meir, S.; Tucker, M.L. Examination of the Abscission-Associated Transcriptomes for Soybean, Tomato, and Arabidopsis Highlights the Conserved Biosynthesis of an Extensible Extracellular Matrix and Boundary Layer. Front. Plant Sci. 2015, 6, 1109. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Chun, J.P.; Tucker, M.L. Transcriptional Regulation of Abscission Zones. Plants 2019, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, N.; Nishitani, K. Cryogenian Origin and Subsequent Diversification of the Plant Cell-Wall Enzyme XTH Family. Plant Cell Physiol. 2021, 62, 1874–1889. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Anderson, C.T.; Cao, J. Polygalacturonase45 Cleaves Pectin and Links Cell Proliferation and Morphogenesis to Leaf Curvature in Arabidopsis Thaliana. Plant J. 2021, 106, 1493–1508. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Feng, C.; Wang, Y.; Sun, Y.; Peng, X.; Xiao, Y.; Zhang, X.; Zhou, X.; Jiao, J.; Wang, W.; et al. Genome-Wide Identification of the Xyloglucan Endotransglucosylase/Hydrolase (XTH) and Polygalacturonase (PG) Genes and Characterization of Their Role in Fruit Softening of Sweet Cherry. Int. J. Mol. Sci. 2021, 22, 12331. [Google Scholar] [CrossRef]
- Xu, J.; Chen, L.; Sun, H.; Wusiman, N.; Sun, W.; Li, B.; Gao, Y.; Kong, J.; Zhang, D.; Zhang, X.; et al. Crosstalk between Cytokinin and Ethylene Signaling Pathways Regulates Leaf Abscission in Cotton in Response to Chemical Defoliants. J. Exp. Bot. 2019, 70, 1525–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meir, S.; Philosoph-Hadas, S.; Riov, J.; Tucker, M.L.; Patterson, S.E.; Roberts, J.A. Re-Evaluation of the Ethylene-Dependent and -Independent Pathways in the Regulation of Floral and Organ Abscission. J. Exp. Bot. 2019, 70, 1461–1467. [Google Scholar] [CrossRef]
- Tieman, D.M.; Ciardi, J.A.; Taylor, M.G.; Klee, H.J. Members of the Tomato LeEIL (EIN3-like) Gene Family Are Functionally Redundant and Regulate Ethylene Responses throughout Plant Development. Plant J. 2001, 26, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Kućko, A.; Wilmowicz, E.; Pokora, W.; Alché, J.D.D. Disruption of the Auxin Gradient in the Abscission Zone Area Evokes Asymmetrical Changes Leading to Flower Separation in Yellow Lupine. Int. J. Mol. Sci. 2020, 21, 3815. [Google Scholar] [CrossRef]
- Liang, Y.; Jiang, C.; Liu, Y.; Gao, Y.; Lu, J.; Aiwaili, P.; Fei, Z.; Jiang, C.Z.; Hong, B.; Ma, C.; et al. Auxin Regulates Sucrose Transport to Repress Petal Abscission in Rose (Rosa Hybrida). Plant Cell 2020, 32, 3485–3499. [Google Scholar] [CrossRef]
- Marciniak, K.; Kućko, A.; Wilmowicz, E.; Świdziński, M.; Przedniczek, K.; Kopcewicz, J. Gibberellic Acid Affects the Functioning of the Flower Abscission Zone in Lupinus Luteus via Cooperation with the Ethylene Precursor Independently of Abscisic Acid. J. Plant Physiol. 2018, 229, 170–174. [Google Scholar] [CrossRef]
- Ma, X.; Li, C.; Huang, X.; Wang, H.; Wu, H.; Zhao, M.; Li, J. Involvement of HD-ZIP I Transcription Factors LcHB2 and LcHB3 in Fruitlet Abscission by Promoting Transcription of Genes Related to the Biosynthesis of Ethylene and ABA in Litchi. Tree Physiol. 2019, 39, 1600–1613. [Google Scholar] [CrossRef]
- Du, M.; Li, Y.; Tian, X.; Duan, L.; Zhang, M.; Tan, W.; Xu, D.; Li, Z. The Phytotoxin Coronatine Induces Abscission-Related Gene Expression and Boll Ripening during Defoliation of Cotton. PLoS ONE 2014, 9, e97652. [Google Scholar] [CrossRef] [Green Version]
