
Hyperthermia and Serotonin: The Quest for a “Better Cyproheptadine”


Abstract
:1. Introduction
- decreasing thermogenesis ↓ (reducing metabolic rate and brown adipose tissue (BAT) catabolism). BAT is richly innervated by sympathetic efferent fibers and β3 receptor activation induces BAT thermogenesis [1];
- diverting heat to the periphery (cholinergic vasodilation);
- increasing cooling (sweat production by the sudoriparous cholinergic glands and evaporation);
- decreasing motor activity (↑ parasympathetic drive).
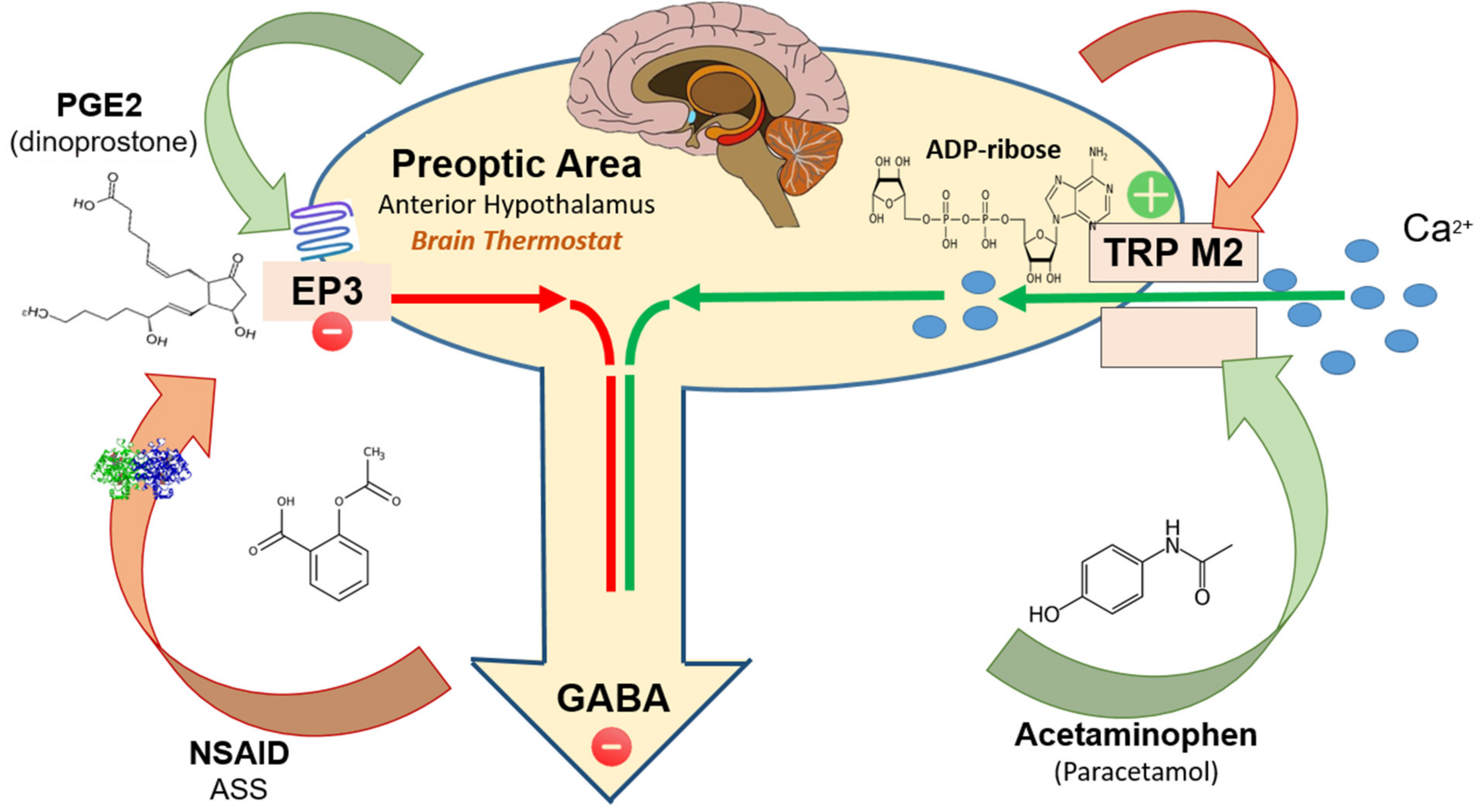
2. TRPM2-Activators
3. EP3-Activators
4. Exogenous TRPM2-Inhibitors
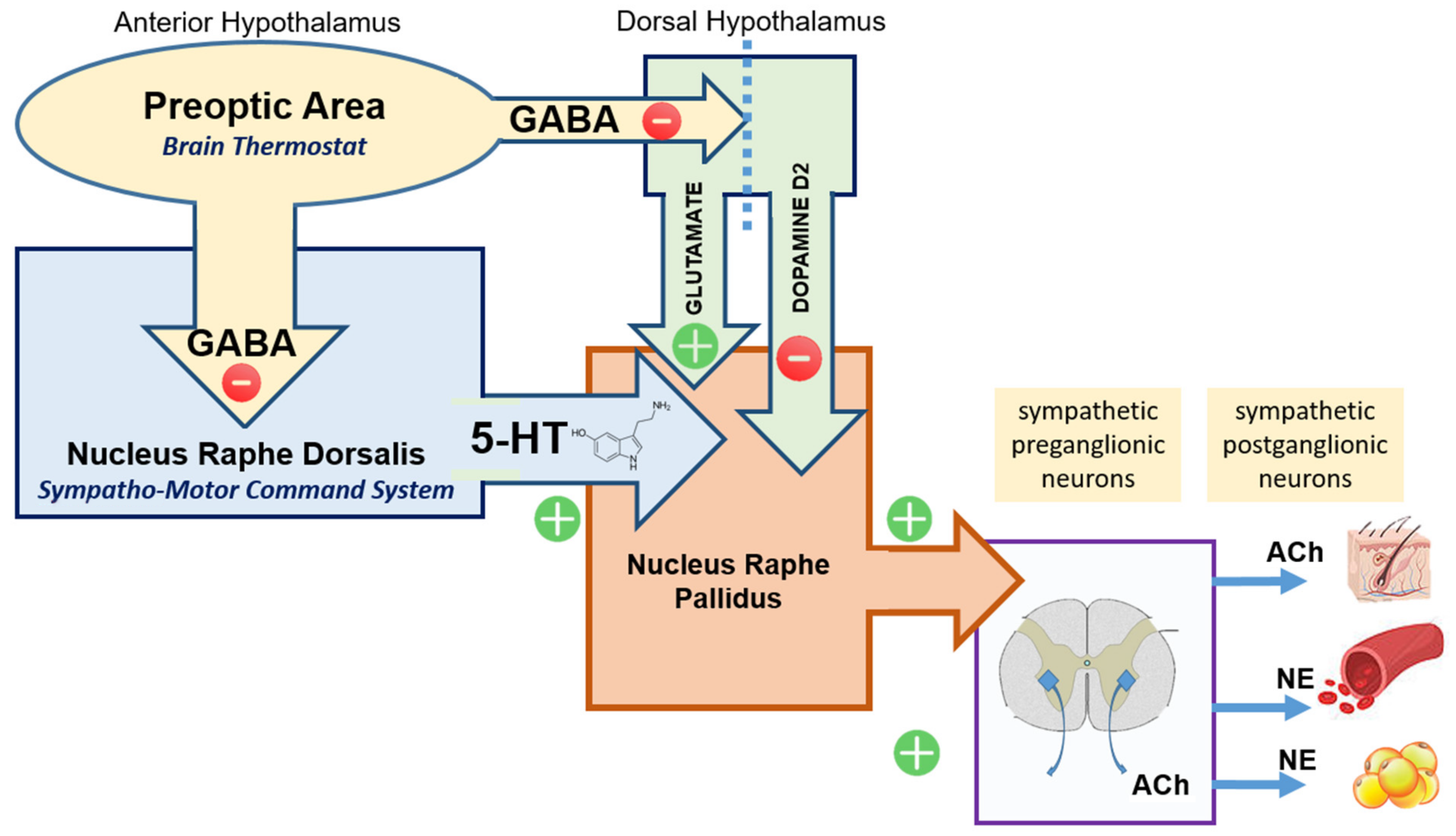
- Output from the preoptic area (POA): The preoptic area of the anterior hypothalamus sends inhibitory efferent output to a number of partner and subaltern sites co-responsible for controlling body temperature;
- Dorsal Hypothalamic Area (DHA): Glutamatergic neurons located in the dorsal hypothalamus are under tonic inhibition from POA. When activated (disinhibited) these neurons cause an increase in body temperature via cascade activation of subaltern downstream structures (raphe pallidus). In contrast, dopaminergic input (via D2 receptors) from the dorsal hypothalamus to raphe pallidus reduces thermogenesis [27];
- Nucleus Raphe Pallidus (NRP): The term raphe refers to a ridge that separates two symmetrical parts of the body, and was used in the naming of the raphe nuclei because this collection of nuclei are clustered around the midline of the brainstem. They are considered part of the reticular formation. The raphe nuclei are the primary location in the brain for serotonin production, and the serotonin synthesized here is distributed throughout the entire central nervous system. While the NRP receives input from the dorsal hypothalamic area, the main input is from the Nucleus Raphe Dorsalis (NRD);
- Nucleus Raphe Dorsalis (NRD) represents the largest population of serotoninergic neurons in the brain. Functionally NRD is viewed as the main Sympatho-Motor Command System. Serotonergic neurons from the sympatho-motor command system [28] project to the nucleus raphe pallidus (NRP) and induce sympathetic activation [29,30,31,32,33] (Figure 2).
5. Neurotransmitters
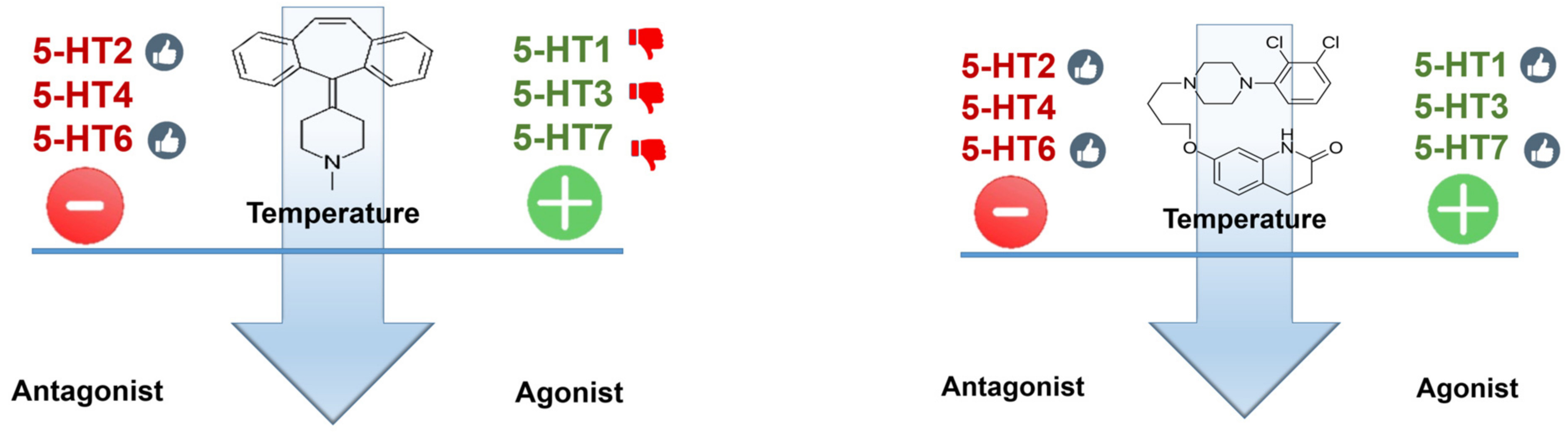
5.1. 5-HT1A Receptors
