
The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years?

Abstract
:1. The Hypothalamus and Sleep: A Brief History
2. Discovery of the Sleep-Active VLPO
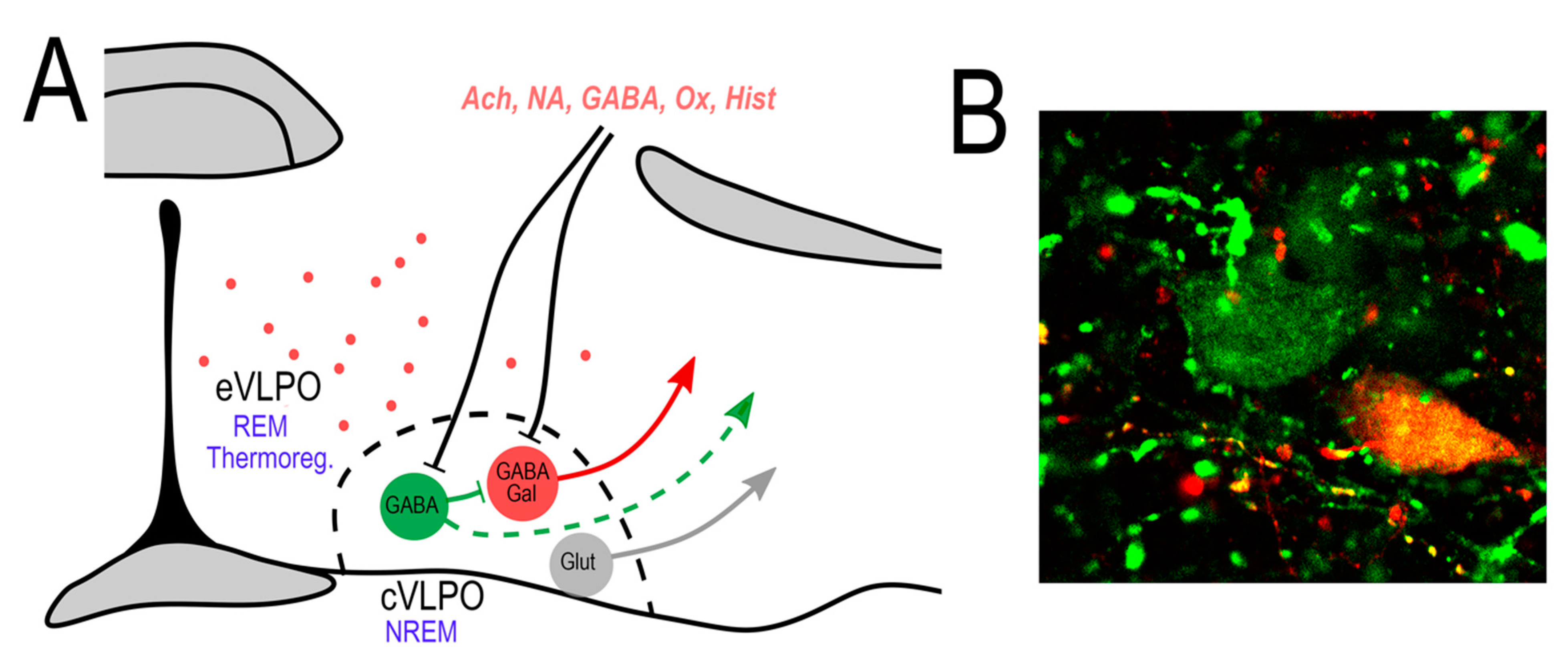
3. VLPO—Inputs and Outputs
4. Cellular Anatomy of the VLPO
4.1. The GABAergic VLPO
4.2. The Galaninergic VLPO
4.3. The Glutamatergic VLPO
5. VLPO Regulation of Behavioral State: Circuit Basis of Sleep and Arousal
Sleep Promoting Signals in VLPO—The Role of Adenosine and Prostaglandin D2
6. VLPO and Other Physiological Processes
6.1. VLPO and Thermoregulation
6.2. VLPO as Metabolic Sensor
6.3. VLPO and Anesthesia
7. On the Horizon for VLPO Research
8. Summary

{kind=link}
| The discovery of VLPO as sleep-promoting nucleus | |
| Afferent and efferent inputs and cellular composition | |
| Electrophysiology and pharmacological studies | In vitro electrophysiology studies |
| Regulation of behavioral states | |
| Other physiological processes | Thermoregulation:
|
| The VLPO nucleus | |
| Galaninergic neurons |
|
| GABAergic neurons | |
| Glutamatergic neurons |
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Von Economo, C. Die Encephalitis Lethargica; Franz Deuticke: Vienna, Austria, 1918. [Google Scholar]
- Von Economo, C. Sleep as a Problem of Localization. J. Nerv. Ment. Dis. 1930, 71, 249–259. [Google Scholar] [CrossRef]
- Hess, W.R. Le sommeil. C. R. Soc. Biol. Paris 1931, 107, 1333–1360. [Google Scholar]
- Ranson, S.W.; Ingram, W.R. Catalepsy Caused by Lesions between the Mamillary Bodies and Third Nerve in the Cat. Am. J. Physiol. 1932, 101, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Ingram, W.R.; Barris, R.W.; Ranson, S.W. Catalepsy: An Experimental Study. Arch. Neurol. Psychiatry 1936, 35, 1175–1197. [Google Scholar] [CrossRef]
- Ranson, S.W. Somnolence Caused by Hypothalamic Lesions in Monkeys. Arch. Neurol. Psychiatr. 1939, 41, 1–23. [Google Scholar] [CrossRef]
- Nauta, W.J. Hypothalamic Regulation of Sleep in Rats; an Experimental Study. J. Neurophysiol. 1946, 9, 285–316. [Google Scholar] [CrossRef]
- McGinty, D.J.; Sterman, M.B. Sleep Suppression after Basal Forebrain Lesions in the Cat. Science 1968, 160, 1253–1255. [Google Scholar] [CrossRef]
- Sherin, J.E.; Shiromani, P.J.; McCarley, R.W.; Saper, C.B. Activation of Ventrolateral Preoptic Neurons during Sleep. Science 1996, 271, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Szymusiak, R.; McGinty, D. Sleep-Waking Discharge of Basal Forebrain Projection Neurons in Cats. Brain Res. Bull. 1989, 22, 423–430. [Google Scholar] [CrossRef]
- Takahashi, K.; Lin, J.S.; Sakai, K. Characterization and Mapping of Sleep-Waking Specific Neurons in the Basal Forebrain and Preoptic Hypothalamus in Mice. Neuroscience 2009, 161, 269–292. [Google Scholar] [CrossRef] [Green Version]
- Gvilia, I.; Xu, F.; McGinty, D.; Szymusiak, R. Homeostatic Regulation of Sleep: A Role for Preoptic Area Neurons. J. Neurosci. 2006, 26, 9426–9433. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Kumar, S.; McGinty, D.; Alam, M.N.; Szymusiak, R. Neuronal Activity in the Preoptic Hypothalamus during Sleep Deprivation and Recovery Sleep. J. Neurophysiol. 2014, 111, 287–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Greco, M.A.; Shiromani, P.; Saper, C.B. Effect of Lesions of the Ventrolateral Preoptic Nucleus on NREM and REM Sleep. J. Neurosci. 2000, 20, 3830–3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigoni, E.; Lu, J.; Vetrivelan, R.; Saper, C.B. Long-Term Synaptic Plasticity Is Impaired in Rats with Lesions of the Ventrolateral Preoptic Nucleus. Eur. J. Neurosci. 2009, 30, 2112–2120. [Google Scholar] [CrossRef] [Green Version]
- Eikermann, M.; Vetrivelan, R.; Grosse-Sundrup, M.; Henry, M.E.; Hoffmann, U.; Yokota, S.; Saper, C.B.; Chamberlin, N.L. The Ventrolateral Preoptic Nucleus Is Not Required for Isoflurane General Anesthesia. Brain Res. 2011, 1426, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Vetrivelan, R.; Fuller, P.M.; Yokota, S.; Lu, J.; Saper, C.B. Metabolic Effects of Chronic Sleep Restriction in Rats. Sleep 2012, 35, 1511–1520. [Google Scholar] [CrossRef]
