
Di-(2-ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial–Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway

 ,
, 
Abstract
:1. Introduction
2. Results
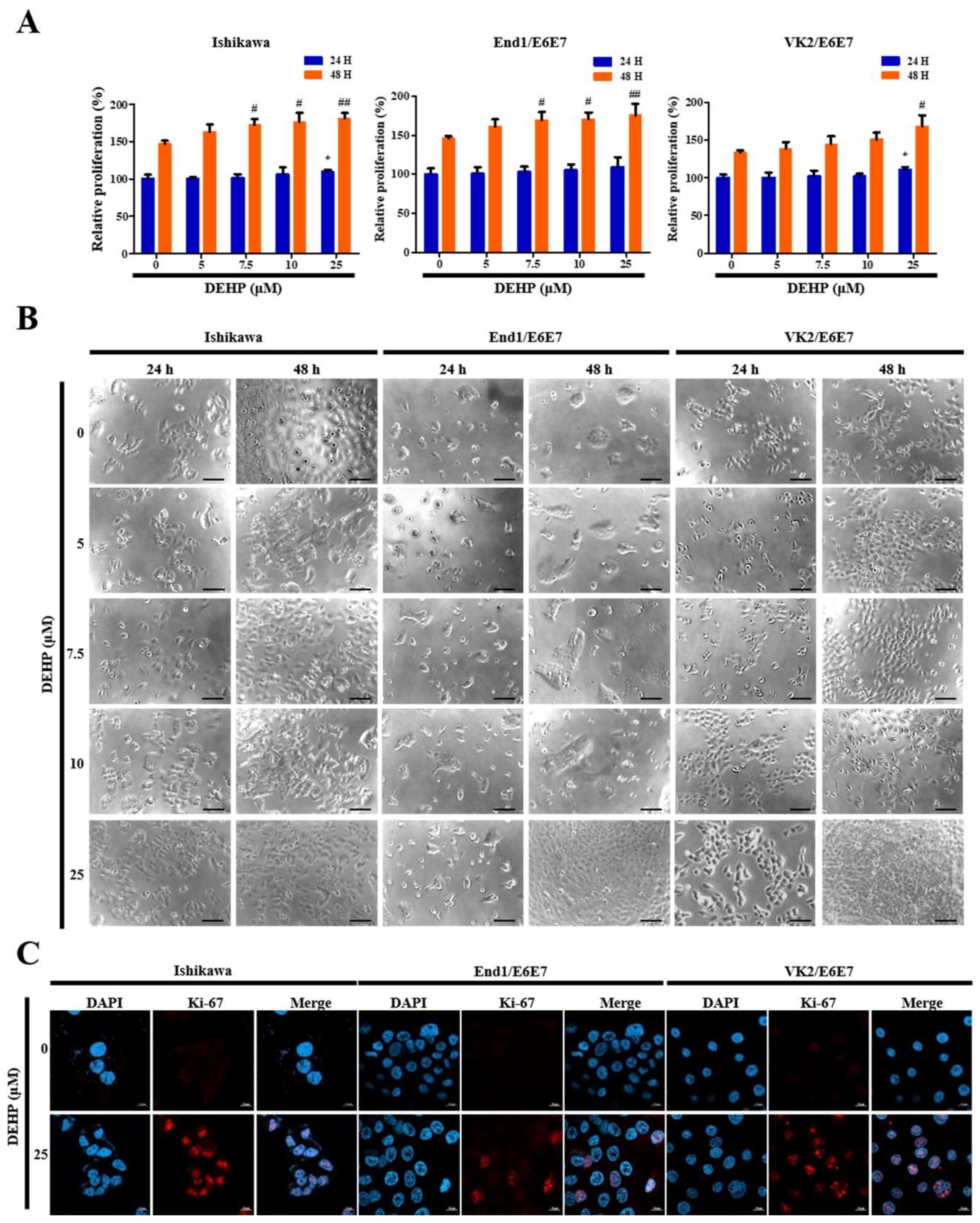
