
The Tiny Companion Matters: The Important Role of Protons in Active Transports in Plants





Abstract
:1. Introduction
2. Phytohormone Transporters
2.1. Overview of ABC Transporters
2.1.1. The Deprotonation of Auxin Molecules in Cytoplasm Implies the Need for Active Transporters
2.1.2. The Deprotonation of ABA Molecules in the Apoplast Implies the Need for Active Transporters
2.2. Overview of MATE Transporters
The Phytohormone Transport Activities of MATE Transporters Could Be Dependent on pH or Electrochemical Proton Gradient
3. Alkaloid Transporters
3.1. ABC Transporter for the Storage of Alkaloids
3.2. MATE Transporter for the Storage of Alkaloids
4. Ion and Ion Chelator Transporters
4.1. ABC Transporters for the Detoxification of Heavy Metal
4.2. The Ion/Ion Chelator Transport Activities of MATE Transporters Are pH Dependent
5. Sugar Transporters
5.1. Classification and Structural Properties
5.2. The SUT Family
5.2.1. The Activities of SUTs Are pH-Dependent
5.2.2. The Role of Sucrose Transport during Stress-Induced Cellular pH Fluctuations
5.3. The MST Family
5.3.1. Classification and Structural Properties
5.3.2. The Activities of MSTs Are pH-Dependent
5.3.3. Paralogs of MSTs Have Differential Expression Patterns to Serve Different Functions
6. Amino Acid Transporters
6.1. Classification and Structural Properties
6.2. Amino Acid Transporters Are Driven by Proton Motive Force
6.3. The Expressions of Amino Acid Transporters Are Stress-Responsive
6.4. Amino Acids as Osmolytes and Their Involvement in Ion Transport Mechanisms during Stress Responses
6.5. Amino Acid Accumulation and Salicylic Acid (SA) Signaling
6.6. Amino Acid Transporters Are Involved in the Regulation of Cellular pH and Rhizospheric pH
6.7. Protons Are the Unneglectable Regulators of Active Transporters under Stresses
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bañó-Polo, M.; Baeza-Delgado, C.; Orzáez, M.; Marti-Renom, M.A.; Abad, C.; Mingarro, I. Polar/ionizable residues in transmembrane segments: Effects on helix-helix packing. PLoS ONE 2012, 7, e44263. [Google Scholar] [CrossRef] [PubMed]
- Van Lehn, R.C.; Zhang, B.; Miller, T.F. Regulation of multispanning membrane protein topology via post-translational annealing. Elife 2015, 4, e08697. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S. PH as a stress signal. Plant Growth Regul. 1999, 29, 87–99. [Google Scholar] [CrossRef]
- Kader, M.A.; Lindberg, S. Cytosolic calcium and pH signaling in plants under salinity stress. Plant Signal. Behav. 2010, 5, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Staiger, C.J. Understanding cytoskeletal dynamics during the plant immune response. Annu. Rev. Phytopathol. 2018, 56, 513–533. [Google Scholar] [CrossRef]
- Cosse, M.; Seidel, T. Plant proton pumps and cytosolic pH-homeostasis. Front. Plant Sci. 2021, 12, 672873. [Google Scholar] [CrossRef]
- Morsomme, P.; Boutry, M. The plant plasma membrane H+-ATPase: Structure, function and regulation. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [Green Version]
- Sze, H.; Li, X.; Palmgren, M.G.; Sze, H.; Li, X.; Palmgrenb, M.G. Energization of plant cell membranes by H+-pumping ATPases: Regulation and biosynthesis. Plant Cell 1999, 11, 677–689. [Google Scholar]
- Haruta, M.; Gray, W.M.; Sussman, M.R. Regulation of the plasma membrane proton pump (H+-ATPase) by phosphorylation. Curr. Opin. Plant Biol. 2016, 28, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Janicka-Russak, M. Plant plasma membrane H+-ATPase in adaptation of plants to abiotic stresses. In Abiotic Stress Response in Plants-Physiological, Biochemical and Genetic Perspectives; Shanker, A., Venkatteswarlu, B., Eds.; Intech Open: London, UK, 2011. [Google Scholar]
- Davies, J.M. Vacuolar energization: Pumps, shunts and stress. J. Exp. Bot. 1997, 48, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, R. Structure, function and regulation of the plant vacuolar H+-translocating ATPase. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Segami, S.; Asaoka, M.; Kinoshita, S.; Fukuda, M.; Nakanishi, Y.; Maeshima, M. Biochemical, structural and physiological characteristics of vacuolar H+-pyrophosphatase. Plant Cell Physiol. 2018, 59, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, Y.; Martinoia, E.; Geisler, M. Plant hormone transporters: What we know and what we would like to know. BMC Biol. 2017, 15, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henikoff, S.; Greene, E.A.; Pietrokovski, S.; Bork, P.; Attwood, T.K.; Hood, L. Gene families: The taxonomy of protein paralogs and chimeras. Science 1997, 278, 5338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Lane, T.S.; Rempe, C.S.; Davitt, J.; Staton, M.E.; Peng, Y.; Soltis, D.E.; Melkonian, M.; Deyholos, M.; Leebens-Mack, J.H.; Chase, M.; et al. Diversity of ABC transporter genes across the plant kingdom and their potential utility in biotechnology. BMC Biotechnol. 2016, 16, 47. [Google Scholar] [CrossRef] [Green Version]
