
Beneficial Effects of Dietary Nitrite on a Model of Nonalcoholic Steatohepatitis Induced by High-Fat/High-Cholesterol Diets in SHRSP5/Dmcr Rats: A Preliminary Study



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Nitrite and Captopril on the Body, Liver, and Heart Weights and Blood Pressure of the Rat NASH Model
2.2. Effects of Nitrite and Captopril on Liver Steatosis in the Rat NASH Model
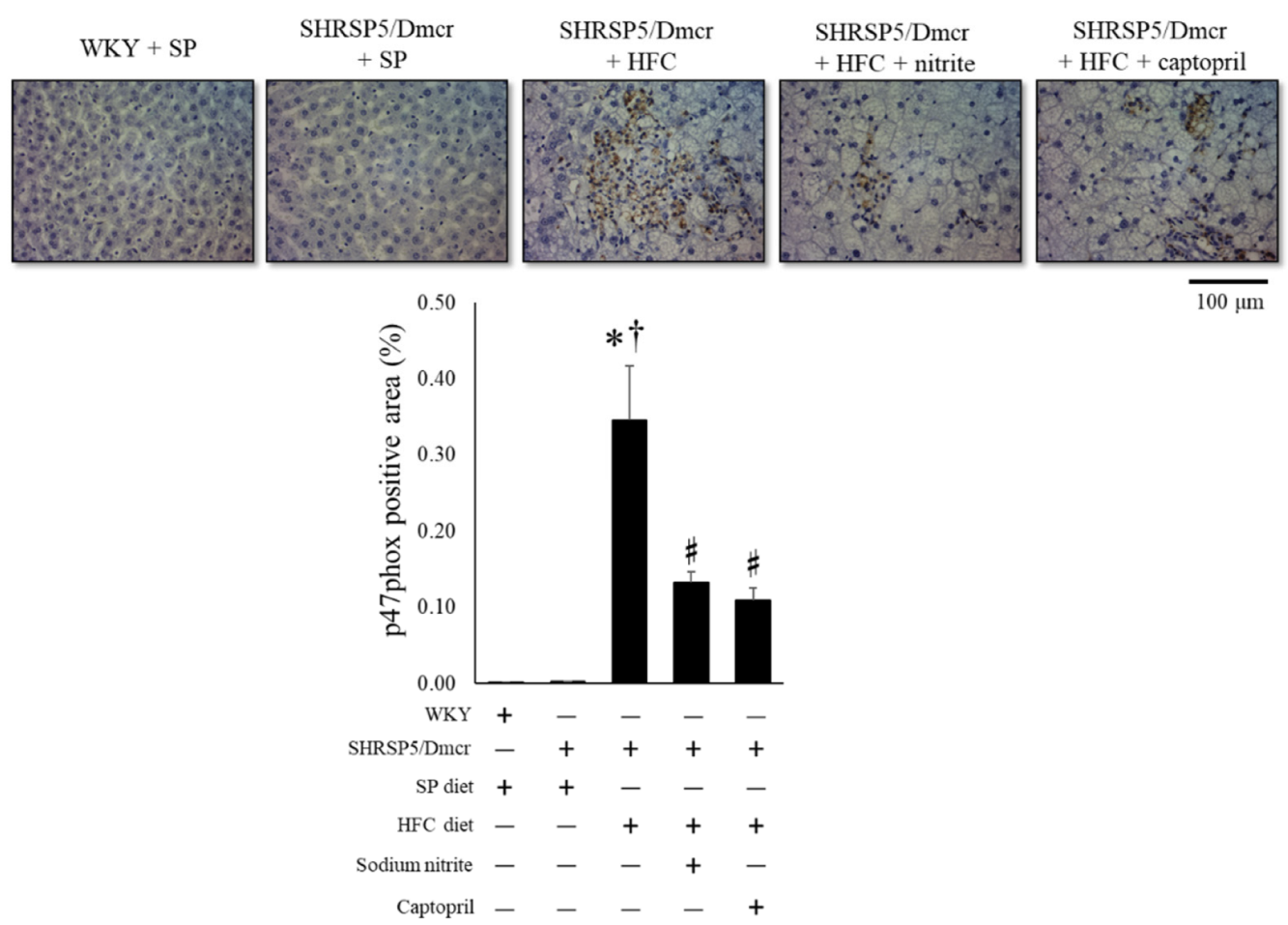
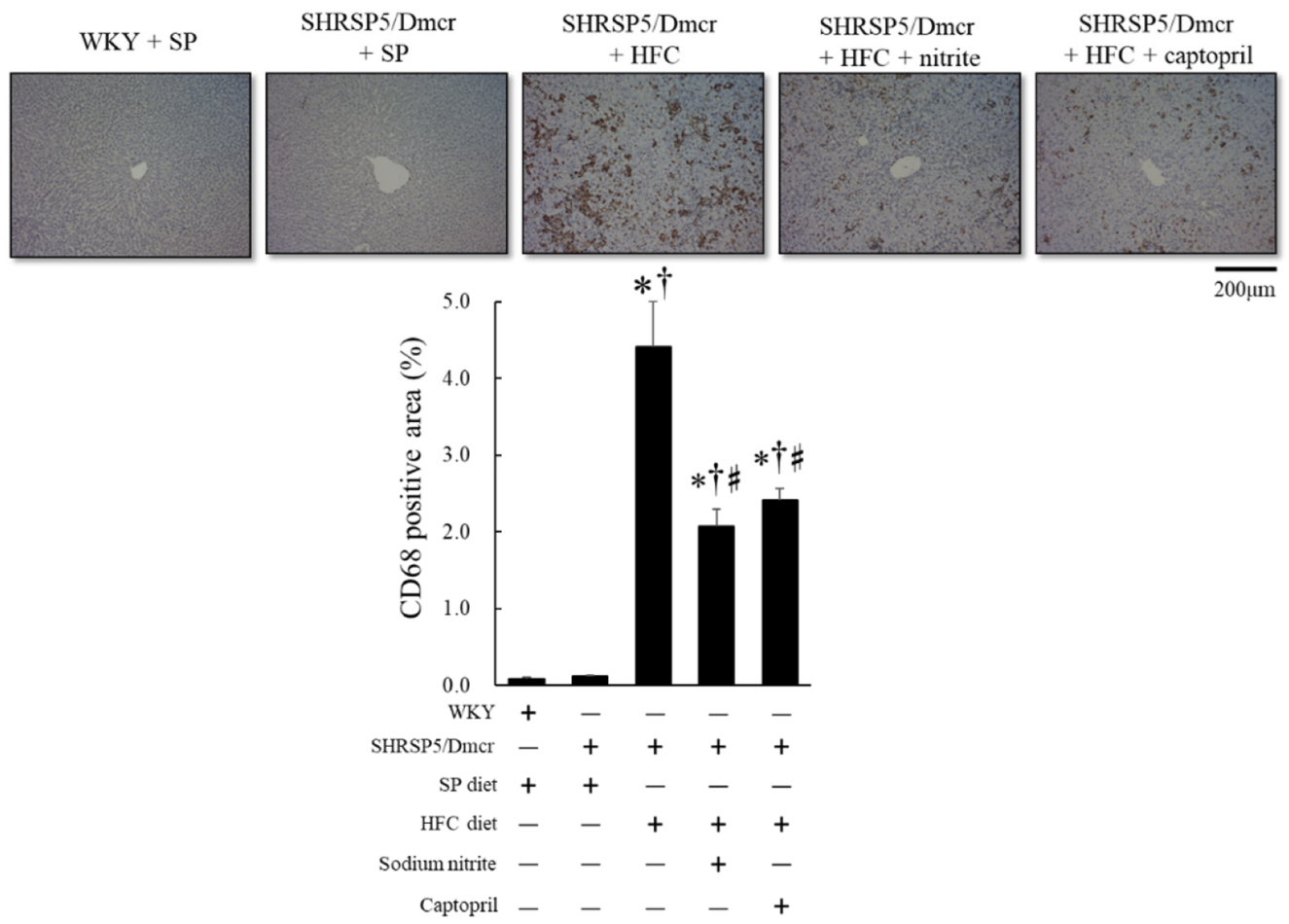
2.3. Effects of Nitrite and Captopril on Liver Inflammation and Fibrosis in the Rat NASH Model
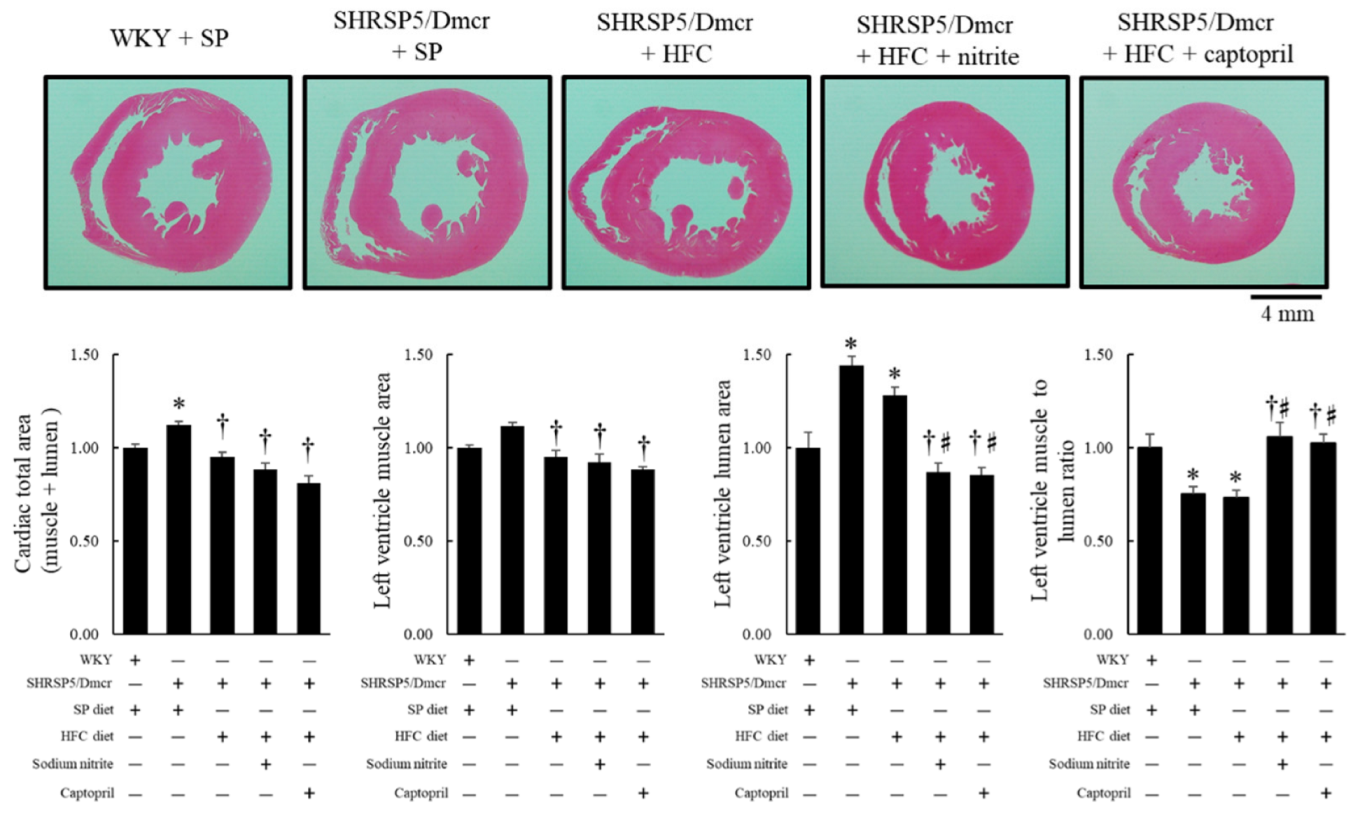
2.4. Effects of Nitrite and Captopril Supplementation on Dilated Left Ventricle (LV) and Coronary Perivascular Fibrosis in the NASH Model
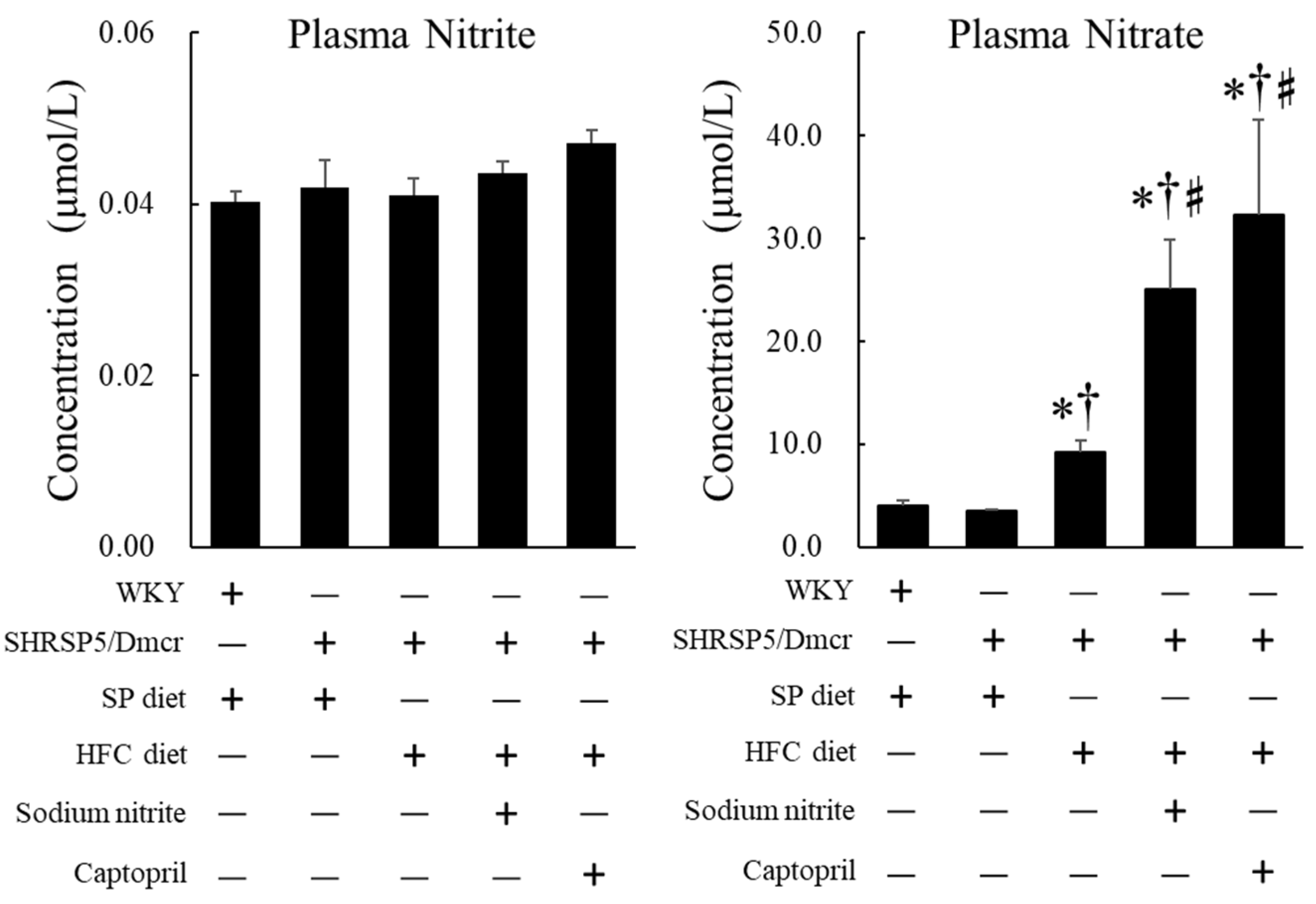
2.5. Effects of Nitrite and Captopril Supplementation on the Plasma Levels of Nitrite and Nitrate in the NASH Model
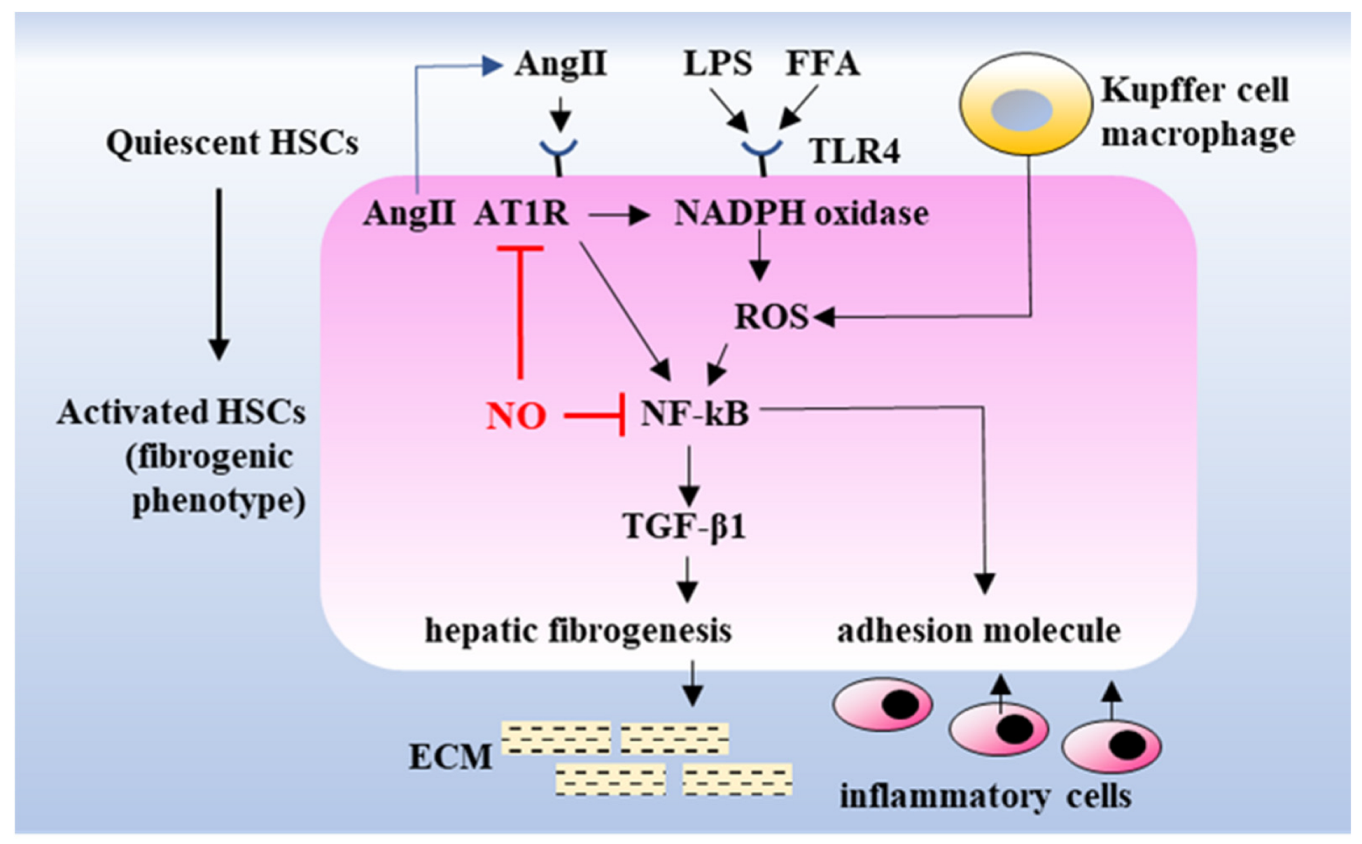
3. Discussion
4. Materials and Methods
4.1. Drug
4.2. Animals
4.3. Arterial Blood Pressure
4.4. Histological Analysis
4.5. Immunohistochemistry Analysis
4.6. RT-PCR
4.7. Nitrite and Nitrate Concentration Measurement
4.8. Plasma AST and ALT Concentration Measurement
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hashimoto, E.; Taniai, M.; Tokushige, K. Characteristics and diagnosis of NAFLD/NASH. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. S4), 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Kumazaki, S.; Yamamoto, S.; Sato, I.; Kitamori, K.; Mori, M.; Yamori, Y.; Hirohata, S. Non-alcoholic steatohepatitis aggravates nitric oxide synthase inhibition-induced arteriosclerosis in SHRSP 5/Dmcr rat model. Int. J. Exp. Pathol. 2018, 99, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Herrera, I.; Kozyra, M.; Zhuge, Z.; Haworth, S.M.; Moretti, C.; Peleli, M.; Caldeira-Dias, M.; Jahandideh, A.; Huirong, H.; de Canpos Cruz, J.; et al. AMP-activated protein kinase activation and NADPH oxidase inhibition by inorganic nitrate and nitrite prevent liver steatosis. Proc. Natl Acad. Sci. USA 2019, 116, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Croft, K.D.; Caparros-Martin, J.; O’Gara, F.; Mori, T.A.; Ward, N.C. Beneficial effects of inorganic nitrate in non-alcoholic fatty liver disease. Arch. Biochem. Biophys. 2021, 711, 109032. [Google Scholar] [CrossRef]
