
Pathophysiological Roles of Actin-Binding Scaffold Protein, Ezrin


Abstract
:1. The Role of Ezrin as the Cross-Linker between Membrane Proteins and the Actin Cytoskeleton
1.1. Direct Interactions with Membrane Proteins
1.2. Indirect Interactions with Membrane Proteins
1.3. The Role of Ezrin as a Regulator of Plasma Membrane Tension
2. The Role of Ezrin as the Regulators of Rho-Related Proteins
2.1. Interactions with Rho Guanine Nucleotide Exchange Factors (Rho-GEFs)
2.2. Interactions with GTPase-Activating Protein (GAP)
2.3. Interactions with Rho GDP-Dissociation Inhibitors (Rho-GDIs)
2.4. Functional Regulation of Ezrin by Rho-Associated Coiled-Coil Containing Kinase (ROCK)
3. The Role of Ezrin as a Scaffold Protein-Recruiting Proteins Involved in Signal Transduction
3.1. EGF-EGFR Signaling
3.2. HGF-Met Signaling
3.3. CD95 (APO-1/Fas)-Mediated Apoptosis
3.4. PI3-Kinase-AKT Signaling
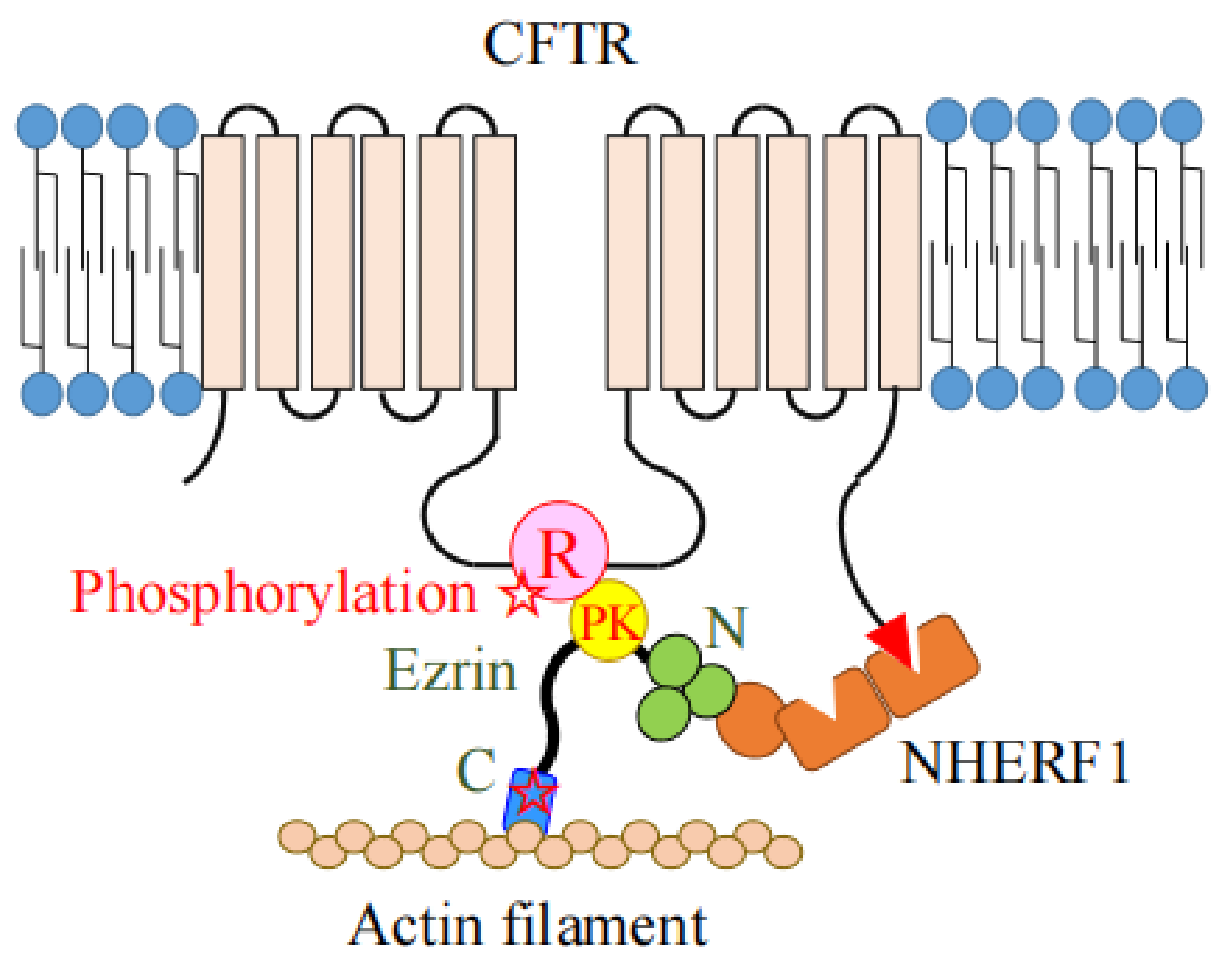
4. The Role of Ezrin as the Protein Kinase A (PKA)-Anchoring Protein (AKAP)
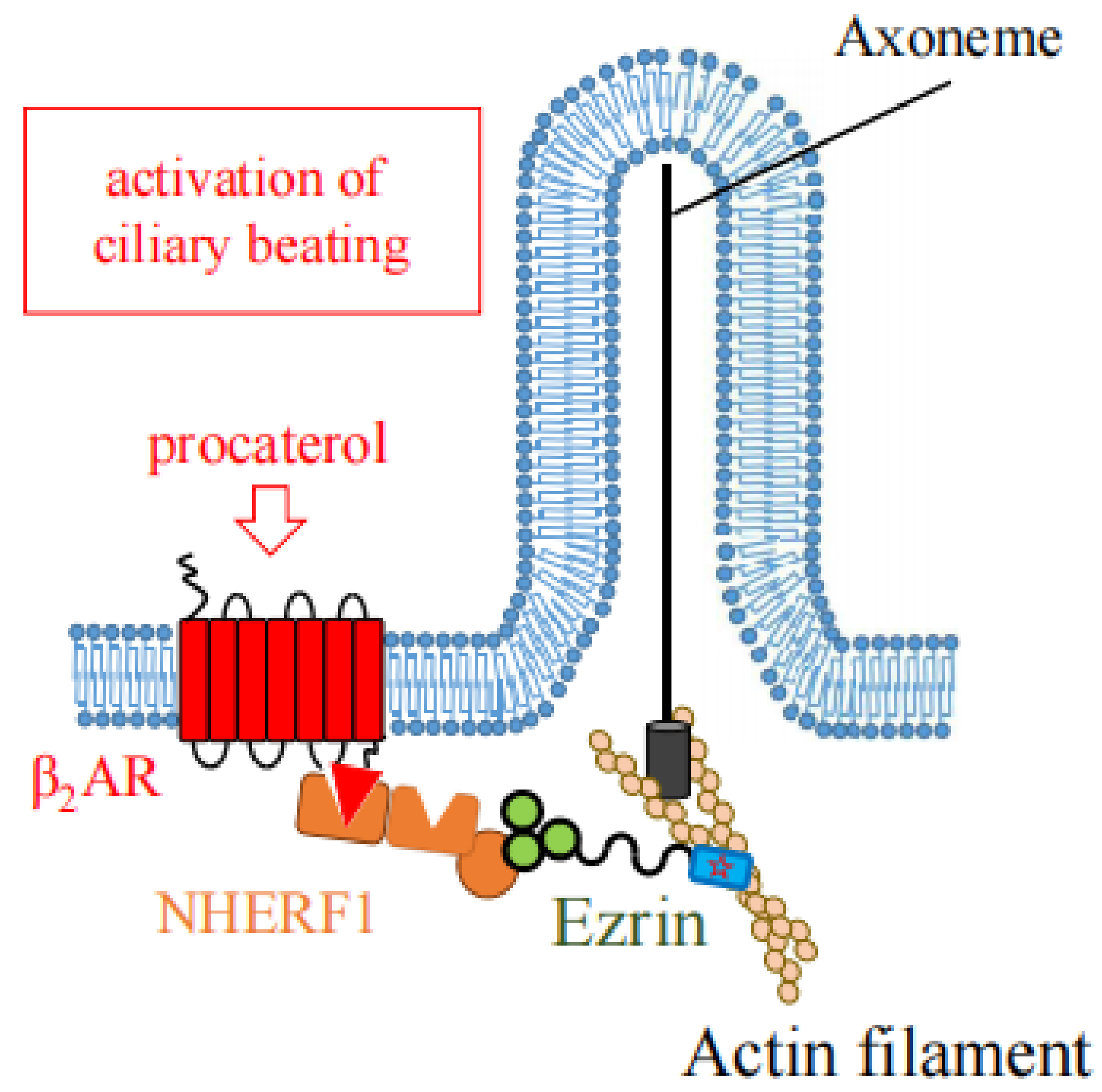
5. The Role of Ezrin in Generation of Cilia, Ciliogenesis
5.1. Roles in Motile Cilia
5.2. Roles in Primary Cilia
6. The Roles of Ezrin in Cancer
7. The Role of Ezrin as the Regulator of mRNA Translation
8. Small Molecules of Ezrin Inhibitor
9. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Functional Roles (Cell/Tissue) |
|---|---|
| Direct Interactions: | |
| Membrane Proteins: | |
| CD44 | Cell surface adhesion receptor involved in cell-cell and cell-matrix interactions [10]. |
| CD43 | Sialoglycoprotein involved in T cell activation (in leukocyte) [11]. |
| ICAM-2 | Ig-like cell adhesion molecule (in leukocyte and endothelial cell) [11]. |
| ICAM-1 | Ig-like cell adhesion molecule (in leukocyte and endothelial cell) [12]. |
| L-selectin | Cell adhesion molecule which induces leukocyte transendothelial migration (in leukocyte) [14]. |
| NHE1 | Transporter exchanging protons for sodium ions (in fibroblast and renal tubule) [16]. |
| P-gp | Multidrug resistance (in intestine, renal tubule and hepatocyte) [17]. |
| ATP11C-b | Phospholipid flippase (in leukocyte) [19]. |
| NHERF1 (EBP50) | Scaffold protein: apical localization of membrane protein [20]. |
| NHERF2 | Scaffold protein: apical localization of membrane protein [21]. |
| Podocalyxin | Sialoglycoprotein which forms the glomerular filtration slits (podocyte) [37]. |
| CD95 | Death receptor (in leukocyte) [50]. |
| Rho-related proteins: | |
| PLEKHG6 | Scaffold protein: apical localization of membrane protein [30]. |
| RhoG | Formation of microvilli and membrane ruffles [30]. |
| ELMO | Ciliary basal body migration [31]. |
| ACAP4 | Localization of the gastric proton pump for gastric acid secretion [32,33]. |
| Rho GDI | Reorganization of actin filaments [36]. |
| AKAP: | |
| PKA | cAMP-dependent protein kinase; parietal cell activation, CFTR-activation [55,56]. |
| Indirect interactions: | |
| Membrane proteins: | |
| CFTR | Chloride channel (in lung, intestine and renal tubule) [22]. |
| NHE3 | Transporter exchanging protons for sodium ions (in gastrointestinal tract and renal tubule [23]. |
| Npt2a | Sodium-dependent phosphate transporter (in renal tubule) [24]. |
