
Dynamics of Reactive Carbonyl Species in Pea Root Nodules in Response to Polyethylene Glycol (PEG)-Induced Osmotic Stress

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
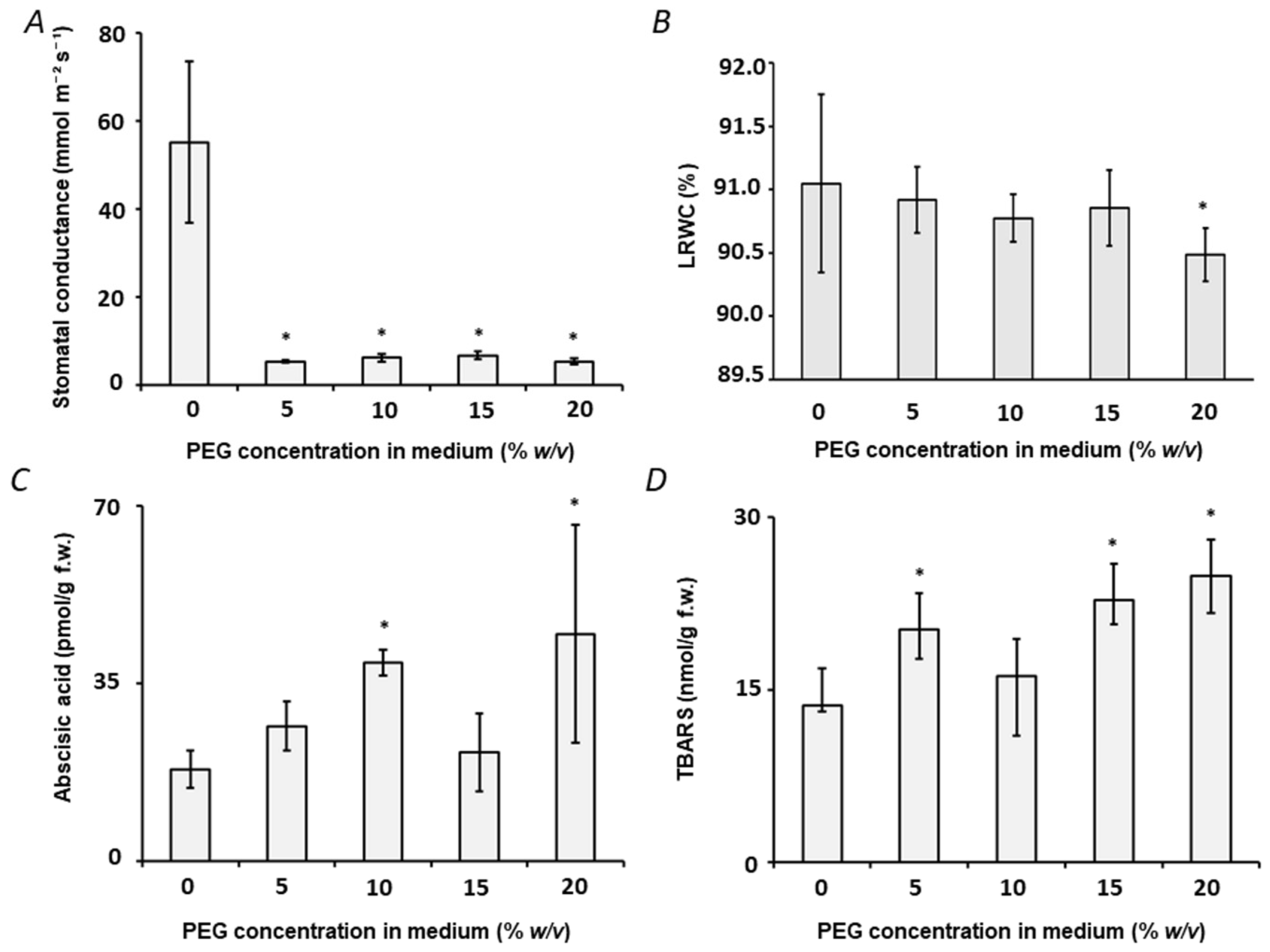
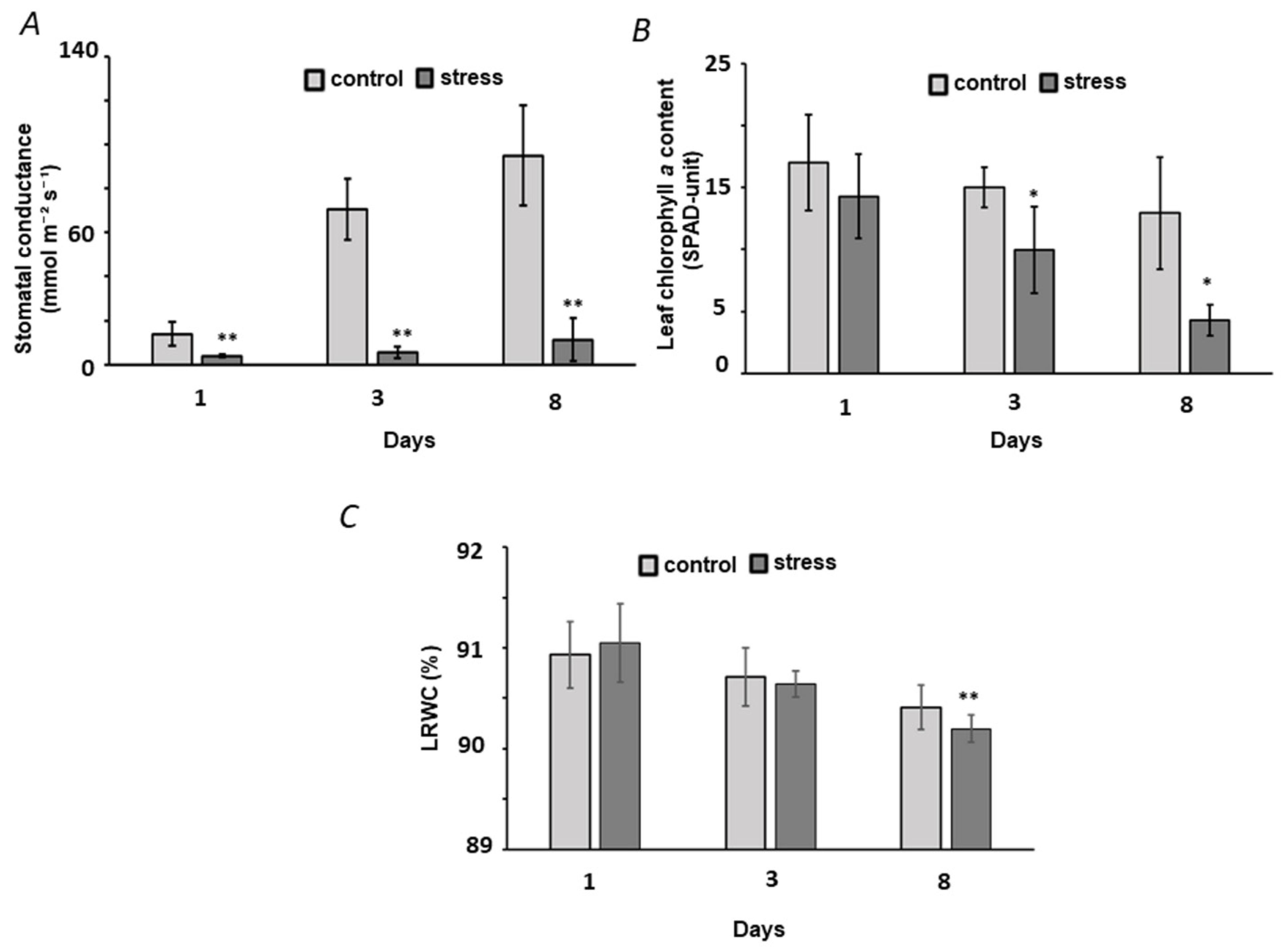
2.1. Establishment of the Drought Stress Model: Plant Growth and Harvesting
2.2. Physiological and Biochemical Characterization of Plant Stress Response
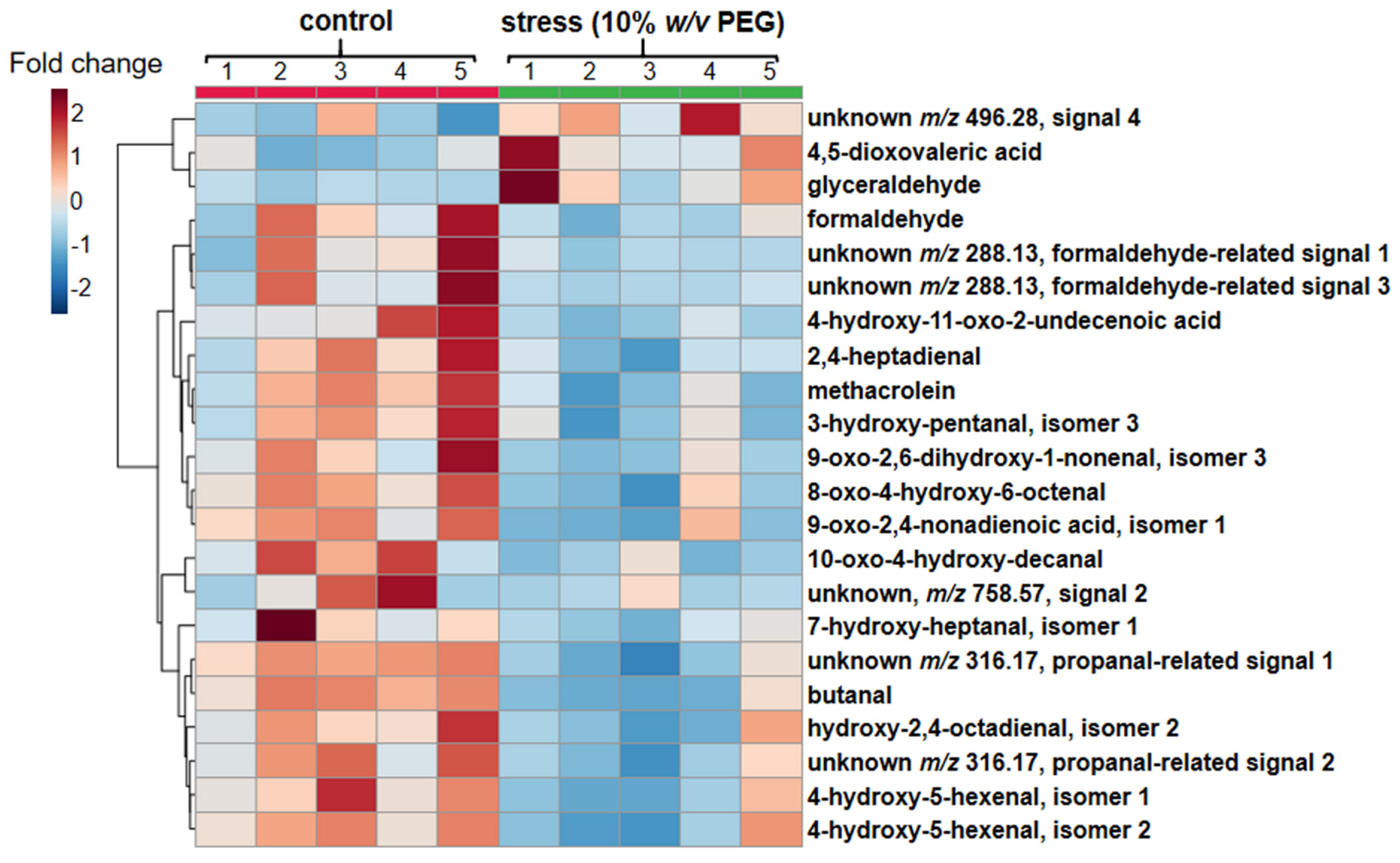
2.3. Quantification of Prospective CHH Derivatives of RCCs by UHPLC-ESI-LIT-Orbitrap-MS
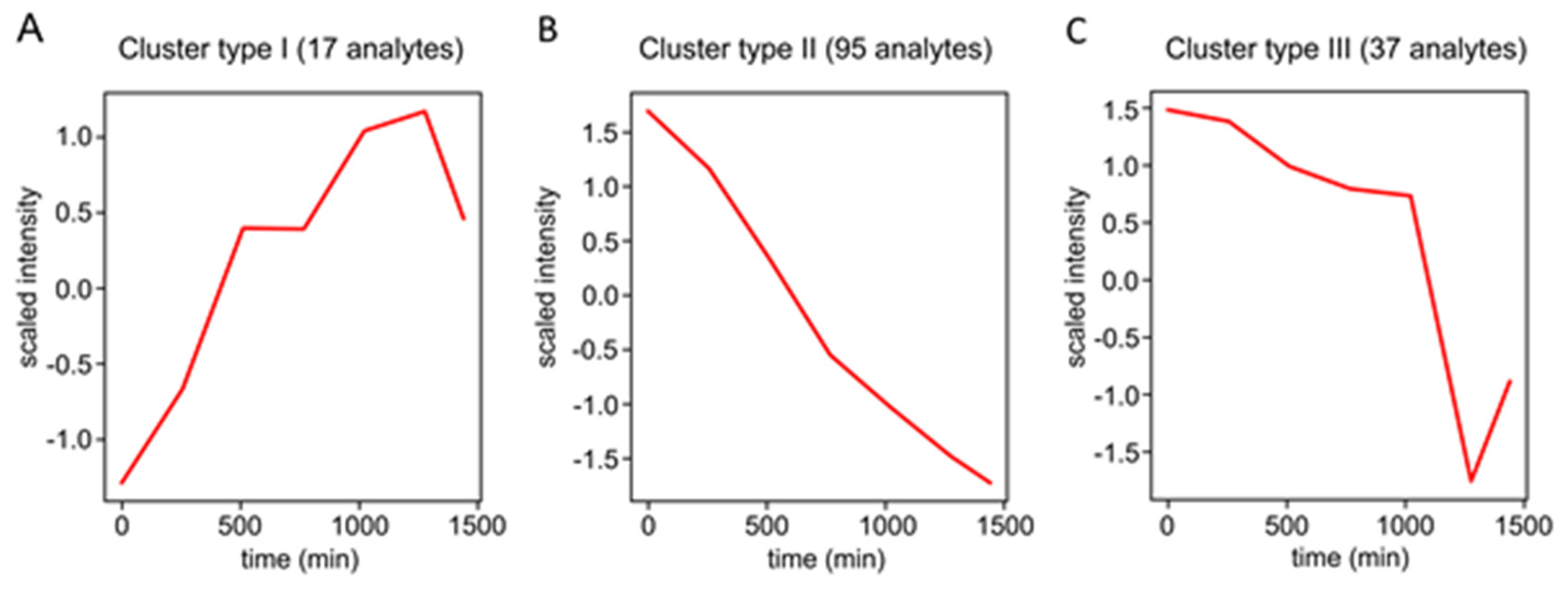
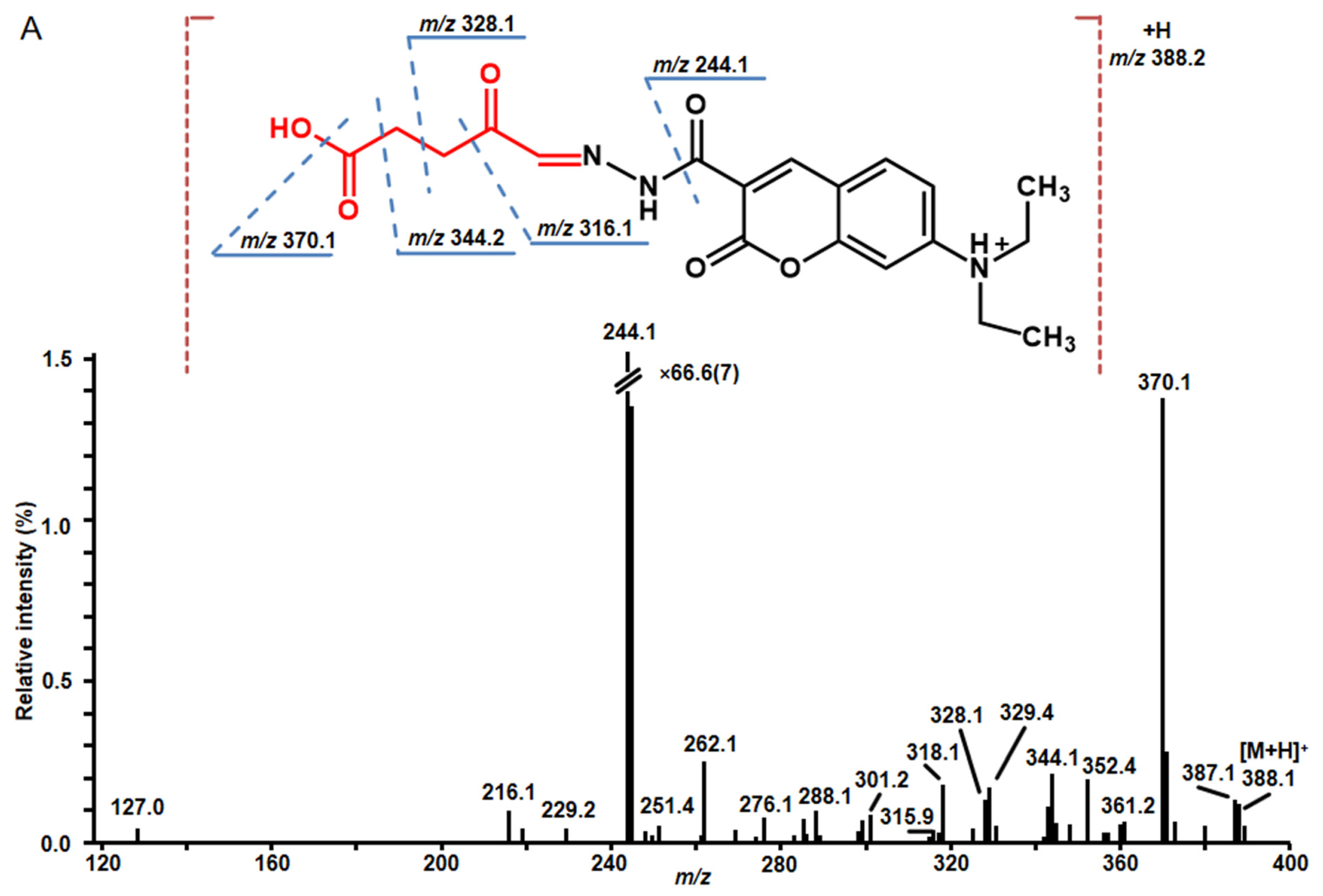
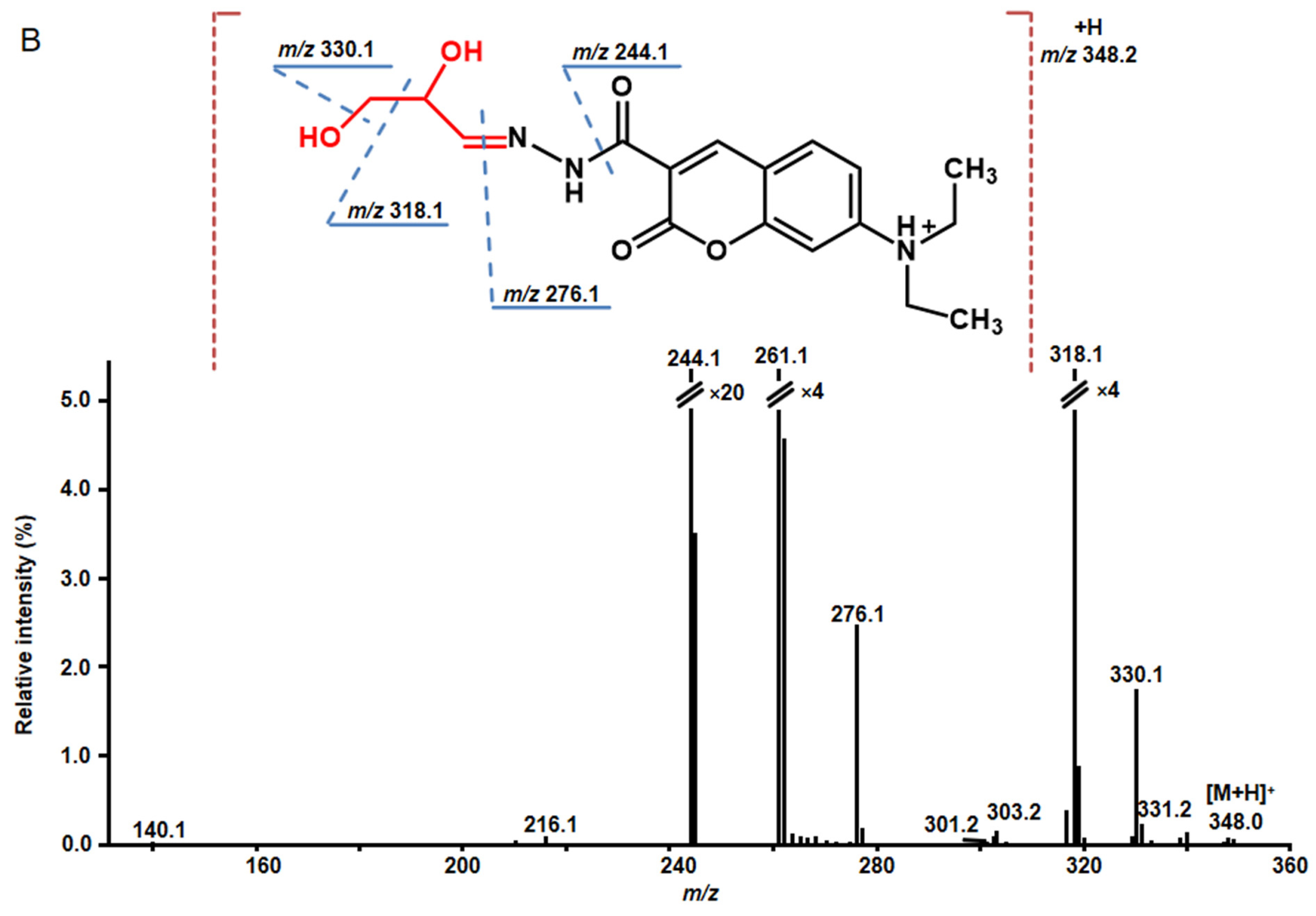
2.4. Structure Characterization of Stress-Regulated Nodule RCCs by MS/MS
3. Discussion
3.1. Establishment of Osmotic Stress in Pea plants
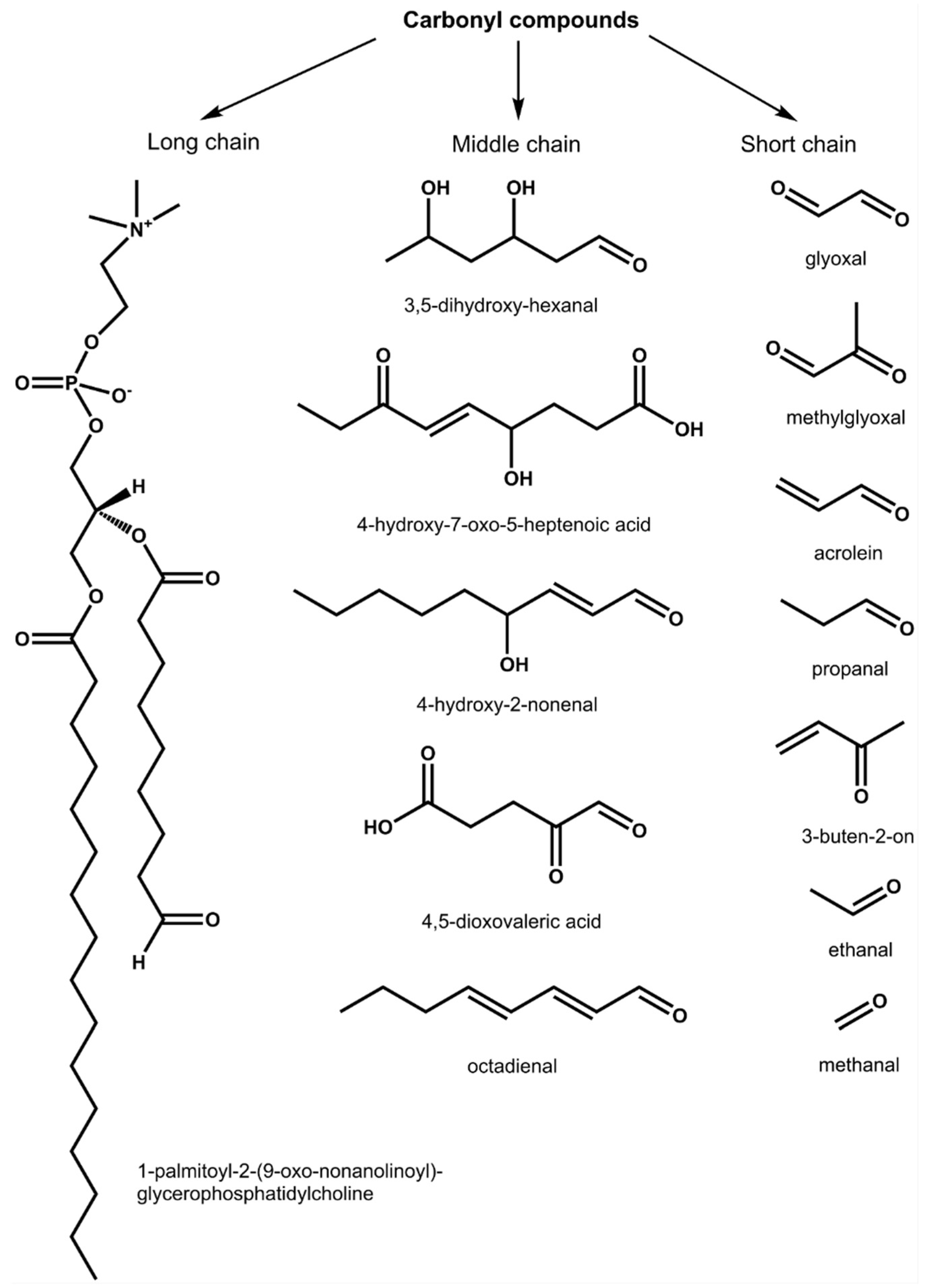
3.2. Analysis of RCCs
3.3. Stress-Related Dynamics of Nodule RCCs
4. Materials and Methods
4.1. Reagents, Plant Material and Rhizobial Culture
4.2. Plant Experiments and the Model of Osmotic Stress
4.3. Physiological and Biochemical Assays
4.4. Extraction and Derivatization of Carbonyl Compounds
4.5. UHPLC-ESI-LIT-Orbitrap-MS
4.6. Annotation and Relative Quantification of Individual RCCs
4.7. Targeted MS/MS Analysis
4.8. Computational Correction of Analyte Degradation/Transformation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| ALA | δ-aminolevulinic acid |
| CHH | 7-(diethylamino)coumarin-3-carbohydrazide |
| DOVA | 4,5-dioxovaleric acid |
| ESI | electrospray ionization |
| FA | formic acid |
| GAP | glyceraldehyde-3-phosphate |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HESI | heated electrospray ionization |
| HPLC | high performance liquid chromatography |
| HSA | human serum albumin |
| JA | jasmonic acid |
| JA-Ile | jasmonyl isoleucine conjugate |
| LC-MS | liquid chromatography-mass spectrometry |
| LC-MS/MS | liquid chromatography-tandem mass-spectrometry |
| LRWC | leaf relative water content |
| MDA | malondialdehyde |
| MS/MS | tandem mass spectrometry |
| NAD+ | nicotinamide adenine dinucleotide |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| OPDA | 12-oxophytodienic acid |
| PEG | polyethylene glycol |
| RCCs | reactive carbonyl compounds |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| RP-UHPLC-MS/MS | reverse phase ultra-high performance liquid chromatography-tandem mass spectrometry |
| RSD | relative standard deviation |
| SPE | solid phase extraction |
| TBA | thiobarbituric acid |
| TBARS | thiobarbituric acid reactive substances |
| UHPLC-ESI-LIT-Orbitrap-MS | ultra-high performance liquid chromatography—electrospray ionization—linear ion trap-Orbitrap mass spectrometry |
| tR | retention time |
| XIC | extracted ion chromatogram |
References
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [Green Version]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passioura, J. The Drought Environment: Physical, Biological and Agricultural Perspectives. J. Exp. Bot. 2007, 58, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.R.; Veneklaas, E.; Polania, J.; Rao, I.M.; Beebe, S.E.; Merchant, A. Field Drought Conditions Impact Yield but Not Nutritional Quality of the Seed in Common Bean (Phaseolus vulgaris L.). PLoS ONE 2019, 14, e0217099. [Google Scholar] [CrossRef] [Green Version]
- García-León, D.; Contreras, S.; Hunink, J. Comparison of Meteorological and Satellite-Based Drought Indices as Yield Predictors of Spanish Cereals. Agric. Water Manag. 2019, 213, 388–396. [Google Scholar] [CrossRef]
- Osmolovskaya, N.; Shumilina, J.; Kim, A.; Didio, A.; Grishina, T.; Bilova, T.; Keltsieva, O.A.; Zhukov, V.; Tikhonovich, I.; Tarakhovskaya, E.; et al. Methodology of Drought Stress Research: Experimental Setup and Physiological Characterization. Int. J. Mol. Sci. 2018, 19, 4089. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Bhardwaj, R.; Sharma, R.; Kapoor, D.; Kohli, S.; Kumar, V.; Kaur, P. Hormonal Regulation of Drought Stress in Plants. In Water Stress and Crop Plants; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 582–599. ISBN 978-1-119-05445-0. [Google Scholar]