- Vetrivelan, R.; Kong, D.; Ferrari, L.L.; Arrigoni, E.; Madara, J.C.; Bandaru, S.S.; Lowell, B.B.; Lu, J.; Saper, C.B. Melanin-Concentrating Hormone Neurons Specifically Promote Rapid Eye Movement Sleep in Mice. Neuroscience 2016, 336, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Anaclet, C.; Lin, J.S.; Vetrivelan, R.; Krenzer, M.; Vong, L.; Fuller, P.M.; Lu, J. Identification and Characterization of a Sleep-Active Cell Group in the Rostral Medullary Brainstem. J. Neurosci. 2012, 32, 17970–17976. [Google Scholar] [CrossRef]
- Anaclet, C.; Ferrari, L.; Arrigoni, E.; Bass, C.E.; Saper, C.B.; Lu, J.; Fuller, P.M. The GABAergic Parafacial Zone Is a Medullary Slow Wave Sleep-Promoting Center. Nat. Neurosci. 2014, 17, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.A.; Kostin, A.; Siegel, J.; McGinty, D.; Szymusiak, R.; Alam, M.N. Characteristics of Sleep-Active Neurons in the Medullary Parafacial Zone in Rats. Sleep 2018, 41, zsy130. [Google Scholar] [CrossRef]
- Gong, H.; McGinty, D.; Guzman-Marin, R.; Chew, K.T.; Stewart, D.; Szymusiak, R. Activation of C-Fos in GABAergic Neurones in the Preoptic Area during Sleep and in Response to Sleep Deprivation. J. Physiol. 2004, 556, 935–946. [Google Scholar] [CrossRef] [PubMed]
- McGinty, D.; Szymusiak, R. Hypothalamic Regulation of Sleep and Arousal. Front. Biosci. 2003, 8, s1074–s1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaclet, C.; Fuller, P.M. Brainstem Regulation of Slow-Wave-Sleep. Curr. Opin. Neurobiol. 2017, 44, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M. Wake-Sleep Circuitry: An Overview. Curr. Opin. Neurobiol. 2017, 44, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Sherin, J.E.; Elmquist, J.K.; Torrealba, F.; Saper, C.B. Innervation of Histaminergic Tuberomammillary Neurons by GABAergic and Galaninergic Neurons in the Ventrolateral Preoptic Nucleus of the Rat. J. Neurosci. 1998, 18, 4705–4721. [Google Scholar] [CrossRef] [Green Version]
- Gaus, S.E.; Strecker, R.E.; Tate, B.A.; Parker, R.A.; Saper, C.B. Ventrolateral Preoptic Nucleus Contains Sleep-Active, Galaninergic Neurons in Multiple Mammalian Species. Neuroscience 2002, 115, 285–294. [Google Scholar] [CrossRef]
- Kroeger, D.; Absi, G.; Gagliardi, C.; Bandaru, S.S.; Madara, J.C.; Ferrari, L.L.; Arrigoni, E.; Munzberg, H.; Scammell, T.E.; Saper, C.B.; et al. Galanin Neurons in the Ventrolateral Preoptic Area Promote Sleep and Heat Loss in Mice. Nat. Commun. 2018, 9, 4129. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.C.; Bjorkum, A.A.; Gaus, S.E.; Lu, J.; Scammell, T.E.; Saper, C.B. Afferents to the Ventrolateral Preoptic Nucleus. J. Neurosci. 2002, 22, 977–990. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.; Weber, F.; Zhong, P.; Tan, C.L.; Nguyen, T.N.; Beier, K.T.; Hormann, N.; Chang, W.C.; Zhang, Z.; Do, J.P.; et al. Identification of Preoptic Sleep Neurons Using Retrograde Labelling and Gene Profiling. Nature 2017, 545, 477–481. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, J.R.; Bambah-Mukku, D.; Eichhorn, S.W.; Vaughn, E.; Shekhar, K.; Perez, J.D.; Rubinstein, N.D.; Hao, J.; Regev, A.; Dulac, C.; et al. Molecular, Spatial, and Functional Single-Cell Profiling of the Hypothalamic Preoptic Region. Science 2018, 362, eaau5324. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, S.; Jang, I.S.; Nabekura, J.; Akaike, N. Alpha 2-Adrenoceptor-Mediated Presynaptic Modulation of GABAergic Transmission in Mechanically Dissociated Rat Ventrolateral Preoptic Neurons. J. Neurophysiol. 