| β2AR | Adrenergic receptor (in airway) [25]. |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, S.D.; Keep, N.H. The 2.7 A crystal structure of the activated FERM domain of moesin: An analysis of structural changes on activation. Biochemistry 2001, 40, 7061–7068. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Yoshida, S.; Hatano, R.; Asano, S. Pathophysiological Roles of Ezrin/Radixin/Moesin Proteins. Biol. Pharm. Bull. 2017, 40, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Pelaseyed, T.; Bretscher, A. Regulation of actin-based apical structures on epithelial cells. J. Cell Sci. 2018, 131, jcs221853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehon, R.G.; McClatchey, A.I.; Bretscher, A. Organizing the cell cortex: The role of ERM proteins. Nat. Rev. Mol. Cell Biol. 2010, 11, 276–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andréoli, C.; Martin, M.; Le Borgne, R.; Reggio, H.; Mangeat, P. Ezrin has properties to self-associate at the plasma membrane. J. Cell Sci. 1994, 107, 2509–2521. [Google Scholar] [CrossRef] [PubMed]
- Barret, C.; Roy, C.; Montcourrier, P.; Mangeat, P.; Niggli, V. Mutagenesis of the phosphatidylinositol 4,5-bisphosphate (PIP2) binding site in the NH2-terminal domain of ezrin correlates with its altered cellular distribution. J. Cell Biol. 2000, 151, 1067–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirao, M.; Sato, N.; Kondo, T.; Yonemura, S.; Monden, M.; Sasaki, T.; Takai, Y.; Tsukita, S.; Tsukita, S. Regulation mechanism of ERM (Ezrin/Radixin/Moesin) protein/plasma membrane association: Possible involvement of phosphatidylinositol turnover and Rho-dependent signaling pathway. J. Cell Biol. 1996, 135, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Viswanatha, R.; Ohouo, P.Y.; Smolka, M.B.; Bretscher, A. Local phosphocycling mediated by LOK/SLK restricts ezrin function to the apical aspect of epithelial cells. J. Cell Biol. 2012, 199, 969–984. [Google Scholar] [CrossRef] [Green Version]
- Zaman, R.; Lombardo, A.; Sauvanet, C.; Viswanatha, R.; Awad, V.; Bonomo, L.E.R.; McDermitt, D.; Bretscher, A. Effector-mediated ERM activation locally inhibits RhoA activity to shape the apical cell domain. J. Cell Biol. 2021, 220, e202007146. [Google Scholar] [CrossRef]
- Tsukita, S.; Oishi, K.; Sato, N.; Sagara, J.; Kawai, A.; Tsukita, S. ERM Family Members as Molecular Linkers between the Cell Surface Glycoprotdn CD44 and Actin-based Cytoskeletons. J. Cell Biol. 1994, 126, 391–401. [Google Scholar] [CrossRef]
- Yonemura, S.; Hirao, M.; Doi, Y.; Takahashi, N.; Kondo, T.; Tsukita, S.; Tsukita, S. Ezrin/Radixin/Moesin (ERM) Proteins Bind to a Positively Charged Amino Acid Cluster in the Juxta-Membrane Cytoplasmic Domain of CD44, CD43, and ICAM-2. J. Cell Biol. 1998, 140, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Heiska, L.; Alfthan, K.; Grönholm, M.; Vilja, P.; Vaherii, A.; Carpén, O. Association of Ezrin with Intercellular Adhesion Molecule-1 and -2 (ICAM-1 and ICAM-2). J. Biol. Chem. 1998, 273, 21893–21900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, S.A.; Vega, A.; Riedl, M.; Collins, R.F.; Ostrowski, P.P.; Woods, E.C.; Bertozzi, C.R.; Tammi, M.I.; Lidke, D.S.; Johnson, P.; et al. Transmembrane Pickets Connect Cyto- and Pericellular Skeletons Forming Barriers to Receptor Engagement. Cell 2018, 172, 305–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rey-Gallardo, A.; Tomlins, H.; Joachim, J.; Rahman, I.; Kitscha, P.; Frudd, K.; Parsons, M.; Ivetic, A. Sequential binding of ezrin and moesin to L-selectin regulates monocyte protrusive behavior during transendothelial migration. J. Cell Biol. 2018, 131, jcs215541. [Google Scholar]
- Gowrishankar, K.; Ghosh, S.; Saha, S.; Rumamol, C.; Mayor, S.; Rao, M. Active Remodeling of Cortical Actin Regulates Spatiotemporal Organization of Cell Surface Molecules. Cell 2012, 149, 1353–1367. [Google Scholar] [CrossRef] [Green Version]
- Denker, S.P.; Huang, D.C.; Orlowski, J.; Furthmayr, H.; Barber, D.L. Direct Binding of the Na–H Exchanger NHE1 to ERM Proteins Regulates the Cortical Cytoskeleton and Cell Shape Independently of H+ Translocation. Mol. Cell 2000, 6, 1425–1436. [Google Scholar] [CrossRef]