- Frolov, A.; Bilova, T.; Paudel, G.; Berger, R.; Balcke, G.U.; Birkemeyer, C.; Wessjohann, L.A. Early Responses of Mature Arabidopsis thaliana Plants to Reduced Water Potential in the Agar-Based Polyethylene Glycol Infusion Drought Model. J. Plant Physiol. 2017, 208, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Shumilina, J.; Kusnetsova, A.; Tsarev, A.; Janse van Rensburg, H.C.; Medvedev, S.; Demidchik, V.; Van den Ende, W.; Frolov, A. Glycation of Plant Proteins: Regulatory Roles and Interplay with Sugar Signalling? Int. J. Mol. Sci. 2019, 20, 2366. [Google Scholar] [CrossRef] [Green Version]
- Soboleva, A.; Schmidt, R.; Vikhnina, M.; Grishina, T.; Frolov, A. Maillard Proteomics: Opening New Pages. Int. J. Mol. Sci. 2017, 18, 2677. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Luqueño, F.; Dendooven, L.; Munive, A.; Corlay-Chee, L.; Serrano-Covarrubias, L.M.; Espinosa-Victoria, D. Micro-Morphology of Common Bean (Phaseolus vulgaris L.) Nodules Undergoing Senescence. Acta Physiol. Plant 2008, 30, 545–552. [Google Scholar] [CrossRef]
- Shumilina, J.; Gorbach, D.; Popova, V.; Tsarev, A.; Kuznetsova, A.; Grashina, M.; Dorn, M.; Lukasheva, E.; Osmolovskaya, N.; Romanovskaya, E.; et al. Protein Glycation and Drought Response of Pea (Pisum sativum L.) Root Nodule Proteome: A Proteomics Approach. Biol. Commun. 2021, 66, 210–224. [Google Scholar] [CrossRef]
- Govindasamy, V.; George, P.; Aher, L.; Ramesh, S.V.; Thangasamy, A.; Anandan, S.; Raina, S.K.; Kumar, M.; Rane, J.; Annapurna, K.; et al. Comparative Conventional and Phenomics Approaches to Assess Symbiotic Effectiveness of Bradyrhizobia Strains in Soybean (Glycine max L. Merrill) to Drought. Sci. Rep. 2017, 7, 6958. [Google Scholar] [CrossRef] [PubMed]
- Serraj, R.; Sinclair, T.R.; Purcell, L.C. Symbiotic N2 Fixation Response to Drought. J. Exp. Bot. 1999, 50, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Milic, I.; Hoffmann, R.; Fedorova, M. Simultaneous Detection of Low and High Molecular Weight Carbonylated Compounds Derived from Lipid Peroxidation by Electrospray Ionization-Tandem Mass Spectrometry. Anal. Chem. 2013, 85, 156–162. [Google Scholar] [CrossRef]
- Altomare, A.; Baron, G.; Gianazza, E.; Banfi, C.; Carini, M.; Aldini, G. Lipid Peroxidation Derived Reactive Carbonyl Species in Free and Conjugated Forms as an Index of Lipid Peroxidation: Limits and Perspectives. Redox Biol. 2021, 42, 101899. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein Carbonylation as a Major Hallmark of Oxidative Damage: Update of Analytical Strategies. Mass Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef]
- Bollineni, R.C.; Fedorova, M.; Blüher, M.; Hoffmann, R. Carbonylated Plasma Proteins As Potential Biomarkers of Obesity Induced Type 2 Diabetes Mellitus. J. Proteome Res. 2014, 13, 5081–5093. [Google Scholar] [CrossRef]
- Mano, J.; Biswas, M.S.; Sugimoto, K. Reactive Carbonyl Species: A Missing Link in ROS Signaling. Plants 2019, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Semchyshyn, H.M. Reactive Carbonyl Species In Vivo: Generation and Dual Biological Effects. Sci. World J. 2014, 2014, 417842. [Google Scholar] [CrossRef] [Green Version]
- Biswas, M.S.; Mano, J. Lipid Peroxide-Derived Reactive Carbonyl Species as Mediators of Oxidative Stress and Signaling. Front. Plant Sci. 2021, 12, 720867. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile C6-Aldehydes and Allo-Ocimene Activate Defense Genes and Induce Resistance against Botrytis Cinerea in Arabidopsis thaliana. Plant Cell Physiol. 2005, 46, 1093–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alméras, E.; Stolz, S.; Vollenweider, S.; Reymond, P.; Mène-Saffrané, L.; Farmer, E.E. Reactive Electrophile Species Activate Defense Gene Expression in Arabidopsis. Plant J. 2003, 34, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Chételat, A.; Reymond, P.; Farmer, E.E. Selective and Powerful Stress Gene Expression in Arabidopsis in Response to Malondialdehyde. Plant J. 2004, 37, 877–888. [Google Scholar] [CrossRef]