2003, 89, 1640–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallopin, T.; Luppi, P.H.; Cauli, B.; Urade, Y.; Rossier, J.; Hayaishi, O.; Lambolez, B.; Fort, P. The Endogenous Somnogen Adenosine Excites a Subset of Sleep-Promoting Neurons via A2A Receptors in the Ventrolateral Preoptic Nucleus. Neuroscience 2005, 134, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Sangare, A.; Dubourget, R.; Geoffroy, H.; Gallopin, T.; Rancillac, A. Serotonin Differentially Modulates Excitatory and Inhibitory Synaptic Inputs to Putative Sleep-Promoting Neurons of the Ventrolateral Preoptic Nucleus. Neuropharmacology 2016, 109, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, F.; Claverie, D.; Lardant, E.; Varin, C.; Hardy, E.; Walter, A.; Canini, F.; Rouach, N.; Rancillac, A. Neuropeptide S Promotes Wakefulness through the Inhibition of Sleep-Promoting Ventrolateral Preoptic Nucleus Neurons. Sleep 2020, 43, zsz189. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Li, J.; Ye, J.H. Histamine Regulates Activities of Neurons in the Ventrolateral Preoptic Nucleus. J. Physiol. 2010, 588, 4103–4116. [Google Scholar] [CrossRef]
- Liang, Y.; Shi, W.; Xiang, A.; Hu, D.; Wang, L.; Zhang, L. The NAergic Locus Coeruleus-Ventrolateral Preoptic Area Neural Circuit Mediates Rapid Arousal from Sleep. Curr. Biol. 2021, 31, 3729–3742.e5. [Google Scholar] [CrossRef]
- Kask, K.; Langel, U.; Bartfai, T. Galanin—A Neuropeptide with Inhibitory Actions. Cell Mol. Neurobiol. 1995, 15, 653–673. [Google Scholar] [CrossRef]
- Pieribone, V.A.; Xu, Z.Q.; Zhang, X.; Grillner, S.; Bartfai, T.; Hokfelt, T. Galanin Induces a Hyperpolarization of Norepinephrine-Containing Locus Coeruleus Neurons in the Brainstem Slice. Neuroscience 1995, 64, 861–874. [Google Scholar] [CrossRef]
- Schonrock, B.; Busselberg, D.; Haas, H.L. Properties of Tuberomammillary Histamine Neurones and Their Response to Galanin. Agents Actions 1991, 33, 135–137. [Google Scholar] [CrossRef]
- Xu, Z.Q.; Zhang, X.; Pieribone, V.A.; Grillner, S.; Hökfelt, T. Galanin-5-Hydroxytryptamine Interactions: Electrophysiological, Immunohistochemical and in Situ Hybridization Studies on Rat Dorsal Raphe Neurons with a Note on Galanin R1 and R2 Receptors. Neuroscience 1998, 87, 79–94. [Google Scholar] [CrossRef]
- Goforth, P.B.; Leinninger, G.M.; Patterson, C.M.; Satin, L.S.; Myers, M.G. Leptin Acts via Lateral Hypothalamic Area Neurotensin Neurons to Inhibit Orexin Neurons by Multiple GABA-Independent Mechanisms. J. Neurosci. 2014, 34, 11405–11415. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.H.; Aston-Jones, G.; Akaoka, H.; Chouvet, G.; Jouvet, M. Afferent Projections to the Rat Locus Coeruleus Demonstrated by Retrograde and Anterograde Tracing with Cholera-Toxin B Subunit and Phaseolus Vulgaris Leucoagglutinin. Neuroscience 1995, 65, 119–160. [Google Scholar] [CrossRef]
- Lim, A.S.; Ellison, B.A.; Wang, J.L.; Yu, L.; Schneider, J.A.; Buchman, A.S.; Bennett, D.A.; Saper, C.B. Sleep Is Related to Neuron Numbers in the Ventrolateral Preoptic/Intermediate Nucleus in Older Adults with and without Alzheimer’s Disease. Brain 2014, 137, 2847–2861. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B. The Intermediate Nucleus in Humans: Cytoarchitecture, Chemoarchitecture, and Relation to Sleep, Sex, and Alzheimer Disease. Handb. Clin. Neurol. 2021, 179, 461–469. [Google Scholar] [CrossRef]
- Lu, J.; Bjorkum, A.A.; Xu, M.; Gaus, S.E.; Shiromani, P.J.; Saper, C.B. Selective Activation of the Extended Ventrolateral Preoptic Nucleus during Rapid Eye Movement Sleep. J. Neurosci. 2002, 22, 4568–4576. [Google Scholar] [CrossRef]
- Koyama, Y.; Hayaishi, O. Firing of Neurons in the Preoptic/Anterior Hypothalamic Areas in Rat: Its Possible Involvement in Slow Wave Sleep and Paradoxical Sleep. Neurosci. Res. 1994, 19, 31–38. [Google Scholar] [CrossRef]
- Osaka, T.; Matsumura, H. Noradrenaline Inhibits Preoptic Sleep-Active Neurons through Alpha 2-Receptors in the Rat. Neurosci. Res. 1995, 21, 323–330. [Google Scholar] [CrossRef]
- Szymusiak, R.; Alam, N.; Steininger, T.L.; McGinty, D. Sleep-Waking Discharge Patterns of Ventrolateral Preoptic/Anterior Hypothalamic Neurons in Rats. Brain Res. 1998, 803, 178–188. [Google Scholar] [CrossRef]
- Lu, J.; Sherman, D.; Devor, M.; Saper, C.B. A Putative Flip-Flop Switch for Control of REM Sleep. Nature 2006, 441, 589–594. [Google Scholar] [CrossRef]
