
Eugenol β-Amino/β-Alkoxy Alcohols with Selective Anticancer Activity

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
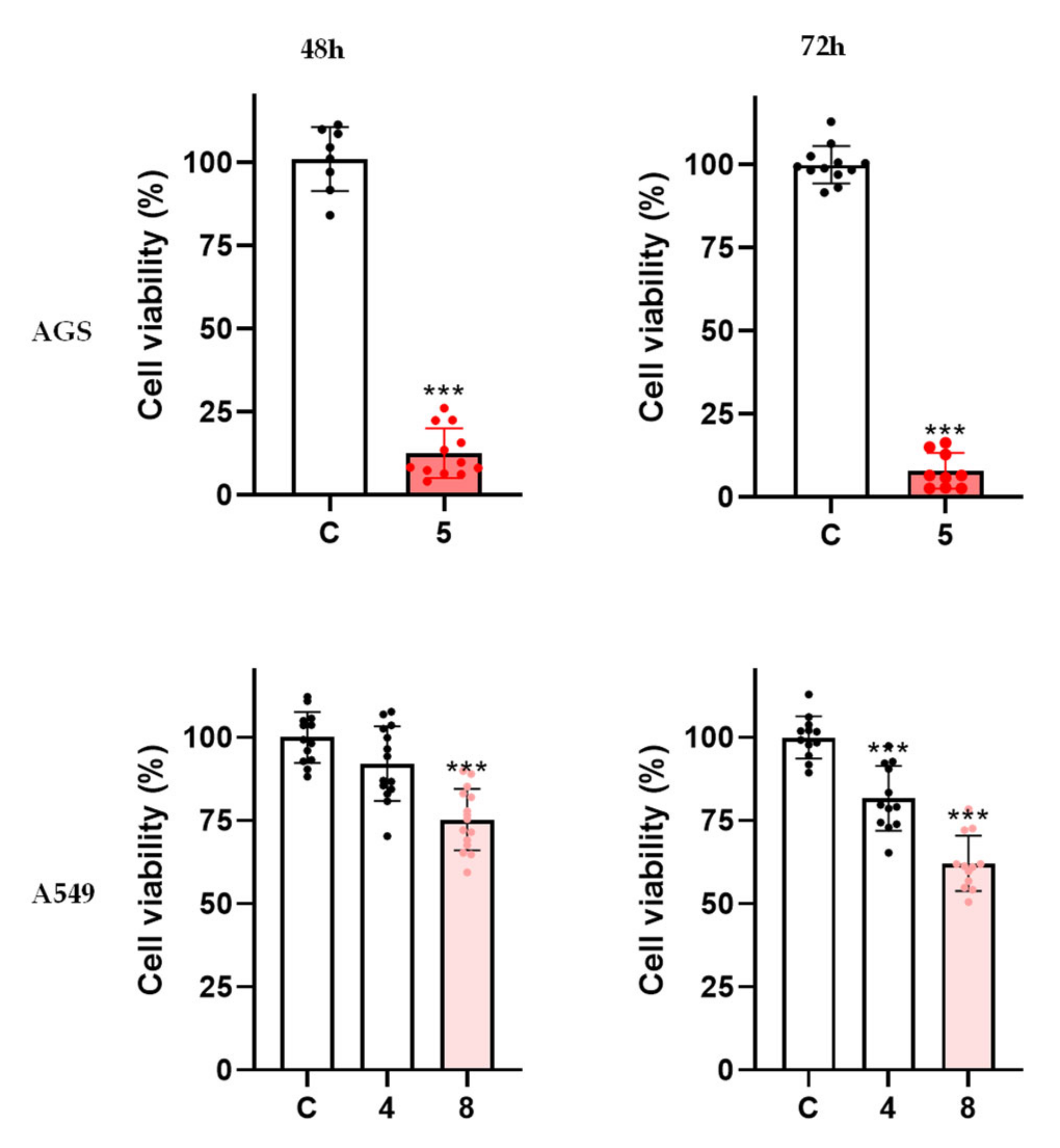
2. Results and Discussion
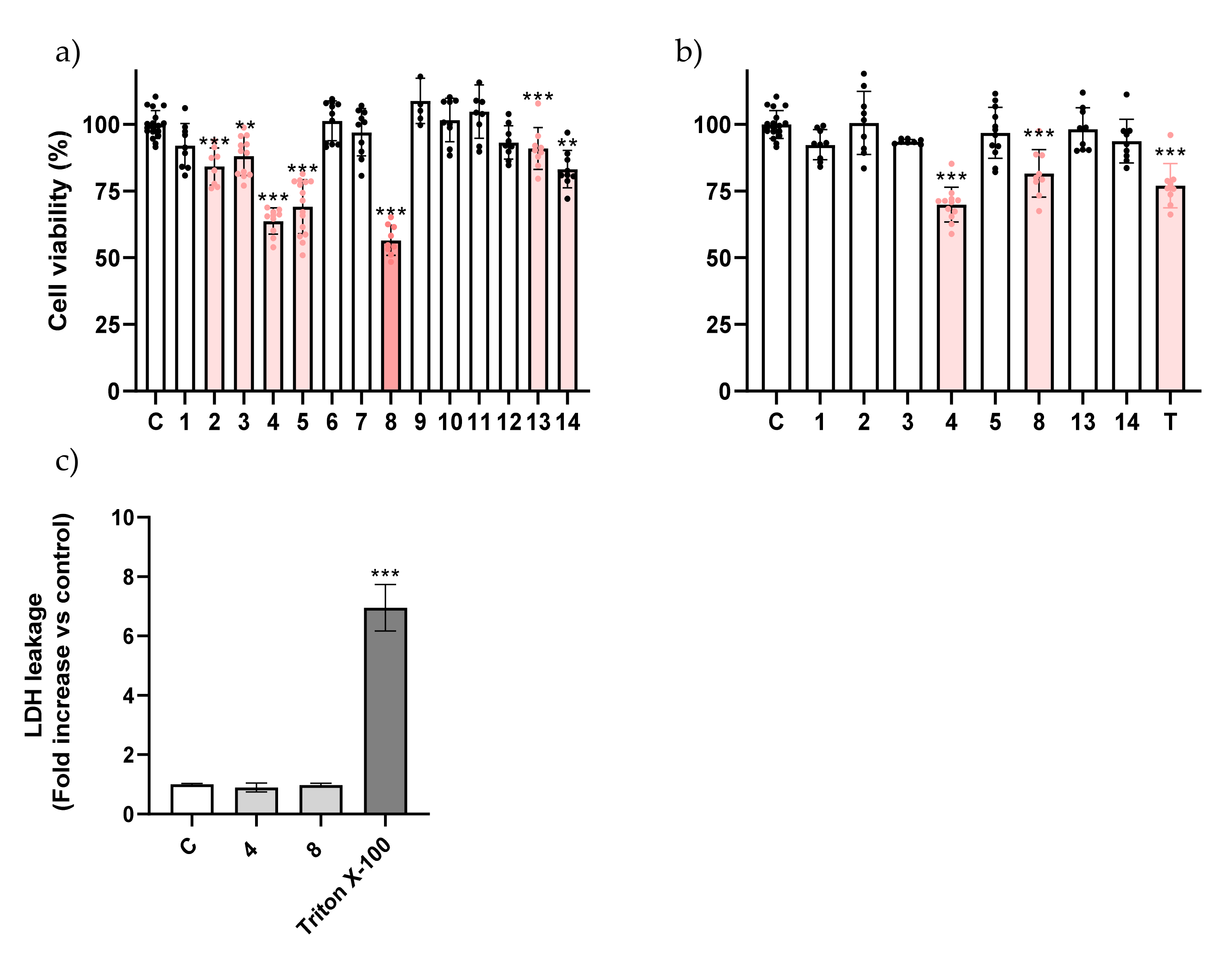