- Luciani, F.; Molinari, A.; Lozupone, F.; Calcabrini, A.; Lugini, L.; Stringaro, A.; Puddu, P.; Arancia, G.; Cianfriglia, M.; Fais, S. P-glycoprotein-actin association through ERM family proteins: A role in P-glycoprotein function in human cells of lymphoid origin. Blood 2002, 99, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Kobori, T.; Tameishi, M.; Tanaka, C.; Urashima, Y.; Obata, T. Subcellular distribution of ezrin/radixin/moesin and their roles in the cell surface localization and transport function of P-glycoprotein in human colon adenocarcinoma LS180 cells. PLoS ONE 2021, 16, e0250889. [Google Scholar] [CrossRef]
- Inoue, H.; Takatsu, H.; Hamamoto, A.; Takayama, M.; Nakabuchi, R.; Muranaka, Y.; Yagi, T.; Nakayama, K.; Shin, H.-W. The interaction of ATP11C-b with ezrin contributes to its polarized localization. J. Cell Sci. 2021, 134, jcs258523. [Google Scholar] [CrossRef]
- Reczek, D.; Berryman, M.; Bretscher, A. Identification of EBP50: A PDZ-containing Phosphoprotein that Associates with Members of the Ezrin-Radixin-Moesin Family. J. Cell Biol. 1997, 139, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T. Podocyte cytoskeleton is connected to the integral membrane protein podocalyxin through Na+/H+-exchanger regulatory factor 2 and ezrin. Clin. Exp. Nephrol. 2003, 7, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Short, D.; Trotter, K.W.; Reczek, D.; Kreda, S.M.; Bretscher, A.; Boucher, R.C.; Stutts, M.J.; Milgram, S.L. An Apical PDZ Protein Anchors the Cystic Fibrosis Transmembrane Conductance Regulator to the Cytoskeleton. J. Biol. Chem. 1998, 273, 19797–19801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamprecht, G.; Weinman, E.L.; Yun, C.-H.C. The role of NHERF and E3KARP in the cAMP-mediated inhibition of NHE3. J. Biol. Chem. 1998, 273, 29972–29978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatano, R.; Fujii, E.; Segawa, H.; Mukaisho, K.; Matsubara, M.; Miyamoto, K.; Hattori, T.; Sugihara, H.; Asano, S. Ezrin, a membrane cytoskeletal cross-linker, is essential for the regulation of phosphate and calcium homeostasis. Kidney Int. 2013, 83, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Nakayama, S.; Saito, D.; Kogiso, H.; Yasuoka, K.; Marunaka, M.; Nakahari, T.; Asano, S. Ezrin knockdown reduces procaterol-stimulated ciliary beating without morphological changes in mouse airway cilia. J. Cell Sci. 2022, 135, jcs259201. [Google Scholar] [CrossRef]
- Pontes, B.; Monzo, P.; Gauthier, N.C. Membrane tension: A challenging but universal physical parameter in cell biology. Semin. Cell Dev. Biol. 2017, 71, 30–41. [Google Scholar] [CrossRef]
- Brückner, B.R.; Pietuch, A.; Nehls, S.; Rother, J.; Janshof, A. Ezrin is a Major Regulator of Membrane Tension in Epithelial Cells. Sci. Rep. 2015, 5, 14700. [Google Scholar] [CrossRef] [Green Version]
- De Belly, H.; Stubb, A.; Yanagida, A.; Labouesse, C.; Jones, P.H.; Paluch, E.K.; Chalut, K.J. Membrane Tension Gates ERK-Mediated Regulation of Pluripotent Cell Fate. Cell Stem Cell 2021, 28, 273–284. [Google Scholar] [CrossRef]
- Ivetic, A.; Ridley, A.J. Ezrin/radixin/moesin proteins and Rho GTPase signaling in leucocytes. Immunology 2004, 112, 165–176. [Google Scholar] [CrossRef]
- D’Angelo, R.; Aresta, S.; Blangy, A.; Del Maestro, L.; Louvard, D.; Arpin, M. Interaction of ezrin with the novel guanine nucleotide exchange factor PLEKHG6 promotes RhoG-dependent apical cytoskeleton rearrangements in epithelial cells. Mol. Biol. Cell. 2007, 18, 4780–4793. [Google Scholar] [CrossRef] [Green Version]
