
The Combination of Intestinal Alkaline Phosphatase Treatment with Moderate Physical Activity Alleviates the Severity of Experimental Colitis in Obese Mice via Modulation of Gut Microbiota, Attenuation of Proinflammatory Cytokines, Oxidative Stress Biomarkers and DNA Oxidative Damage in Colonic Mucosa

 , , , ,
, , , ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
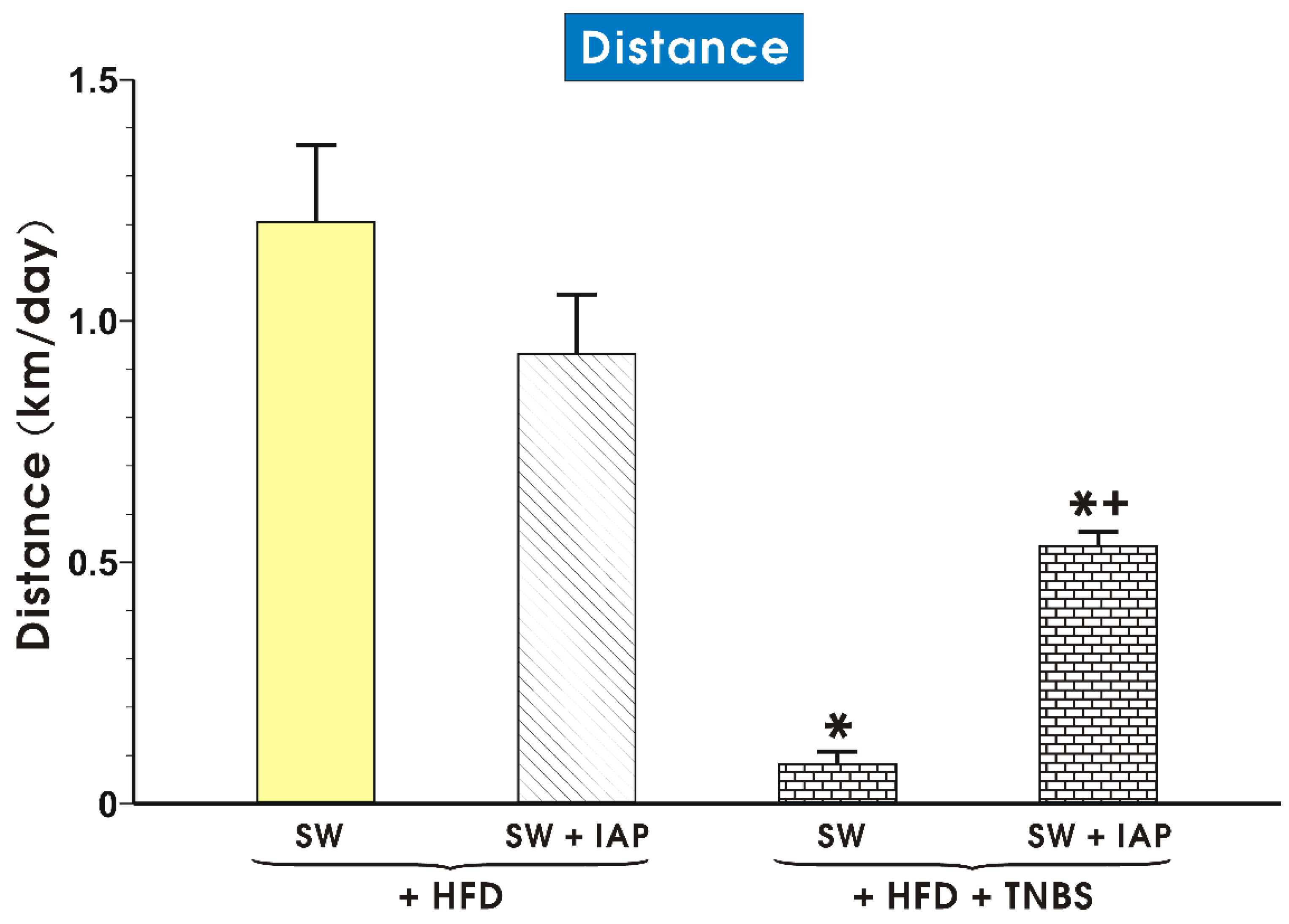
2.1. Effects of IAP Administration with or without SW Exercise on Running Distance, Body Weight and Relative Grip Strength in HFD Fed Mice with Colitis
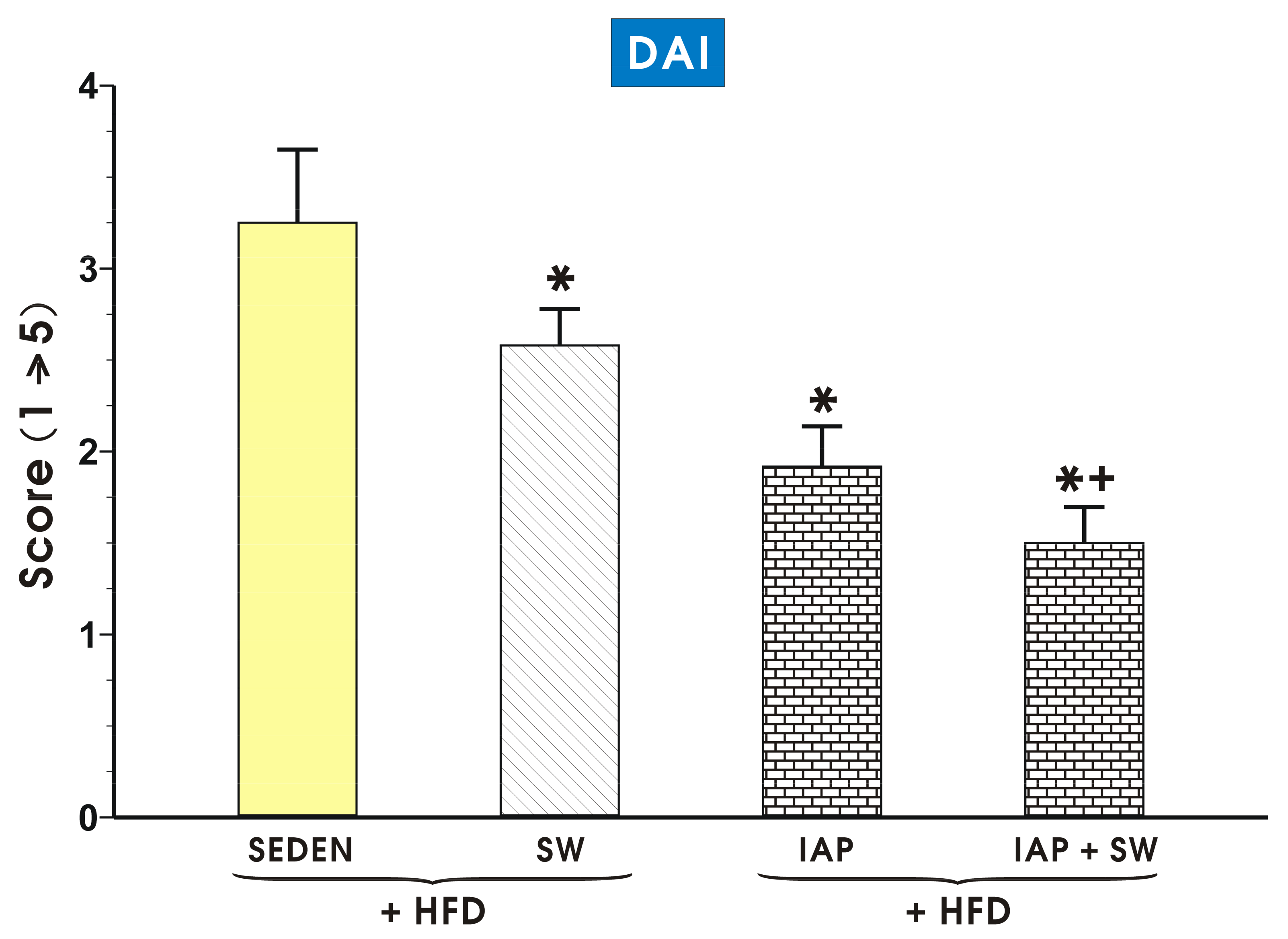
2.2. The Effect of Voluntary Exercise Combined with IAP Administration on DAI Activity and Macroscopic and Microscopic Appearance of Colonic Mucosa in Mice with Colitis
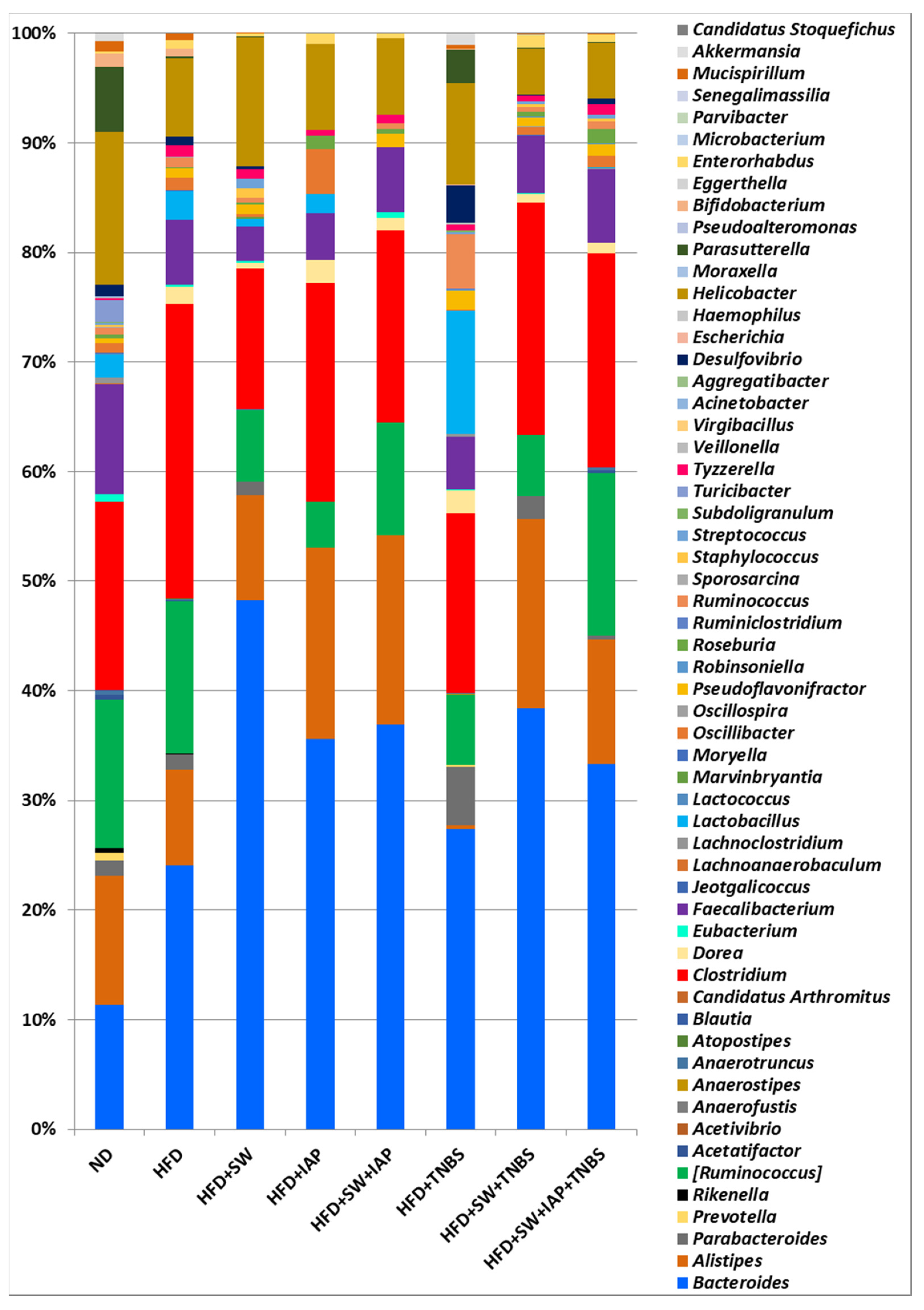
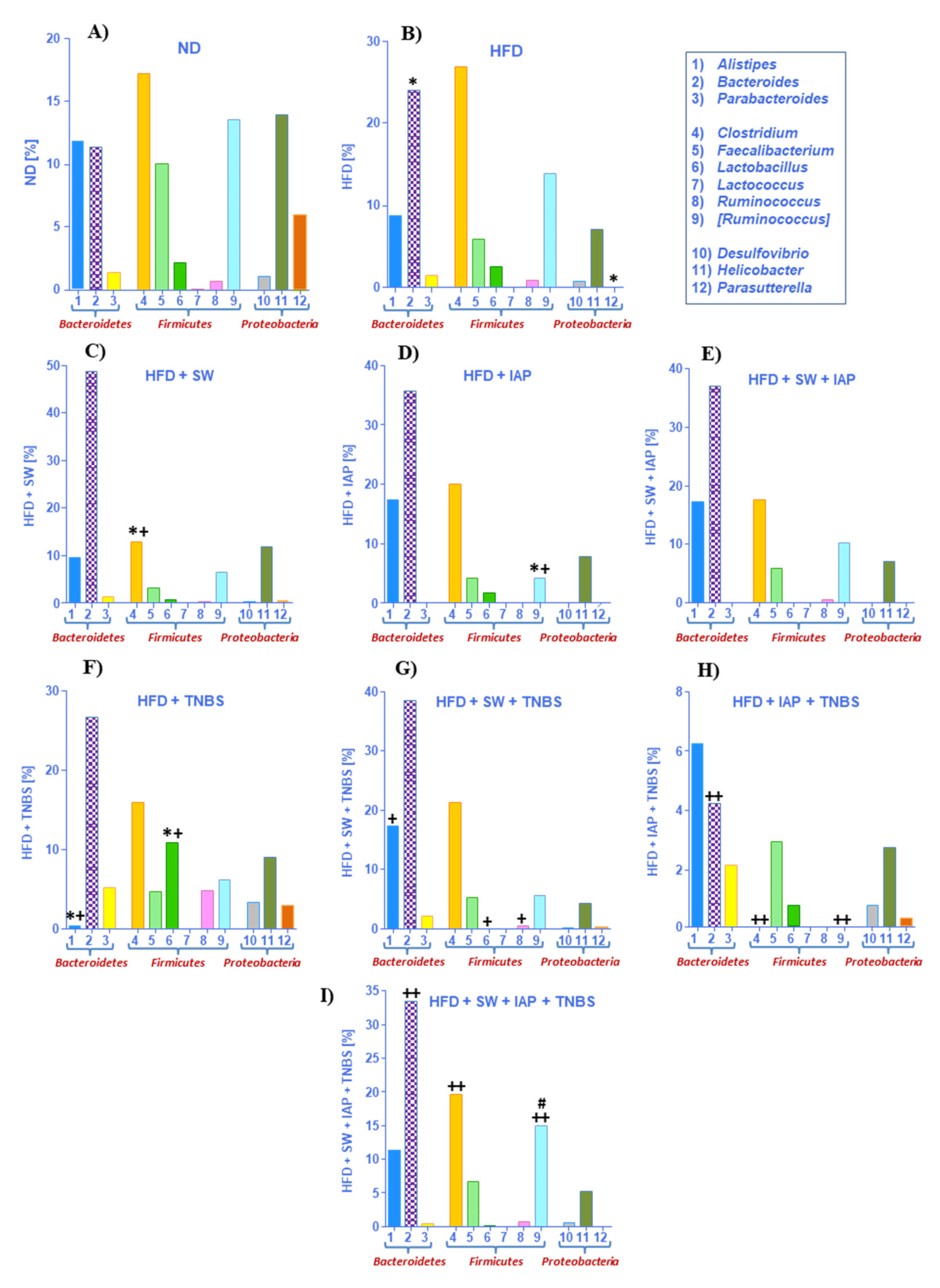
2.3. Changes in Gut Microbiome in Mice with TNBS Colitis with or without Voluntary Physical Activity on SW, IAP Administration Alone or the Combination of SW and IAP
2.4. Effects of Voluntary Exercise Combined with IAP Administration on the Mucosal Colonic Content of MDA plus 4-HNE, GSH, SOD Activity and 8-OHdG Concentration in Mice with Experimental Colitis
2.5. Concentration of IL-2, IL-6, IL-10, IL-12p70, IL-17a, TNF-α, MCP-1 and Leptin in Plasma in Mice with Experimental Colitis, Exercising on SW and Administered with IAP Alone or in Combination with SW
3. Discussion
4. Materials and Methods
4.1. Animals and Diets
4.2. Experimental Design
4.3. Grip Strength Test
4.4. Experimental Colitis Induction
4.5. Macroscopic and Microscopic Changes in the Colonic Mucosa Assessment
4.6. Next Generation Sequencing of Gut Microbiome
4.6.1. DNA Isolation
4.6.2. 16s rRNA Sequencing and Data Analyses
4.7. Lipid Peroxidation Determination
4.8. Total Glutathione (GSH + GSSG) Concentration Measurement
4.9. Superoxide Dismutase (SOD) Activity Determination
4.10. Measurement of 8-Hydroxy-2′-deoxyguanosine (8-OHdG) Concentration
4.11. Luminex Microbeads Fluorescent Assays
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bing, X.; Crusius, J.B.A.; Meuwissen, S.G.M.; Peña, A.S. Inflammatory bowel disease: Definition, epidemiology, etiologic aspects, and immunogenetic studies. World J. Gastroenterol. 1998, 4, 446. [Google Scholar] [CrossRef]
- Abraham, C.; Cho, J.H. Inflammatory Bowel Disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Bilski, J.; Mazur-Bialy, A.; Wojcik, D.; Magierowski, M.; Surmiak, M.; Kwiecien, S.; Magierowska, K.; Hubalewska-Mazgaj, M.; Sliwowski, Z.; Brzozowski, T. Effect of forced physical activity on the severity of experimental colitis in normal weight and obese mice. Involvement of oxidative stress and proinflammatory biomarkers. Nutrients 2019, 11, 1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilski, J.; Brzozowski, B.; Mazur-Bialy, A.; Sliwowski, Z.; Brzozowski, T. The role of physical exercise in inflammatory bowel disease. Biomed Res. Int. 2014, 2014, 429031. [Google Scholar] [CrossRef]
- Barbosa, M.M.C.; Vicentini, F.A.; Castro-Ghizoni, C.V.; Lameira, O.A.; Sa-Nakanishi, A.B.; Bracht, L.; Peralta, R.M.; Natali, M.R.M.; Bracht, A.; Comar, J.F. Copaiba Oil Decreases Oxidative Stress and Inflammation But not Colon Damage in Rats with TNBS-Induced Colitis. Endocr. Metab. Immune Disord.-Drug Targets 2018, 18, 268–280. [Google Scholar] [CrossRef]
- Choghakhori, R.; Abbasnezhad, A.; Hasanvand, A.; Amani, R. Inflammatory cytokines and oxidative stress biomarkers in irritable bowel syndrome: Association with digestive symptoms and quality of life. Cytokine 2017, 93, 34–43. [Google Scholar] [CrossRef]