- Martinola, E.; Grill, E.; Tommaslnl, R.; Kreuz, K.; Amrhein, N. ATP-dependent glutathione S-conjugate “export” pump in the vacuolar membrane of plants. Nature 1993, 364, 247–249. [Google Scholar] [CrossRef]
- Do, T.H.T.; Martinoia, E.; Lee, Y. Functions of ABC transporters in plant growth and development. Curr. Opin. Plant Biol. 2018, 41, 32–38. [Google Scholar] [CrossRef]
- Hwang, J.; Song, W.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 338–355. [Google Scholar] [CrossRef] [Green Version]
- Ha, T.; Do, T.; Choi, H.; Palmgren, M.; Martinoia, E.; Hwang, J.-U.; Lee, Y. Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes. Proc. Natl. Acad. Sci. USA 2019, 116, 25. [Google Scholar]
- Footitt, S.; Slocombe, S.P.; Larner, V.; Kurup, S.; Wu, Y.; Larson, T.; Graham, I.; Baker, A.; Holdsworth, M. Control of germination and lipid mobilization by COMATOSE, the Arabidopsis homologue of human ALDP. EMBO J. 2002, 21, 2912–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz, H.-H.; Scharnewski, M.; Feussner, K.; Feussner, I.; Flügge, U.-I.; Fulda, M.; Gierth, M. The ABC transporter PXA1 and peroxisomal β-oxidation are vital for metabolism in mature leaves of Arabidopsis during extended darkness. Plant Cell 2009, 21, 2733–2749. [Google Scholar] [CrossRef] [Green Version]
- Dahuja, A.; Kumar, R.R.; Sachdev, A.; Watts, A.; Singh, B.; Goswami, S.; Sachdev, A.; Praveen, S. Role of ATP-binding cassette transporters in maintaining plant homeostasis under abiotic and biotic stresses. Physiol. Plant. 2021, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-Y.; Park, J.; Mendoza-Cózatl, D.G.; Suter-Grotemeyer, M.; Shim, D.; Hörtensteiner, S.; Geisler, M.; Weder, B.; Rea, P.A.; Rentsch, D.; et al. Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc. Natl. Acad. Sci. USA 2010, 49, 21187–21192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Voithenberg, L.V.; Park, J.; Stübe, R.; Lux, C.; Lee, Y.; Philippar, K. A novel prokaryote-type ECF/ABC transporter module in chloroplast metal hoemostasis. Front. Plant Sci. 2019, 10, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crouzet, J.; Roland, J.; Peeters, E.; Trombik, T.; Ducos, E.; Nader, J.; Boutry, M. NtPDR1, a plasma membrane ABC transporter from Nicotiana tabacum, is involved in diterpene transport. Plant Mol. Biol. 2013, 82, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Zažímalová, E.; Murphy, A.S.; Yang, H.; Hoyerová, K.; Hošek, P. Auxin transporters-Why so many? Cold Spring Harb. Perspect. Biol. 2010, 2, a001552. [Google Scholar]
- Geisler, M. Getting to the right side. Plant Physiol. 2016, 172, 2081. [Google Scholar] [CrossRef] [PubMed]
- Cecchetti, V.; Brunetti, P.; Napoli, N.; Fattorini, L.; Altamura, M.M.; Costantino, P.; Cardarelli, M. ABCB1 and ABCB19 auxin transporters have synergistic effects on early and late Arabidopsis anther development. J. Integr. Plant Biol. 2015, 57, 1089–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, B.; Murphy, A.S.; Spalding, E.P. Multidrug Resistance-like genes of Arabidopsis required for auxin transport and auxin-mediated development. Plant Cell 2001, 13, 2441–2454. [Google Scholar]
- Yang, H.; Murphy, A.S. Functional expression and characterization of Arabidopsis ABCB, AUX 1 and PIN auxin transporters in Schizosaccharomyces pombe. Plant J. 2010, 59, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.; Subudhi, P.K. Comprehensive analysis and expression profiling of the OsLAX and OsABCB auxin transporter gene families in rice (Oryza sativa) under phytohormone stimuli and abiotic stresses. Front. Plant Sci. 2016, 7, 593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribba, T.; Garrido-Vargas, F.; O’Brien, J.A. Auxin-mediated responses under salt stress: From developmental regulation to biotechnological applications. J. Exp. Biol. 2020, 71, 3843–3853. [Google Scholar] [CrossRef] [PubMed]