- Michelotti, G.A.; Machado, M.V.; Diehl, A.M. NAFLD, NASH and liver cancer. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 656–665. [Google Scholar] [CrossRef]
- Kitamori, K.; Naito, H.; Tamada, H.; Kobayashi, M.; Miyazawa, D.; Yasui, Y.; Sonoda, K.; Tsuchikura, S.; Yasui, N.; Ikeda, K.; et al. Development of novel rat model for high-fat and high-cholesterol diet-induced steatohepatitis and severe fibrosis progression in SHRSP5/Dmcr. Environ. Health Prev. Med. 2012, 17, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Horai, Y.; Utsumi, H.; Ono, Y.; Kishimoto, T.; Ono, Y.; Fukunari, A. Pathological characterization and morphometric analysis of hepatic lesions in SHRSP5/Dmcr, an experimental non-alcoholic steatohepatitis model, induced by high-fat and high-cholesterol diet. Int. J. Exp. Pathol. 2016, 97, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Kumazaki, S.; Kusunoki, K.; Inoue, T.; Maeda, Y.; Usui, S.; Shinohata, R.; Ohtsuki, T.; Hirohata, S.; Kusachi, S.; et al. A high-fat and high-cholesterol diet induces cardiac fibrosis, vascular endothelial, and left ventricular diastolic dysfunction in SHRSP5/Dmcr rats. J. Atheroscler. Thromb. 2018, 25, 439–453. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wong, G.L.H.; Yip, T.C.F.; Tse, Y.K.; Liang, L.Y.; Hui, V.W.K.; Lin, H.; Li, G.L.; Lai, J.C.T.; Chan, H.L.Y.; et al. Angiotensin-converting enzyme inhibitors prevent liver-related events in nonalcoholic fatty liver disease. Hepatology 2021, 1–14. [Google Scholar] [CrossRef]
- Toblli, J.E.; Muñoz, M.C.; Cao, G.; Mella, J.; Pereyra, L.; Mastai, R. ACE inhibition and AT1 receptor blockade prevent fatty liver and fibrosis in obese zucker rats. Obesity 2008, 16, 770–776. [Google Scholar] [CrossRef]