- Hsieh, K.-C.; Gvilia, I.; Kumar, S.; Uschakov, A.; McGinty, D.; Alam, M.N.; Szymusiak, R. C-Fos Expression in Neurons Projecting from the Preoptic and Lateral Hypothalamic Areas to the Ventrolateral Periaqueductal Gray in Relation to Sleep States. Neuroscience 2011, 188, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Vanini, G.; Bassana, M.; Mast, M.; Mondino, A.; Cerda, I.; Phyle, M.; Chen, V.; Colmenero, A.V.; Hambrecht-Wiedbusch, V.S.; Mashour, G.A. Activation of Preoptic GABAergic or Glutamatergic Neurons Modulates Sleep-Wake Architecture, but Not Anesthetic State Transitions. Curr. Biol. 2020, 30, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaclet, C.; Pedersen, N.P.; Ferrari, L.L.; Venner, A.; Bass, C.E.; Arrigoni, E.; Fuller, P.M. Basal Forebrain Control of Wakefulness and Cortical Rhythms. Nat. Commun. 2015, 6, 8744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venner, A.; Anaclet, C.; Broadhurst, R.Y.; Saper, C.B.; Fuller, P.M. A Novel Population of Wake-Promoting GABAergic Neurons in the Ventral Lateral Hypothalamus. Curr. Biol. 2016, 26, 2137–2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, N.P.; Ferrari, L.; Venner, A.; Wang, J.L.; Abbott, S.B.G.; Vujovic, N.; Arrigoni, E.; Saper, C.B.; Fuller, P.M. Supramammillary Glutamate Neurons Are a Key Node of the Arousal System. Nat. Commun. 2017, 8, 1405. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, D.-R.; Dong, H.; Chen, L.; Lu, J.; Lazarus, M.; Cherasse, Y.; Chen, G.-H.; Qu, W.-M.; Huang, Z.-L. Medial Parabrachial Nucleus Is Essential in Controlling Wakefulness in Rats. Front. Neurosci. 2021, 15, 645877. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Fuller, P.M.; Pedersen, N.P.; Lu, J.; Scammell, T.E. Sleep State Switching. Neuron 2010, 68, 1023–1042. [Google Scholar] [CrossRef] [Green Version]
- Scammell, T.E.; Arrigoni, E.; Lipton, J.O. Neural Circuitry of Wakefulness and Sleep. Neuron 2017, 93, 747–765. [Google Scholar] [CrossRef] [Green Version]
- Gallopin, T.; Fort, P.; Eggermann, E.; Cauli, B.; Luppi, P.H.; Rossier, J.; Audinat, E.; Muhlethaler, M.; Serafin, M. Identification of Sleep-Promoting Neurons in Vitro. Nature 2000, 404, 992–995. [Google Scholar] [CrossRef]
- Gallopin, T.; Luppi, P.H.; Rambert, F.A.; Frydman, A.; Fort, P. Effect of the Wake-Promoting Agent Modafinil on Sleep-Promoting Neurons from the Ventrolateral Preoptic Nucleus: An in Vitro Pharmacologic Study. Sleep 2004, 27, 19–25. [Google Scholar]
- Williams, R.H.; Chee, M.J.; Kroeger, D.; Ferrari, L.L.; Maratos-Flier, E.; Scammell, T.E.; Arrigoni, E. Optogenetic-Mediated Release of Histamine Reveals Distal and Autoregulatory Mechanisms for Controlling Arousal. J. Neurosci. 2014, 34, 6023–6029. [Google Scholar] [CrossRef]
- Venner, A.; De Luca, R.; Sohn, L.T.; Bandaru, S.S.; Verstegen, A.M.J.; Arrigoni, E.; Fuller, P.M. An Inhibitory Lateral Hypothalamic-Preoptic Circuit Mediates Rapid Arousals from Sleep. Curr. Biol. 2019, 29, 4155–4168. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.T.; Chen, J.; Han, B.; Meng, Q.C.; Veasey, S.C.; Beck, S.G.; Kelz, M.B. Direct Activation of Sleep-Promoting VLPO Neurons by Volatile Anesthetics Contributes to Anesthetic Hypnosis. Curr. Biol. 2012, 22, 2008–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osaka, T.; Matsumura, H. Noradrenergic Inputs to Sleep-Related Neurons in the Preoptic Area from the Locus Coeruleus and the Ventrolateral Medulla in the Rat. Neurosci. Res. 1994, 19, 39–50. [Google Scholar] [CrossRef]
- Saint-Mleux, B.; Eggermann, E.; Bisetti, A.; Bayer, L.; Machard, D.; Jones, B.E.; Muhlethaler, M.; Serafin, M. Nicotinic Enhancement of the Noradrenergic Inhibition of Sleep-Promoting Neurons in the Ventrolateral Preoptic Area. J. Neurosci. 2004, 24, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methippara, M.M.; Alam, M.N.; Szymusiak, R.; McGinty, D. Effects of Lateral Preoptic Area Application of Orexin-A on Sleep-Wakefulness. Neuroreport 2000, 11, 3423–3426. [Google Scholar] [CrossRef]