5.2. 5-HT2 Receptors
5.3. 5-HT3 Receptors
5.4. 5-HT4 Receptor
5.5. 5-HT5 Receptors
5.6. 5-HT6 Receptors
5.7. 5-HT7 Receptors
6. Cyproheptadine
7. Conclusions
Funding
Conflicts of Interest
References
- Carpentier, A.C.; Blondin, D.P.; Virtanen, K.A.; Richard, D.; Haman, F.; Turcotte, É.E. Brown Adipose Tissue Energy Metabolism in Humans. Front. Endocrinol. 2018, 9, 447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siemens, J.; Kamm, G.B. Cellular populations and thermosensing mechanisms of the hypothalamic thermoregulatory center. Pflüg. Arch.—Eur. J. Physiol. 2018, 470, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Kraft, R.; Harteneck, C. The mammalian melastatin-related transient receptor potential cation channels: An overview. Pflüg. Arch.—Eur. J. Physiol. 2005, 451, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Kheradpezhouh, E.; Barritt, G.J.; Rychkov, G.Y. Curcumin inhibits activation of TRPM2 channels in rat hepatocytes. Redox Biol. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Duncan, L.M.; Deeds, J.; Hunter, J.; Shao, J.; Holmgren, L.M.; A Woolf, E.; Tepper, R.I.; Shyjan, A.W. Down-regulation of the novel gene melastatin correlates with potential for melanoma metastasis. Cancer Res. 1998, 58, 1515–1520. [Google Scholar]
- Montell, C. Physiology, Phylogeny, and Functions of the TRP Superfamily of Cation Channels. Sci. STKE Signal Transduct. Knowl. Environ. 2001, 2001, re1. [Google Scholar] [CrossRef] [PubMed]
- Ali, E.S.; Rychkov, G.Y.; Barritt, G.J. TRPM2 Non-Selective Cation Channels in Liver Injury Mediated by Reactive Oxygen Species. Antioxidants 2021, 10, 1243. [Google Scholar] [CrossRef]
- Kamm, G.B.; Siemens, J. The TRPM2 channel in temperature detection and thermoregulation. Temperature 2016, 4, 21–23. [Google Scholar] [CrossRef] [Green Version]
- Song, K.; Wang, H.; Kamm, G.B.; Pohle, J.; Reis, F.D.C.; Heppenstall, P.; Wende, H.; Siemens, J. The TRPM2 channel is a hypothalamic heat sensor that limits fever and can drive hypothermia. Science 2016, 353, 1393–1398. [Google Scholar] [CrossRef]
- Voronova, I.P. 5-HT Receptors and Temperature Homeostasis. Biomolecules 2021, 11, 1914. [Google Scholar] [CrossRef]