- Taki, N.; Sasaki-Sekimoto, Y.; Obayashi, T.; Kikuta, A.; Kobayashi, K.; Ainai, T.; Yagi, K.; Sakurai, N.; Suzuki, H.; Masuda, T.; et al. 12-Oxo-Phytodienoic Acid Triggers Expression of a Distinct Set of Genes and Plays a Role in Wound-Induced Gene Expression in Arabidopsis. Plant Physiol. 2005, 139, 1268–1283. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, Y.; Furutera, A.; Seki, K.; Toyoda, Y.; Tanaka, K.; Sugimoto, Y. Malondialdehyde Generated from Peroxidized Linolenic Acid Causes Protein Modification in Heat-Stressed Plants. Plant Physiol. Biochem. 2008, 46, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, M.A.; Kim, A.; Peñuelas, M.; Ihling, C.; Griesser, E.; Hoffmann, R.; Fedorova, M.; Frolov, A.; Becana, M. Protein Carbonylation and Glycation in Legume Nodules. Plant Physiol. 2018, 177, 1510–1528. [Google Scholar] [CrossRef] [Green Version]
- Fritzsche, S.; Billig, S.; Rynek, R.; Abburi, R.; Tarakhovskaya, E.; Leuner, O.; Frolov, A.; Birkemeyer, C. Derivatization of Methylglyoxal for LC-ESI-MS Analysis—Stability and Relative Sensitivity of Different Derivatives. Molecules 2018, 23, 2994. [Google Scholar] [CrossRef] [Green Version]
- Henning, C.; Glomb, M.A. Pathways of the Maillard Reaction under Physiological Conditions. Glycoconj. J. 2016, 33, 499–512. [Google Scholar] [CrossRef]
- Krajewski, L.C.; Lobodin, V.V.; Robbins, W.K.; Jin, P.; Bota, G.; Marshall, A.G.; Rodgers, R.P. Method for Isolation and Detection of Ketones Formed from High-Temperature Naphthenic Acid Corrosion. Energy Fuels 2017, 31, 10674–10679. [Google Scholar] [CrossRef]
- Milic, I.; Fedorova, M. Derivatization and Detection of Small Aliphatic and Lipid-Bound Carbonylated Lipid Peroxidation Products by ESI-MS. Methods Mol. Biol. 2015, 1208, 3–20. [Google Scholar] [CrossRef]
- Paudel, G.; Bilova, T.; Schmidt, R.; Greifenhagen, U.; Berger, R.; Tarakhovskaya, E.; Stöckhardt, S.; Balcke, G.U.; Humbeck, K.; Brandt, W.; et al. Osmotic Stress Is Accompanied by Protein Glycation in Arabidopsis thaliana L. J. Exp. Bot. 2016, 67, 6283–6295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.S.; Abraham, T.K.; Jaya, D.S. Studies on the Changes in Lipid Peroxidation and Antioxidants in Drought Stress Induced Cowpea (Vigna unguiculata L.) Varieties. J. Environ. Biol. 2008, 29, 689–691. [Google Scholar]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The Physiology of Plant Responses to Drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Egert, M.; Tevini, M. Influence of Drought on Some Physiological Parameters Symptomatic for Oxidative Stress in Leaves of Chives (Allium schoenoprasum). Environ. Exp. Bot. 2002, 48, 43–49. [Google Scholar] [CrossRef]
- Sergiev, I.; Todorova, D.; Shopova, E.A.; Jankauskienė, J.; Jankovska-Bortkevič, E.; Jurkonienė, S. Exogenous Auxin Type Compounds Amend PEG-Induced Physiological Responses Of Pea Plants 10.1016/J.SCIENTA.2019.01.015. Chemistry 2019, 248, 200–205. [Google Scholar]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The Effect of Drought and Ultraviolet Radiation on Growth and Stress Markers in Pea and Wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Basal, O.; Szabó, A.; Veres, S. Physiology of Soybean as Affected by PEG-Induced Drought Stress. Curr. Plant Biol. 2020, 22, 100135. [Google Scholar] [CrossRef]
- Leonova, T.; Popova, V.; Tsarev, A.; Henning, C.; Antonova, K.; Rogovskaya, N.; Vikhnina, M.; Baldensperger, T.; Soboleva, A.; Dinastia, E.; et al. Does Protein Glycation Impact on the Drought-Related Changes in Metabolism and Nutritional Properties of Mature Pea (Pisum sativum L.) Seeds? Int. J. Mol. Sci. 2020, 21, 567. [Google Scholar] [CrossRef] [Green Version]
- González, E.M.; Wienkoop, S.; Staudinger, C.; Lyon, D.; Gil-Quintana, E. Medicago truncatula: Local Response of the Root Nodule Proteome to Drought Stress. In The Model Legume Medicago truncatula; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 1096–1101. ISBN 978-1-119-40914-4. [Google Scholar]
- Hossain, M.A.; Liu, F.; Burritt, D.J.; Fujita, M.; Huang, B. Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Academic Press: Cambridge, MA, USA, 2020; ISBN 978-0-12-817893-5. [Google Scholar]
- Zwiewka, M.; Bielach, A.; Tamizhselvan, P.; Madhavan, S.; Ryad, E.E.; Tan, S.; Hrtyan, M.N.; Dobrev, P.; Vankovï, R.; Friml, J.; et al. Root Adaptation to H2O2-Induced Oxidative Stress by ARF-GEF BEN1- and Cytoskeleton-Mediated PIN2 Trafficking. Plant Cell Physiol. 2019, 60, 255–273. [Google Scholar] [CrossRef]
- Liu, X.; Jeffries, H.E.; Sexton, K.G. Hydroxyl Radical and Ozone Initiated Photochemical Reactions of 1,3-Butadiene. Atmos. Environ. 1999, 33, 3005–3022. [Google Scholar] [CrossRef]