- Mavanji, V.; Perez-Leighton, C.E.; Kotz, C.M.; Billington, C.J.; Parthasarathy, S.; Sinton, C.M.; Teske, J.A. Promotion of Wakefulness and Energy Expenditure by Orexin-A in the Ventrolateral Preoptic Area. Sleep 2015, 38, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- De Luca, R.; Park, D.; Bandaru, S.S. Orexin Mediates Feed-Forward Inhibition of VLPO Sleep-Active Neurons—A Mechanism for Controlling Arousal. Sleep 2017, 40, A50. [Google Scholar] [CrossRef] [Green Version]
- McCarren, H.S.; Chalifoux, M.R.; Han, B.; Moore, J.T.; Meng, Q.C.; Baron-Hionis, N.; Sedigh-Sarvestani, M.; Contreras, D.; Beck, S.G.; Kelz, M.B. A2-Adrenergic Stimulation of the Ventrolateral Preoptic Nucleus Destabilizes the Anesthetic State. J. Neurosci. 2014, 34, 16385–16396. [Google Scholar] [CrossRef]
- Huang, Z.L.; Urade, Y.; Hayaishi, O. Prostaglandins and Adenosine in the Regulation of Sleep and Wakefulness. Curr. Opin. Pharmacol. 2007, 7, 33–38. [Google Scholar] [CrossRef]
- Urade, Y.; Hayaishi, O. Prostaglandin D2 and Sleep/Wake Regulation. Sleep Med. Rev. 2011, 15, 411–418. [Google Scholar] [CrossRef]
- Lazarus, M.; Chen, J.-F.; Huang, Z.-L.; Urade, Y.; Fredholm, B.B. Adenosine and Sleep. Handb. Exp. Pharmacol. 2019, 253, 359–381. [Google Scholar] [CrossRef] [PubMed]
- Porkka-Heiskanen, T.; Strecker, R.E.; Thakkar, M.; Bjorkum, A.A.; Greene, R.W.; McCarley, R.W. Adenosine: A Mediator of the Sleep-Inducing Effects of Prolonged Wakefulness. Science 1997, 276, 1265–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ram, A.; Pandey, H.P.; Matsumura, H.; Kasahara-Orita, K.; Nakajima, T.; Takahata, R.; Satoh, S.; Terao, A.; Hayaishi, O. CSF Levels of Prostaglandins, Especially the Level of Prostaglandin D2, Are Correlated with Increasing Propensity towards Sleep in Rats. Brain Res. 1997, 751, 81–89. [Google Scholar] [CrossRef]
- Pentreath, V.W.; Rees, K.; Owolabi, O.A.; Philip, K.A.; Doua, F. The Somnogenic T Lymphocyte Suppressor Prostaglandin D2 Is Selectively Elevated in Cerebrospinal Fluid of Advanced Sleeping Sickness Patients. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 795–799. [Google Scholar] [CrossRef]
- Jordan, W.; Tumani, H.; Cohrs, S.; Eggert, S.; Rodenbeck, A.; Brunner, E.; Rüther, E.; Hajak, G. Prostaglandin D Synthase (Beta-Trace) in Healthy Human Sleep. Sleep 2004, 27, 867–874. [Google Scholar] [CrossRef]
- Porkka-Heiskanen, T.; Kalinchuk, A.V. Adenosine, Energy Metabolism and Sleep Homeostasis. Sleep Med. Rev. 2011, 15, 123–135. [Google Scholar] [CrossRef]
- Scammell, T.E.; Gerashchenko, D.Y.; Mochizuki, T.; McCarthy, M.T.; Estabrooke, I.V.; Sears, C.A.; Saper, C.B.; Urade, Y.; Hayaishi, O. An Adenosine A2a Agonist Increases Sleep and Induces Fos in Ventrolateral Preoptic Neurons. Neuroscience 2001, 107, 653–663. [Google Scholar] [CrossRef]
- Chamberlin, N.L.; Arrigoni, E.; Chou, T.C.; Scammell, T.E.; Greene, R.W.; Saper, C.B. Effects of Adenosine on Gabaergic Synaptic Inputs to Identified Ventrolateral Preoptic Neurons. Neuroscience 2003, 119, 913–918. [Google Scholar] [CrossRef]
- Morairty, S.; Rainnie, D.; McCarley, R.; Greene, R. Disinhibition of Ventrolateral Preoptic Area Sleep-Active Neurons by Adenosine: A New Mechanism for Sleep Promotion. Neuroscience 2004, 123, 451–457. [Google Scholar] [CrossRef]
- Narumiya, S.; Ogorochi, T.; Nakao, K.; Hayaishi, O. Prostaglandin D2 in Rat Brain, Spinal Cord and Pituitary: Basal Level and Regional Distribution. Life Sci. 1982, 31, 2093–2103. [Google Scholar] [CrossRef]
- Ogorochi, T.; Narumiya, S.; Mizuno, N.; Yamashita, K.; Miyazaki, H.; Hayaishi, O. Regional Distribution of Prostaglandins D2, E2, and F2 Alpha and Related Enzymes in Postmortem Human Brain. J. Neurochem. 1984, 43, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Ueno, R.; Ishikawa, Y.; Nakayama, T.; Hayaishi, O. Prostaglandin D2 Induces Sleep When Microinjected into the Preoptic Area of Conscious Rats. Biochem. Biophys. Res. Commun. 1982, 109, 576–582. [Google Scholar] [CrossRef]