- Sharp, T.; Barnes, N.M. Central 5-HT receptors and their function; present and future. Neuropharmacology 2020, 177, 108155. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, T. Activators of TRPM2: Getting it right. J. Gen. Physiol. 2015, 145, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Tóth, B.; Iordanov, I.; Csanády, L. Ruling out pyridine dinucleotides as true TRPM2 channel activators reveals novel direct agonist ADP-ribose-2′-phosphate. J. Gen. Physiol. 2015, 145, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, R.; Liu, Z.; Kraus, W.L. PARPs and ADP-ribosylation: Recent advances linking molecular functions to biological outcomes. Genes Dev. 2017, 31, 101–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kheradpezhouh, E.; Ma, L.; Morphett, A.; Barritt, G.J.; Rychkov, G.Y. TRPM2 channels mediate acetaminophen-induced liver damage. Proc. Natl. Acad. Sci. USA 2014, 111, 3176–3181. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, N.; Kohno, T. Analgesic Effect of Acetaminophen: A Review of Known and Novel Mechanisms of Action. Front. Pharmacol. 2020, 11, 580289. [Google Scholar] [CrossRef]
- Raffa, R.B.; Codd, E.E. Lack of binding of acetaminophen to 5-HT receptor or uptake sites (or eleven other binding/uptake assays). Life Sci. 1996, 59, PL37–PL40. [Google Scholar] [CrossRef]
- Lazarus, M.; Yoshida, K.; Coppari, R.; Bass, C.E.; Mochizuki, T.; Lowell, B.B.; Saper, C.B. EP3 prostaglandin receptors in the median preoptic nucleus are critical for fever responses. Nat. Neurosci. 2007, 10, 1131–1133. [Google Scholar] [CrossRef]
- Hu, H.-Z.; Gu, Q.; Wang, C.; Colton, C.K.; Tang, J.; Kinoshita-Kawada, M.; Lee, L.-Y.; Wood, J.D.; Zhu, M.X. 2-Aminoethoxydiphenyl Borate Is a Common Activator of TRPV1, TRPV2, and TRPV3. J. Biol. Chem. 2004, 279, 35741–35748. [Google Scholar] [CrossRef] [Green Version]
- Togashi, K.; Inada, H.; Tominaga, M. Inhibition of the transient receptor potential cation channel TRPM2 by 2-aminoethoxydiphenyl borate (2-APB). J. Cereb. Blood Flow Metab. 2008, 153, 1324–1330. [Google Scholar] [CrossRef]
- Chen, G.-L.; Zeng, B.; Eastmond, S.; Elsenussi, S.E.; Boa, A.N.; Xu, S.-Z. Pharmacological comparison of novel synthetic fenamate analogues with econazole and 2-APB on the inhibition of TRPM2 channels. J. Cereb. Blood Flow Metab. 