- Cronin, O.; Barton, W.; Moran, C.; Sheehan, D.; Whiston, R.; Nugent, H.; McCarthy, Y.; Molloy, C.B.; O’Sullivan, O.; Cotter, P.D.; et al. Moderate-intensity aerobic and resistance exercise is safe and favorably influences body composition in patients with quiescent Inflammatory Bowel Disease: A randomized controlled cross-over trial. BMC Gastroenterol. 2019, 19, 29. [Google Scholar] [CrossRef]
- Friedrich, M.; Pohin, M.; Powrie, F. Cytokine Networks in the Pathophysiology of Inflammatory Bowel Disease. Immunity 2019, 50, 992–1006. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Chen, D.; Zheng, P.; Yu, J.; He, J.; Mao, X.; Yu, B. The Bidirectional Interactions between Resveratrol and Gut Microbiota: An Insight into Oxidative Stress and Inflammatory Bowel Disease Therapy. Biomed Res. Int. 2019, 2019, 5403761. [Google Scholar] [CrossRef]
- Liu, G.; Yan, W.; Ding, S.; Jiang, H.; Ma, Y.; Wang, H.; Fang, J. Effects of IRW and IQW on oxidative stress and gut microbiota in dextran sodium sulfate-induced colitis. Cell. Physiol. Biochem. 2018, 51, 441–451. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Nikkhah-Bodaghi, M.; Maleki, I.; Agah, S.; Hekmatdoost, A. Zingiber officinale and oxidative stress in patients with ulcerative colitis: A randomized, placebo-controlled, clinical trial. Complement. Ther. Med. 2019, 43, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Paramsothy, S.; Nielsen, S.; Kamm, M.A.; Deshpande, N.P.; Faith, J.J.; Clemente, J.C.; Paramsothy, R.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; et al. Specific Bacteria and Metabolites Associated with Response to Fecal Microbiota Transplantation in Patients with Ulcerative Colitis. Gastroenterology 2019, 156, 1440–1454.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxid. Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Luo, Y.; Lu, Y.; Wang, D.; Wang, T.; Pu, W.; Wang, Y. Maggot Extracts Alleviate Inflammation and Oxidative Stress in Acute Experimental Colitis via the Activation of Nrf2. Oxid. Med. Cell. Longev. 2019, 2019, 4703253. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Ma, Y.; Ding, S.; Jiang, H.; Fang, J. Effects of Melatonin on Intestinal Microbiota and Oxidative Stress in Colitis Mice. Biomed Res. Int. 2018, 2018, 2607679. [Google Scholar] [CrossRef] [Green Version]
- Duff, W.; Haskey, N.; Potter, G.; Alcorn, J.; Hunter, P.; Fowler, S. Non-pharmacological therapies for inflammatory bowel disease: Recommendations for self-care and physician guidance. World J. Gastroenterol. 2018, 24, 3055–3070. [Google Scholar] [CrossRef]
- Hashash, J.G.; Binion, D.G. Exercise and Inflammatory Bowel Disease: Insights into Etiopathogenesis and Modification of Clinical Course. Gastroenterol. Clin. N. Am. 2017, 46, 895–905. [Google Scholar] [CrossRef]
- Marchioni Beery, R.M.; Li, E.; Fishman, L.N. Impact of pediatric inflammatory bowel disease diagnosis on exercise and sports participation: Patient and parent perspectives. World J. Gastroenterol. 2019, 25, 4493–4501. [Google Scholar] [CrossRef]
- Mazur-Bialy, A.I.; Bilski, J.; Wojcik, D.; Brzozowski, B.; Surmiak, M.; Hubalewska-Mazgaj, M.; Chmura, A.; Magierowski, M.; Magierowska, K.; Mach, T.; et al. Beneficial effect of voluntary exercise on experimental colitis in mice fed a High-Fat diet: The role of irisin, adiponectin and proinflammatory biomarkers. Nutrients 2017, 9, 410. [Google Scholar] [CrossRef] [Green Version]
