
Dynamics of Plasma and Urinary Extracellular DNA in Acute Kidney Injury

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Concentrations of Creatinine and Urea in the Plasma
2.2. Concentrations of ecDNA in the Plasma
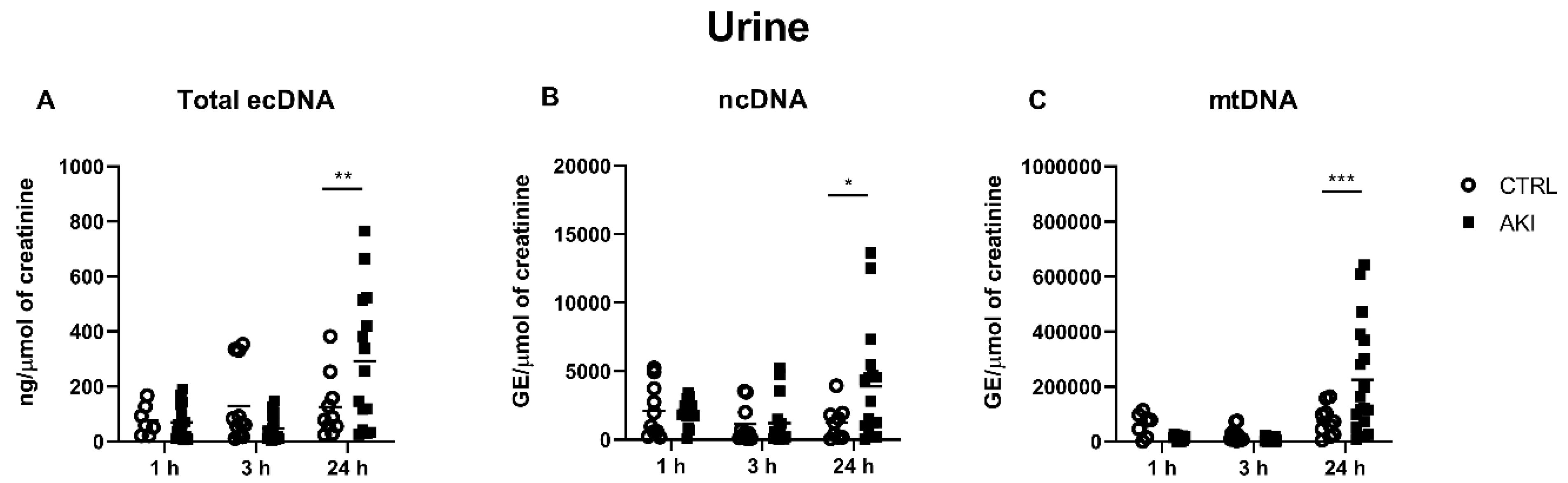
2.3. Concentrations of ecDNA in the Urine
2.4. Correlations between the AKI Functional Markers and DNA Measurements
3. Discussion
4. Methods
4.1. Study Design
4.2. Biochemical Analysis
4.3. DNA Isolation and Quantification
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- K.A.K.I. Working Group. Summary of recommendation statements. Kidney Int. Suppl. 2012, 2, 8–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lameire, N.H.; Bagga, A.; Cruz, D.; De Maeseneer, J.; Endre, Z.; Kellum, J.; Liu, K.D.; Mehta, R.L.; Pannu, N.; Van Biesen, W.; et al. Acute kidney injury: An increasing global concern. Lancet 2013, 382, 170–179. [Google Scholar] [CrossRef]
- Hoste, E.A.J.; Kellum, J.A.; Selby, N.M.; Zarbock, A.; Palevsky, P.M.; Bagshaw, S.M.; Goldstein, S.L.; Cerdá, J.; Chawla, L.S. Global epidemiology and outcomes of acute kidney injury. Nat. Rev. Nephrol. 2018, 14, 607–625. [Google Scholar] [CrossRef]
- Sarnak, M.J.; Amann, K.; Bangalore, S.; Cavalcante, J.L.; Charytan, D.; Craig, J.; Gill, J.S.; Hlatky, M.A.; Jardine, A.G.; Landmesser, U.; et al. Chronic kidney disease and coronary artery disease: JACC state-of-the-art review. J. Am. Coll. Cardiol. 2019, 74, 1823–1838. [Google Scholar] [CrossRef]
- Ronco, C.; Bellomo, R.; Kellum, J.A. Acute kidney injury. Lancet 2019, 394, 1949–1964. [Google Scholar] [CrossRef]
- Parikh, C.R.; Mansour, S.G. Perspective on clinical application of biomarkers in AKI. J. Am. Soc. Nephrol. 2017, 28, 1677–1685. [Google Scholar] [CrossRef] [Green Version]
- Celec, P.; Vlková, B.; Lauková, L.; Babickova, J.; Boor, P. Cell-free DNA: The role in pathophysiology and as a biomarker in kidney diseases. Expert Rev. Mol. Med. 2018, 20, e1. [Google Scholar] [CrossRef]
- Salvi, S.; Casadio, V. Urinary cell-free DNA: Potential and applications. Methods Mol. Biol. 2019, 1909, 201–209. [Google Scholar] [CrossRef]
- Grabuschnig, S.; Bronkhorst, A.J.; Holdenrieder, S.; Rodriguez, I.R.; Schliep, K.P.; Schwendenwein, D.; Ungerer, V.; Sensen, C.W. Putative origins of cell-free DNA in humans: A review of active and passive nucleic acid release mechanisms. Int. J. Mol. Sci. 2020, 21, 8062. [Google Scholar] [CrossRef]
- Raup-Konsavage, W.M.; Wang, Y.; Wang, W.W.; Feliers, D.; Ruan, H.; Reeves, W.B. Neutrophil peptidyl arginine deiminase-4 has a pivotal role in ischemia/reperfusion-induced acute kidney injury. Kidney Int. 2018, 93, 365–374. [Google Scholar] [CrossRef]
- Hu, Q.; Ren, J.; Ren, H.; Wu, J.; Wu, X.; Liu, S.; Wang, G.; Gu, G.; Guo, K.; Li, J. Urinary mitochondrial DNA identifies renal dysfunction and mitochondrial damage in sepsis-induced acute kidney injury. Oxid. Med. Cell. Longev. 2018, 2018, 8074936. [Google Scholar] [CrossRef]