- Ku, Y.-S.; Sintaha, M.; Cheung, M.-Y.; Lam, H.-M. Plant hormone signaling crosstalks between biotic and abiotic stress responses. Int. J. Mol. Sci. 2018, 19, 3206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Intertissue signal transfer of abscisic acid from vascular. Plant Physiol. 2014, 164, 1587–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.; Koshiba, T. Transport of ABA from the site of biosynthesis to the site of action. J. Plant Res. 2011, 124, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Geilfus, C.-M. The pH of the apoplast: Dynamic factor with functional impact under stress. Mol. Plant 2017, 10, 1371–1386. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Miyaji, T.; Yabuuchi, H.; Shimizu, H.; Sugimoto, E.; Kamiya, A.; Moriyama, Y.; Shinozaki, K. ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc. Natl. Acad. Sci. USA 2010, 107, 2361–2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.; Hwang, J.; Lee, M.; Kim, Y.-Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krattinger, S.G.; Kang, J.; Bräunlich, S.; Boni, R.; Chauhan, H.; Selter, L.L.; Robinson, M.D.; Schmid, M.W.; Wiederhold, E.; Hensel, G.; et al. Abscisic acid is a substrate of the ABC transporter encoded by the durable wheat disease resistance gene Lr34. New Phytol. 2019, 223, 853–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omote, H.; Hiasa, M.; Matsumoto, T.; Otsuka, M.; Moriyama, Y. The MATE proteins as fundamental transporters of metabolic and xenobiotic organic cations. Trends Pharmacol. Sci. 2006, 27, 11. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, K.; Shitan, N.; Yazaki, K. The multidrug and toxic compound extrusion (MATE) family in plants. Plant Biotechnol. 2014, 31, 417–430. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, H.; Pan, Y.; Yu, Y.; Luan, S.; Li, L. A DTX/MATE-type transporter facilitates abscisic acid efflux and modulates ABA sensitivity and drought tolerance in Arabidopsis. Mol. Plant 2014, 7, 1522–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.; Davies, W.J. Xylem sap pH increase: A drought signal received at the apoplastic face of the guard cell that involves the suppression of saturable abscisic acid uptake by the epidermal Symplast. Plant Physiol. 1997, 113, 559–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, M.; Wang, B.; Aryal, B.; Garcion, C.; Abou-Mansour, E.; Heck, S.; Geisler, M.; Mauch, F.; Nawrath, C.; Métraux, J.-P. Export of salicylic acid from the chloroplast requires the multidrug and toxin extrusion-like transporter EDS5. Plant Physiol. 2013, 162, 1815–1821. [Google Scholar] [CrossRef] [Green Version]
- Nawrath, C.; Métraux, J.-P. Salicylic acid induction-deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin after pathogen inoculation. Plant Cell 1999, 11, 1393–1404. [Google Scholar] [PubMed] [Green Version]
- Lefevere, H.; Bauters, L.; Gheysen, G. Salicylic acid biosynthesis in plants. Front. Plant Sci. 2020, 11, 338. [Google Scholar] [CrossRef] [PubMed]
- Nawrath, C.; Heck, S.; Parinthawong, N.; Métraux, J.-P. EDS5, an essential component of salicylic acid-dependent signaling for disease resistance in Arabidopsis, is a member of the MATE transporter family. Plant Cell 2002, 14, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.; Blake-Kalff, M.; Davies, T.G.E. Detoxification of xenobiotics by plants: Chemical modification and vacuolar compartmentation. Trends Plant Sci. 1997, 2, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Biol. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Bennett, B.Y.R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Shitan, N. Secondary metabolites in plants: Transport and self-tolerance mechanisms. Biosci. Biotechnol. Biochem. 2016, 80, 1283–1293. [Google Scholar] [CrossRef] [Green Version]
- Shitan, N.; Bazin, I.; Dan, K.; Obata, K.; Kigawa, K.; Ueda, K.; Sato, F.; Forestier, C.; Yazaki, K. Involvement of CjMDR1, a plant multidrug-resistance-type ATP-binding cassette protein, in alkaloid transport in Coptis japonica. Proc. Natl. Acad. Sci. USA 2003, 100, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Morita, M.; Shitan, N.; Sawada, K.; Van Mongtagu, M.C.E.; Inzéc, D.; Rischer, H.; Goossens, A.; Oksman-caldentey, K.; Moriyama, Y.; Yazaki, K. Vacuolar transport of nicotine is mediated by a multidrug and toxic compound extrusion (MATE) transporter in Nicotiana tabacum. Proc. Natl. Acad. Sci. USA 2009, 106, 2447–2452. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Song, W.-Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef]
- Song, W.-Y.; Yamaki, T.; Yamaji, N.; Ko, D.; Jung, K.H.; Fujii-Kashino, M.; An, G.; Martinoia, E.; Lee, Y.; Ma, J.F. A rice ABC transporter, OsABCC1, reduces arsenic accumulation in the grain. Proc. Natl. Acad. Sci. USA 2014, 111, 15699–15704. [Google Scholar] [CrossRef] [Green Version]
- Song, W.-Y.; Park, J.; Eisenach, C.; Maeshima, M.; Lee, Y.; Martinoia, E. ABC transporters and heavy metals. In Plant ABC Transporters; Geisler, M., Ed.; Springer International Publishing: Cham, Switzerland, 2014; pp. 1–17. ISBN 9783319065113. [Google Scholar]
- Doshi, R.; Mcgrath, A.P.; Piñeros, M.; Szewczyk, P.; Garza, D.M.; Kochian, L.V.; Chang, G. Functional characterization and discovery of modulators of SbMATE, the agronomically important aluminium tolerance transporter from Sorghum bicolor. Sci. Rep. 2017, 7, 17996. [Google Scholar] [CrossRef] [Green Version]