- Szalai, Z.; Szász, A.; Nagy, I.; Puskás, L.G.; Kupai, K.; Király, A.; Berkó, A.M.; Pósa, A.; Strifler, G.; Baráth, Z.; et al. Anti-inflammatory effect of recreational exercise in TNBS-Induced colitis in rats: Role of NOS/HO/MPO system. Oxid. Med. Cell. Longev. 2014, 2014, 925981. [Google Scholar] [CrossRef] [PubMed]
- Elia, J.; Kane, S. Adult inflammatory bowel disease, physical rehabilitation, and structured exercise. Inflamm. Bowel Dis. 2018, 24, 2543–2549. [Google Scholar] [CrossRef] [PubMed]
- Millán, J.L. Mammalian Alkaline Phosphatases; Wiley: Hoboken, NJ, USA, 2006; ISBN 9783527310791. [Google Scholar]
- Shifrin, D.A.; McConnell, R.E.; Nambiar, R.; Higginbotham, J.N.; Coffey, R.J.; Tyska, M.J. Enterocyte microvillus-derived vesicles detoxify bacterial products and regulate epithelial-microbial interactions. Curr. Biol. 2012, 22, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Hu, D.; Huo, H.; Zhang, W.; Adiliaghdam, F.; Morrison, S.; Ramirez, J.M.; Gul, S.S.; Hamarneh, S.R.; Hodin, R.A. Intestinal alkaline phosphatase regulates tight junction protein levels. J. Am. Coll. Surg. 2016, 222, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Bilski, J.; Wojcik, D.; Danielak, A.; Mazur-Bialy, A.; Magierowski, M.; Tønnesen, K.; Brzozowski, B.; Surmiak, M.; Magierowska, K.; Pajdo, R.; et al. Alternative Therapy in the Prevention of Experimental and Clinical Inflammatory Bowel Disease. Impact of Regular Physical Activity, Intestinal Alkaline Phosphatase and Herbal Products. Curr. Pharm. Des. 2020, 26, 2936–2950. [Google Scholar] [CrossRef]
- Kaliannan, K.; Hamarneh, S.R.; Economopoulos, K.P.; Alam, S.N.; Moaven, O.; Patel, P.; Malo, N.S.; Ray, M.; Abtahi, S.M.; Muhammad, N.; et al. Intestinal alkaline phosphatase prevents metabolic syndrome in mice. Proc. Natl. Acad. Sci. USA 2013, 110, 7003–7008. [Google Scholar] [CrossRef] [Green Version]
- Malo, M.S. A High Level of Intestinal Alkaline Phosphatase Is Protective Against Type 2 Diabetes Mellitus Irrespective of Obesity. EBioMedicine 2015, 2, 2016–2023. [Google Scholar] [CrossRef] [Green Version]
- Molnár, K.; Vannay, Á.; Sziksz, E.; Bánki, N.F.; Győrffy, H.; Arató, A.; Dezsőfi, A.; Veres, G. Decreased mucosal expression of intestinal alkaline phosphatase in children with coeliac disease. Virchows Arch. 2012, 460, 157–161. [Google Scholar] [CrossRef]
- Parlato, M.; Charbit-Henrion, F.; Pan, J.; Romano, C.; Duclaux-Loras, R.; Le Du, M.; Warner, N.; Francalanci, P.; Bruneau, J.; Bras, M.; et al. Human ALPI deficiency causes inflammatory bowel disease and highlights a key mechanism of gut homeostasis. EMBO Mol. Med. 2018, 10, 49–61. [Google Scholar] [CrossRef]
- Whitehouse, J.S.; Riggle, K.M.; Purpi, D.P.; Mayer, A.N.; Pritchard, K.A.; Oldham, K.T.; Gourlay, D.M. The protective role of intestinal alkaline phosphatase in necrotizing enterocolitis. J. Surg. Res. 2010, 163, 79–85. [Google Scholar] [CrossRef]
- De Lisle, R.C.; Mueller, R.; Boyd, M. Impaired mucosal barrier function in the small intestine of the cystic fibrosis mouse. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, G.; Muise, A.M. Monogenic Intestinal Epithelium Defects and the Development of Inflammatory Bowel Disease. Physiology 2018, 33, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Vercalsteren, E.; Vranckx, C.; Lijnen, H.R.; Hemmeryckx, B.; Scroyen, I. Adiposity and metabolic health in mice deficient in intestinal alkaline phosphatase. Adipocyte 2018, 7, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malo, M.S.; Nasrin Alam, S.; Mostafa, G.; Zeller, S.J.; Johnson, P.V.; Mohammad, N.; Chen, K.T.; Moss, A.K.; Ramasamy, S.; Faruqui, A.; et al. Intestinal alkaline phosphatase preserves the normal homeostasis of gut microbiota. Gut 2010, 59, 1476–1484. [Google Scholar] [CrossRef]