2.1. Eugenol β-Amino Alcohol Derivatives Are More Cytotoxic Than Their β-Alkoxy Alcohol Counterparts
2.2. The Toxicity of Eugenol Derivatives Is Selective for Cancer Cells
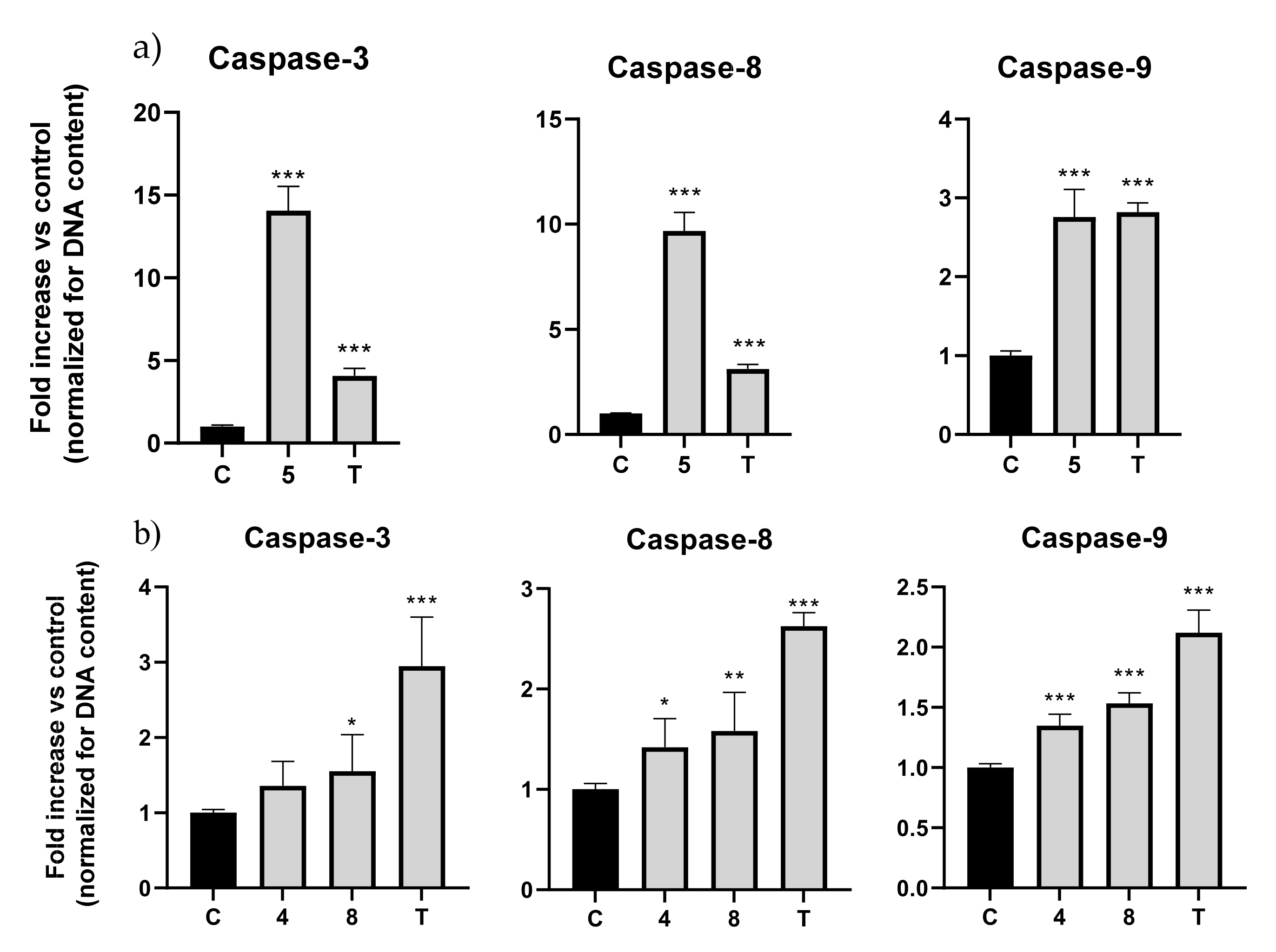