- Kobayashi, J.; Ohtake, K.; Uchida, H. NO-rich diet for lifestyle-related diseases. Nutrients 2015, 7, 4911–4937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapil, V.; Khambata, R.S.; Jones, D.A.; Rathod, K.; Primus, C.; Massimo, G.; Fukuto, J.M.; Ahluwalia, A. The noncanonical pathway for in vivo nitric oxide generation: The nitrate-nitrite-nitric oxide pathway. Pharmacol. Rev. 2020, 72, 692–766. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J. Lifestyle-mediated nitric oxide boost to prevent SARS-CoV-2 infection: A perspective. Nitric Oxide 2021, 115, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Akiba, J.; Matsui, T.; Nakamura, K.; Hisamoto, T.; Abe, M.; Ikezono, Y.; Wada, F.; Iwamoto, H.; Nakamura, T.; et al. Pigment epithelium-derived factor (PEDF) prevents hepatic fat storage, inflammation, and fibrosis in dietary steatohepatitis of mice. Dig. Dis. Sci. 2017, 62, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Epshteyn, V.; Morrison, K.; Krishnaswamy, P.; Kazanegra, R.; Clopton, P.; Mudaliar, S.; Edelman, S.; Henry, R.; Maisel, A. Utility of B-type natriuretic peptide (BNP) as a screen for left ventricular dysfunction in patients with diabetes. Diabetes Care 2003, 26, 2081–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, K.C.; Bernier, S.G.; Jacobson, S.; Liu, G.; Zhang, P.Y.; Sarno, R.; Catanzano, V.; Currie, M.G.; Masferrer, J.L. sGC stimulator praliciguat suppresses stellate cell fibrotic transformation and inhibits fibrosis and inflammation in models of NASH. Proc. Natl Acad. Sci. USA 2019, 116, 11057–11062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefano, J.T.; Cogliati, B.; Santos, F.; Lima, V.M.R.; Mazo, D.C.; Matte, U.; Alvares-da-Silva, M.R.; Silveira, T.R.; Carrilho, F.J.; Oliveira, C.P.M.S. S-nitroso-N-acetylcysteine induces de-differentiation of activated hepatic stellate cells and promotes antifibrotic effects in vitro. Nitric Oxide 2011, 25, 360–365. [Google Scholar] [CrossRef]
- Mann, D.A.; Marra, F. Fibrogenic signalling in hepatic stellate cells. J. Hepatol. 2010, 52, 949–950. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Higashi, T.; Friedman, S.L.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef]