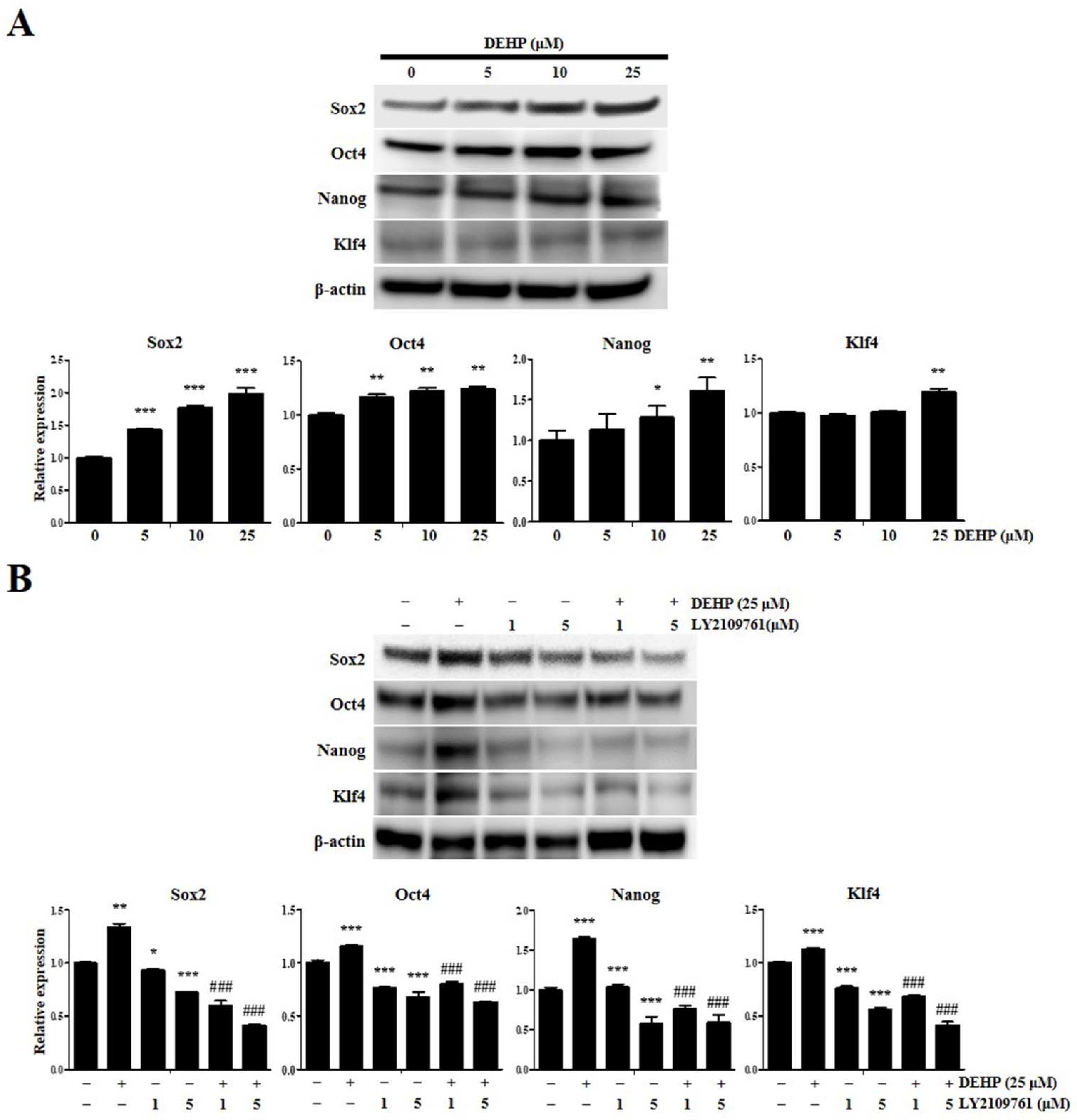
2.1. DEHP Promotes Human EEECs Proliferation
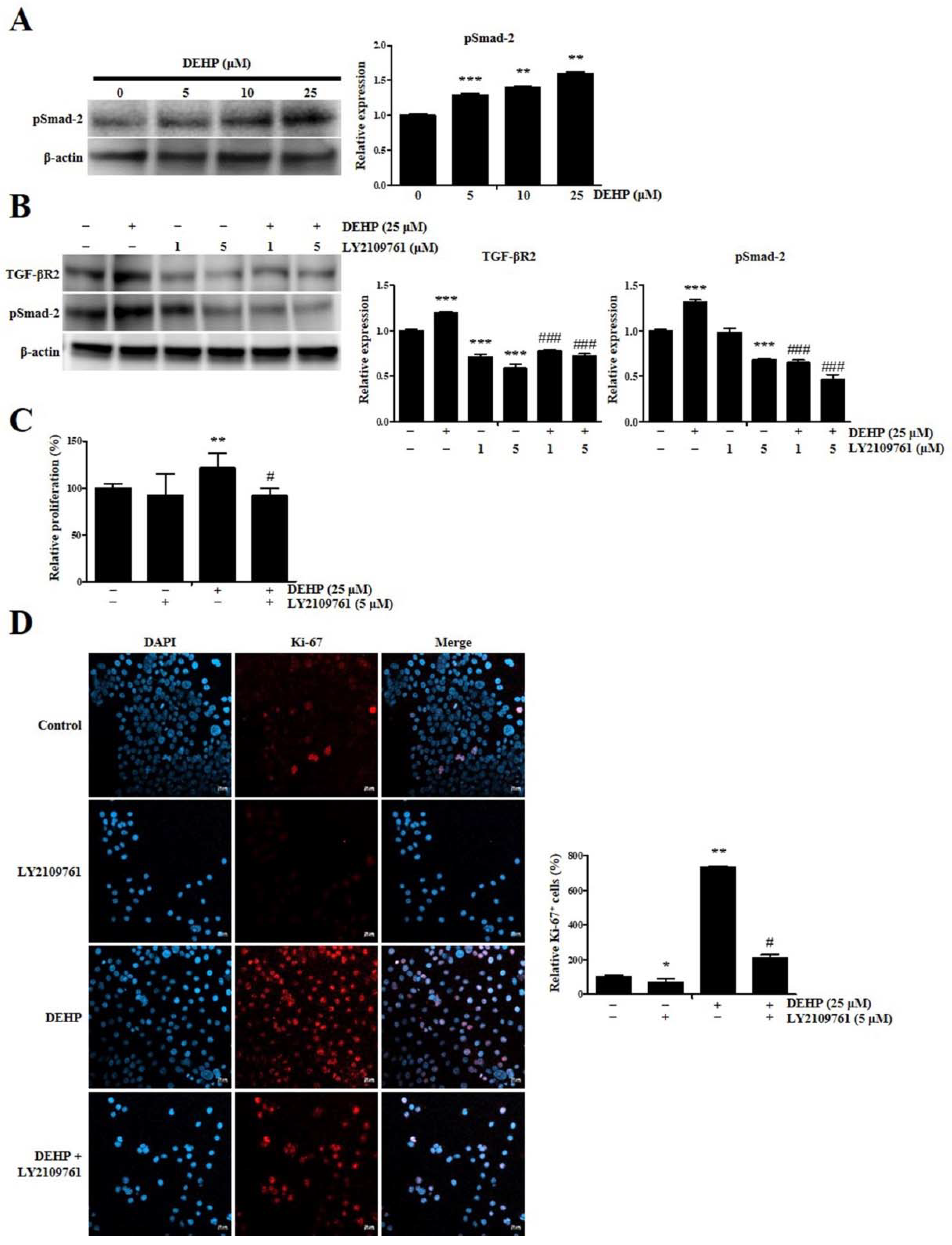