2012, 167, 1232–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinamard, R.; Simard, C.; Del Negro, C. Flufenamic acid as an ion channel modulator. Pharmacol. Ther. 2013, 138, 272–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klose, C.; Straub, I.; Riehle, M.; Ranta, F.; Krautwurst, D.; Ullrich, S.; Meyerhof, W.; Harteneck, C. Fenamates as TRP channel blockers: Mefenamic acid selectively blocks TRPM3. J. Cereb. Blood Flow Metab. 2011, 162, 1757–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, T.; Yamamoto, S.; Umehara, N.; Mori, Y.; Wakamori, M.; Shimizu, S. Protective Effects of Duloxetine against Cerebral Ischemia-Reperfusion Injury via Transient Receptor Potential Melastatin 2 Inhibition. J. Pharmacol. Exp. Ther. 2019, 368, 246–254. [Google Scholar] [CrossRef] [Green Version]
- De Donatis, D.; Florio, V.; Porcelli, S.; Saria, A.; Mercolini, L.; Serretti, A.; Conca, A. Duloxetine plasma level and antidepressant response. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 92, 127–132. [Google Scholar] [CrossRef]
- Nedahl, M.; Johansen, S.S.; Linnet, K. Reference Brain/Blood Concentrations of Citalopram, Duloxetine, Mirtazapine and Sertraline. J. Anal. Toxicol. 2017, 42, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Conceição Furber, E.P.S.; Mota, C.M.D.; Veytsman, E.; Morrison, S.F.; Madden, C.J. Dopaminergic input from the posterior hypothalamus to the raphe pallidus area inhibits brown adipose tissue thermogenesis. Am. J. Physiol. Integr. Comp. Physiol. 2021, 321, R938–R950. [Google Scholar] [CrossRef]
- Kerman, I.A.; Akil, H.; Watson, S.J. Rostral Elements of Sympatho-motor Circuitry: A Virally Mediated Transsynaptic Tracing Study. J. Neurosci. 2006, 26, 3423–3433. [Google Scholar] [CrossRef]
- Hermann, D.M.; Luppi, P.-H.; Peyron, C.; Hinckel, P.; Jouvet, M. Afferent projections to the rat nuclei raphe magnus, raphe pallidus and reticularis gigantocellularis pars α demonstrated by iontophoretic application of choleratoxin (subunit b). J. Chem. Neuroanat. 1997, 13, 1–21. [Google Scholar] [CrossRef]
- Ootsuka, Y.; Blessing, W.W. 5-Hydroxytryptamine 1A Receptors Inhibit Cold-Induced Sympathetically Mediated Cutaneous Vasoconstriction in Rabbits. J. Physiol. 2003, 552, 303–314. [Google Scholar] [CrossRef]