- Epting, D.; Slanchev, K.; Boehlke, C.; Hoff, S.; Loges, N.T.; Yasunaga, T.; Indorf, L.; Nestel, S.; Lienkamp, S.S.; Omran, H.; et al. The Rac1 regulator ELMO controls basal body migration and docking in multiciliated cells through interaction with Ezrin. Development 2015, 142, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Deng, H.; Wang, D.; Zhou, J.; Huang, Y.; Zhao, X.; Yu, X.; Wang, M.; Wang, F.; Ward, T.; et al. Phospho-regulated ACAP4-Ezrin interaction is essential for histamine-stimulated parietal cell secretion. J. Biol. Chem. 2010, 285, 18769–18780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Yao, P.Y.; Jiang, J.; Zhang, Y.; Su, Z.; Yao, W.; Wang, X.; Gui, P.; Mullen, M.; Henry, C.; et al. MST4 kinase phosphorylates ACAP4 protein to orchestrate apical membrane remodeling during gastric acid secretion. J. Biol. Chem. 2017, 292, 16174–16187. [Google Scholar] [CrossRef] [Green Version]
- Tamura, A.; Kikuchi, S.; Hata, M.; Katsuno, T.; Matsui, T.; Hayashi, H.; Suzuki, Y.; Noda, T.; Tsukita, S. Achlorhydria by ezrin knockdown: Defects in the formation/expansion of apical canaliculi in gastric parietal cells. J. Cell Biol. 2005, 169, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Yamamoto, H.; Tetsui, T.; Kobayakawa, Y.; Hatano, R.; Mukaisho, K.; Hattori, T.; Sugihara, H.; Asano, S. Effects of ezrin knockdown on the structure of gastric glandular epithelia. J. Physiol. Sci. 2016, 66, 53–65. [Google Scholar] [CrossRef]
- Takahashi, K.; Sasaki, T.; Mammoto, A.; Tsukita, S.; Tsukita, S.; Takai, Y. Direct interaction of the Rho GDP dissociation inhibitor with Ezrin/Radixin/Moesin initiates the activation of the Rho small G protein. J. Biol. Chem. 1997, 272, 23365–23371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmieder, S.; Nagai, M.; Orlando, R.A.; Takeda, T.; Farquhar, M.G. Podocalyxin activates RhoA and induces actin reorganization through NHERF1 and ezrin in MDCK cells. J. Am. Soc. Nephrol. 2004, 15, 2289–2298. [Google Scholar] [CrossRef] [Green Version]
- Leung, T.; Manser, E.; Tan, L.; Lim, L. A novel serine/threonine kinase binding the Ras-related RhoA GTPase which translocates the kinase to peripheral membranes. J. Biol. Chem. 1995, 270, 29051–29054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, O.; Fujisawa, K.; Ishizaki, T.; Saito, Y.; Nakao, K.; Narumiya, S. ROCK-I and ROCK-II, two isoforms of Rho-associated coiled-coil forming protein serine/threonine kinase in mice. FEBS Lett. 1996, 392, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Totsukawa, G.; Yamakita, Y.; Yamashiro, S.; Hartshorne, D.J.; Sasaki, Y.; Matsumura, F. Distinct roles of ROCK (Rho-kinase) and MLCK in spatial regulation of MLC phosphorylation for assembly of stress fibers and focal adhesions in 3T3 fibroblasts. J. Cell Biol. 2000, 150, 797–806. [Google Scholar] [CrossRef]
- Quang, C.; Gautreau, A.; Arpin, M.; Treisman, R. Ezrin function is required for ROCK-mediated fibroblast transformation by the Net and Dbl oncogenes. EMBO J. 2000, 19, 4565–4576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, T.; Cooper, J.A. Epidermal growth factor induces rapid tyrosine phosphorylation of proteins in A431 human tumor cells. Cell 1981, 24, 741–752. [Google Scholar] [CrossRef]
- Bretscher, A. Rapid phosphorylation and reorganization of ezrin and spectrin accompany morphological changes induced in A-431 cells by epidermal growth factor. J. Cell Biol. 1989, 108, 921–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saygideğer-Kont, Y.; Minas, T.Z.; Jones, H.; Hour, S.; Çelik, H.; Temel, I.; Han, J.; Atabey, N.; Erkizan, H.V.; Toretsky, J.A.; et al. Ezrin Enhances EGFR Signaling and Modulates Erlotinib Sensitivity in Non-Small Cell Lung Cancer Cells. Neoplasia 2016, 18, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Gherardi, E.; Sandin, S.; Petoukhov, M.V.; Finch, J.; Youles, M.E.; Ofverstedt, L.G.; Miguel, R.N.; Blundell, T.L.; Woude, G.F.V.; Skoglund, U.; et al. Structural basis of hepatocyte growth factor/scatter factor and MET signalling. Proc. Natl. Acad. Sci. USA 2006, 103, 4046–4051. [Google Scholar] [CrossRef] [Green Version]