- Yang, F.; Miao, L.-F. Adaptive Responses to Progressive Drought Stress in Two Poplar Species Originating from Different Altitudes. Silva Fenn. 2010, 44, 23–37. [Google Scholar] [CrossRef] [Green Version]
- Mitran, M.I.; Nicolae, I.; Tampa, M.; Mitran, C.I.; Caruntu, C.; Sarbu, M.I.; Ene, C.D.; Matei, C.; Georgescu, S.R.; Popa, M.I. Reactive Carbonyl Species as Potential Pro-Oxidant Factors Involved in Lichen Planus Pathogenesis. Metabolites 2019, 9, 213. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, L.; Dong, F.; Gao, J.; Galbraith, D.W.; Song, C.-P. Hydrogen Peroxide Is Involved in Abscisic Acid-Induced Stomatal Closure in Vicia faba. Plant Physiol. 2001, 126, 1438–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, J.; Turner, N.C.; Khan, T.; Du, Y.-L.; Xiong, J.-L.; Colmer, T.D.; Devilla, R.; Stefanova, K.; Siddique, K.H.M. Response of Chickpea (Cicer arietinum L.) to Terminal Drought: Leaf Stomatal Conductance, Pod Abscisic Acid Concentration, and Seed Set. J. Exp. Bot. 2017, 68, 1973–1985. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D.; Rothmann, S.; Schneider, J.Y.; Suchy, M.-T.; Trettin, A.; Modun, D.; Stuke, N.; Maassen, N.; Frölich, J.C. Development, Validation and Biomedical Applications of Stable-Isotope Dilution GC–MS and GC–MS/MS Techniques for Circulating Malondialdehyde (MDA) after Pentafluorobenzyl Bromide Derivatization: MDA as a Biomarker of Oxidative Stress and Its Relation to 15(S)-8-Iso-Prostaglandin F2α and Nitric Oxide (NO). J. Chromatogr. B 2016, 1019, 95–111. [Google Scholar] [CrossRef]
- Tang, Y.; Zhao, Y.; Wang, P.; Sang, S. Simultaneous Determination of Multiple Reactive Carbonyl Species in High Fat Diet-Induced Metabolic Disordered Mice and the Inhibitory Effects of Rosemary on Carbonyl Stress. J. Agric. Food Chem. 2021. [Google Scholar] [CrossRef] [PubMed]
- Griesser, E.; Vemula, V.; Raulien, N.; Wagner, U.; Reeg, S.; Grune, T.; Fedorova, M. Cross-Talk between Lipid and Protein Carbonylation in a Dynamic Cardiomyocyte Model of Mild Nitroxidative Stress. Redox Biol. 2017, 11, 438–455. [Google Scholar] [CrossRef]
- Mano, J.; Tokushige, K.; Mizoguchi, H.; Fujii, H.; Khorobrykh, S. Accumulation of Lipid Peroxide-Derived, Toxic α,β-Unsaturated Aldehydes (E)-2-Pentenal, Acrolein and (E)-2-Hexenal in Leaves under Photoinhibitory Illumination. Plant Biotechnol. 2010, 27, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Kamleh, M.A.; Ebbels, T.M.D.; Spagou, K.; Masson, P.; Want, E.J. Optimizing the Use of Quality Control Samples for Signal Drift Correction in Large-Scale Urine Metabolic Profiling Studies. Anal Chem. 2012, 84, 2670–2677. [Google Scholar] [CrossRef]
- Sánchez-Illana, Á.; Pérez-Guaita, D.; Cuesta-García, D.; Sanjuan-Herráez, J.D.; Vento, M.; Ruiz-Cerdá, J.L.; Quintás, G.; Kuligowski, J. Model Selection for Within-Batch Effect Correction in UPLC-MS Metabolomics Using Quality Control-Support Vector Regression. Anal. Chim. Acta 2018, 1026, 62–68. [Google Scholar] [CrossRef]
- Islam, M.M.; Ye, W.; Matsushima, D.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, S.; Mano, J.; Murata, Y. Reactive Carbonyl Species Mediate ABA Signaling in Guard Cells. Plant Cell Physiol. 2016, 57, 2552–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, J.; Figueiredo, M.; Cavalcanti, F.R.; Ferreira-Silva, S. Legume Nodule Oxidative Stress and N2 Fixation Efficiency. Microb. Ecol. Trop. Soils 2011, 1, 9–78. [Google Scholar]
- Gogorcena, Y.; Iturbe-Ormaetxe, I.; Escuredo, P.R.; Becana, M. Antioxidant Defenses against Activated Oxygen in Pea Nodules Subjected to Water Stress. Plant Physiol. 1995, 108, 753–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldridge, K.C.; Zavala, J.; Surratt, J.; Sexton, K.G.; Contreras, L.M. Cellular RNA Is Chemically Modified by Exposure to Air Pollution Mixtures. Inhal. Toxicol. 2015, 27, 74–82. [Google Scholar] [CrossRef]
- Mano, J.; Kanameda, S.; Kuramitsu, R.; Matsuura, N.; Yamauchi, Y. Detoxification of Reactive Carbonyl Species by Glutathione Transferase Tau Isozymes. Front. Plant Sci. 2019, 10, 487. [Google Scholar] [CrossRef] [Green Version]