- Marinova, K.; Pourcel, L.; Weder, B.; Schwarz, M.; Barron, D.; Routaboul, J.-M.; Debeaujon, I.; Klein, M. The Arabidopsis MATE transporter TT12 acts as a vacuolar flavonoid/H+- antiporter active in proanthocyanidin-accumulating cells of the seed coat. Plant Cell 2007, 19, 2023–2038. [Google Scholar] [CrossRef] [Green Version]
- Durrett, T.P.; Gassmann, W.; Rogers, E.E. The FRD3-mediated efflux of citrate into the root vasculature is necessary for efficient iron translocation. Plant Physiol. 2007, 144, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; He, Z.; Pandey, G.K.; Tsuchiya, T.; Luan, S. Functional cloning and characterization of a plant efflux carrier for multidrug and heavy metal detoxification. J. Biol. Chem. 2002, 277, 5360–5368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Wei, J.; Li, D.; Kong, X.; Rengel, Z.; Chen, L.; Yang, Y.; Cui, X.; Chen, Q. The role of the plasma membrane H+-ATPase in plant responses to aluminum toxicity. Front. Plant Sci. 2017, 8, 1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. Molecular mechanisms for coping with Al toxicity in plants. Int. J. Mol. Sci. 2019, 20, 1551. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhao, F.; Tang, R.; Yu, Y.; Song, J.; Wang, Y.; Li, L. Two tonoplast MATE proteins function as turgor-regulating chloride channels in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 144, E2036–E2045. [Google Scholar] [CrossRef] [Green Version]
- Misra, V.A.; Wafula, E.K.; Wang, Y.; DePamphilis, C.W.; Timko, M.P. Genome-wide identification of MST, SUT and SWEET family sugar transporters in root parasitic angiosperms and analysis of their expression during host parasitism. BMC Plant Biol. 2019, 19, 196. [Google Scholar] [CrossRef]
- Doidy, J.; Vidal, U.; Lemoine, R. Sugar transporters in Fabaceae, featuring SUT MST and SWEET families of the model plant Medicago truncatula and the agricultural crop Pisum sativum. PLoS ONE 2019, 14, e0223173. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, Y.; Wei, Q.; Wan, H.; Sun, C. Phylogenetic relationships of sucrose transporters (SUTs) in plants and genome-wide characterization of SUT genes in Orchidaceae reveal roles in floral organ development. PeerJ 2021, 9, e11961. [Google Scholar] [CrossRef]
- Drew, D.; North, R.A.; Nagarathinam, K.; Tanabe, M. Structures and general transport mechanisms by the major facilitator superfamily (MFS). Chem. Rev. 2021, 121, 5289–5335. [Google Scholar] [CrossRef]
- Jeena, G.S.; Kumar, S.; Shukla, R.K. Structure, evolution and diverse physiological roles of SWEET sugar transporters in plants. Plant Mol. Biol. 2019, 100, 351–365. [Google Scholar] [CrossRef]
- Eom, J.; Chen, L.; Sosso, D.; Julius, B.T.; Lin, I.W.; Qu, X.; Braun, D.M.; Frommer, W.B. SWEETs, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalonde, S.; Frommer, W.B. SUT sucrose and MST monosaccharide transporter inventory of the Selaginella genome. Front. Plant Sci. 2012, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nookaraju, A.; Upadhyaya, C.P.; Pandey, S.K.; Young, K.E.; Hong, S.J.; Park, S.K.; Park, S.W. Molecular approaches for enhancing sweetness in fruits and vegetables. Sci. Hortic. 2010, 127, 1–15. [Google Scholar] [CrossRef]
- Julius, B.T.; Leach, K.A.; Tran, T.M.; Mertz, R.A.; Braun, D.M. Sugar transporters in plants: New insights and discoveries. Plant Cell Physiol. 2017, 58, 1442–1460. [Google Scholar] [CrossRef] [Green Version]
- Schulz, A.; Beyhl, D.; Marten, I.; Wormit, A.; Neuhaus, E.; Poschet, G.; Büttner, M.; Schneider, S.; Sauer, N.; Hedrich, R. Proton-driven sucrose symport and antiport are provided by the vacuolar transporters SUC4 and TMT1/2. Plant J. 2011, 68, 129–136. [Google Scholar] [CrossRef]
- Weise, A.; Barker, L.; Kühn, C.; Lalonde, S.; Buschmann, H.; Frommer, W.B.; Ward, J.M. A new subfamily of sucrose transporters, SUT4, with low affinity/high capacity localized in enucleate sieve elements of plants. Plant Cell 2000, 12, 1345–1355. [Google Scholar] [CrossRef] [Green Version]
- Carpaneto, A.; Geiger, D.; Bamberg, E.; Sauer, N.; Fromm, J.; Hedrich, R. Phloem-localized, proton-coupled sucrose carrier ZmSUT1 mediates sucrose efflux under the control of the sucrose gradient and the proton motive force. J. Biol. Chem. 2005, 280, 21437–21443. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.; Liu, M.; Zhang, L.; Ruan, Y.; Ding, R.; Ji, Y.; Zhang, N.; Zhang, S.; Farmer, J.; Wang, C. Arabidopsis AtSUC2 and AtSUC4, encoding sucrose transporters, are required for abiotic stress tolerance in an ABA-dependent pathway. Physiol. Plant. 2015, 153, 119–136. [Google Scholar] [CrossRef]