- Matsumura, H.; Nakajima, T.; Osaka, T.; Satoh, S.; Kawase, K.; Kubo, E.; Kantha, S.S.; Kasahara, K.; Hayaishi, O. Prostaglandin D2-Sensitive, Sleep-Promoting Zone Defined in the Ventral Surface of the Rostral Basal Forebrain. Proc. Natl. Acad. Sci. USA 1994, 91, 11998–12002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, S.; Matsumura, H.; Suzuki, F.; Hayaishi, O. Promotion of Sleep Mediated by the A2a-Adenosine Receptor and Possible Involvement of This Receptor in the Sleep Induced by Prostaglandin D2 in Rats. Proc. Natl. Acad. Sci. USA 1996, 93, 5980–5984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, A.; Eguchi, N.; Kimura, K.; Kiyohara, Y.; Qu, W.M.; Huang, Z.L.; Mochizuki, T.; Lazarus, M.; Kobayashi, T.; Kaneko, T.; et al. Dominant Localization of Prostaglandin D Receptors on Arachnoid Trabecular Cells in Mouse Basal Forebrain and Their Involvement in the Regulation of Non-Rapid Eye Movement Sleep. Proc. Natl. Acad. Sci. USA 2001, 98, 11674–11679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onoe, H.; Ueno, R.; Fujita, I.; Nishino, H.; Oomura, Y.; Hayaishi, O. Prostaglandin D2, a Cerebral Sleep-Inducing Substance in Monkeys. Proc. Natl. Acad. Sci. USA 1988, 85, 4082–4086. [Google Scholar] [CrossRef] [Green Version]
- Pandey, H.P.; Ram, A.; Matsumura, H.; Hayaishi, O. Concentration of Prostaglandin D2 in Cerebrospinal Fluid Exhibits a Circadian Alteration in Conscious Rats. Biochem. Mol. Biol. Int. 1995, 37, 431–437. [Google Scholar]
- Beuckmann, C.T.; Lazarus, M.; Gerashchenko, D.; Mizoguchi, A.; Nomura, S.; Mohri, I.; Uesugi, A.; Kaneko, T.; Mizuno, N.; Hayaishi, O.; et al. Cellular Localization of Lipocalin-Type Prostaglandin D Synthase (Beta-Trace) in the Central Nervous System of the Adult Rat. J. Comp. Neurol. 2000, 428, 62–78. [Google Scholar] [CrossRef]
- Hayaishi, O.; Urade, Y.; Eguchi, N.; Huang, Z.L. Genes for Prostaglandin d Synthase and Receptor as Well as Adenosine A2A Receptor Are Involved in the Homeostatic Regulation of Nrem Sleep. Arch. Ital. Biol. 2004, 142, 533–539. [Google Scholar]
- Scammell, T.; Gerashchenko, D.; Urade, Y.; Onoe, H.; Saper, C.; Hayaishi, O. Activation of Ventrolateral Preoptic Neurons by the Somnogen Prostaglandin D2. Proc. Natl. Acad. Sci. USA 1998, 95, 7754–7759. [Google Scholar] [CrossRef] [Green Version]
- Gerashchenko, D.; Beuckmann, C.T.; Kanaoka, Y.; Eguchi, N.; Gordon, W.C.; Urade, Y.; Bazan, N.G.; Hayaishi, O. Dominant Expression of Rat Prostanoid DP Receptor MRNA in Leptomeninges, Inner Segments of Photoreceptor Cells, Iris Epithelium, and Ciliary Processes. J. Neurochem. 1998, 71, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alföldi, P.; Rubicsek, G.; Cserni, G.; Obál, F. Brain and Core Temperatures and Peripheral Vasomotion during Sleep and Wakefulness at Various Ambient Temperatures in the Rat. Pflugers Arch. 1990, 417, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Landolt, H.P.; Moser, S.; Wieser, H.G.; Borbély, A.A.; Dijk, D.J. Intracranial Temperature across 24-Hour Sleep-Wake Cycles in Humans. Neuroreport 1995, 6, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Klerman, E.B.; Sakurai, T.; Scammell, T.E. Elevated Body Temperature during Sleep in Orexin Knockout Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R533–R540. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.N.; McGinty, D.; Szymusiak, R. Neuronal Discharge of Preoptic/Anterior Hypothalamic Thermosensitive Neurons: Relation to NREM Sleep. Am. J. Physiol. 1995, 269, R1240–R1249. [Google Scholar] [CrossRef]
- Gong, H.; Szymusiak, R.; King, J.; Steininger, T.; McGinty, D. Sleep-Related c-Fos Protein Expression in the Preoptic Hypothalamus: Effects of Ambient Warming. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R2079–R2088. [Google Scholar] [CrossRef]
- Nakamura, K. Central Circuitries for Body Temperature Regulation and Fever. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1207–R1228. [Google Scholar] [CrossRef] [Green Version]
- Saper, C.B.; Romanovsky, A.A.; Scammell, T.E. Neural Circuitry Engaged by Prostaglandins during the Sickness Syndrome. Nat. Neurosci. 2012, 15, 1088–1095. [Google Scholar] [CrossRef]
- Harding, E.C.; Franks, N.P.; Wisden, W. The Temperature Dependence of Sleep. Front. Neurosci. 2019, 13, 336. [Google Scholar] [CrossRef]