2.2. DEHP Enhances Human Endometrial Epithelial Cell Proliferation through TGF-β Signaling Pathway
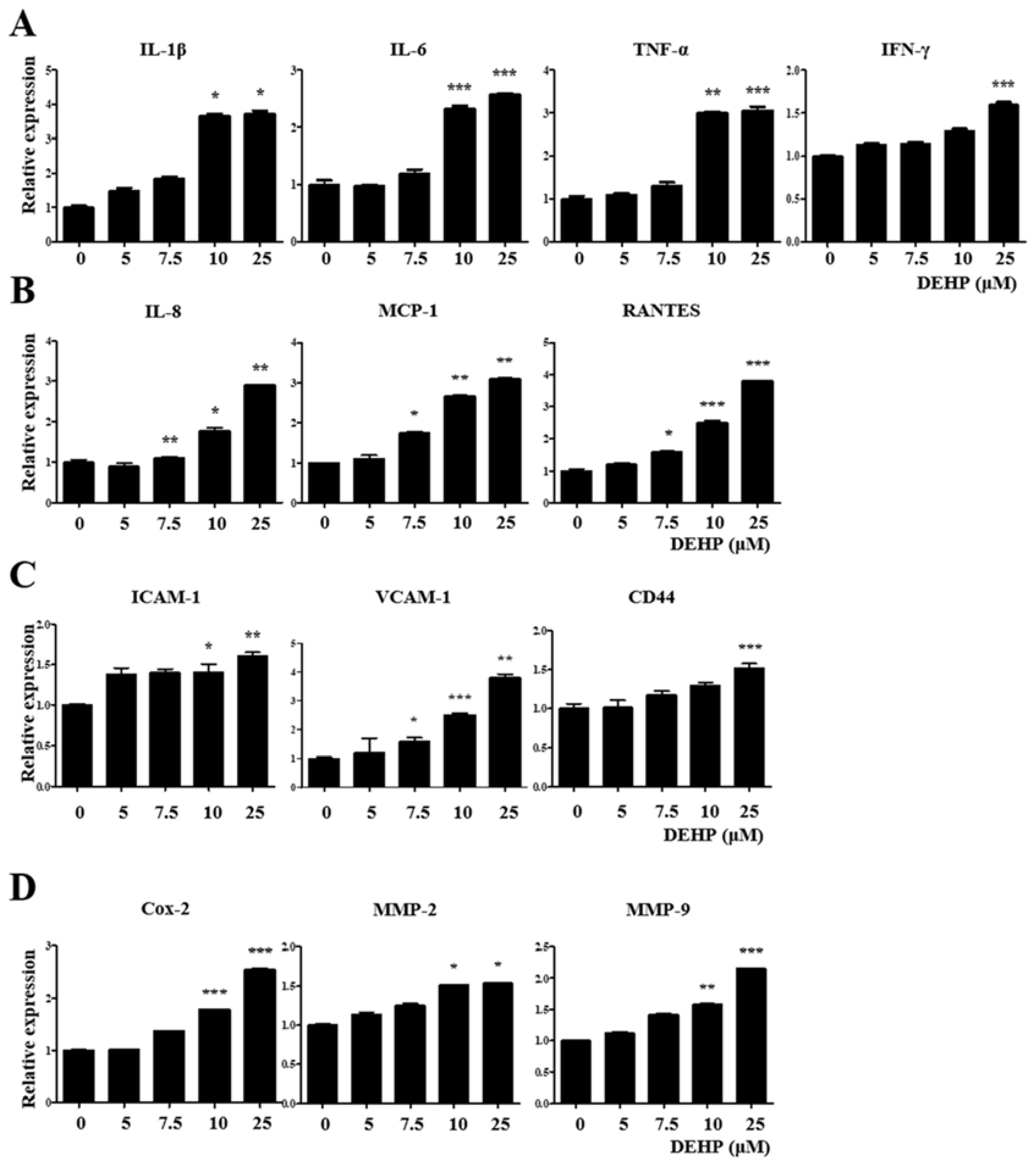
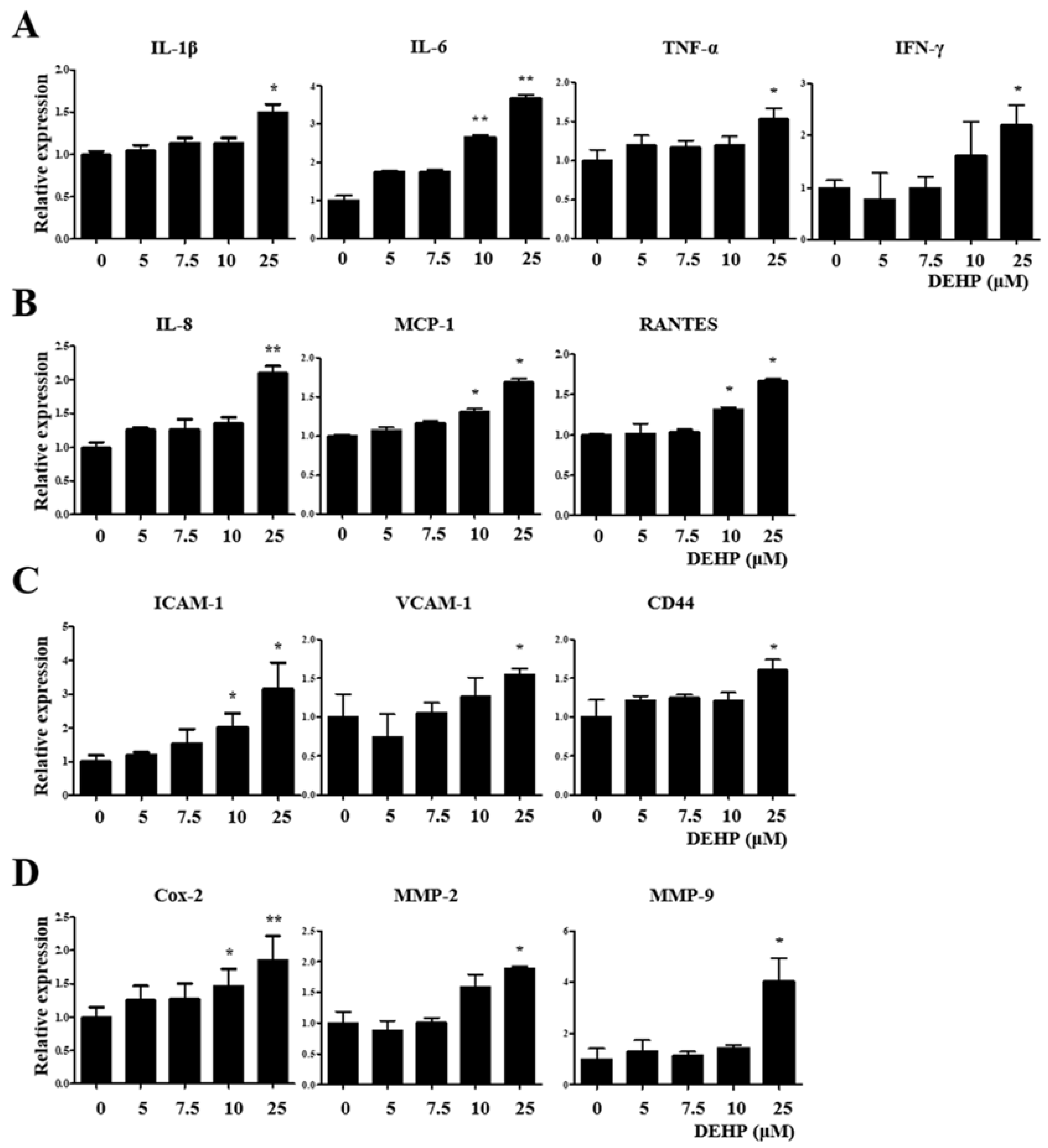