2.3. Eugenol β-Amino Alcohols Activate Several Caspase Isoforms
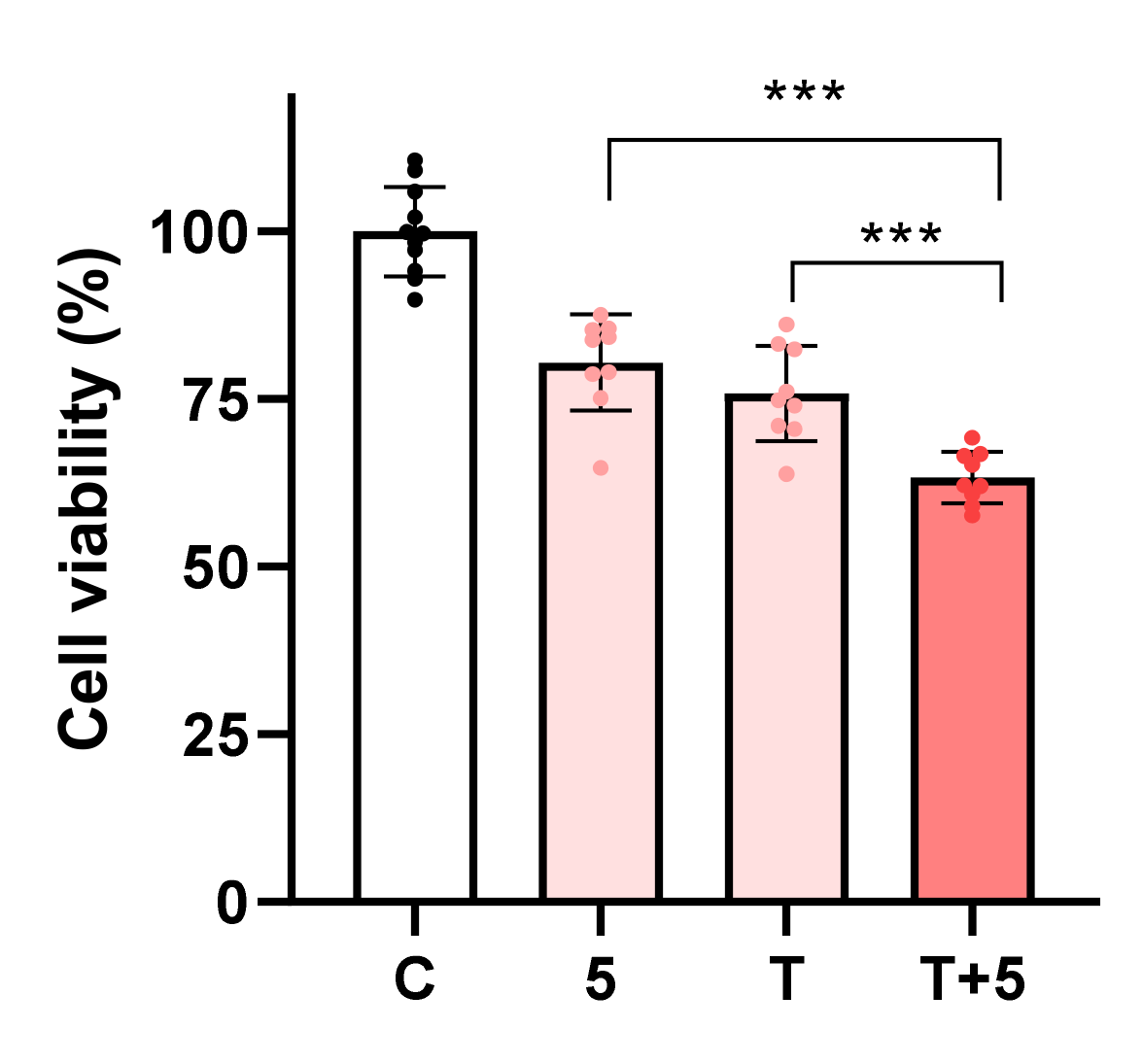
2.4. β-Amino Alcohol 5 Enhances the Toxicity of Topotecan
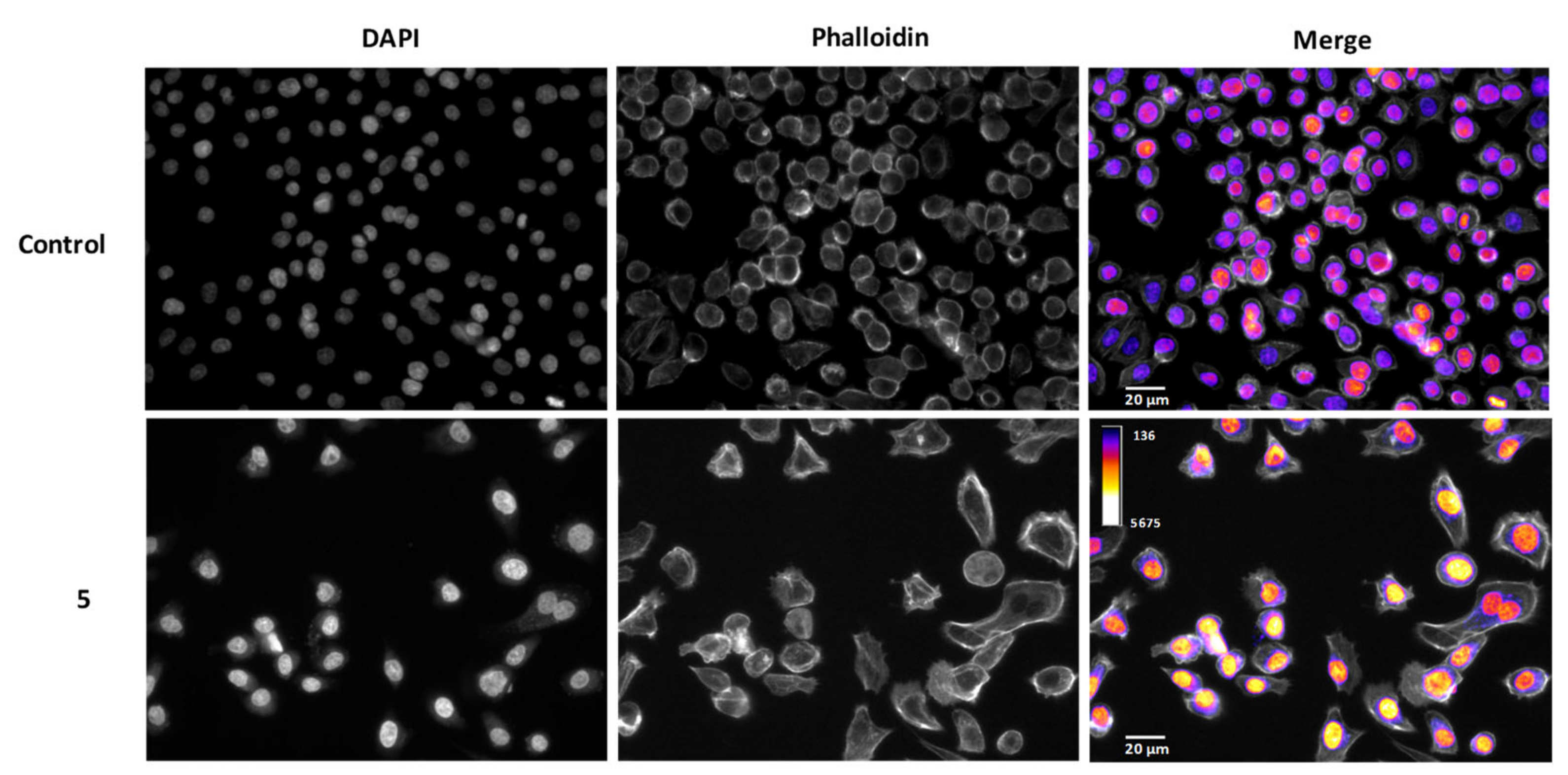
2.5. Derivatives 5 and 8, but Not 4, Have a Time-Dependent Effect
2.6. Structure-Activity Relationship
3. Materials and Methods
3.1. Reagents
3.2. Synthesis of Eugenol Derivatives
3.3. Cell Culture
3.4. Cell Viability Assessment
3.5. LDH Assay
3.6. Morphology Assement
3.7. Caspase Activity Assay
3.8. DNA Quantification
3.9. Chemometric Analysis
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 11 November 2021).