- Jonsson, J.R.; Clouston, A.D.; Ando, Y.; Kelemen, L.I.; Horn, M.J.; Adamson, M.D.; Purdie, D.M.; Powell, E.E. Angiotensin-converting enzyme inhibition attenuates the progressionof rat hepatic fibrosis. Gastroenterology 2001, 121, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Croquet, V.; Moal, F.; Veal, N.; Wang, J.; Oberti, F.; Roux, J.; Vuillemin, E.; Gallois, Y.; Douay, O.; Chappard, D.; et al. Hemodynamic and antifibrotic effects of losartan in rats with liver fibrosis and/orportal hypertension. J. Hepatol. 2002, 37, 773–780. [Google Scholar] [CrossRef]
- Yokohama, S.; Yoneda, M.; Haneda, M.; Okamoto, S.; Okada, M.; Aso, K.; Hasegawa, T.; Tokusashi, Y.; Miyokawa, N.; Nakamura, K. Therapeutic efficacy of an angiotensin II receptor antagonist in patientswith nonalcoholic steatohepatitis. Hepatology 2004, 40, 1222–1225. [Google Scholar] [CrossRef] [PubMed]
- Yatabe, J.; Sanada, H.; Yatabe, M.S.; Hashimoto, S.; Yoneda, M.; Felder, R.A.; Jose, P.A.; Watanabe, T. Angiotensin II type 1 receptor blocker attenuates the activation of ERK and NADPH oxidase by mechanical strain in mesangial cells in the absence of angiotensin II. Am. J. Physiol. Renal Physiol. 2009, 296, F1052–F1060. [Google Scholar] [CrossRef] [Green Version]
- Opie, L.H.; Commerford, P.J.; Gersh, B.J.; Pfeffer, M.A. Controversies in ventricular remodelling. Lancet 2006, 367, 356–367. [Google Scholar] [CrossRef]
- Heymes, C.; Bendall, J.K.; Ratajczak, P.; Cave, A.C.; Samuel, J.L.; Hasenfuss, G.; Shah, A.M. Increased myocardial NADPH oxidase activity in human heart failure. J. Am. Coll. Cardiol. 2003, 41, 2164–2171. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.J.; Vapaatalo, H.; Mervaala, E. Angiotensin II and vascular inflammation. Med. Sci. Monit. 2005, 11, RA194–RA205. [Google Scholar]
- Sonoda, K.; Ohtake, K.; Uchida, H.; Ito, J.; Uchida, M.; Natsume, H.; Tamada, H.; Kobayashi, J. Dietary nitrite supplementation attenuates cardiac remodeling in L-NAME-induced hypertensive rats. Nitric Oxide 2017, 67, 1–9. [Google Scholar] [CrossRef]