- Wang, D.; Liu, H.; Wang, H.; Zhang, P.; Shi, C. A novel sucrose transporter gene IbSUT4 involves in plant growth and response to abiotic stress through the ABF-dependent ABA signaling pathway in Sweetpotato. BMC Plant Biol. 2020, 20, 157. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Knight, M.R.; Trewavas, A.J.; Sattelmacher, B.; Plieth, C. Self-reporting Arabidopsis expressing pH and [Ca2+] indicators unveil ion dynamics in the cytoplasm and in the apoplast under abiotic stress. Plant Physiol. 2004, 134, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.P.; Ward, J.M.; Sharkey, T.D. Phaseolus vulgaris SUT1.1 is a high affinity sucrose-proton co-transporter. Plant Direct 2020, 4, e00260. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Zhang, R. High temperature effects on electron and proton circuits of photosynthesis. J. Integr. Plant Biol. 2010, 52, 712–722. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, S.; Yunjuan, R.; Chen, S.; Liesche, J. Regulation of sucrose transporters and phloem loading in response to environmental cues. Plant Physiol. 2018, 176, 930–945. [Google Scholar] [CrossRef] [Green Version]
- Raghavendra, A.S.; Yin, Z.-H.; Heber, U. Light-dependent pH changes in leaves of C4 plants Comparison of the pH response to carbon dioxide and oxygen with that of C3 plants. Planta 1993, 189, 278–287. [Google Scholar] [CrossRef]
- Slewinski, T.L. Diverse functional roles of monosaccharide transporters and their homologs in vascular plants: A physiological perspective. Mol. Plant 2011, 4, 641–662. [Google Scholar] [CrossRef]
- Paulsen, P.A.; Custódio, T.F.; Pedersen, B.P. Crystal structure of the plant symporter STP10 illuminates sugar uptake mechanism in monosaccharide transporter superfamily. Nat. Commun. 2019, 10, 407. [Google Scholar] [CrossRef]
- Bavnhøj, L.; Paulsen, P.A.; Flores-Canales, J.C.; Schiøtt, B.; Pedersen, B.P. Molecular mechanism of sugar transport in plants unveiled by structures of glucose/H+ symporter STP10. Nat. Plants 2021, 7, 1409–1419. [Google Scholar] [CrossRef]
- Klepek, Y.; Geiger, D.; Stadler, R.; Klebl, F.; Landouar-arsivaud, L. Arabidopsis POLYOL TRANSPORTER5, a new member of the monosaccharide transporter-like superfamily, mediates H+-symport of numerous substrates, including myo-inositol, glycerol, and ribose. Plant Cell 2005, 17, 204–218. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Meng, D.; Piñeros, M.A.; Mao, Y.; Dandekar, A.M.; Cheng, L. A sugar transporter takes up both hexose and sucrose for sorbitol-modulated in vitro pollen tube growth in apple. Plant Cell 2020, 32, 449–469. [Google Scholar] [CrossRef] [Green Version]
- Schubert, M.; Koteyeva, N.K.; Wabnitz, P.W.; Santos, P.; Büttner, M.; Sauner, N.; Demchenko, K.; Pawlowski, K. Plasmodesmata distribution and sugar partitioning in nitrogen-fixing root nodules of Datisca glomerata. Planta 2011, 233, 139–152. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Kanai, M.; Osakabe, Y.; Ohiraki, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monosaccharide absorption activity of Arabidopsis roots depends on expression profiles of transporter genes under high salinity conditions. J. Biol. Chem. 2011, 286, 43577–43586. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; An, B.; Zhang, Y.; Yang, J.; Li, S.; Sun, T.; Li, Y. Sugar transporter proteins (STPs) in Gramineae crops: Comparative analysis, phylogeny, evolution, and expression profiling. Cells 2019, 8, 560. [Google Scholar] [CrossRef] [Green Version]
- Muratore, C.; Espen, L.; Prinsi, B. Nitrogen uptake in plants: The plasma membrane root transport systems from a physiological and proteomic perspective. Plants 2021, 10, 681. [Google Scholar] [CrossRef]
- Feng, H.; Fan, X.; Miller, A.J.; Xu, G. Plant nitrogen uptake and assimilation: Regulation of cellular pH homeostasis. J. Exp. Bot. 2020, 71, 4380–4392. [Google Scholar] [CrossRef] [Green Version]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino acid transporters in plants: Identification and function. Plants 2020, 9, 972. [Google Scholar] [CrossRef]
- Yang, G.; Wei, Q.; Huang, H.; Xia, J. Amino acid transporters in plant cells: A brief review. Plants 2020, 9, 967. [Google Scholar] [CrossRef]
- Ortiz-Lopez, A.; Chang, H.-C.; Bush, D.R. Amino acid transporters in plants. Biochim. Biophys. Acta 2000, 1465, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Okumoto, S.; Pilot, G. Amino acid export in plants: A missing link in nitrogen cycling. Mol. Plant 2011, 4, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Dinkeloo, K.; Boyd, S.; Pilot, G. Update on amino acid transporter functions and on possible amino acid sensing mechanisms in plants. Semin. Cell Dev. Biol. 2018, 74, 105–113. [Google Scholar] [CrossRef]