- Morrison, S.F.; Nakamura, K. Central Neural Pathways for Thermoregulation. Front. Biosci. 2011, 16, 74–104. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.F. Central Neural Control of Thermoregulation and Brown Adipose Tissue. Auton. Neurosci. 2016, 196, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.-D.; Yang, W.Z.; Gao, C.; Fu, X.; Zhang, W.; Zhou, Q.; Chen, W.; Ni, X.; Lin, J.-K.; Yang, J.; et al. A Hypothalamic Circuit That Controls Body Temperature. Proc. Natl. Acad. Sci. USA 2017, 114, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; McKinley, M.J.; McAllen, R.M. Roles of Two Preoptic Cell Groups in Tonic and Febrile Control of Rat Tail Sympathetic Fibers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1248–R1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; McKinley, M.J.; McAllen, R.M. Preoptic-Raphé Connections for Thermoregulatory Vasomotor Control. J. Neurosci. 2011, 31, 5078–5088. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Li, X.; Cano, G.; Lazarus, M.; Saper, C.B. Parallel Preoptic Pathways for Thermoregulation. J. Neurosci. 2009, 29, 11954–11964. [Google Scholar] [CrossRef]
- Machado, N.L.S.; Bandaru, S.S.; Abbott, S.B.G.; Saper, C.B. EP3R-Expressing Glutamatergic Preoptic Neurons Mediate Inflammatory Fever. J. Neurosci. 2020, 40, 2573–2588. [Google Scholar] [CrossRef]
- Varin, C.; Rancillac, A.; Geoffroy, H.; Arthaud, S.; Fort, P.; Gallopin, T. Glucose Induces Slow-Wave Sleep by Exciting the Sleep-Promoting Neurons in the Ventrolateral Preoptic Nucleus: A New Link between Sleep and Metabolism. J. Neurosci. 2015, 35, 9900–9911. [Google Scholar] [CrossRef] [Green Version]
- Koster, J.C.; Marshall, B.A.; Ensor, N.; Corbett, J.A.; Nichols, C.G. Targeted Overactivity of Beta Cell K(ATP) Channels Induces Profound Neonatal Diabetes. Cell 2000, 100, 645–654. [Google Scholar] [CrossRef] [Green Version]
- Parton, L.E.; Ye, C.P.; Coppari, R.; Enriori, P.J.; Choi, B.; Zhang, C.-Y.; Xu, C.; Vianna, C.R.; Balthasar, N.; Lee, C.E.; et al. Glucose Sensing by POMC Neurons Regulates Glucose Homeostasis and Is Impaired in Obesity. Nature 2007, 449, 228–232. [Google Scholar] [CrossRef]
- Yoon, N.A.; Diano, S. Hypothalamic Glucose-Sensing Mechanisms. Diabetologia 2021, 64, 985–993. [Google Scholar] [CrossRef]
- Burdakov, D.; Jensen, L.T.; Alexopoulos, H.; Williams, R.H.; Fearon, I.M.; O’Kelly, I.; Gerasimenko, O.; Fugger, L.; Verkhratsky, A. Tandem-Pore K+ Channels Mediate Inhibition of Orexin Neurons by Glucose. Neuron 2006, 50, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Danguir, J.; Nicolaidis, S. Dependence of Sleep on Nutrients’ Availability. Physiol. Behav. 1979, 22, 735–740. [Google Scholar] [CrossRef]
- Yamanaka, A.; Beuckmann, C.T.; Willie, J.T.; Hara, J.; Tsujino, N.; Mieda, M.; Tominaga, M.; Yagami, K.; Sugiyama, F.; Goto, K.; et al. Hypothalamic Orexin Neurons Regulate Arousal According to Energy Balance in Mice. Neuron 2003, 38, 701–713. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, J.B.; Omori, T.; Guan, Z.; Vgontzas, A.N.; Bixler, E.O.; Fang, J. Sleep Is Increased in Mice with Obesity Induced by High-Fat Food. Physiol. Behav. 2006, 87, 255–262. [Google Scholar] [CrossRef]
- Northeast, R.C.; Vyazovskiy, V.V.; Bechtold, D.A. Eat, Sleep, Repeat: The Role of the Circadian System in Balancing Sleep-Wake Control with Metabolic Need. Curr. Opin. Physiol. 2020, 15, 183–191. [Google Scholar] [CrossRef]
- Lydic, R.; Biebuyck, J.F. Sleep Neurobiology: Relevance for Mechanistic Studies of Anaesthesia. Br. J. Anaesth. 1994, 72, 506–508. [Google Scholar] [CrossRef]
- Mashour, G.A.; Hudetz, A.G. Bottom-Up and Top-Down Mechanisms of General Anesthetics Modulate Different Dimensions of Consciousness. Front. Neural. Circuits 2017, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.E.; Guo, T.Z.; Lu, J.; Saper, C.B.; Franks, N.P.; Maze, M. The Sedative Component of Anesthesia Is Mediated by GABA(A) Receptors in an Endogenous Sleep Pathway. Nat. Neurosci. 2002, 5, 979–984. [Google Scholar] [CrossRef]