- Schieffer, B.; Wirger, A.; Meybrunn, M.; Seitz, S.; Holtz, J.; Riede, U.N.; Drexler, H. Comparative effects of chronic angiotensin-converting enzyme inhibition and angiotensin II type 1 receptor blockade on cardiac remodeling after myocardial infarction in the rat. Circulation 1994, 89, 2273–2282. [Google Scholar] [CrossRef] [Green Version]
- Borém, L.M.A.; Neto, J.F.R.; Brandi, I.V.; Lelis, D.F.; Santos, S.H.S. The role of the angiotensin II type I receptor blocker telmisartan in the treatment of non-alcoholic fatty liver disease: A brief review. Hypertens. Res. 2018, 41, 394–405. [Google Scholar] [CrossRef]
- Ackermann, A.; Fernández-Alfonso, M.S.; Sánchez de Rojas, R.; Ortega, T.; Paul, M.; González, C. Modulation of angiotensin-converting enzyme by nitric oxide. Br. J. Pharmacol. 1998, 124, 291–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiki, T.; Usui, M.; Kato, M.; Funakoshi, Y.; Ito, K.; Egashira, K.; Takeshita, A. Downregulation of angiotensin II type 1 receptor gene transcription by nitric oxide. Hypertension 1998, 31, 342–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M.; Egashira, K.; Usui, M.; Ichiki, T.; Tomita, H.; Shimokawa, H.; Rakugi, H.; Takeshita, A. Cardiac angiotensin II receptors are upregulated by long-term inhibition of nitric oxide synthesis in rats. Circ. Res. 1998, 83, 743–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunyady, L.; Catt, K.J. Pleiotropic AT1 receptor signaling pathways mediating physiological and pathogenic actions of angiotensin II. Mol. Endocrinol. 2006, 20, 953–970. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Kitajima, N.; Saiki, S.; Nakaya, M.; Kurose, H. Regulation of angiotensin II receptor signaling by cysteine modification of NF-κβ. Nitric Oxide 2011, 24, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Kupfahl, C.; Pink, D.; Friedrich, K.; Zurbrügg, H.R.; Neuss, M.; Warnecke, C.; Fielitz, J.; Graf, K.; Fleck, E.; Regitz-Zagrosek, V. Angiotensin II directly increases transforming growth factor beta1 and osteopontin and indirectly affects collagen mRNA expression in the human heart. Cardiovasc. Res. 2000, 46, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Dobaczewski, M.; Chen, W.; Frangogiannis, N.G. Transforming growth factor (TGF)-β signaling in cardiac remodeling. J. Mol. Cell. Cardiol. 