- Blowman, K.; Magalhaes, M.; Lemos, M.F.L.; Cabral, C.; Pires, I.M. Anticancer properties of essential oils and other natural products. Evid. Based Complement Altern. Med. 2018, 2018, 3149362. [Google Scholar] [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F.T.; Zhou, T.T.; Liu, B.; Bao, J.K. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Prolif. 2012, 45, 487–498. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Kaufman, T.S. The multiple faces of eugenol. A versatile starting material and building block for organic and bio-organic synthesis and a convenient precursor toward bio-based fine chemicals. J. Braz. Chem. Soc. 2015, 26, 1055–1085. [Google Scholar] [CrossRef]
- Baker, B.P.; Grant, J.A. Active Ingredients Eligible for Minimum Risk Pesticide Use: Overview of the Profiles; New York State IPM Program: Geneva, NY, USA, 2018. [Google Scholar]
- Barboza, J.N.; da Silva Maia Bezerra Filho, C.; Silva, R.O.; Medeiros, J.V.R.; de Sousa, D.P. An overview on the anti-inflammatory potential and antioxidant profile of eugenol. Oxid. Med. Cell Longev. 2018, 2018, 3957262. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, D.P.; Militao, G.C.G.; de Morais, M.C.; de Sousa, D.P. The dual antioxidant/prooxidant effect of eugenol and its action in cancer development and treatment. Nutrients 2017, 9, 1367. [Google Scholar] [CrossRef] [Green Version]
- Jaganathan, S.K.; Supriyanto, E. Antiproliferative and molecular mechanism of eugenol-induced apoptosis in cancer cells. Molecules 2012, 17, 6290–6304. [Google Scholar] [CrossRef]
- Sarkar, A.; Bhattacharjee, S.; Mandal, D.P. Induction of apoptosis by eugenol and capsaicin in human gastric cancer AGS cells--Elucidating the role of p53. Asian Pac. J. Cancer Prev. 2015, 16, 6753–6759. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, M.J.G.; Pereira, R.B.; Pereira, D.M.; Fortes, A.G.; Castanheira, E.M.S.; Gonçalves, M.S.T. New eugenol derivatives with enhanced insecticidal activity. Int. J. Mol. Sci. 2020, 21, 9257. [Google Scholar] [CrossRef]
- Carrasco, A.H.; Espinoza, C.L.; Cardile, V.; Gallardo, C.; Cardona, W.; Lombardo, L.; Catalán, M.K.; Cuellar, F.M.; Russo, A. Eugenol and its synthetic analogues inhibit cell growth of human cancer cells (Part I). J. Braz. Chem. Soc. 2008, 19, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, V.; Ferreres, F.; Macedo, T.; Gil-Izquierdo, A.; Oliveira, A.P.; Gomes, N.G.M.; Araujo, L.; Pereira, D.M.; Andrade, P.B.; Valentao, P. Activation of caspase-3 in gastric adenocarcinoma AGS cells by Xylopia aethiopica (Dunal) A. Rich. fruit and characterization of its phenolic fingerprint by HPLC-DAD-ESI(Ion Trap)-MS(n) and UPLC-ESI-QTOF-MS(2). Food Res. Int. 2021, 141, 110121. [Google Scholar] [CrossRef]
- Ngabire, D.; Seong, Y.A.; Patil, M.P.; Niyonizigiye, I.; Seo, Y.B.; Kim, G.D. Induction of apoptosis and G1 phase cell cycle arrest by Aster incisus in AGS gastric adenocarcinoma cells. Int. J. Oncol. 2018, 53, 2300–2308. [Google Scholar] [CrossRef] [Green Version]