- Nelson, L.E.; Lu, J.; Guo, T.; Saper, C.B.; Franks, N.P.; Maze, M. The Alpha2-Adrenoceptor Agonist Dexmedetomidine Converges on an Endogenous Sleep-Promoting Pathway to Exert Its Sedative Effects. Anesthesiology 2003, 98, 428–436. [Google Scholar] [CrossRef] [Green Version]
- Li, K.Y.; Guan, Y.; Krnjević, K.; Ye, J.H. Propofol Facilitates Glutamatergic Transmission to Neurons of the Ventrolateral Preoptic Nucleus. Anesthesiology 2009, 111, 1271–1278. [Google Scholar] [CrossRef] [Green Version]
- Tung, A.; Szafran, M.J.; Bluhm, B.; Mendelson, W.B. Sleep Deprivation Potentiates the Onset and Duration of Loss of Righting Reflex Induced by Propofol and Isoflurane. Anesthesiology 2002, 97, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Saper, C.B.; Chou, T.C.; Scammell, T.E. The Sleep Switch: Hypothalamic Control of Sleep and Wakefulness. Trends Neurosci. 2001, 24, 726–731. [Google Scholar] [CrossRef]
- Szymusiak, R.; Gvilia, I.; McGinty, D. Hypothalamic Control of Sleep. Sleep Med. 2007, 8, 291–301. [Google Scholar] [CrossRef]
- Steininger, T.L.; Gong, H.; McGinty, D.; Szymusiak, R. Subregional Organization of Preoptic Area/Anterior Hypothalamic Projections to Arousal-Related Monoaminergic Cell Groups. J. Comp. Neurol. 2001, 429, 638–653. [Google Scholar] [CrossRef]
- Scharbarg, E.; Daenens, M.; Lemaitre, F.; Geoffroy, H.; Guille-Collignon, M.; Gallopin, T.; Rancillac, A. Astrocyte-Derived Adenosine Is Central to the Hypnogenic Effect of Glucose. Sci. Rep. 2016, 6, 19107. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, F.; Gómez-Extremera, M.; Bernaola-Galván, P.; Vetrivelan, R.; Saper, C.B.; Scammell, T.E.; Ivanov, P.C. Critical Dynamics and Coupling in Bursts of Cortical Rhythms Indicate Non-Homeostatic Mechanism for Sleep-Stage Transitions and Dual Role of VLPO Neurons in Both Sleep and Wake. J. Neurosci. 2020, 40, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Vetrivelan, R.; Saper, C.B.; Fuller, P.M. Armodafinil-Induced Wakefulness in Animals with Ventrolateral Preoptic Lesions. Nat. Sci. Sleep 2014, 6, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.L.; Cooke, E.K.; Leib, D.E.; Lin, Y.-C.; Daly, G.E.; Zimmerman, C.A.; Knight, Z.A. Warm-Sensitive Neurons That Control Body Temperature. Cell 2016, 167, 47–59.e15. [Google Scholar] [CrossRef] [Green Version]
- Szymusiak, R. Body Temperature and Sleep. Handb. Clin. Neurol. 2018, 156, 341–351. [Google Scholar] [CrossRef]
- Rothhaas, R.; Chung, S. Role of the Preoptic Area in Sleep and Thermoregulation. Front. Neurosci. 2021, 15, 664781. [Google Scholar] [CrossRef]
- Harding, E.C.; Franks, N.P.; Wisden, W. Sleep and Thermoregulation. Curr. Opin. Physiol. 2020, 15, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.L.; Knight, Z.A. Regulation of Body Temperature by the Nervous System. Neuron 2018, 98, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Luo, Z.; Zhang, Y.; Zhang, Y.; Wang, Y.; Cao, S.; Fu, B.; Yang, H.; Zhang, L.; Zhou, W.; et al. GABAergic Ventrolateral Pre-optic Nucleus Neurons Are Involved in the Mediation of the Anesthetic Hypnosis Induced by Propofol. Mol. Med. Rep. 2017, 16, 3179–3186. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arrigoni, E.; Fuller, P.M. The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? Int. J. Mol. Sci. 2022, 23, 2905. https://doi.org/10.3390/ijms23062905
Arrigoni E, Fuller PM. The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? International Journal of Molecular Sciences. 2022; 23(6):2905. https://doi.org/10.3390/ijms23062905
Chicago/Turabian StyleArrigoni, Elda, and Patrick M. Fuller. 2022. "The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years?" International Journal of Molecular Sciences 23, no. 6: 2905. https://doi.org/10.3390/ijms23062905
APA StyleArrigoni, E., & Fuller, P. M. (2022). The Sleep-Promoting Ventrolateral Preoptic Nucleus: What Have We Learned over the Past 25 Years? International Journal of Molecular Sciences, 23(6), 2905. https://doi.org/10.3390/ijms23062905