- Rusyniak, D.E.; Zaretskaia, M.V.; Zaretsky, D.V.; DiMicco, J.A. 3,4-Methylenedioxymethamphetamine- and 8-Hydroxy-2-di-n-propylamino-tetralin-Induced Hypothermia: Role and Location of 5-Hydroxytryptamine 1A Receptors. J. Pharmacol. Exp. Ther. 2007, 323, 477–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ootsuka, Y.; Blessing, W.W. Activation of slowly conducting medullary raphe-spinal neurons, including serotonergic neurons, increases cutaneous sympathetic vasomotor discharge in rabbit. Am. J. Physiol. Integr. Comp. Physiol. 2005, 288, R909–R918. [Google Scholar] [CrossRef] [PubMed]
- Cerri, M.; Zamboni, G.; Tupone, D.; Dentico, D.; Luppi, M.; Martelli, D.; Perez, E.; Amici, R. Cutaneous vasodilation elicited by disinhibition of the caudal portion of the rostral ventromedial medulla of the free-behaving rat. Neuroscience 2010, 165, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Yamawaki, S. A pharmacological study of veratrine-induced hyperthermia in the rat: A model of neuro-leptic malignant syndrome. Hiroshima J. Med. Sci. 1989, 38, 173–181. [Google Scholar]
- Ramage, A.G. Influence of 5-HT1A receptor agonists on sympathetic and parasympathetic nerve activity. J. Cardiovasc. Pharmacol. 1990, 15 (Suppl. S7), S75–S85. [Google Scholar] [CrossRef]
- Jordan, D. Vagal control of the heart: Central serotonergic (5-HT) mechanisms. Exp. Physiol. 2005, 90, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Audero, E.; Coppi, E.; Mlinar, B.; Rossetti, T.; Caprioli, A.; Al Banchaabouchi, M.; Corradetti, R.; Gross, C. Sporadic Autonomic Dysregulation and Death Associated with Excessive Serotonin Autoinhibition. Science 2008, 321, 130–133. [Google Scholar] [CrossRef]
- Ramage, A.G.; Villalón, C.M. 5-hydroxytryptamine and cardiovascular regulation. Trends Pharmacol. Sci. 2008, 29, 472–481. [Google Scholar] [CrossRef]
- Restrepo, B.; Martín, M.L.; San Román, L.; Morán, A. Peripheral 5-HT1Aand 5-HT7Serotonergic Receptors Modulate Parasympathetic Neurotransmission in Long-Term Diabetic Rats. Exp. Diabetes Res. 2010, 2010, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Ramage, A.G.; Koss, M.C. Pharmacological studies of 8-OH-DPAT-induced pupillary dilation in anesthetized rats. Eur. J. Pharmacol. 2004, 489, 207–213. [Google Scholar] [CrossRef]
- Fanciullacci, M.; Sicuteri, R.; Alessandri, M.; Geppetti, P. Buspirone, but not sumatriptan, induces miosis in humans: Relevance for a serotoninergic pupil control*. Clin. Pharmacol. Ther. 1995, 57, 349–355. [Google Scholar] [CrossRef]