- Videira, R.A.; Andrade, P.B.; Monteiro, L.S.; Valentão, P.; Ferreira, P.M.T.; Pereira, D.M. Toxicity and structure-activity relationship (SAR) of α,β-dehydroamino acids against human cancer cell lines. Toxicol. Vitr. 2018, 47, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollmannsberger, C.; Mross, K.; Jakob, A.; Kanz, L.; Bokemeyer, C. Topotecan—A novel topoisomerase I inhibitor: Pharmacology and clinical experience. Oncology 1999, 56, 1–12. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Hemaiswarya, S.; Doble, M. Combination of phenylpropanoids with 5-fluorouracil as anti-cancer agents against human cervical cancer (HeLa) cell line. Phytomedicine 2013, 20, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Fadilah, F.; Andrajati, R.; Arsianti, A.; Paramita, R.I.; Erlina, L.; Yanuar, A. Synthesis and in vitro activity of eugenyl benzoate derivatives as BCL-2 inhibitor in colorectal cancer with QSAR and molecular docking approach. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bendre, R.S.; Rajput, J.D.; Bagul, S.D.; Karandikar, P. Outlooks on medicinal properties of eugenol and its synthetic derivatives. Nat. Prod. Chem. Res. 2016, 4, 1000212. [Google Scholar] [CrossRef]
- He, L.; Jagtap, P.G.; Kingston, D.G.; Shen, H.J.; Orr, G.A.; Horwitz, S.B. A common pharmacophore for Taxol and the epothilones based on the biological activity of a taxane molecule lacking a C-13 side chain. Biochemistry 2000, 39, 3972–3978. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.F.M.; Monte, F.J.Q.; de Lemos, T.L.G.; do Nascimento, P.G.G.; de Medeiros Costa, A.K.; de Paiva, L.M.M. Eugenol derivatives: Synthesis, characterization, and evaluation of antibacterial and antioxidant activities. Chem. Cent. J. 2018, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, R.B.; Pinto, N.F.S.; Fernandes, M.J.G.; Vieira, T.F.; Rodrigues, A.R.O.; Pereira, D.M.; Sousa, S.F.; Castanheira, E.M.S.; Fortes, A.G.; Gonçalves, M.S.T. Amino alcohols from eugenol as potential semisynthetic insecticides: Chemical, biological, and computational insights. Molecules 2021, 26, 6616. [Google Scholar] [CrossRef] [PubMed]
- Coelho, C.; Pereira, R.; Vieira, T.; Teixeira, C.; Fernandes, M.; Rodrigues, A.; Pereira, D.; Sousa, S.; Fortes, A.; Castanheira, E.; et al. Synthesis, Computational and Nanoencapsulation Studies on Eugenol-Derived Insecticides. Preprints 2022. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 14 | 10 | 11 | 12 | 13 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Parent/Interm | β-Amino Alcohols | β-Alkoxy Alcohols | |||||||||||
| Molecular weight | 164.20 | 180.20 | 273.33 | 298.34 | 309.45 | 253.34 | 303.36 | 287.36 | 265.35 | 352.23 | 226.27 | 212.24 | 254.33 | 274.32 |
| cLogP | 2.27 | 1.21 | 2.17 | 2.01 | 3.60 | 1.61 | 2.10 | 2.42 | 1.86 | 2.90 | 1.34 | 0.94 | 2.08 | 2.45 |
| cLogS | −2.05 | −1.686 | −2.704 | −3.477 | −3.117 | −1.984 | −2.722 | −2.68 | −1.791 | −3.538 | −1.633 | −1.333 | −2.12 | −2.662 |
| H-Acceptors | 2 | 3 | 4 | 5 | 4 | 4 | 5 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| H-Donors | 1 | 1 | 3 | 3 | 3 | 3 | 3 | 2 | 2 | 3 | 2 | 2 | 2 | 2 |
| Total Surface Area | 140.17 | 146.04 | 219.46 | 241.17 | 265.52 | 206.89 | 241.72 | 230.56 | 213.84 | 238.09 | 184.26 | 170.5 | 205.43 | 218 |