- Naba, A.; Reverdy, C.; Louvard, D.; Arpin, M. Spatial recruitment and activation of the Fes kinase by ezrin promotes HGF-induced cell scattering. EMBO J. 2008, 27, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Mak, H.; Naba, A.; Varma, S.; Shick, C.; Day, A.; SenGupta, S.K.; Arpin, M.; Eliott, B.E. Ezrin phosphorylation on tyrosine 477 regulates invasion and metastasis of breast cancer cells. BMC Cancer 2012, 12, 82. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Qin, Y.; Zuo, Q.; Bhatnagar, K.; Xiong, J.; Merlino, G.; Yu, Y. Ezrin mediates both HGF/Met autocrine and non-autocrine signaling-induced metastasis in melanoma. Int. J. Cancer 2018, 142, 1652–1663. [Google Scholar] [CrossRef]
- Fais, S.; De Milito, A.; Lozupone, F. The role of FAS to ezrin association in FAS-mediated apoptosis. Apoptosis 2005, 10, 941–947. [Google Scholar] [CrossRef]
- Lozupone, F.; Lugini, L.; Matarrese, P.; Luciani, F.; Federici, C.; Iessi, E.; Margutti, P.; Stassi, G.; Malorni, W.; Fais, S. Identification and relevance of the CD95-binding domain in the N-terminal region of ezrin. J. Biol. Chem. 2004, 279, 9199–9207. [Google Scholar] [CrossRef] [Green Version]
- Algeciras-Schimnich, A.; Le Shen, L.; Barnhart, B.C.; Murmann, A.E.; Burkhardt, J.K.; Peter, M.E. Molecular Ordering of the Initial Signaling Events of CD95. Mol. Cell Biol. 2002, 22, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-H.; Park, J.; Lee, J.-S.; Lee, H.-H. Effects of 1alpha, 25-dihydroxyvitamin D3 on programmed cell death of Ishikawa endometrial cancer cells through ezrin phosphorylation. J. Obstet. Gynaecol. 2017, 37, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Hébert, M.; Potin, S.; Sebbagh, M.; Bertoglio, J.; Bréard, J.; Hamelin, J. Rho-ROCK-dependent ezrin-radixin-moesin phosphorylation regulates Fas-mediated apoptosis in Jurkat cells. J. Immunol. 2008, 181, 5963–5973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautreau, A.; Poullet, P.; Jouvard, D.; Arpin, M. Ezrin, a plasma membrane-microfilament linker, signals cell survival through the phosphatidylinositol 3-kinase/Akt pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 7300–7305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dransfield, D.T.; Bradford, A.J.; Smith, J.; Martin, M.; Roy, C.; Mangeat, P.H.; Goldenring, J.R. Ezrin is a cyclic AMP-dependent protein kinase anchoring protein. EMBO J. 1997, 16, 5–43. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Hug, M.J.; Bradbury, N.A.; Frizzell, R.A. Protein kinase A associates with cystic fibrosis transmembrane conductance regulator via an interaction with ezrin. J. Biol. Chem. 2000, 275, 14360–14366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pidoux, G.; Gerbaud, P.; Dompierre, J.; Lygren, B.; Solstad, T.; Evain-Brion, D.; Taskén, K. A PKA-ezrin-Cx43 signaling complex controls gap junction communication and thereby trophoblast cell fusion. J. Cell Sci. 2014, 127, 4172–4185. [Google Scholar] [PubMed] [Green Version]
- Dukic, A.R.; Haugen, L.H.; Pidoux, G.; Leithe, E.; Bakke, O.; Tasken, K. A protein kinase A-ezrin complex regulates connexin 43 gap junction communication in liver epithelial cells. Cell Signal. 2017, 32, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ruppelt, A.; Mosenden, R.; Grönholm, M.; Aandahl, E.M.; Tobin, D.; Carlson, C.R.; Abrahamsen, H.; Herberg, F.W.; Carpén, O.; Taskén, K. Inhibition of T cell activation by cyclic adenosine 5’-monophosphate requires lipid raft targeting of protein kinase A type I by the A-kinase anchoring protein ezrin. J. Immunol. 2007, 179, 5159–5168. [Google Scholar] [CrossRef] [Green Version]