- Danielak, A.; Wojcik, D.; Mazur-Bialy, A.; Surmiak, M.; Bilski, J.; Targosz, A.; Magierowski, M.; Chmura, A.; Strzalka, M.; Krzysiek-Maczka, G.; et al. Intestinal Alkaline Phosphatase Combined with Voluntary Physical Activity Alleviates Experimental Colitis in Obese Mice. Involvement of Oxidative Stress, Myokines, Adipokines and Proinflammatory Biomarkers. Antioxidants 2021, 10, 240. [Google Scholar] [CrossRef]
- Elson, C.O.; Cong, Y.; McCracken, V.J.; Dimmitt, R.A.; Lorenz, R.G.; Weaver, C.T. Experimental models of inflammatory bowel disease reveal innate, adaptive, and regulatory mechanisms of host dialogue with the microbiota. Immunol. Rev. 2005, 206, 260–276. [Google Scholar] [CrossRef]
- Estaki, M.; Morck, D.W.; Ghosh, S.; Quin, C.; Pither, J.; Barnett, J.A.; Gill, S.K.; Gibson, D.L. Physical Activity Shapes the Intestinal Microbiome and Immunity of Healthy Mice but Has No Protective Effects against Colitis in MUC2−/− Mice. mSystems 2020, 5, e00515-20. [Google Scholar] [CrossRef]
- Silva, P. Mateus Preclinical Study in Vivo for New Pharmacological Approaches in Inflammatory Bowel Disease: A Systematic Review of Chronic Model of TNBS-Induced Colitis. J. Clin. Med. 2019, 8, 1574. [Google Scholar] [CrossRef] [Green Version]
- Werkstetter, K.J.; Ullrich, J.; Schatz, S.B.; Prell, C.; Koletzko, B.; Koletzko, S. Lean body mass, physical activity and quality of life in paediatric patients with inflammatory bowel disease and in healthy controls. J. Crohn’s Colitis 2012, 6, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Gui, X.; Gui, X.; Köbel, M.; Ferraz, J.G.P.; Iacucci, M.; Ghosh, S.; Liu, S.; Ou, Y.; Perizzolo, M.; Winkfein, R.J.; et al. Histological and molecular diversity and heterogeneity of precancerous lesions associated with inflammatory bowel diseases. J. Clin. Pathol. 2020, 73, 391–402. [Google Scholar] [CrossRef]
- Singh, S.B.; Carroll-Portillo, A.; Coffman, C.; Ritz, N.L.; Lin, H.C. Intestinal Alkaline Phosphatase Exerts Anti-Inflammatory Effects Against Lipopolysaccharide by Inducing Autophagy. Sci. Rep. 2020, 10, 3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, A.K.; Hamarneh, S.R.; Mohamed, M.M.R.; Ramasamy, S.; Yammine, H.; Patel, P.; Kaliannan, K.; Alam, S.N.; Muhammad, N.; Moaven, O.; et al. Intestinal alkaline phosphatase inhibits the proinflammatory nucleotide uridine diphosphate. Am. J. Physiol.-Gastrointest. Liver Physiol. 2013, 304, G597–G604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramasamy, S.; Nguyen, D.D.; Eston, M.A.; Nasrin Alam, S.; Moss, A.K.; Ebrahimi, F.; Biswas, B.; Mostafa, G.; Chen, K.T.; Kaliannan, K.; et al. Intestinal alkaline phosphatase has beneficial effects in mouse models of chronic colitis. Inflamm. Bowel Dis. 2011, 17, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.W.; Kim, J.H.; Lee, C.; Im, J.P.; Kim, J.S. Intestinal alkaline phosphatase ameliorates experimental colitis via toll-like receptor 4-dependent pathway. Eur. J. Pharmacol. 2018, 820, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, R.; Uchida, K.; Fujimaki, H.; Miyagi, M.; Inoue, G.; Sekiguchi, H.; Murata, K.; Takata, K.; Kawakubo, A.; Takaso, M. Elevated leptin levels induce inflammation through IL-6 in skeletal muscle of aged female rats. BMC Musculoskelet. Disord. 2019, 20, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilski, J.; Mazur-Bialy, A.I.; Brzozowski, B.; Magierowski, M.; Jasnos, K.; Krzysiek-Maczka, G.; Urbanczyk, K.; Ptak-Belowska, A.; Zwolinska-Wcislo, M.; Mach, T.; et al. Moderate exercise training attenuates the severity of experimental rodent colitis: The importance of crosstalk between adipose tissue and skeletal muscles. Mediators Inflamm. 2015, 2015, 605071. [Google Scholar] [CrossRef] [PubMed]