- Jack, D.L.; Paulsen, I.T.; Saier, J. The amino acid/polyamine/organocation (APC) superfamily of transporters specific for amino acids, polyamines and organocations. Microbiology 2000, 146, 1797–1814. [Google Scholar] [CrossRef] [Green Version]
- Young, G.B.; Jack, D.L.; Smith, D.W.; Saier, M.H., Jr. The amino acid/auxin: Proton symport permease family. Biochim. Biophys. Acta (BBA)-Biomembr. 1999, 1415, 306–322. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Pratelli, R.; Yu, S.; Shelley, B.; Collakova, E.; Pilot, G. Detailed characterization of the UMAMIT proteins provides insight into their evolution, amino acid transport properties, and role in the plant. J. Exp. Bot. 2021, 72, 6400–6417. [Google Scholar] [CrossRef]
- Williams, L.E.; Nelson, S.J.; Hall, J.L. Characterization of solute transport in plasma membrane vesicles isolated from cotyledons of Ricinus communis L.-II. Evidence for a proton-coupled mechanism for sucrose and amino acid uptake. Planta 1990, 182, 540–545. [Google Scholar] [CrossRef]
- Li, Z.C.; Bush, D.R. Structural determinants in substrate recognition by proton-amino acid symports in plasma membrane vesicles isolated from sugar beet leaves. Arch. Biochem. Biophys. 1992, 294, 519–526. [Google Scholar] [CrossRef]
- Fischer, W.N.; Kwart, M.; Hummel, S.; Frommer, W.B. Substrate specificity and expression profile of amino acid transporters (AAPs) in Arabidopsis. J. Biol. Chem. 1995, 270, 16315–16320. [Google Scholar] [CrossRef] [Green Version]
- Okumoto, S.; Koch, W.; Tegeder, M.; Fischer, W.N.; Biehl, A.; Leister, D.; Stierhof, Y.D.; Frommer, W.B. Root phloem-specific expression of the plasma membrane amino acid proton co-transporter AAP3. J. Exp. Bot. 2004, 55, 2155–2168. [Google Scholar] [CrossRef] [Green Version]
- Montamat, F.; Maurousset, L.; Tegeder, M.; Frommer, W.; Delrot, S. Cloning and expression of amino acid transporters from broad bean. Plant Mol. Biol. 1999, 41, 259–268. [Google Scholar] [CrossRef]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef] [Green Version]
- Rentsch, D.; Hirner, B.; Schmelzer, E.; Frommer, W.B. Salt stress-induced proline transporters and salt stress-repressed broad specificity amino acid permeases identified by suppression of a yeast amino acid permease-targeting mutant. Plant Cell 1996, 8, 1437–1446. [Google Scholar]
- Wang, J.J.; Hou, Q.Q.; Li, P.H.; Yang, L.; Sun, X.C.; Benedito, V.A.; Wen, J.Q.; Chen, B.B.; Mysore, K.S.; Zhao, J. Diverse functions of multidrug and toxin extrusion (MATE) transporters in citric acid efflux and metal homeostasis in Medicago truncatula. Plant J. 2017, 90, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Ma, H.; Yu, L.; Wang, X.; Zhao, J. Genome-wide survey and expression analysis of amino acid transporter gene family in rice (Oryza sativa L.). PLoS ONE 2012, 7, e49210. [Google Scholar] [CrossRef]
- Wan, Y.; King, R.; Mitchell, R.A.C.; Hassani-Pak, K.; Hawkesford, M.J. Spatiotemporal expression patterns of wheat amino acid transporters reveal their putative roles in nitrogen transport and responses to abiotic stress. Sci. Rep. 2017, 7, 5461. [Google Scholar] [CrossRef]
- Ueda, A.; Shi, W.; Sanmiya, K.; Shono, M.; Takabe, T. Functional analysis of salt-inducible proline transporter of barley roots. Plant Cell Physiol. 2001, 42, 1282–1289. [Google Scholar] [CrossRef] [Green Version]
- Tian, R.; Yang, Y.; Chen, M. Genome-wide survey of the amino acid transporter gene family in wheat (Triticum aestivum L.): Identification, expression analysis and response to abiotic stress. Int. J. Biol. Macromol. 2020, 162, 1372–1387. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, S.; Zhou, T.; Yue, C.; Hua, Y.; Huang, J. Genome-wide identification of Brassicaceae B-BOX genes and molecular characterization of their transcriptional responses to various nutrient stresses in allotetraploid rapeseed. BMC Plant Biol. 2021, 21, 288. [Google Scholar] [CrossRef]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [Green Version]
- El Moukhtari, A.; Cabassa-Hourton, C.; Farissi, M.; Savouré, A. How does proline treatment promote salt stress tolerance during crop plant development? Front. Plant Sci. 2020, 11, 1127. [Google Scholar] [CrossRef]
- Ben Ahmed, C.; Magdich, S.; Ben Rouina, B.; Sensoy, S.; Boukhris, M.; Ben Abdullah, F. Exogenous proline effects on water relations and ions contents in leaves and roots of young olive. Amino Acids 2011, 40, 565–573. [Google Scholar] [CrossRef]
- Huang, Y.; Bie, Z.; Liu, Z.; Zhen, A.; Wang, W. Protective role of proline against salt stress is partially related to the improvement of water status and peroxidase enzyme activity in cucumber. Soil Sci. Plant Nutr. 2009, 55, 698–704. [Google Scholar] [CrossRef] [Green Version]