- Klemenhagen, K.C.; Gordon, J.A.; David, D.J.; Hen, R.; Gross, C.T. Increased Fear Response to Contextual Cues in Mice Lacking the 5-HT1A Receptor. Neuropsychopharmacol. 2005, 31, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Guscott, M.R.; Egan, E.; Cook, G.P.; Stanton, J.A.; Beer, M.S.; Rosahl, T.W.; Hartmann, S.; Kulagowski, J.; McAllister, G.; Fone, K.C.; et al. The hypothermic effect of 5-CT in mice is mediated through the 5-HT7 receptor. Neuropharmacology 2003, 44, 1031–1037. [Google Scholar] [CrossRef]
- Hagan, J.J.; Price, G.W.; Jeffrey, P.; Deeks, N.J.; Stean, T.; Piper, D.; Smith, M.I.; Upton, N.; Medhurst, A.D.; Middlemiss, D.N.; et al. Characterization of SB-269970-A, a selective 5-HT7receptor antagonist. J. Cereb. Blood Flow Metab. 2000, 130, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, P.B.; Kelly, L.; Mazur, C.; Lovenberg, T.; Sutcliffe, J.G.; Bonaventure, P. 8-OH-DPAT acts on both 5-HT1A and 5-HT7 receptors to induce hypothermia in rodents. Eur. J. Pharmacol. 2004, 487, 125–132. [Google Scholar] [CrossRef]
- Thomas, D.R.; Melotto, S.; Massagrande, M.; Gribble, A.D.; Jeffrey, P.; Stevens, A.J.; Deeks, N.J.; Eddershaw, P.J.; Fenwick, S.H.; Riley, G.; et al. SB-656104-A, a novel selective 5-HT7receptor antagonist, modulates REM sleep in rats. J. Cereb. Blood Flow Metab. 2003, 139, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.F. Activation of 5-HT1A receptors in raphe pallidus inhibits leptin-evoked increases in brown adipose tissue thermogenesis. Am. J. Physiol. Integr. Comp. Physiol. 2004, 286, R832–R837. [Google Scholar] [CrossRef] [Green Version]
- Gillman, P.K. The serotonin syndrome and its treatment. J. Psychopharmacol. 1999, 13, 100–109. [Google Scholar] [CrossRef]
- Francescangeli, J.; Vaida, S.; Bonavia, A.S. Perioperative Diagnosis and Treatment of Serotonin Syndrome Following Administration of Methylene Blue. Am. J. Case Rep. 2016, 17, 347–351. [Google Scholar] [CrossRef]
- Petroianu, G.A.; Lorke, D.E. The Role of Serotonin in Singultus: A Review. Front. Neurosci. 2020, 14, 629. [Google Scholar] [CrossRef]
- Alefishat, E.; Aloum, L.; Baltatu, O.C.; Petroianu, G.A. The action of aripiprazole and brexpiprazole at the receptor level in singultus. J. Integr. Neurosci. 2021, 20, 247–254. [Google Scholar] [CrossRef]