- Cornez, I.; Tasken, K. Spatiotemporal control of cyclic AMP immunomodulation through the PKA-Csk inhibitory pathway is achieved by anchoring to an Ezrin-EBP50-PAG scaffold in effector T cells. FEBS Lett. 2010, 584, 2681–2688. [Google Scholar] [CrossRef] [Green Version]
- Deming, P.B.; Campbell, S.L.; Stone, J.B.; Rivard, R.L.; Mercier, A.L.; Howe, A.K. Anchoring of Protein Kinase A by ERM (Ezrin-Radixin-Moesin) Proteins Is Required for Proper Netrin Signaling through DCC (Deleted in Colorectal Cancer). J. Biol. Chem. 2015, 290, 5783–5796. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.S.; Griffin, J.N.; Date, P.P.; Liem, K.F., Jr.; Khokha, M.K. WDR5 Stabilizes Actin Architecture to Promote Multiciliated Cell Formation. Dev. Cell. 2018, 46, 595–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; You, Y.; Spoor, M.S.; Richer, E.J.; Kudva, V.V.; Paige, R.C.; Seiler, M.P.; Liebler, J.M.; Zabner, J.; Plopper, C.G.; et al. Foxj1 is required for apical localization of ezrin in airway epithelial cells. J. Cell Sci. 2003, 116, 4935–4945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomperts, B.N.; Gong-Cooper, X.; Hackett, B.P. Foxj1 regulates basal body anchoring to the cytoskeleton of ciliated pulmonary epithelial cells. J. Cell Sci. 2004, 117, 1329–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomperts, B.N.; Kim, L.J.; Flaherty, S.A.; Hackett, B.P. IL-13 Regulates Cilia Loss and foxj1 Expression in Human Airway Epithelium. Am. J. Respir. Cell Mol. Biol. 2007, 37, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Jin, M.; Hu, R.; Wang, H.; Zhang, F.; Yuan, S.; Cao, Y. The Joubert Syndrome Protein Inpp5e Controls Ciliogenesis by Regulating Phosphoinositides at the Apical Membrane. J. Am. Soc. Nephrol. 2017, 28, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Elliott, B.E.; Meens, J.A.; SenGupta, S.K.; Louvard, D.; Arpin, M. The membrane cytoskeletal crosslinker ezrin is required for metastasis of breast carcinoma cells. Breast Cancer Res. 2005, 7, R365–R373. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Kong, J.; Lin, Z.; Yang, Y.; Jin, T.; Xu, M.; Gun, J.; Chen, L. Ezrin promotes breast cancer progression by modulating AKT signals. Br. J. Cancer 2019, 120, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Ling, Z.-Q.; Mukaisho, K.; Yamamoto, H.; Chen, K.-H.; Asano, S.; Araki, Y.; Sugihara, H.; Mao, W.-M.; Hattori, T. Initiation of malignancy by duodenal contents reflux and the role of ezrin in developing esophageal squamous cell carcinoma. Cancer Sci. 2010, 101, 624–630. [Google Scholar] [CrossRef]
- Saito, S.; Yamamoto, H.; Mukaisho, K.; Sato, S.; Higo, T.; Hattori, T.; Yamamoto, G.; Sugihara, H. Mechanisms underlying cancer progression caused by ezrin overexpression in tongue squamous cell carcinoma. PLoS ONE 2013, 8, e54881. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-Q.; Chen, G.-P.; Wu, T.; Yan, J.-P.; Zhou, J.-Y. Expression and clinical significance of ezrin in non-small-cell lung cancer. Clin. Lung Cancer 2012, 13, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Bal, N.; Yildirim, S.; Nursal, T.Z.; Bolat, F.; Kayaselcuk, F. Association of ezrin expression in intestinal and diffuse gastric carcinoma with clinicopathological parameters and tumor type. World J. Gastroenterol. 2007, 13, 3726–3729. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, K.; Sakamoto, H.; Rutherford, T.; Chen, Z.; Satoh, K.; Naftolin, F. Ezrin, a membrane-cytoskeletal linking protein, is involved in the process of invasion of endometrial cancer cells. Cancer Lett. 1999, 147, 31–38. [Google Scholar] [CrossRef]