- Lone, M.I.; Kueh, J.S.H.; Wyn Jones, R.G.; Bright, S.W.J. Influence of proline and glycinebetaine on salt tolerance of cultured barley embryos. J. Exp. Bot. 1987, 38, 479–490. [Google Scholar] [CrossRef]
- Buell, C.R.; Joardar, V.; Lindeberg, M.; Selengut, J.; Paulsen, I.T.; Gwinn, M.L.; Dodson, R.J.; Deboy, R.T.; Durkin, A.S.; Kolonay, J.F.; et al. The complete genome sequence of the Arabidopsis and tomato pathogen Pseudomonas syringae pv. tomato DC3000. Proc. Natl. Acad. Sci. USA 2003, 100, 10181–10186. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Ji, Y.; Bhuiyan, N.H.; Pilot, G.; Selvaraj, G.; Zou, J.; Wei, Y. Amino acid homeostasis modulates salicylic acid-associated redox status and defense responses in Arabidopsis. Plant Cell 2010, 22, 3845–3863. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Postel, S.; Kemmerling, B.; Ludewig, U. Altered growth and improved resistance of Arabidopsis against Pseudomonas syringae by overexpression of the basic amino acid transporter AtCAT1. Plant Cell Environ. 2014, 37, 1404–1414. [Google Scholar] [CrossRef]
- Song, J.T.; Lu, H.; Mcdowell, J.M.; Greenberg, J.T. A key role for ALD1 in activation of local and systemic defenses in Arabidopsis. Plant J. 2004, 40, 200–212. [Google Scholar] [CrossRef]
- Song, J.T.; Lu, H.; Greenberg, J.T. Divergent roles in Arabidopsis thaliana development and defense of two homologous genes, Aberrant Growth and Death2 and Agd2-Like Defense Response Protein1, encoding novel aminotransferases. Plant Cell 2004, 16, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Näsholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2008, 182, 31–48. [Google Scholar] [CrossRef]
- Raven, J.A.; Smith, F.A. Nitrogen assimilation and transport in vacuolar land plants in relation to intraceuular pH regulation. New Phytol. 1976, 76, 415–431. [Google Scholar] [CrossRef]
- Snowden, C.J.; Thomas, B.; Baxter, C.J.; Smith, J.A.C.; Sweetlove, L.J. A tonoplast Glu/Asp/GABA exchanger that affects tomato fruit amino acid composition. Plant J. 2015, 81, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Ramesh, S.A.; Gilliham, M.; Tyerman, S.D.; Bose, J. Role of TaALMT1 malate-GABA transporter in alkaline pH tolerance of wheat. Plant Cell Environ. 2020, 43, 2443–2459. [Google Scholar] [CrossRef]
- Rolin, D.; Baldet, P.; Just, D.; Chevalier, C.; Biran, M.; Raymond, P. NMR study of low subcellular pH during the development of cherry tomato fruit. Aust. J. Plant Physiol. 2000, 27, 61–69. [Google Scholar]
- Grewer, C.; Gameiro, A.; Zhang, Z.; Tao, Z.; Braams, S.; Rauen, T. Glutamate forward and reverse transport: From molecular mechanism to transporter-mediated release after ischemia. IUBMB Life 2008, 60, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.; Koshy, C.; Yildiz, Ö.; Ziegler, C. Alternating-access mechanism in conformationally asymmetric trimers of the betaine transporter BetP. Nature 2012, 490, 126–130. [Google Scholar] [CrossRef]
- Yernool, D.; Boudker, O.; Jin, Y.; Gouaux, E. Structure of a glutamate transporter homologue from Pyrococcus horikoshii. Nature 2004, 431, 811–818. [Google Scholar] [CrossRef] [Green Version]
- Nogia, P.; Pati, P.K. Plant secondary metabolite transporters: Diversity, functionality, and their modulation. Front. Plant Sci. 2021, 12, 758202. [Google Scholar] [CrossRef]


{kind=link}
| Transporter Type | Transporter Name | Transport Activity | pH Dependence | Biological Significance | References |
|---|---|---|---|---|---|
| MATE | Nt-JAT1 | Mediates nicotine influx into the vacuole | Makes use of pH gradient across vacuolar membrane, with pH inside vacuole lower than in cytosol | Storage of toxic compound | [57] |
| AtDTX50 | Mediates ABA efflux under drought stress | Most active at pH 7 when compared among pH 6, 7, and 8 | Promotes ABA efflux under drought stress when the pH of the xylem sap becomes closer to neutral | [46] | |
| AtDTX33 | Mediates Cl− influx into vacuoles; mutant impaired in stomatal opening | Most active at vacuolar pH 5 when compared among vacuolar pH 5, 6, and 7 | Promotes Cl− influx into the vacuole, which is more acid than the cytosol, for turgor regulation | [67] | |
| AtDTX35 | Mediates Cl− influx into vacuoles; mutant impaired in stomatal opening | Most active at vacuolar pH 5 when compared among vacuolar pH 5, 6, and 7 | Promotes Cl− influx into the vacuole, which is more acid than the cytosol, for turgor regulation | [67] | |
| AtEDS5 | Mediates SA efflux from chloroplast to cytoplasm | The transport activity is driven by the proton gradient across the biological membrane | The efflux of SA from chloroplast to cytosol promotes stress tolerance | [48,51] | |