- Murphy, T.J.; Murnane, K.S. The serotonin 2C receptor agonist WAY-163909 attenuates ketamine-induced hypothermia in mice. Eur. J. Pharmacol. 2019, 842, 255–261. [Google Scholar] [CrossRef]
- Sévoz-Couche, C.; Wang, Y.; Ramage, A.G.; Spyer, K.M.; Jordan, D. In vivo modulation of nucleus tractus solitarius (NTS) neurones by activation of 5-hydroxytryptamine(2) receptors in rats. Neuropharmacology 2000, 39, 2006–2016. [Google Scholar] [CrossRef]
- Naumenko, V.S.; Kondaurova, E.M.; Popova, N.K. Central 5-HT3 receptor-induced hypothermia in mice: Interstrain differences and comparison with hypothermia mediated via 5-HT1A receptor. Neurosci. Lett. 2009, 465, 50–54. [Google Scholar] [CrossRef]
- Kandasamy, S.B. Effect of ondansetron and ICS 205-930 on radiation-induced hypothermia in rats. Radiat. Res. 1997, 147, 741. [Google Scholar] [CrossRef]
- Conductier, G.; Dusticier, N.; Lucas, G.; Côté, F.; Debonnel, G.; Daszuta, A.; Dumuis, A.; Nieoullon, A.; Hen, R.; Bockaert, J.; et al. Adaptive changes in serotonin neurons of the raphe nuclei in 5-HT4receptor knock-out mouse. Eur. J. Neurosci. 2006, 24, 1053–1062. [Google Scholar] [CrossRef]
- Lucas, G.; Debonnel, G. 5-HT4receptors exert a frequency-related facilitatory control on dorsal raphé nucleus 5-HT neuronal activity. Eur. J. Neurosci. 2002, 16, 817–822. [Google Scholar] [CrossRef]
- Lucas, G.; Compan, V.; Charnay, Y.; Neve, R.L.; Nestler, E.J.; Bockaert, J.; Barrot, M.; Debonnel, G. Frontocortical 5-HT4 receptors exert positive feedback on serotonergic activity: Viral transfections, subacute and chronic treatments with 5-HT4 agonists. Biol. Psychiatry 2005, 57, 918–925. [Google Scholar] [CrossRef]
- Brouard, J.T.; Schweimer, J.V.; Houlton, R.; Burnham, K.E.; Quérée, P.; Sharp, T. Pharmacological Evidence for 5-HT6 Receptor Modulation of 5-HT Neuron Firing in Vivo. ACS Chem. Neurosci. 2015, 6, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Emeneses, A. Serotonin, neural markers, and memory. Front. Pharmacol. 2015, 6, 143. [Google Scholar] [CrossRef]
- Pithadia, A.B.; Jain, S.M. 5-Hydroxytryptamine Receptor Subtypes and their Modulators with Therapeutic Potentials. J. Clin. Med. Res. 2009, 1, 72–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, R.; Grysman, N.; Gold, J.; Patel, K.; Grossberg, G.T. The role of 5 HT6-receptor antagonists in Alzheimer’s disease: An update. Expert Opin. Investig. Drugs 2018, 27, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Hedlund, P.B.; Sutcliffe, J.G. Functional, molecular and pharmacological advances in 5-HT7 receptor research. Trends Pharmacol. Sci. 2004, 25, 481–486. [Google Scholar] [CrossRef]