- Suganuma, N.; Yamamoto, Y. Carbon Metabolism Related to Nitrogen Fixation in Soybean Root Nodules. Soil Sci. Plant Nutr. 1987, 33, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, D.; Kannan, M.; Reddy, A.R. A Root Proteomics-Based Insight Reveals Dynamic Regulation of Root Proteins under Progressive Drought Stress and Recovery in Vigna radiata (L.) Wilczek. Planta 2011, 233, 1111–1127. [Google Scholar] [CrossRef]
- Yu, J.; Sun, H.; Zhang, J.; Hou, Y.; Zhang, T.; Kang, J.; Wang, Z.; Yang, Q.; Long, R. Analysis of Aldo–Keto Reductase Gene Family and Their Responses to Salt, Drought, and Abscisic Acid Stresses in Medicago truncatula. Int. J. Mol. Sci. 2020, 21, 754. [Google Scholar] [CrossRef] [Green Version]
- Dalton, D.A.; Boniface, C.; Turner, Z.; Lindahl, A.; Kim, H.J.; Jelinek, L.; Govindarajulu, M.; Finger, R.E.; Taylor, C.G. Physiological Roles of Glutathione S-Transferases in Soybean Root Nodules. Plant Physiol. 2009, 150, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I. The Roles of Reactive Carbonyl Species in Induction of Antioxidant Defence and ROS Signalling in Extreme Halophytic Model Eutrema parvulum and Glycophytic Model Arabidopsis thaliana. Environ. Exp. Bot. 2019, 160, 81–91. [Google Scholar] [CrossRef]
- Cao, X.; Zhu, C.; Zhong, C.; Zhang, J.; Wu, L.; Jin, Q.; Ma, Q. Nitric Oxide Synthase-Mediated Early Nitric Oxide Burst Alleviates Water Stress-Induced Oxidative Damage in Ammonium-Supplied Rice Roots. BMC Plant Biol. 2019, 19, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamoros, M.A.; Becana, M. Molecular Responses of Legumes to Abiotic Stress: Post-Translational Modifications of Proteins and Redox Signaling. J. Exp. Bot. 2021, 72, 5876–5892. [Google Scholar] [CrossRef]
- Fahim, J.R.; Attia, E.Z.; Kamel, M.S. The Phenolic Profile of Pea (Pisum sativum): A Phytochemical and Pharmacological Overview. Phytochem. Rev. 2019, 18, 173–198. [Google Scholar] [CrossRef]
- Hidalgo, F.J.; Delgado, R.M.; Zamora, R. Protective Effect of Phenolic Compounds on Carbonyl-Amine Reactions Produced by Lipid-Derived Reactive Carbonyls. Food Chem. 2017, 229, 388–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colzani, M.; Regazzoni, L.; Criscuolo, A.; Baron, G.; Carini, M.; Vistoli, G.; Lee, Y.-M.; Han, S.-I.; Aldini, G.; Yeum, K.-J. Isotopic Labelling for the Characterisation of HNE-Sequestering Agents in Plant-Based Extracts and Its Application for the Identification of Anthocyanidins in Black Rice with Giant Embryo. Free Radic. Res. 2018, 52, 896–906. [Google Scholar] [CrossRef]
- Ho, C.-T.; Wang, M. Dietary Phenolics as Reactive Carbonyl Scavengers: Potential Impact on Human Health and Mechanism of Action. J. Tradit. Complement. Med. 2013, 3, 139–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soboleva, A.; Vikhnina, M.; Grishina, T.; Frolov, A. Probing Protein Glycation by Chromatography and Mass Spectrometry: Analysis of Glycation Adducts. Int. J. Mol. Sci. 2017, 18, 2557. [Google Scholar] [CrossRef] [Green Version]
- Flores-Tinoco, C.E.; Tschan, F.; Fuhrer, T.; Margot, C.; Sauer, U.; Christen, M.; Christen, B. Co-Catabolism of Arginine and Succinate Drives Symbiotic Nitrogen Fixation. Mol. Syst. Biol. 2020, 16, 9419. [Google Scholar] [CrossRef]
- Ramos, M.L.G.; Parsons, R.; Sprent, J.I. Differences in Ureide and Amino Acid Composition of Water Stressed Soybean Inoculated with Either Bradyrhizobium japonicum (CB1809) or B. Elkanii (29W). In Biological Nitrogen Fixation for the 21st Century: Proceedings of the 11th International Congress on Nitrogen Fixation, Institut Pasteur, Paris, France, 20–25 July 1997; Elmerich, C., Kondorosi, A., Newton, W.E., Eds.; Current Plant Science and Biotechnology in Agriculture; Springer Netherlands: Dordrecht, The Netherlands, 1998; p. 521. ISBN 978-94-011-5159-7. [Google Scholar]
- López-Gómez, M.; Hidalgo-Castellanos, J.; Iribarne, C.; Lluch, C. Proline Accumulation Has Prevalence over Polyamines in Nodules of Medicago sativa in Symbiosis with Sinorhizobium meliloti during the Initial Response to Salinity. Plant Soil 2014, 374, 149–159. [Google Scholar] [CrossRef]
- Bertrand, A.; Bipfubusa, M.; Dhont, C.; Chalifour, F.-P.; Drouin, P.; Beauchamp, C.J. Rhizobial Strains Exert a Major Effect on the Amino Acid Composition of Alfalfa Nodules under NaCl Stress. Plant Physiol. Biochem. 2016, 108, 344–352. [Google Scholar] [CrossRef]