- Khanna, C.; Wan, X.; Bose, S.; Cassaday, R.; Olomu, O.; Mendoza, A.; Yeung, C.; Gorlick, R.; Hewitt, S.; Helman, L.J. The membrane-cytoskeleton linker ezrin is necessary for osteosarcoma metastasis. Nat. Med. 2004, 10, 182–186. [Google Scholar] [CrossRef]
- Yu, Y.; Khan, J.; Khanna, C.; Helman, L.; Meltzer, P.S.; Merlino, G. Expression profiling identifies the cytoskeletal organizer ezrin and the developmental homeoprotein Six-1 as key metastatic regulators. Nat. Med. 2004, 10, 175–181. [Google Scholar] [CrossRef]
- Weng, W.-H.; Ahlen, J.; Astrom, K.; Lui, W.-O.; Larsson, C. Prognostic impact of immunohistochemical expression of ezrin in highly malignant soft tissue sarcomas. Clin. Cancer Res. 2005, 11, 6198–6204. [Google Scholar] [CrossRef] [Green Version]
- Ilmonen, S.; Vaheri, A.; Asko-Seljavaara, S.; Carpen, O. Ezrin in primary cutaneous melanoma. Mod. Pathol. 2005, 18, 503–510. [Google Scholar] [CrossRef]
- Geiger, K.D.; Stoldt, P.; Schlote, W.; Derouiche, A. Ezrin Immunoreactivity Is Associated with Increasing Malignancy of Astrocytic Tumors but Is Absent in Oligodendrogliomas. Am. J. Pathol. 2000, 157, 1785–1793. [Google Scholar] [CrossRef]
- Athanasopoulou, A.; Aroukatos, P.; Nakas, D.; Repanti, M.; Papadaki, H.; Bravou, V. Decreased ezrin and paxillin expression in human urothelial bladder tumors correlate with tumor progression. Urol. Oncol. 2013, 31, 836–842. [Google Scholar] [CrossRef]
- Song, Y.; Ma, X.; Zhang, M.; Wang, M.; Wang, G.; Ye, Y.; Xia, W. Ezrin Mediates Invasion and Metastasis in Tumorigenesis. Front. Cell Dev. Biol. 2020, 8, 588801. [Google Scholar]
- Li, J.; Wei1, K.; Yu, H.; Jin, D.; Wang, G.; Yu, B. Prognostic Value of Ezrin in Various Cancers: A Systematic Review and Updated Meta-analysis. Sci. Rep. 2015, 5, 17903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoskin, V.; Ghaffari, A.; Elliott, B.E. Ezrin, more than a metastatic detERMinant? Oncotarget 2019, 10, 6755–6757. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, A.; Fujita, H.; Kosaka, T.; Minato, H.; Kiyokawa, E. Clinicopathological significance of heterogeneic ezrin expression in poorly differentiated clusters of colorectal cancers. Cancer Sci. 2019, 110, 2667–2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lin, Z.; Sun, L.; Fan, S.; Huang, Z.; Zhang, D.; Yang, Z.; Li, J.; Chen, W. Akt/Ezrin Tyr353/NF-kB pathway regulates EGF-induced EMT and metastasis in tongue squamous cell carcinoma. Br. J. Cancer 2014, 110, 695–705. [Google Scholar] [CrossRef]
- Chen, M.-J.; Gao, X.-J.; Xu, L.-N.; Liu, X.-H.; Liu, L.-X. Ezrin is required for epithelial-mesenchymal transition induced by TGF-β1 in A549 cells. Int. J. Oncol. 2014, 45, 1515–1522. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.A.; Harrison, G.; Mansel, R.E.; Jiang, W.G. The role of the CD44/ezrin complex in cancer metastasis. Crit. Rev. Oncol. Hematol. 2003, 46, 165–186. [Google Scholar] [CrossRef]
- Herrlich, P.; Morrison, H.; Sleeman, J.; Orian-Rousseau, V.; König, H.; Weg-Remers, S.; Ponta, H. CD44 acts both as a growth- and invasiveness-promoting molecule and as a tumor-suppressing cofactor. Ann. N. Y. Acad. Sci. 2000, 910, 106–118. [Google Scholar] [CrossRef]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev. 2002, 16, 3074–3086. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Bhushan, B.; Mars, W.M.; Bowen, W.; Tao, J.; Orr, A.; Stoops, J.; Yu, Y.; Luo, J.; Duncan, A.W.; et al. Phosphorylated Ezrin (Thr567) Regulates Hippo Pathway and Yes-Associated Protein (Yap) in Liver. Am. J. Pathol. 2020, 190, 1427–1437. [Google Scholar] [CrossRef]
- Quan, C.; Sun, J.; Lin, Z.; Jin, T.; Dong, B.; Meng, Z.; Piao, J. Ezrin promotes pancreatic cancer cell proliferation and invasion through activating the Akt/mTOR pathway and inducing YAP translocation. Cancer Manag. Res. 2019, 11, 6553–6566. [Google Scholar] [CrossRef] [Green Version]