2011, 51, 600–606. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Bachetti, T.; Comini, L.; Pasini, E.; Cargnoni, A.; Curello, S.; Ferrari, R. Ace-inhibition with quinapril modulates the nitric oxide pathway in normotensive rats. J. Mol. Cell. Cardiol. 2001, 33, 395–403. [Google Scholar] [CrossRef]
- Minato, T.; Tsutsumi, M.; Tsuchishima, M.; Hayashi, N.; Saito, T.; Matsue, Y.; Toshikuni, N.; Arisawa, T.; George, J. Binge alcohol consumption aggravates oxidative stress and promotes pathogenesis of NASH from obesity-induced simple steatosis. Mol. Med. 2014, 20, 490–502. [Google Scholar] [CrossRef]
- Montenegro, M.F.; Sundqvist, M.L.; Nihlén, C.; Hezel, M.; Carlström, M.; Weitzberg, E.; Lundberg, J.O. Profound differences between humans and rodents in the ability to concentrate salivary nitrate: Implications for translational research. Redox Biol. 2016, 10, 206–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, N.S.; Calvert, J.W.; Elrod, J.W.; Gundewar, S.; Ji, S.Y.; Lefer, D.J. Dietary nitrite supplementation protects against myocardial ischemia-reperfusion injury. Proc. Natl. Acad. Sci. USA 2007, 104, 19144–19149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, A.A. Infantile methemoglobinemia: Reexamining the role of drinking water nitrates. Environ. Health Perspect. 1999, 107, 583–586. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J. Effect of diet and gut environment on the gastrointestinal formation of N-nitroso compounds: A review. Nitric Oxide 2018, 73, 66–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, C.E.; Khatri, J.; Maskell, P.; Odongerel, C.; Webb, A.J. It is rocket science—Why dietary nitrate is hard to ‘beet’! Part II: Further mechanisms and therapeutic potential of the nitrate-nitrite-NO pathway. Br. J. Clin. Pharmacol. 2017, 83, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, J. Chewing well During meals may benefit health via the Enterosalivary nitrate-nitrite-nitric oxide pathway. J. Gastroenterol. Hepatol. Res. 2019, 8, 2882–2885. [Google Scholar] [CrossRef]