- Stidham, R.W.; Higgins, P.D.R. Colorectal Cancer in Inflammatory Bowel Disease. Clin. Colon Rectal Surg. 2018, 31, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.G.; Leonel, A.J.; Aguilar, E.C.; Batista, N.V.; Alves, A.C.; Coimbra, C.C.; Ferreira, A.V.M.; De Faria, A.M.C.; Cara, D.C.; Alvarez Leite, J.I. The combination of high-fat diet-induced obesity and chronic ulcerative colitis reciprocally exacerbates adipose tissue and colon inflammation. Lipids Health Dis. 2011, 10, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benveniste, E.N. Cytokines. In Encyclopedia of the Neurological Sciences; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 921–925. ISBN 9780123851574. [Google Scholar]
- Kleemann, R.; Zadelaar, S.; Kooistra, T. Cytokines and atherosclerosis: A comprehensive review of studies in mice. Cardiovasc. Res. 2008, 79, 360–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochumon, S.; Al Madhoun, A.; Al-Rashed, F.; Thomas, R.; Sindhu, S.; Al-Ozairi, E.; Al-Mulla, F.; Ahmad, R. Elevated adipose tissue associated IL-2 expression in obesity correlates with metabolic inflammation and insulin resistance. Sci. Rep. 2020, 10, 16364. [Google Scholar] [CrossRef] [PubMed]
- Balmus, I.; Ciobica, A.; Trifan, A.; Stanciu, C. The implications of oxidative stress and antioxidant therapies in Inflammatory Bowel Disease: Clinical aspects and animal models. Saudi J. Gastroenterol. 2016, 22, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Gruber, L.; Kisling, S.; Lichti, P.; Martin, F.-P.; May, S.; Klingenspor, M.; Lichtenegger, M.; Rychlik, M.; Haller, D. High Fat Diet Accelerates Pathogenesis of Murine Crohn’s Disease-Like Ileitis Independently of Obesity. PLoS ONE 2013, 8, e71661. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.-S.; Choi, M.-J. Effects of Isoflavone Supplementation on Lipid Profiles and Antioxidant Enzyme Activities in Growing Rats Fed High Fat Diet. Clin. Nutr. Res. 2019, 8, 296. [Google Scholar] [CrossRef]
- de Oliveira Formiga, R.; Alves Júnior, E.B.; Vasconcelos, R.C.; Guerra, G.C.B.; de Araújo, A.A.; de Carvalho, T.G.; Garcia, V.B.; de Araújo Junior, R.F.; Gadelha, F.A.A.F.; Vieira, G.C.; et al. P-cymene and rosmarinic acid ameliorate tnbs-induced intestinal inflammation upkeeping zo-1 and muc-2: Role of antioxidant system and immunomodulation. Int. J. Mol. Sci. 2020, 21, 5870. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Shi, F.; Zhu, J.; Shao, Y.; Gong, W.; Zhou, G.; Wu, H.; She, J.; Shi, W. Piperine, a functional food alkaloid, exhibits inhibitory potential against TNBS-induced colitis via the inhibition of IκB-α/NF-κB and induces tight junction protein (claudin-1, occludin, and ZO-1) signaling pathway in experimental mice. Hum. Exp. Toxicol. 2020, 39, 477–491. [Google Scholar] [CrossRef]
- Manzo, L.P.; De-Faria, F.M.; Dunder, R.J.; Rabelo-Socca, E.A.; Consonni, S.R.; De Almeida, A.C.A.; Souza-Brito, A.R.M.; Luiz-Ferreira, A. Royal jelly and its dual role in TNBS colitis in mice. Sci. World J. 2015, 2015, 956235. [Google Scholar] [CrossRef] [Green Version]