- Clementi, A.; Virzì, G.M.; Brocca, A.; Pastori, S.; De Cal, M.; Marcante, S.; Granata, A.; Ronco, C. The role of cell-free plasma DNA in critically ill patients with sepsis. Blood Purif. 2016, 41, 34–40. [Google Scholar] [CrossRef]
- Likhvantsev, V.V.; Landoni, G.; Grebenchikov, O.A.; Skripkin, Y.V.; Zabelina, T.S.; Zinovkina, L.A.; Prikhodko, A.S.; Lomivorotov, V.V.; Zinovkin, R.A. Nuclear DNA as predictor of acute kidney injury in patients undergoing coronary artery bypass graft: A pilot study. J. Cardiothorac. Vasc. Anesth. 2017, 31, 2080–2085. [Google Scholar] [CrossRef]
- Merkle, J.; Daka, A.; Deppe, A.C.; Wahlers, T.; Paunel-Görgülü, A. High levels of cell-free DNA accurately predict late acute kidney injury in patients after cardiac surgery. PLoS ONE 2019, 14, e0218548. [Google Scholar] [CrossRef]
- Filippone, E.J.; Farber, J.L. The monitoring of donor-derived cell-free DNA in kidney transplantation. Transplantation 2021, 105, 509–516. [Google Scholar] [CrossRef]
- Bronkhorst, A.J.; Ungerer, V.; Diehl, F.; Anker, P.; Dor, Y.; Fleischhacker, M.; Gahan, P.B.; Hui, L.; Holdenrieder, S.; Thierry, A.R. Towards systematic nomenclature for cell-free DNA. Hum. Genet. 2021, 140, 565–578. [Google Scholar] [CrossRef]
- Homolová, J.; Janovičová, Ľ.; Konečná, B.; Vlková, B.; Celec, P.; Tóthová, Ľ.; Bábíčková, J. plasma concentrations of extracellular DNA in acute kidney injury. Diagnostics 2020, 10, 152. [Google Scholar] [CrossRef] [Green Version]
- Okubo, K.; Kurosawa, M.; Kamiya, M.; Urano, Y.; Suzuki, A.; Yamamoto, K.; Hase, K.; Homma, K.; Sasaki, J.; Miyauchi, H.; et al. Macrophage extracellular trap formation promoted by platelet activation is a key mediator of rhabdomyolysis-induced acute kidney injury. Nat. Med. 2018, 24, 232–238. [Google Scholar] [CrossRef]
- Nakazawa, D.; Kumar, S.V.; Marschner, J.; Desai, J.; Holderied, A.; Rath, L.; Kraft, F.; Lei, Y.; Fukasawa, Y.; Moeckel, G.; et al. Histones and neutrophil extracellular traps enhance tubular necrosis and remote organ injury in ischemic AKI. J. Am. Soc. Nephrol. 2017, 28, 1753–1768. [Google Scholar] [CrossRef] [Green Version]
- Jansen, M.P.; Emal, D.; Teske, G.J.; Dessing, M.C.; Florquin, S.; Roelofs, J.J. Release of extracellular DNA influences renal ischemia reperfusion injury by platelet activation and formation of neutrophil extracellular traps. Kidney Int. 2017, 91, 352–364. [Google Scholar] [CrossRef] [Green Version]
- Dvm, V.C.D.; Goggs, R.; Hansen, C.; Dvm, C.W.F.; Letendre, J.-A.; Wakshlag, J.J. Serum myoglobin, creatine kinase, and cell-free DNA in endurance sled dogs and sled dogs with clinical rhabdomyolysis. J. Vet. Emerg. Crit. Care 2018, 28, 310–316. [Google Scholar] [CrossRef]