- Leyser, O. Auxin Signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The Role of Gibberellin Signalling in Plant Responses to Abiotic Stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Singh, A.P.; Sane, A.P. Differential and Reciprocal Regulation of Ethylene Pathway Genes Regulates Petal Abscission in Fragrant and Non-Fragrant Roses. Plant Sci. 2019, 280, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of Plant Growth by Cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Fujisawa, M.; Shima, Y.; Ito, Y. Expression Profiling of Tomato Pre-Abscission Pedicels Provides Insights into Abscission Zone Properties Including Competence to Respond to Abscission Signals. BMC Plant Biol. 2013, 13, 40. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Wen, Z.; Hou, Q.; Qiao, G.; Yang, K.; Hong, Y.; Wen, X. Cross-Talk between Transcriptome, Phytohormone and HD-ZIP Gene Family Analysis Illuminates the Molecular Mechanism Underlying Fruitlet Abscission in Sweet Cherry (Prunus Avium L). BMC Plant Biol. 2021, 21, 173. [Google Scholar] [CrossRef]
- Qi, F.; Zhang, F. Cell Cycle Regulation in the Plant Response to Stress. Front. Plant Sci. 2019, 10, 1765. [Google Scholar] [CrossRef] [Green Version]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. An Apple NAC Transcription Factor Enhances Salt Stress Tolerance by Modulating the Ethylene Response. Physiol. Plant 2018, 164, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.M.; Zhang, X.W.; Liu, Y.W.; Li, K.; Tan, Q.; Zhou, S.; Wang, G.; Zhou, C.J. A WRKY Transcription Factor, TaWRKY42-B, Facilitates Initiation of Leaf Senescence by Promoting Jasmonic Acid Biosynthesis. BMC Plant Biol. 2020, 20, 444. [Google Scholar] [CrossRef]
- Huang, R.; Liu, D.; Huang, M.; Ma, J.; Li, Z.; Li, M.; Sui, S. CpWRKY71, a WRKY Transcription Factor Gene of Wintersweet (Chimonanthus Praecox), Promotes Flowering and Leaf Senescence in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 5325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhao, Q.; Zeng, D.; Xu, J.; Zhou, H.; Wang, F.; Ma, N.; Li, Y. RhMYB108, an R2R3-MYB Transcription Factor, Is Involved in Ethylene- and JA-Induced Petal Senescence in Rose Plants. Hortic. Res. 2019, 6, 131. [Google Scholar] [CrossRef] [Green Version]
- Suttle, J.C. Disruption of the Polar Auxin Transport System in Cotton Seedlings Following Treatment with the Defoliant Thidiazuron. Plant Physiol. 1988, 86, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Shi, Z.; Jiang, Y.; Jiang, L.; Qi, M.; Xu, T.; Li, T. A Family of Auxin Conjugate Hydrolases from Solanum Lycopersicum and Analysis of Their Roles in Flower Pedicel Abscission. BMC Plant Biol. 2019, 19, 233. [Google Scholar] [CrossRef] [PubMed]
- Cin, V.D.; Boschetti, A.; Dorigoni, A.; Ramina, A. Benzylaminopurine Application on Two Different Apple Cultivars (Malus Domestica) Displays New and Unexpected Fruitlet Abscission Features. Ann. Bot. 2007, 99, 1195–1202. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Khare, S.; Trivedi, P.K.; Nath, P. Effect of Ethylene, 1-MCP, ABA and IAA on Break Strength, Cellulase and Polygalacturonase Activities during Cotton Leaf Abscission. S. Afr. J. Bot. 2008, 74, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Shkolnik-Inbar, D.; Bar-Zvi, D. ABI4 Mediates Abscisic Acid and Cytokinin Inhibition of Lateral Root Formation by Reducing Polar Auxin Transport in Arabidopsis. Plant Cell 2010, 22, 3560–3573. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Xing, L.; Wang, X.; Hou, Y.J.; Gao, J.; Wang, P.; Duan, C.G.; Zhu, X.; Zhu, J.K. The ABA Receptor PYL8 Promotes Lateral Root Growth by Enhancing MYB77-Dependent Transcription of Auxin-Responsive Genes. Sci. Signal 2014, 7, ra53. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Meng, J.; Deng, L.; Wang, Y.; Liu, H.; Yao, J.L.; Nieuwenhuizen, N.J.; Wang, Z.; Zeng, W. Diverse Functions of IAA-Leucine Resistant PpILR1 Provide a Genic Basis for Auxin-Ethylene Crosstalk During Peach Fruit Ripening. Front. Plant Sci. 2021, 12, 882. [Google Scholar] [CrossRef]
- Bastajian, N.; Friesen, H.; Andrews, B.J. Bck2 Acts through the MADS Box Protein Mcm1 to Activate Cell-Cycle-Regulated Genes in Budding Yeast. PLoS Genet. 2013, 9, e1003507. [Google Scholar] [CrossRef] [Green Version]