- Ummus, R.E.; Onuki, J.; Dörnemann, D.; Medeiros, M.H.; Di Mascio, P. Measurement of 4,5-Dioxovaleric Acid by High-Performance Liquid Chromatography and Fluorescence Detection. J. Chromatogr. B Biomed. Sci. Appl. 1999, 729, 237–243. [Google Scholar] [CrossRef]
- Bechara, E.J.H.; Ramos, L.D.; Stevani, C.V. 5-Aminolevulinic Acid: A Matter of Life and Caveats. J. Photochem. Photobiol. 2021, 7, 100036. [Google Scholar] [CrossRef]
- Di Mascio, P.; Teixeira, P.C.; Onuki, J.; Medeiros, M.H.G.; Dörnemann, D.; Douki, T.; Cadet, J. DNA Damage by 5-Aminolevulinic and 4,5-Dioxovaleric Acids in the Presence of Ferritin. Arch. Biochem. Biophys. 2000, 373, 368–374. [Google Scholar] [CrossRef]
- Brumm, P.J.; Thomas, G.A.; Friedmann, H.C. The Role of 4,5-Dioxovaleric Acid in Porphyrin and Vitamin B12 Formation by Clostridia. Biochem. Biophys. Res. Commun. 1982, 104, 814–822. [Google Scholar] [CrossRef]
- Ramos, M.L.G.; Gordon, A.J.; Minchin, F.R.; Sprent, J.I.; Parsons, R. Effect of Water Stress on Nodule Physiology and Biochemistry of a Drought Tolerant Cultivar of Common Bean (Phaseolus vulgaris L.). Ann. Bot. 1999, 83, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Nadler, K.D.; Avissar, Y.J. Heme Synthesis in Soybean Root Nodules: I. On the Role of Bacteroid δ-Aminolevulinic Acid Synthase and δ-Aminolevulinic Acid Dehydrase in the Synthesis of the Heme of Leghemoglobin 1. Plant Physiol. 1977, 60, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Sangwan, I.; O’Brian, M.R. Characterization of δ-Aminolevulinic Acid Formation in Soybean Root Nodules 1. Plant Physiol. 1992, 98, 1074–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Jiménez, F.; Collera Zúñiga, O.; Castells García, Y.; Cárdenas, J.; Cuevas, G. Experimental and Theoretical Study of the Products from the Spontaneous Dimerization of DL- and D-Glyceraldehyde. J. Braz. Chem. Soc. 2005, 16, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Muronetz, V.I.; Melnikova, A.K.; Barinova, K.V.; Schmalhausen, E.V. Inhibitors of Glyceraldehyde 3-Phosphate Dehydrogenase and Unexpected Effects of Its Reduced Activity. Biochem. Mosc. 2019, 84, 1268–1279. [Google Scholar] [CrossRef]
- Muronetz, V.I.; Melnikova, A.K.; Saso, L.; Schmalhausen, E. Influence of Oxidative Stress on Catalytic and Non-Glycolytic Functions of Glyceraldehyde-3-Phosphate Dehydrogenase. Curr. Med. Chem. 2020, 27, 2040–2058. [Google Scholar] [CrossRef]
- Peralta, D.; Bronowska, A.K.; Morgan, B.; Dóka, É.; Van Laer, K.; Nagy, P.; Gräter, F.; Dick, T.P. A Proton Relay Enhances H2O2 Sensitivity of GAPDH to Facilitate Metabolic Adaptation. Nat. Chem. Biol. 2015, 11, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Al-Motawa, M.; Thornalley, P.J. Protein Glycation in Plants—An Under-Researched Field with Much Still to Discover. Int. J. Mol. Sci. 2020, 21, 3942. [Google Scholar] [CrossRef] [PubMed]
- Jahn, M.; Spiteller, G. Oxidation of D-(-)-Ribose with H2O2 and Lipid Hydroperoxides. Z. Nat. C 1996, 51, 870–876. [Google Scholar] [CrossRef]
- Guo, L.; Devaiah, S.P.; Narasimhan, R.; Pan, X.; Zhang, Y.; Zhang, W.; Wang, X. Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenases Interact with Phospholipase DΔ to Transduce Hydrogen Peroxide Signals in the Arabidopsis Response to Stress. Plant Cell 2012, 24, 2200–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.d.M.; Ye, W.; Akter, F.; Rhaman, M.S.; Matsushima, D.; Munemasa, S.; Okuma, E.; Nakamura, Y.; Biswas, M.d.S.; Mano, J.; et al. Reactive Carbonyl Species Mediate Methyl Jasmonate-Induced Stomatal Closure. Plant Cell Physiol. 2020, 61, 1788–1797. [Google Scholar] [CrossRef]
- Sulima, A.S.; Zhukov, V.A.; Kulaeva, O.A.; Vasileva, E.N.; Borisov, A.Y.; Tikhonovich, I.A. New Sources of Sym2A Allele in the Pea (Pisum sativum L.) Carry the Unique Variant of Candidate LysM-RLK Gene LykX. PeerJ 2019, 7, e8070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida Silva, M.; Jifon, J.L.; Sharma, V.; da Silva, J.A.G.; Caputo, M.M.; Damaj, M.B.; Guimarães, E.R.; Ferro, M.I.T. Use of Physiological Parameters in Screening Drought Tolerance in Sugarcane Genotypes. Sugar Tech 2011, 13, 191–197. [Google Scholar] [CrossRef]
- Morison, J.I.L. Increasing Atmospheric CO2 and Stomata. New Phytol. 2001, 149, 154–156. [Google Scholar] [CrossRef]