- Kozaki, Y.; Umetsu, R.; Mizukami, Y.; Yamamura, A.; Kitamori, K.; Tsuchikura, S.; Ikeda, K.; Yamori, Y. Peripheral gene expression profile of mechanical hyperalgesia induced by repeated cold stress in SHRSP5/Dmcr rats. J. Physiol. Sci. 2015, 65, 417–425. [Google Scholar] [CrossRef]
- Sweet, J.G.; Chan, S.L.; Cipolla, M.J. Effect of hypertension and carotid occlusion on brain parenchymal arteriole structure and reactivity. J. Appl. Physiol. 2015, 119, 817–823. [Google Scholar] [CrossRef] [Green Version]
- Djekoun-Bensoltane, S.; Kammerer, M.; Larhantec, M.; Pilet, N.; Thorin, C. Nitrate and nitrite concentrations in rabbit saliva: Comparison with rat saliva. Environ. Toxicol. Pharmacol. 2007, 23, 132–134. [Google Scholar] [CrossRef]
- Cockburn, A.; Brambilla, G.; Fernández, M.L.; Arcella, D.; Bordajandi, L.R.; Cottrill, B.; van Peteghem, C.; Dorne, J.L. Nitrite in feed: From animal health to human health. Toxicol. Appl. Pharmacol. 2013, 270, 209–217. [Google Scholar] [CrossRef]
- Buelke-Sam, J.; Holson, J.F.; Bazare, J.J.; Young, J.F. Comparative stability of physiological parameters during sustained anesthesia in rats. Lab. Anim. Sci. 1978, 28, 157–162. [Google Scholar] [PubMed]
- Tamada, H.; Naito, H.; Kitamori, K.; Hayashi, Y.; Yamagishi, N.; Kato, M.; Nakajima, T. Efficacy of dietary lipid control in healing high-fat and high-cholesterol diet-induced fibrotic steatohepatitis in rats. PLoS ONE 2016, 11, e0145939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loennechen, J.P.; Støylen, A.; Beisvag, V.; Wisløff, U.; Ellingsen, O. Regional expression of endothelin-1, ANP, IGF-1, and LV wall stress in the infarcted rat heart. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H2902–H2910. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.W.; Huang, Y.W.; Xia, Q.; Xu, Q.W. Combined effects of ramipril and angiotensin II receptor blocker TCV116 on rat congestive heart failure after myocardial infarction. Chin. Med. J. 2005, 118, 146–154. [Google Scholar] [PubMed]