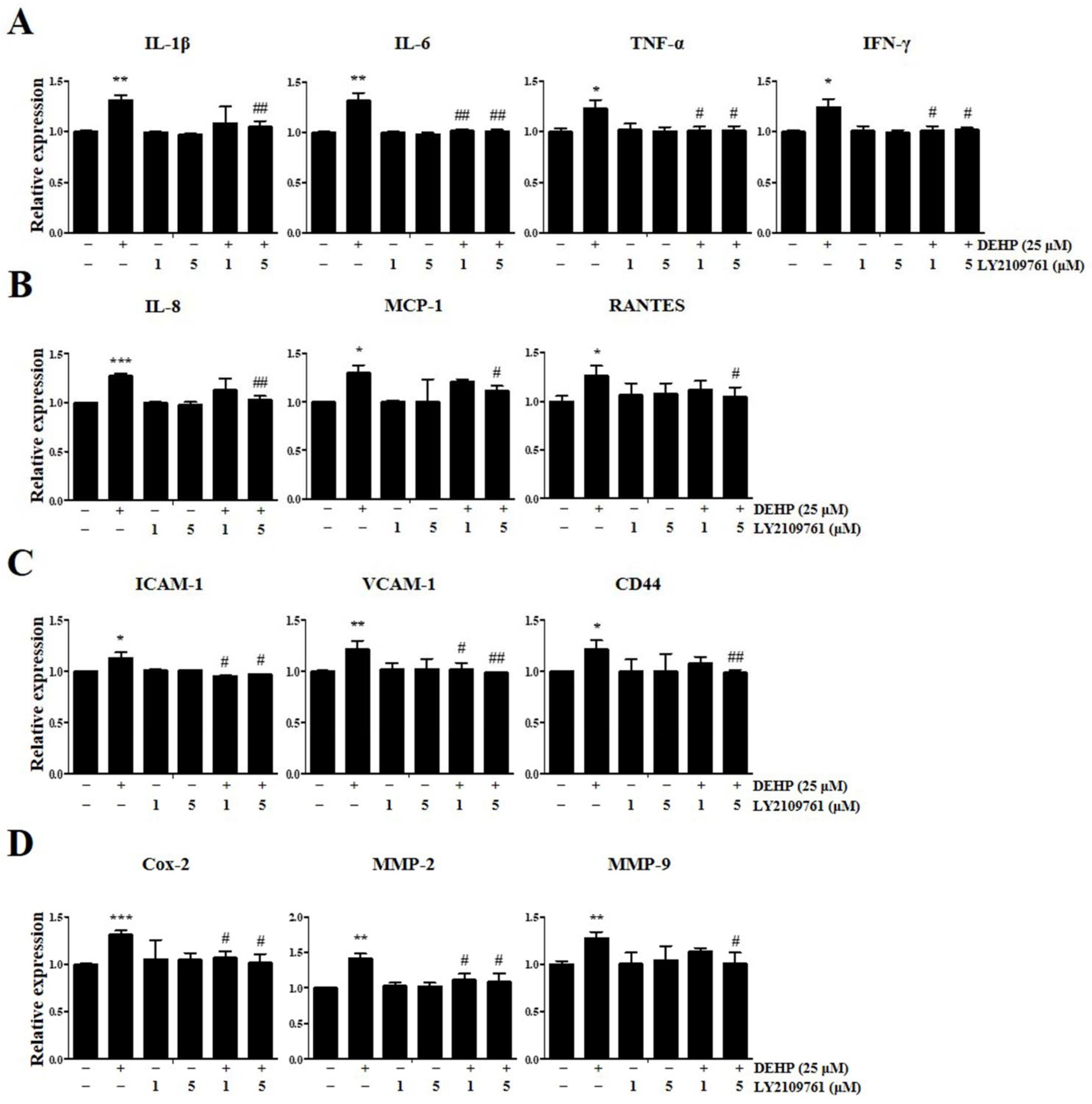
2.3. DEHP Triggers Inflammatory and Immunological Responses through the TGF-β