- Chantseva, V.; Bilova, T.; Smolikova, G.; Frolov, A.; Medvedev, S. 3D-Clinorotation Induces Specific Alterations in Metabolite Profiles of Germinating Brassica napus L. Seeds. Biol. Commun. 2019, 64, 55–74. [Google Scholar] [CrossRef]
- Balcke, G.U.; Handrick, V.; Bergau, N.; Fichtner, M.; Henning, A.; Stellmach, H.; Tissier, A.; Hause, B.; Frolov, A. An UPLC-MS / MS Method for Highly Sensitive High-Throughput Analysis of Phytohormones in Plant Tissues. Plant Methods 2012, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- Petitjean, F.; Ketterlin, A.; Gançarski, P. A Global Averaging Method for Dynamic Time Warping, with Applications to Clustering. Pattern Recognit. 2011, 44, 678–693. [Google Scholar] [CrossRef]
- Tavenard, R.; Faouzi, J.; Vandewiele, G.; Divo, F.; Androz, G.; Holtz, C.; Payne, M.; Yurchak, R.; Rußwurm, M.; Kolar, K.; et al. Tslearn, a machine learning toolkit for time series data. J. Mach. Learn. Res. 2020, 21, 1–6. [Google Scholar]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array Programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-Learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Waskom, M.L. Seaborn: Statistical Data Visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | tR (Min) | m/z [M + H]+ Observed | m/z [M + H]+ Calculated | Elemental Composition | Fragmentation Patterns (m/z (%)) | Error (ppm) | Fold Change | Assignment |
|---|---|---|---|---|---|---|---|---|
| 1 * | 3.7 | 348.1555 | 348.1554 | C17H22N3O5 | 216.1 (6), 261.1 (100), 262.1 (31), 276.1 (19), 301.2 (3), 303.2 (9.4), 318.1 (94), 319.2 (6), 330.1 (13), 331.2 (3) | −0.3 | 2.7↑ | glyceraldehyde |
| 2 * | 6.2 | 388.1507 | 388.1503 | C19H22N3O6 | 216.0 (7.1), 262.2 (18.6), 276.1 (5.7), 288.1 (7.1), 301.0 (6.4), 315.9 (2.9), 318.1 (13.6), 328.1 (9.3), 343.2 (7.9), 344.1 (15.7), 352.4 (15.0), 360.0 (4.6), 370.1 (100), 371.1 (20.0) | −1.0 | 2.1↑ | 4,5-dioxovaleric acid |
| 3 * | 6.6 | 288.1344 | 288.1344 | C15H18N3O3 | 216.2 (7), 217.1 (18), 218.2 (40), 225.2 (12), 232.1 (100), 258.3 (9), 260.2 (40), 261.3 (65.7), 262.2 (89), 271.3 (6), 276.0 (12) | 0.0 | 1.5↓ | formaldehyde |
| 4 * | 7.9 | 316.1655 | 316.1656 | C17H22N3O3 | 216.1 (18), 218.2 (58), 219.1 (24), 258.2 (100), 260.1 (42), 272.4 (47), 298.2 (67), 299.4 (76) | 0.3 | 1.7↓ | unknown m/z 316.17, propanal-related signal 1 |
| 5 | 8.2 | 372.1914 | 372.1917 | C20H26N3O4 | 218.1 (0.05), 244.1 (30), 261.1 (0.5), 276.1 (16), 302.2 (0.6), 315.2 (0.25), 328.3 (0.1), 342.2 (0.3), 353.0 (2.5), 354.2 (100) | 0.8 | 1.6↓ | 4-hydroxy-5-hexenal, isomer 1 |
| 6 * | 8.3 | 316.1655 | 316.1656 | C17H22N3O3 | 217.1 (9), 218.2 (3), 232.2 (14), 256.3 (16), 258.2 (100), 260.2 (25), 262.2 (21), 298.1 (20), 299.3 (38), 300.2 (29) | 0.3 | 2.0↓ | unknown m/z 316.17 propanal-related signal 2 |
| 7 | 8.3 | 398.2074 | 398.2073 | C22H28N3O4 | 123.0 (3.3), 138.0 (0.5), 244.1 (16), 260.2 (0.7), 276.2 (1.4), 298.1 (0.22), 311.1 (0.35), 316.2 (0.3), 328.2 (0.37), 337.2 (2.2), 340.2 (0.4), 356.2 (1.1), 363.2 (1.8), 366.2 (1.2), 380.2 (100), 381.2 (15), 397.3 (0.6) | −0.3 | 1.6↓ | hydroxy-2,4-octadienal |
| 8 | 8.4 | 372.1917 | 372.1917 | C20H26N3O4 | 216.2 (0.18), 244.1 (55), 261.3 (0.3), 262.3 (0.55), 276.1 (100), 300.3 (0.15), 313.9 (0.1), 329.7 (0.17), 342.2 (1.8), 354.2 (2.4), 355.2 (0.35) | 0.0 | 1.6↓ | 4-hydroxy-5-hexenal, isomer 2 |
| 9 * | 8.5 | 328.1653 | 328.1656 | C18H22N3O3 | 216.1 (32), 232.1 (8), 256.1 (2), 260.1 (100), 271.1 (2), 283.2 (10), 300.1 (18), 311.2 (6) | 0.9 | 2.1↓ | methacrolein |
| 10 | 8.5 | 360.1915 | 360.1915 | C19H26N3O4 | 216.1 (0.04), 232.3 (0.02), 244.1 (1.05), 261.2 (0.06), 276.1 (0.08), 297.1 (1.17), 302.2 (0.08), 314.1 (1.24), 315.2 (0.36), 327.0 (1.13), 328.2 (100) | 0.6 | 1.8↓ | 3-hydroxy-pentanal |
| 11 * | 8.5 | 414.2021 | 414.2022 | C22H28N3O5 | 139.1 (1), 276.1 (22), 288.2 (7), 302.2 (1) 316.1 (100), 317.1 (10), 340.2 (1), 354.2 (11), 380.2 (4), 396.2 (62) | 0.2 | 2.2↓ | 8-oxo-4-hydroxy-6-octenal |
| 12 * | 8.5 | 426.2021 | 426.2022 | C23H28N3O5 | 122.1 (0.5), 166.0 (0.5), 218.1 (0.5), 262.2 (3.3), 276.1 (11), 298.1 (0.5), 330.2 (1.4), 352.2 (1.4), 354.2 (7.6), 366.2 (2.4), 380.2 (100), 381.2 (22), 382.2 (45), 383.3 (11), 394.2 (1.4), 408.2 (40) | 0.2 | 2.7↓ | 9-oxo-2,4-nonadienoic acid |