- Liao, J.; Seril, D.N.; Lu, G.G.; Zhang, M.; Toyokuni, S.; Yang, A.L.; Yang, G.Y. Increased susceptibility of chronic ulcerative colitis-induced carcinoma development in DNA repair enzyme Ogg1 deficient mice. Mol. Carcinog. 2008, 47, 638–646. [Google Scholar] [CrossRef] [Green Version]
- Flint, H.J. The impact of nutrition on the human microbiome. Nutr. Rev. 2012, 70, S10–S13. [Google Scholar] [CrossRef]
- Ferranti, E.P.; Dunbar, S.B.; Dunlop, A.L.; Corwin, E.J. Things you didn’t know about: The human gut microbiome. J. Cardiovasc. Nurs. 2014, 29, 479–481. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Novakovic, M.; Rout, A.; Kingsley, T.; Kirchoff, R.; Singh, A.; Verma, V.; Kant, R.; Chaudhary, R. Role of gut microbiota in cardiovascular diseases. World J. Cardiol. 2020, 12, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Rook, G.A.W.; Raison, C.L.; Lowry, C.A. Childhood microbial experience, immunoregulation, inflammation, and adult susceptibility to psychosocial stressors and depression. In Inflammation and Immunity in Depression: Basic Science and Clinical Applications; Elsevier: Amsterdam, The Netherlands, 2018; pp. 17–44. ISBN 9780128110737. [Google Scholar]
- Borody, T.J.; Paramsothy, S.; Agrawal, G. Fecal microbiota transplantation: Indications, methods, evidence, and future directions. Curr. Gastroenterol. Rep. 2013, 15, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Groot, P.F.; Frissen, M.N.; de Clercq, N.C.; Nieuwdorp, M. Fecal microbiota transplantation in metabolic syndrome: History, present and future. Gut Microbes 2017, 8, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Vendrik, K.E.W.; Ooijevaar, R.E.; de Jong, P.R.C.; Laman, J.D.; van Oosten, B.W.; van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal Microbiota Transplantation in Neurological Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef] [Green Version]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916. [Google Scholar] [CrossRef]
- Laukens, D.; Brinkman, B.M.; Raes, J.; De Vos, M.; Vandenabeele, P. Heterogeneity of the gut microbiome in mice: Guidelines for optimizing experimental design. FEMS Microbiol. Rev. 2015, 40, 117–132. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-Induced Obesity Is Linked to Marked but Reversible Alterations in the Mouse Distal Gut Microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.C.; LePard, K.J.; Kwak, J.W.; Stancukas, M.C.; Laskowski, S.; Dougherty, J.; Moulton, L.; Glawe, A.; Wang, Y.; Leone, V.; et al. Exercise prevents weight gain and alters the gut microbiota in a mouse model of high fat diet-induced obesity. PLoS ONE 2014, 9, e92193. [Google Scholar] [CrossRef]
- Angelakis, E.; Armougom, F.; Million, M.; Raoult, D. The relationship between gut microbiota and weight gain in humans. Future Microbiol. 2012, 7, 91–109. [Google Scholar] [CrossRef]
- Yun, Y.; Kim, H.N.; Kim, S.E.; Heo, S.G.; Chang, Y.; Ryu, S.; Shin, H.; Kim, H.L. Comparative analysis of gut microbiota associated with body mass index in a large Korean cohort. BMC Microbiol. 2017, 17, 151. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.W.; Gwak, H.-J.; Moon, S.; Rho, M.; Ryu, J.-H. Functional dynamics of bacterial species in the mouse gut microbiome revealed by metagenomic and metatranscriptomic analyses. PLoS ONE 2020, 15, e0227886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, T.M.; Holster, S.; Wall, R.; König, J.; Brummer, R.J.; de Vos, W.M. Correlating the Gut Microbiome to Health and Disease. In The Gut-Brain Axis Dietary, Probiotic, and Prebiotic Interventions on the Microbiota; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 261–291. ISBN 9780128025444. [Google Scholar]