- Ramos, M.V.; Mejias, M.P.; Sabbione, F.; Fernandez-Brando, R.J.; Santiago, A.P.; Amaral, M.M.; Exeni, R.; Trevani, A.; Palermo, M.S. Induction of neutrophil extracellular traps in shiga toxin-associated hemolytic uremic syndrome. J. Innate Immun. 2016, 8, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Moon, H.; Lee, Y.H.; Seo, J.-W.; Kim, Y.G.; Moon, J.-Y.; Kim, J.S.; Jeong, K.-H.; Lee, T.W.; Ihm, C.-G.; et al. Clinical relevance of cell-free mitochondrial DNA during the early postoperative period in kidney transplant recipients. Sci. Rep. 2019, 9, 18607. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.P.B.; Pulskens, W.P.C.; Uil, M.; Claessen, N.; Nieuwenhuizen, G.; Standaar, D.; Hau, C.M.; Nieuwland, R.; Florquin, S.; Bemelman, F.J.; et al. Urinary mitochondrial DNA associates with delayed graft function following renal transplantation. Nephrol. Dial. Transplant. 2020, 35, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Arruda, N.M.; Braz, L.G.; Nogueira, F.R.; Souza, K.M.; Aun, A.G.; Figueiredo, D.B.; Lara, J.R.; Silva, M.A.P.; Golim, M.A.; de Carvalho, L.R.; et al. Inflammation and DNA damage induction in surgical patients maintained with desflurane anesthesia. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 846, 403073. [Google Scholar] [CrossRef] [PubMed]
- Delogu, G.; Moretti, S.; Famularo, G.; Antonucci, A.; Signore, L.; Marcellini, S.; Bosco, L.L.; De Simone, C. Circulating neutrophils exhibit enhanced apoptosis associated with mitochondrial dysfunctions after surgery under general anaesthesia. Acta Anaesthesiol. Scand. 2001, 45, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Brand, J.-M.; Frohn, C.; Luhm, J.; Kirchner, H.; Schmucker, P. Early alterations in the number of circulating lymphocyte subpopulations and enhanced proinflammatory immune response during opioid-based general anesthesia. Shock 2003, 20, 213–217. [Google Scholar] [CrossRef]
- Lo, Y.M.D.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.; Wainscoat, J.S. Presence of fetal DNA in maternal plasma and serum. Lancet 1997, 350, 485–487. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jančuška, A.; Potočárová, A.; Kovalčíková, A.G.; Podracká, Ľ.; Bábíčková, J.; Celec, P.; Tóthová, Ľ. Dynamics of Plasma and Urinary Extracellular DNA in Acute Kidney Injury. Int. J. Mol. Sci. 2022, 23, 3402. https://doi.org/10.3390/ijms23063402
Jančuška A, Potočárová A, Kovalčíková AG, Podracká Ľ, Bábíčková J, Celec P, Tóthová Ľ. Dynamics of Plasma and Urinary Extracellular DNA in Acute Kidney Injury. International Journal of Molecular Sciences. 2022; 23(6):3402. https://doi.org/10.3390/ijms23063402
Chicago/Turabian StyleJančuška, Alexander, Alena Potočárová, Alexandra Gaál Kovalčíková, Ľudmila Podracká, Janka Bábíčková, Peter Celec, and Ľubomíra Tóthová. 2022. "Dynamics of Plasma and Urinary Extracellular DNA in Acute Kidney Injury" International Journal of Molecular Sciences 23, no. 6: 3402. https://doi.org/10.3390/ijms23063402
APA StyleJančuška, A., Potočárová, A., Kovalčíková, A. G., Podracká, Ľ., Bábíčková, J., Celec, P., & Tóthová, Ľ. (2022). Dynamics of Plasma and Urinary Extracellular DNA in Acute Kidney Injury. International Journal of Molecular Sciences, 23(6), 3402. https://doi.org/10.3390/ijms23063402