| 13 | 8.8 | 444.2120 | 444.2127 | C23H30N3O6 | 183.1 (0.15), 244.1 (44), 262.2 (0.52), 276.1 (100), 300.2 (0.3), 302.3 (0.4), 316.2 (5), 332.3 (0.5), 346.2 (0.15), 356.1 (0.3), 368.1 (0.4), 370.2 (0.6), 384.2 (0.4), 408.2 (1.9), 426.3 (4.9) | 1.6 | 2.1↓ | 9-oxo-2,6-dihydroxy-1-nonenal |
| 14 | 8.8 | 472.2450 | 472.2439 | C25H34N3O6 | 139.0 (11), 154.1 (0.2), 175.1 (0.3), 193.1 (1.6), 212.1 (1.8), 244.1 (18), 261.0 (0.8), 276.1 (8), 316.2 (4.4), 330.3 (0.4), 352.2 (0.4), 380.3 (0.4), 394.3 (1.2), 396.2 (2), 418.2 (3), 427.2 (4.7), 436.2 (12.6), 437.3 (3.2), 454.2 (100), 455.2 (11.8) | −2.3 | 3.8↓ | 4-hydroxy-11-oxo-2-undecenoic acid |
| 15 * | 8.9 | 330.1811 | 330.1812 | C18H24N3O3 | 216.1 (1.2), 218.1 (2.4), 232.2 (1.2), 258.3 (0.5), 260.1 (3.6), 261.1 (31), 262.1 (3.1), 276.3 (0.5), 287.2 (2.4), 301.2 (1.2), 302.3 (1.1), 312.1 (7.4), 313.2 (100), 314.1 (3.1) | 0.3 | 1.6↓ | butanal |
| 16 | 9.4 | 388.2226 | 388.2229 | C21H30N3O4 | 216.1 (0.9), 232.1 (0.7), 244.1 (10), 261.2 (0.5), 273.2 (0.3), 276.2 (0.3), 302.2 (3.4), 316.2 (0.2), 332.2 (0.6), 344.1 (0.4), 356.2 (100), 370.3 (6), 371.2 (28) | 0.8 | 2.0↓ | 7-hydroxy-heptanal |
| 17 * | 9.9 | 288.1346 | 288.1344 | C15H18N3O3 | 178.2 (11), 202.2 (14), 216.1 (32), 217.1 (19), 225.2 (16), 230.2 (32), 232.2 (63), 260.1 (100), 261.1 (46), 262.2 (63), 270.1 (35) | −0.7 | 5.3↓ | Unknown m/z 288.13, formaldehyde-related signal 1 |
| 18 | 10.1 | 496.2807 | 496.2802 | C28H38N3O5 | 218.2 (2.1), 244.1 (100), 261.2 (1.1), 276.2 (10.5), 316.1 (57.9), 340.3 (2.1), 351.3 (3.7), 368.1 (2.1), 380.2 (51.6), 381.2 (10.5), 396.2 (6.3), 436.2 (12.1), 438.3 (24.2), 460.1 (4.2), 478.3 (34.7) | −1.0 | 1.7↑ | unknown, m/z 496.28, signal 4 |
| 19 * | 10.2 | 368.1964 | 368.1968 | C21H26N3O3 | 108.0 (100), 201.3 (0.5), 218.2 (8.4), 261.1 (16.8), 276.1 (18.9), 302.1 (9.5), 324.3 (5.8), 333.3 (3.7), 339.0 (1.1), 348.2 (4.2), 350.3 (13.2), 351.3 (17.9), 352.2 (8.9) | 1.1 | 1.7↓ | 2,4-heptadienal |
| 20 * | 10.3 | 444.2495 | 444.2490 | C24H34N3O5 | 184.1 (20.9), 218.1 (3.5), 262.1 (4.0), 276.1 (4.7), 316.1 (4.0), 346.1 (1.2), 356.2 (2.3), 370.3 (1.2), 384.2 (17.4), 401.2 (4.7), 412.2 (100), 425.3 (20.9), 426.2 (15.1), 427.2 (31.4) | −1.1 | 1.9↓ | 10-oxo-4-hydroxy-decanal |
| 21 * | 11.0 | 288.1344 | 288.1344 | C15H18N3O3 | 176.1 (9), 177.1 (19), 178.4 (8), 190.2 (8), 204.3 (13), 216.2 (50), 229.3 (9), 231.0 (20), 232.1 (75), 233.1 (28), 260.3 (66), 261.2 (91), 262.2 (100), 271.1 (31) | 0.0 | 2.1↓ | Unknown m/z 288.13, formaldehyde-related signal 2 |
| 22 | 13.5 | 758.5685 | 758.5667 | C40H79N4O7P | 244.1 (9), 281.1 (3), 307.3 (3), 325.2 (12), 327.2 (14), 353.3 (4), 387.3 (4), 423.3 (6), 453.2 (22), 455.3 (30), 475.2 (63), 477.2 (100), 478.2 (20), 479.3 (5), 501.4 (4), 529.5 (3), 573.5 (3), 601.5 (24), 603.6 (70), 611.5 (3), 629.6 (40), 646.6 (13), 671.6 (10), 699.6 (20), 701.6 (11), 713.5 (6), 723.6 (8), 739.6 (13), 740.6 (30), 741.5 (22), 759.5 (46) | −2.4 | 4.4↓ | unknown, m/z 758.57, signal 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soboleva, A.; Frolova, N.; Bureiko, K.; Shumilina, J.; Balcke, G.U.; Zhukov, V.A.; Tikhonovich, I.A.; Frolov, A. Dynamics of Reactive Carbonyl Species in Pea Root Nodules in Response to Polyethylene Glycol (PEG)-Induced Osmotic Stress. Int. J. Mol. Sci. 2022, 23, 2726. https://doi.org/10.3390/ijms23052726
Soboleva A, Frolova N, Bureiko K, Shumilina J, Balcke GU, Zhukov VA, Tikhonovich IA, Frolov A. Dynamics of Reactive Carbonyl Species in Pea Root Nodules in Response to Polyethylene Glycol (PEG)-Induced Osmotic Stress. International Journal of Molecular Sciences. 2022; 23(5):2726. https://doi.org/10.3390/ijms23052726
Chicago/Turabian StyleSoboleva, Alena, Nadezhda Frolova, Kseniia Bureiko, Julia Shumilina, Gerd U. Balcke, Vladimir A. Zhukov, Igor A. Tikhonovich, and Andrej Frolov. 2022. "Dynamics of Reactive Carbonyl Species in Pea Root Nodules in Response to Polyethylene Glycol (PEG)-Induced Osmotic Stress" International Journal of Molecular Sciences 23, no. 5: 2726. https://doi.org/10.3390/ijms23052726
APA StyleSoboleva, A., Frolova, N., Bureiko, K., Shumilina, J., Balcke, G. U., Zhukov, V. A., Tikhonovich, I. A., & Frolov, A. (2022). Dynamics of Reactive Carbonyl Species in Pea Root Nodules in Response to Polyethylene Glycol (PEG)-Induced Osmotic Stress. International Journal of Molecular Sciences, 23(5), 2726. https://doi.org/10.3390/ijms23052726