- Verdu, E.F.; Hayes, C.L.; O’Mahony, S.M. Importance of the Microbiota in Early Life and Influence on Future Health. In The Gut-Brain Axis Dietary, Probiotic, and Prebiotic Interventions on the Microbiota; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 159–184. ISBN 9780128025444. [Google Scholar]
- Sheehan, A.L.; Warren, B.F.; Gear, M.W.L.; Shepherd, N.A. Fat-wrapping in Crohn’s disease: Pathological basis and relevance to surgical practice. Br. J. Surg. 1992, 79, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Van Raemdonck, K.; Umar, S.; Szekanecz, Z.; Zomorrodi, R.K.; Shahrara, S. Impact of obesity on autoimmune arthritis and its cardiovascular complications. Autoimmun. Rev. 2018, 17, 821–835. [Google Scholar] [CrossRef]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus gnavus, a member of the human gut microbiome associated with Crohn’s disease, produces an inflammatory polysaccharide. Proc. Natl. Acad. Sci. USA 2019, 116, 12672–12677. [Google Scholar] [CrossRef] [Green Version]
- Alrafas, H.R.; Busbee, P.B.; Nagarkatti, M.; Nagarkatti, P.S. Resveratrol modulates the gut microbiota to prevent murine colitis development through induction of Tregs and suppression of Th17 cells. J. Leukoc. Biol. 2019, 106, 467–480. [Google Scholar] [CrossRef]
- Satokari, R.; Fuentes, S.; Mattila, E.; Jalanka, J.; De Vos, W.M.; Arkkila, P. Fecal transplantation treatment of antibiotic-induced. Case Rep. Med. 2014, 2014, 13867. [Google Scholar] [CrossRef] [Green Version]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H.J. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef]
- Allen, J.M.; Mailing, L.J.; Cohrs, J.; Salmonson, C.; Fryer, J.D.; Nehra, V.; Hale, V.L.; Kashyap, P.; White, B.A.; Woods, J.A. Exercise training-induced modification of the gut microbiota persists after microbiota colonization and attenuates the response to chemically-induced colitis in gnotobiotic mice. Gut Microbes 2018, 9, 115–130. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, H.Y.; Zhou, H.; Zhan, Q.; Lai, W.; Zeng, Q.; Ren, H.; Xu, D. Moderate-intensity exercise affects gut microbiome composition and influences cardiac function in myocardial infarction mice. Front. Microbiol. 2017, 8, 1687. [Google Scholar] [CrossRef]
- Szatkowski, P.; Krzysciak, W.; Mach, T.; Owczarek, D.; Brzozowski, B.; Szczeklik, K. Nuclear factor-kb-importance, induction of inflammation, and effects of pharmacological modulators in crohn’s disease. J. Physiol. Pharmacol. 2020, 71, 1–13. [Google Scholar]
- Konturek, P.C.; Konturek, K.; Brzozowski, T.; Wojcik, D.; Magierowski, M.; Targosz, A.; Krzysiek-Maczka, G.; Sliwowski, Z.; Strzalka, M.; Magierowska, K.; et al. Participation of the intestinal microbiota in the mechanism of beneficial effect of treatment with synbiotic syngut on experimental colitis under stress conditions. J. Physiol. Pharmacol. 2020, 71, 1–14. [Google Scholar] [CrossRef]
- Castro, B.; Kuang, S. Evaluation of Muscle Performance in Mice by Treadmill Exhaustion Test and Whole-limb Grip Strength Assay. Bio-Protocol 2017, 7, e2237. [Google Scholar] [CrossRef] [PubMed]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kuhl, A.A. A guide to histomorphological evaluation of intestinal inflammation in mouse models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar]