- Ohtake, K.; Ishiyama, Y.; Uchida, H.; Muraki, E.; Kobayashi, J. Dietary nitrite inhibits early glomerular injury in streptozotocin-induced diabetic nephropathy in rats. Nitric Oxide 2007, 17, 75–81. [Google Scholar] [CrossRef]
- Ohtake, K.; Koga, M.; Uchida, H.; Sonoda, K.; Ito, J.; Uchida, M.; Natsume, H.; Kobayashi, J. Oral nitrite ameliorates dextran sulfate sodium-induced acute experimental colitis in mice. Nitric Oxide 2010, 23, 65–73. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sonoda, K.; Kono, Y.; Kitamori, K.; Ohtake, K.; Shiba, S.; Kasono, K.; Kobayashi, J. Beneficial Effects of Dietary Nitrite on a Model of Nonalcoholic Steatohepatitis Induced by High-Fat/High-Cholesterol Diets in SHRSP5/Dmcr Rats: A Preliminary Study. Int. J. Mol. Sci. 2022, 23, 2931. https://doi.org/10.3390/ijms23062931
Sonoda K, Kono Y, Kitamori K, Ohtake K, Shiba S, Kasono K, Kobayashi J. Beneficial Effects of Dietary Nitrite on a Model of Nonalcoholic Steatohepatitis Induced by High-Fat/High-Cholesterol Diets in SHRSP5/Dmcr Rats: A Preliminary Study. International Journal of Molecular Sciences. 2022; 23(6):2931. https://doi.org/10.3390/ijms23062931
Chicago/Turabian StyleSonoda, Kunihiro, Yuka Kono, Kazuya Kitamori, Kazuo Ohtake, Sachiko Shiba, Keizo Kasono, and Jun Kobayashi. 2022. "Beneficial Effects of Dietary Nitrite on a Model of Nonalcoholic Steatohepatitis Induced by High-Fat/High-Cholesterol Diets in SHRSP5/Dmcr Rats: A Preliminary Study" International Journal of Molecular Sciences 23, no. 6: 2931. https://doi.org/10.3390/ijms23062931
APA StyleSonoda, K., Kono, Y., Kitamori, K., Ohtake, K., Shiba, S., Kasono, K., & Kobayashi, J. (2022). Beneficial Effects of Dietary Nitrite on a Model of Nonalcoholic Steatohepatitis Induced by High-Fat/High-Cholesterol Diets in SHRSP5/Dmcr Rats: A Preliminary Study. International Journal of Molecular Sciences, 23(6), 2931. https://doi.org/10.3390/ijms23062931