- Hedlund, P.B.; Danielson, P.E.; Thomas, E.A.; Slanina, K.; Carson, M.J.; Sutcliffe, J.G. No hypothermic response to serotonin in 5-HT 7 receptor knockout mice. Proc. Natl. Acad. Sci. USA 2003, 100, 1375–1380. [Google Scholar] [CrossRef] [Green Version]
- Naumenko, V.S.; Kondaurova, E.M.; Popova, N.K. On the role of brain 5-HT7 receptor in the mechanism of hypothermia: Comparison with hypothermia mediated via 5-HT1A and 5-HT3 receptor. Neuropharmacology 2011, 61, 1360–1365. [Google Scholar] [CrossRef]
- Landry, E.S.; Lapointe, N.P.; Rouillard, C.; Levesque, D.; Hedlund, P.B.; Guertin, P.A. Contribution of spinal 5-HT1Aand 5-HT7receptors to locomotor-like movement induced by 8-OH-DPAT in spinal cord-transected mice. Eur. J. Neurosci. 2006, 24, 535–546. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cyproheptadine | Aripiprazole | Serotonin | |
| 5-HT1A | 60 | 1–10 partial agonist | 4 |
| 5-HT2A | 2 | 4–60 antagonist | 12 |
| 5-HT2B | 2 | 1 antagonist | 10 |
| 5-HT2C | 2 | 15 weak agonist Emax ≈ 10% | 5 |
| 5-HT3 | 230 | 500–700 | 200–600 |
| 5-HT4 | Not available | No effect | 120 |
| 5-HT5 | Not available | No effect | 200–300 |
| 5-HT6 | 140 | 140–700 weak antagonist | 60–120 |
| 5-HT7 | 120 | 40 weak agonist | 2–10 |
| SERT | 4000 | 100 (IC50) | 500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petroianu, G.A. Hyperthermia and Serotonin: The Quest for a “Better Cyproheptadine”. Int. J. Mol. Sci. 2022, 23, 3365. https://doi.org/10.3390/ijms23063365
Petroianu GA. Hyperthermia and Serotonin: The Quest for a “Better Cyproheptadine”. International Journal of Molecular Sciences. 2022; 23(6):3365. https://doi.org/10.3390/ijms23063365
Chicago/Turabian StylePetroianu, Georg A. 2022. "Hyperthermia and Serotonin: The Quest for a “Better Cyproheptadine”" International Journal of Molecular Sciences 23, no. 6: 3365. https://doi.org/10.3390/ijms23063365
APA StylePetroianu, G. A. (2022). Hyperthermia and Serotonin: The Quest for a “Better Cyproheptadine”. International Journal of Molecular Sciences, 23(6), 3365. https://doi.org/10.3390/ijms23063365