- Lyu, T.; Cao, J. Cys2/His2 Zinc-Finger Proteins in Transcriptional Regulation of Flower Development. Int. J. Mol. Sci. 2018, 19, 2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.; Lashbrook, C.C. Stamen Abscission Zone Transcriptome Profiling Reveals New Candidates for Abscission Control: Enhanced Retention of Floral Organs in Transgenic Plants Overexpressing Arabidopsis ZINC FINGER PROTEIN2. Plant Physiol. 2008, 146, 1305–1321. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.; Liu, C.; Yu, H.; Broun, P. Integration of Cytokinin and Gibberellin Signalling by Arabidopsis Transcription Factors GIS, ZFP8 and GIS2 in the Regulation of Epidermal Cell Fate. Development 2007, 134, 2073–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Sun, L.; Zhao, Y.; An, L.; Yan, A.; Meng, X.; Gan, Y. Zinc Finger Protein 6 (ZFP6) Regulates Trichome Initiation by Integrating Gibberellin and Cytokinin Signaling in Arabidopsis Thaliana. New Phytol. 2013, 198, 699–708. [Google Scholar] [CrossRef]
- Guo, P.; Li, Z.; Huang, P.; Li, B.; Fang, S.; Chu, J.; Guo, H. A Tripartite Amplification Loop Involving the Transcription Factor WRKY75, Salicylic Acid, and Reactive Oxygen Species Accelerates Leaf Senescence. Plant Cell 2017, 29, 2854–2870. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Ma, Q.; Zhang, C.; Wang, C.; Wei, H.; Wang, H.; Yu, S. The Cotton GhWRKY91 Transcription Factor Mediates Leaf Senescence and Responses to Drought Stress in Transgenic Arabidopsis Thaliana. Front. Plant Sci. 2019, 10, 1352. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, Z.; Gao, J.; Zhou, X.; Zhu, S.; Wang, X.; Wang, X.; Ren, G.; Kuai, B. The NPR1-WRKY46-WRKY6 Signaling Cascade Mediates Probenazole/Salicylic Acid-Elicited Leaf Senescence in Arabidopsis Thaliana. J. Integr. Plant Biol. 2021, 63, 924–936. [Google Scholar] [CrossRef]
- Liao, W.; Yang, Y.; Li, Y.; Wang, G.; Peng, M. Genome-Wide Identification of Cassava R2R3 MYB Family Genes Related to Abscission Zone Separation after Environmental-Stress-Induced Abscission. Sci. Rep. 2016, 6, 32006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Wang, W. Hormonal Changes in the Grains of Rice Subjected to Water Stress during Grain Filling. Plant Physiol. 2001, 127, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Li, G.; Yi, G.X.; Wang, B.M.; Deng, A.X.; Nan, T.G.; Li, Z.H.; Li, Q.X. Comparison between Conventional Indirect Competitive Enzyme-Linked Immunosorbent Assay (IcELISA) and Simplified IcELISA for Small Molecules. Anal. Chim. Acta 2006, 571, 79–85. [Google Scholar] [CrossRef]
- Xue, J.; Li, Y.; Tan, H.; Yang, F.; Ma, N.; Gao, J. Expression of Ethylene Biosynthetic and Receptor Genes in Rose Floral Tissues during Ethylene-Enhanced Flower Opening. J. Exp. Bot. 2008, 59, 2161–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Wu, Q.; Liao, B.; Yu, K.; Huo, Y.; Meng, L.; Wang, S.; Wang, B.; Du, M.; Tian, X.; et al. Thidiazuron Promotes Leaf Abscission by Regulating the Crosstalk Complexities between Ethylene, Auxin, and Cytokinin in Cotton. Int. J. Mol. Sci. 2022, 23, 2696. https://doi.org/10.3390/ijms23052696
Li F, Wu Q, Liao B, Yu K, Huo Y, Meng L, Wang S, Wang B, Du M, Tian X, et al. Thidiazuron Promotes Leaf Abscission by Regulating the Crosstalk Complexities between Ethylene, Auxin, and Cytokinin in Cotton. International Journal of Molecular Sciences. 2022; 23(5):2696. https://doi.org/10.3390/ijms23052696
Chicago/Turabian StyleLi, Fangjun, Qian Wu, Baopeng Liao, Keke Yu, Yini Huo, Lu Meng, Songman Wang, Baomin Wang, Mingwei Du, Xiaoli Tian, and et al. 2022. "Thidiazuron Promotes Leaf Abscission by Regulating the Crosstalk Complexities between Ethylene, Auxin, and Cytokinin in Cotton" International Journal of Molecular Sciences 23, no. 5: 2696. https://doi.org/10.3390/ijms23052696
APA StyleLi, F., Wu, Q., Liao, B., Yu, K., Huo, Y., Meng, L., Wang, S., Wang, B., Du, M., Tian, X., & Li, Z. (2022). Thidiazuron Promotes Leaf Abscission by Regulating the Crosstalk Complexities between Ethylene, Auxin, and Cytokinin in Cotton. International Journal of Molecular Sciences, 23(5), 2696. https://doi.org/10.3390/ijms23052696