- Quan, C.; Yan, Y.; Qin, Z.; Lin, Z.; Quan, T. Ezrin regulates skin fibroblast size/mechanical properties and YAP-dependent proliferation. J. Cell Commun. Signal. 2018, 12, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Choi, J.; Kim, W.; Dann, P.; Takyar, F.; Gefter, J.V.; Friedman, P.A.; Wysolmerski, J.J. Inhibition of ezrin causes PKCalpha-mediated internalization of erbb2/HER2 tyrosine kinase in breast cancer cells. J. Biol. Chem. 2019, 294, 887–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltz, A.G.; Munschauer, M.; Schwanhäusser, B.; Vasile, A.; Murakawa, Y.; Schueler, M.; Youngs, N.; Penfold-Brown, D.; Drew, K.; Milek, M.; et al. The mRNA-bound proteome and its global occupancy profile on protein-coding transcripts. Mol. Cell. 2012, 46, 674–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castello, A.; Fischer, B.; Eichelbaum, K.; Horos, R.; Beckmann, B.M.; Strein, C.; Davey, N.E.; Humphreys, D.T.; Preiss, T.; Steinmetz, L.M.; et al. Insights into RNA biology from an atlas of mammalian mRNA-binding proteins. Cell 2012, 149, 1393–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çelik, H.; Sajwan, K.P.; Selvanathan, S.P.; Marsh, B.J.; Pai, A.V.; Kont, Y.S.; Han, J.; Minas, T.Z.; Rahim, S.; Erkizan, H.V.; et al. Ezrin Binds to DEAD-Box RNA Helicase DDX3 and Regulates Its Function and Protein Level. Mol. Cell Biol. 2015, 35, 3145–3162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulut, G.; Hong, S.H.; Chen, K.; Beauchamp, E.M.; Rahim, S.; Kosturko, G.W.; Glasgow, E.; Dakshanamurthy, S.; Lee, H.S.; Daar, I.; et al. Small molecule inhibitors of ezrin inhibit the invasive phenotype of osteosarcoma cells. Oncogene 2012, 31, 269–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Ye, C.; Xia, J.; Zhou, Y.; Wu, R. Ezrin T567 phosphorylation regulates migration and invasion of ectopic endometrial stromal cells by changing actin cytoskeleton. Life Sci. 2020, 254, 117681. [Google Scholar] [CrossRef]
- Çelik, H.; Bulut, G.; Han, J.; Graham, G.T.; Minas, T.Z.; Conn, E.J.; Hong, S.-H.; Pauly, G.T.; Hayran, M.; Li, X.; et al. Ezrin Inhibition Up-regulates Stress Response Gene Expression. J. Biol. Chem. 2016, 291, 13257–13270. [Google Scholar] [CrossRef] [Green Version]
- Ghaffari, A.; Hoskin, V.; Turashvili, G.; Varma, S.; Mewburn, J.; Mullins, G.; Greer, P.A.; Kiefer, F.; Day, A.G.; Madarnas, Y.; et al. Intravital imaging reveals systemic ezrin inhibition impedes cancer cell migration and lymph node metastasis in breast cancer. Breast Cancer Res. 2019, 21, 12. [Google Scholar] [CrossRef]
- Çelik, H.; Hong, S.H.; Colón-López, D.D.; Han, J.; Kont, Y.S.; Minas, T.Z.; Swift, M.; Paige, M.; Glasgow, E.; Toretsky, J.A.; et al. Identification of Novel Ezrin Inhibitors Targeting Metastatic Osteosarcoma by Screening Open Access Malaria Box. Mol. Cancer Ther. 2015, 14, 2497–2507. [Google Scholar] [CrossRef] [Green Version]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawaguchi, K.; Asano, S. Pathophysiological Roles of Actin-Binding Scaffold Protein, Ezrin. Int. J. Mol. Sci. 2022, 23, 3246. https://doi.org/10.3390/ijms23063246
Kawaguchi K, Asano S. Pathophysiological Roles of Actin-Binding Scaffold Protein, Ezrin. International Journal of Molecular Sciences. 2022; 23(6):3246. https://doi.org/10.3390/ijms23063246
Chicago/Turabian StyleKawaguchi, Kotoku, and Shinji Asano. 2022. "Pathophysiological Roles of Actin-Binding Scaffold Protein, Ezrin" International Journal of Molecular Sciences 23, no. 6: 3246. https://doi.org/10.3390/ijms23063246
APA StyleKawaguchi, K., & Asano, S. (2022). Pathophysiological Roles of Actin-Binding Scaffold Protein, Ezrin. International Journal of Molecular Sciences, 23(6), 3246. https://doi.org/10.3390/ijms23063246