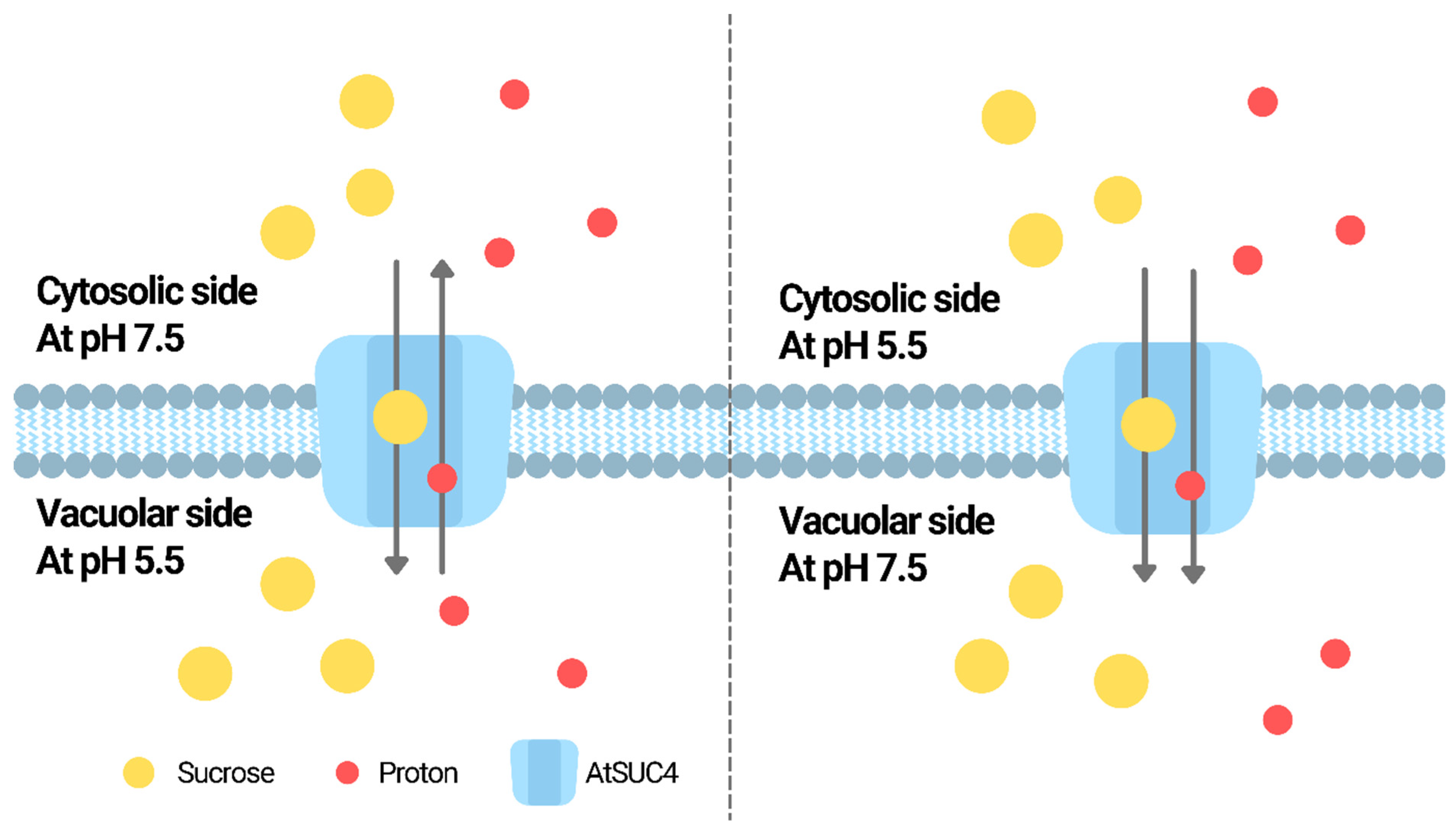
| SUC | AtSUC4 | Mediates vacuolar sucrose storage | Acts as a H+/sucrose antiporter or symporter depending on the pH difference between vacuole lumen and the medium outside | Facilitates sugar distribution under stress; compared to the wild-type, mutants have higher and lower sucrose, fructose and glucose in shoots and roots, respectively, and are more sensitive to salt, osmotic, cold and ABA treatments | [78,79,81] |
| PvSUT1.1 | Exports sucrose from leaf through phloem | Higher activity at lower pH in medium | Involved in sucrose translocation between different tissues of plant, the downregulated expression is possibly associated with the heat susceptibility of the plant | [84] | |
| MST | AtPLT5 | Mediates transport of a large spectrum of polyols | Maximal transport activity at pH 5.5; activity reduced at pH 6.5 and no activity at pH 7 | Proposed to be involved in the retrieval of sugars from the apoplast | [91] |
| MdSTP13a | Mediates transport of a hexose and sucrose for pollen tube growth | Optimal uptake at pH 6 in yeast model | Growth and development | [92] | |
| AtSTP1 | Inducible by salinity but mainly involved in the distribution of monosaccharides under normal conditions; mutant with reduced ability to uptake glucose, fructose and galactose | unknown | Growth and development, adaptation to the environment | [94] | |
| AtSTP13 | Inducible by salinity and ABA treatments; involved in the reabsorption of monosaccharides leaked from damaged cells; mutant with reduced ability to uptake glucose, fructose and galactose | unknown | Growth and development, adaptation to the environment | [94] | |
| ATF | HvProT | Proline transportation during salt stress | pH-dependent; the proline uptake activity o yeast mutant complemented with HvProT was the highest at pH 4.5 among pH 4.5, 5.5, and 6.5 | Adaptation to the environment | [116] |
| AAP | AtLHT1 | Transports a broad spectrum of amino acids; knockout mutant with reduced susceptibility to P. syringae; suspected to be involved in SA pathway | pH gradient dependent | Resistance to biotic stress | [111,125] |
| APC | AtCAT1 | Lysine incorporation; overexpressor more resistant to P. syringae with increased SA level in leaves | unknown | Resistance to biotic stress | [126] |
| SlCAT9 | Exchange of GABA for glutamate and aspartate during fruit ripening | The transport of GABA has been suggested to play a role in regulating cytosolic pH | Growth and development | [131] | |
| ABC | AtABCB1 | Transports auxin; double mutant with Atabcb19 resulted in developmental problem with poor fertility | unknown | Growth and development | [31] |
| AtABCB19 | Transports auxin; double mutant with Atabcb1 resulted in developmental problem with poor fertility | unknown | Growth and development | [31] | |
| AtABCG25 | Exports ABA; mutant with ABA-sensitive phenotype at early growth stage | unknown | Adaptation to the environment | [41] | |
| AtABCG40 | Uptakes ABA; mutant with slow uptake of ABA and insensitivity towards ABA | unknown | Adaptation to the environment | [42] | |
| CjMDR1 | Transports berberine from root to rhizome | unknown | Adaptation to the environment | [56] | |
| Lr34 | Transports ABA; ectopic expression in wheat enhanced the tolerance to rice blast | unknown | Adaptation to the environment | [43] | |
| AtABCC1 | Mediates microsomal uptake of PC and PC conjugates for heavy metal detoxification in vacuole | unknown | Adaptation to the environment | [26] | |
| AtABCC2 | Mediates microsomal uptake of PC and PC conjugates for heavy metal detoxification in vacuole | unknown | Adaptation to the environment | [26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ku, Y.-S.; Cheng, S.-S.; Ng, M.-S.; Chung, G.; Lam, H.-M. The Tiny Companion Matters: The Important Role of Protons in Active Transports in Plants. Int. J. Mol. Sci. 2022, 23, 2824. https://doi.org/10.3390/ijms23052824
Ku Y-S, Cheng S-S, Ng M-S, Chung G, Lam H-M. The Tiny Companion Matters: The Important Role of Protons in Active Transports in Plants. International Journal of Molecular Sciences. 2022; 23(5):2824. https://doi.org/10.3390/ijms23052824
Chicago/Turabian StyleKu, Yee-Shan, Sau-Shan Cheng, Ming-Sin Ng, Gyuhwa Chung, and Hon-Ming Lam. 2022. "The Tiny Companion Matters: The Important Role of Protons in Active Transports in Plants" International Journal of Molecular Sciences 23, no. 5: 2824. https://doi.org/10.3390/ijms23052824
APA StyleKu, Y. -S., Cheng, S. -S., Ng, M. -S., Chung, G., & Lam, H. -M. (2022). The Tiny Companion Matters: The Important Role of Protons in Active Transports in Plants. International Journal of Molecular Sciences, 23(5), 2824. https://doi.org/10.3390/ijms23052824