| Relative PSA | 0.16 | 0.28 | 0.22 | 0.25 | 0.18 | 0.23 | 0.24 | 0.17 | 0.19 | 0.20 | 0.25 | 0.27 | 0.22 | 0.21 |
| Polar Surface Area | 29.46 | 41.99 | 61.72 | 85.51 | 61.72 | 61.72 | 70.95 | 52.93 | 52.93 | 61.72 | 58.92 | 58.92 | 58.92 | 58.92 |
| Druglikeness | −4.64 | −3.79 | 2.50 | −1.78 | −7.76 | 8.04 | 2.50 | 4.23 | 2.66 | 0.71 | −1.41 | −0.33 | −13.87 | −0.05 |
| Non-H Atoms | 12 | 13 | 20 | 22 | 22 | 18 | 22 | 21 | 19 | 21 | 16 | 15 | 18 | 20 |
| Non-C/H Atoms | 2 | 3 | 4 | 5 | 4 | 4 | 5 | 4 | 4 | 5 | 4 | 4 | 4 | 4 |
| Electronegative Atoms | 2 | 3 | 4 | 5 | 4 | 4 | 5 | 4 | 4 | 5 | 4 | 4 | 4 | 4 |
| Stereo Centers | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Rotatable Bonds | 3 | 3 | 6 | 6 | 11 | 6 | 7 | 6 | 5 | 6 | 6 | 5 | 6 | 6 |
| Aromatic Atoms | 6 | 6 | 12 | 12 | 6 | 6 | 12 | 12 | 6 | 12 | 6 | 6 | 6 | 12 |
| sp3-Atoms | 4 | 7 | 7 | 7 | 16 | 12 | 9 | 8 | 13 | 7 | 10 | 9 | 12 | 8 |
| Carbo-Rings | 1 | 1 | 2 | 2 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 1 | 1 | 2 |
| Hetero-Rings | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Non-Aromatic Rings | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Aromatic Rings | 1 | 1 | 2 | 2 | 1 | 1 | 2 | 2 | 1 | 2 | 1 | 1 | 1 | 2 |
| Amines | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| Alkyl-Amines | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Aromatic Amines | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| Basic Nitrogens | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| VDW-Surface | 188.95 | 192.07 | 280.22 | 301.71 | 343.78 | 273.89 | 309.8 | 291.99 | 271.35 | 299.96 | 243.92 | 220.22 | 269.69 | 274.62 |
| VDW-Volume | 174.15 | 179.9 | 280.28 | 300.96 | 347.12 | 276.4 | 307.53 | 301.51 | 281.39 | 299.68 | 233.78 | 215.05 | 273.74 | 273.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, C.; Pereira, R.B.; Pinto, N.F.S.; Coelho, C.M.M.; Fernandes, M.J.G.; Fortes, A.G.; Gonçalves, M.S.T.; Pereira, D.M. Eugenol β-Amino/β-Alkoxy Alcohols with Selective Anticancer Activity. Int. J. Mol. Sci. 2022, 23, 3759. https://doi.org/10.3390/ijms23073759
Teixeira C, Pereira RB, Pinto NFS, Coelho CMM, Fernandes MJG, Fortes AG, Gonçalves MST, Pereira DM. Eugenol β-Amino/β-Alkoxy Alcohols with Selective Anticancer Activity. International Journal of Molecular Sciences. 2022; 23(7):3759. https://doi.org/10.3390/ijms23073759
Chicago/Turabian StyleTeixeira, Cláudia, Renato B. Pereira, Nuno F. S. Pinto, Catarina M. M. Coelho, Maria José G. Fernandes, António G. Fortes, Maria S. T. Gonçalves, and David M. Pereira. 2022. "Eugenol β-Amino/β-Alkoxy Alcohols with Selective Anticancer Activity" International Journal of Molecular Sciences 23, no. 7: 3759. https://doi.org/10.3390/ijms23073759
APA StyleTeixeira, C., Pereira, R. B., Pinto, N. F. S., Coelho, C. M. M., Fernandes, M. J. G., Fortes, A. G., Gonçalves, M. S. T., & Pereira, D. M. (2022). Eugenol β-Amino/β-Alkoxy Alcohols with Selective Anticancer Activity. International Journal of Molecular Sciences, 23(7), 3759. https://doi.org/10.3390/ijms23073759