- Zeber-Lubecka, N.; Kulecka, M.; Ambrozkiewicz, F.; Paziewska, A.; Lechowicz, M.; Konopka, E.; Majewska, U.; Borszewska-Kornacka, M.; Mikula, M.; Cukrowska, B.; et al. Effect of Saccharomyces boulardii and Mode of Delivery on the Early Development of the Gut Microbial Community in Preterm Infants. PLoS ONE 2016, 11, e0150306. [Google Scholar] [CrossRef]
- Magierowska, K.; Korbut, E.; Hubalewska-Mazgaj, M.; Surmiak, M.; Chmura, A.; Bakalarz, D.; Buszewicz, G.; Wójcik, D.; Śliwowski, Z.; Ginter, G.; et al. Oxidative gastric mucosal damage induced by ischemia/reperfusion and the mechanisms of its prevention by carbon monoxide-releasing tricarbonyldichlororuthenium (II) dimer. Free Radic. Biol. Med. 2019, 145, 198–208. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pẽa, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (g) |
|---|---|
| ND | 25.53 ± 0.99 |
| HFD | 43.85 ± 1.68 * |
| HFD + SW | 38.22 ± 2.09 *+ |
| HFD + IAP | 32.03 ± 6.28 *+ |
| HFD + SW + IAP | 31.20 ± 1.64 *+ |
| HFD + TNBS | 37.19 ± 2.68 *+ |
| HFD + SW + TNBS | 34.00 ± 4.28 *+ |
| HFD + IAP + TNBS | 44.81 ± 2.09 + |
| HFD + SW + IAP + TNBS | 35.13 ± 2.39 *+ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojcik-Grzybek, D.; Hubalewska-Mazgaj, M.; Surmiak, M.; Sliwowski, Z.; Dobrut, A.; Mlodzinska, A.; Wojcik, A.; Kwiecien, S.; Magierowski, M.; Mazur-Bialy, A.; et al. The Combination of Intestinal Alkaline Phosphatase Treatment with Moderate Physical Activity Alleviates the Severity of Experimental Colitis in Obese Mice via Modulation of Gut Microbiota, Attenuation of Proinflammatory Cytokines, Oxidative Stress Biomarkers and DNA Oxidative Damage in Colonic Mucosa. Int. J. Mol. Sci. 2022, 23, 2964. https://doi.org/10.3390/ijms23062964
Wojcik-Grzybek D, Hubalewska-Mazgaj M, Surmiak M, Sliwowski Z, Dobrut A, Mlodzinska A, Wojcik A, Kwiecien S, Magierowski M, Mazur-Bialy A, et al. The Combination of Intestinal Alkaline Phosphatase Treatment with Moderate Physical Activity Alleviates the Severity of Experimental Colitis in Obese Mice via Modulation of Gut Microbiota, Attenuation of Proinflammatory Cytokines, Oxidative Stress Biomarkers and DNA Oxidative Damage in Colonic Mucosa. International Journal of Molecular Sciences. 2022; 23(6):2964. https://doi.org/10.3390/ijms23062964
Chicago/Turabian StyleWojcik-Grzybek, Dagmara, Magdalena Hubalewska-Mazgaj, Marcin Surmiak, Zbigniew Sliwowski, Anna Dobrut, Agata Mlodzinska, Adrianna Wojcik, Slawomir Kwiecien, Marcin Magierowski, Agnieszka Mazur-Bialy, and et al. 2022. "The Combination of Intestinal Alkaline Phosphatase Treatment with Moderate Physical Activity Alleviates the Severity of Experimental Colitis in Obese Mice via Modulation of Gut Microbiota, Attenuation of Proinflammatory Cytokines, Oxidative Stress Biomarkers and DNA Oxidative Damage in Colonic Mucosa" International Journal of Molecular Sciences 23, no. 6: 2964. https://doi.org/10.3390/ijms23062964
APA StyleWojcik-Grzybek, D., Hubalewska-Mazgaj, M., Surmiak, M., Sliwowski, Z., Dobrut, A., Mlodzinska, A., Wojcik, A., Kwiecien, S., Magierowski, M., Mazur-Bialy, A., Bilski, J., & Brzozowski, T. (2022). The Combination of Intestinal Alkaline Phosphatase Treatment with Moderate Physical Activity Alleviates the Severity of Experimental Colitis in Obese Mice via Modulation of Gut Microbiota, Attenuation of Proinflammatory Cytokines, Oxidative Stress Biomarkers and DNA Oxidative Damage in Colonic Mucosa. International Journal of Molecular Sciences, 23(6), 2964. https://doi.org/10.3390/ijms23062964