Signaling Pathway in Human EEECs
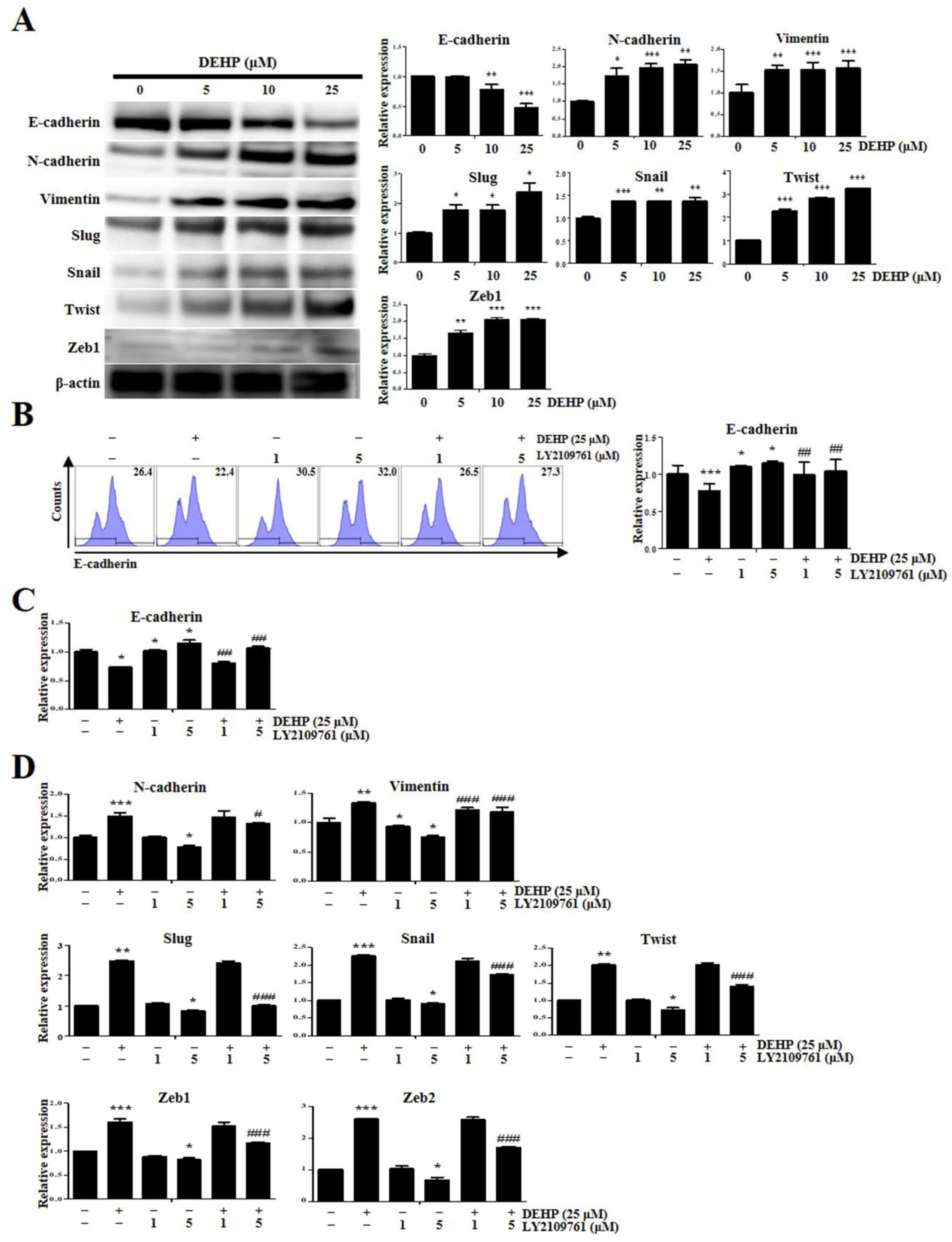
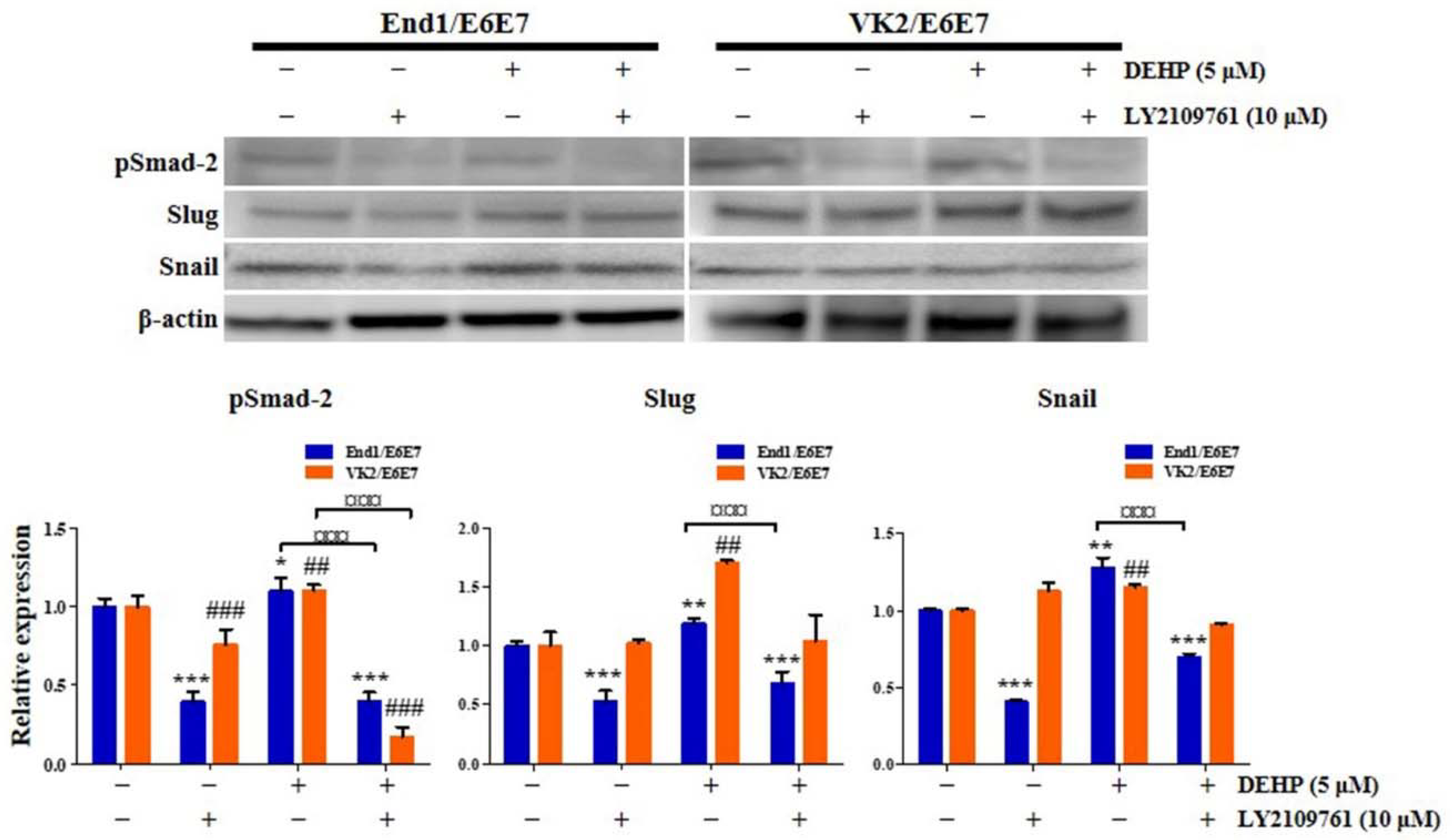
2.4. DEHP Mediates EMT through TGF-β Signaling Pathway in Human Endometrial Epithelial Cells
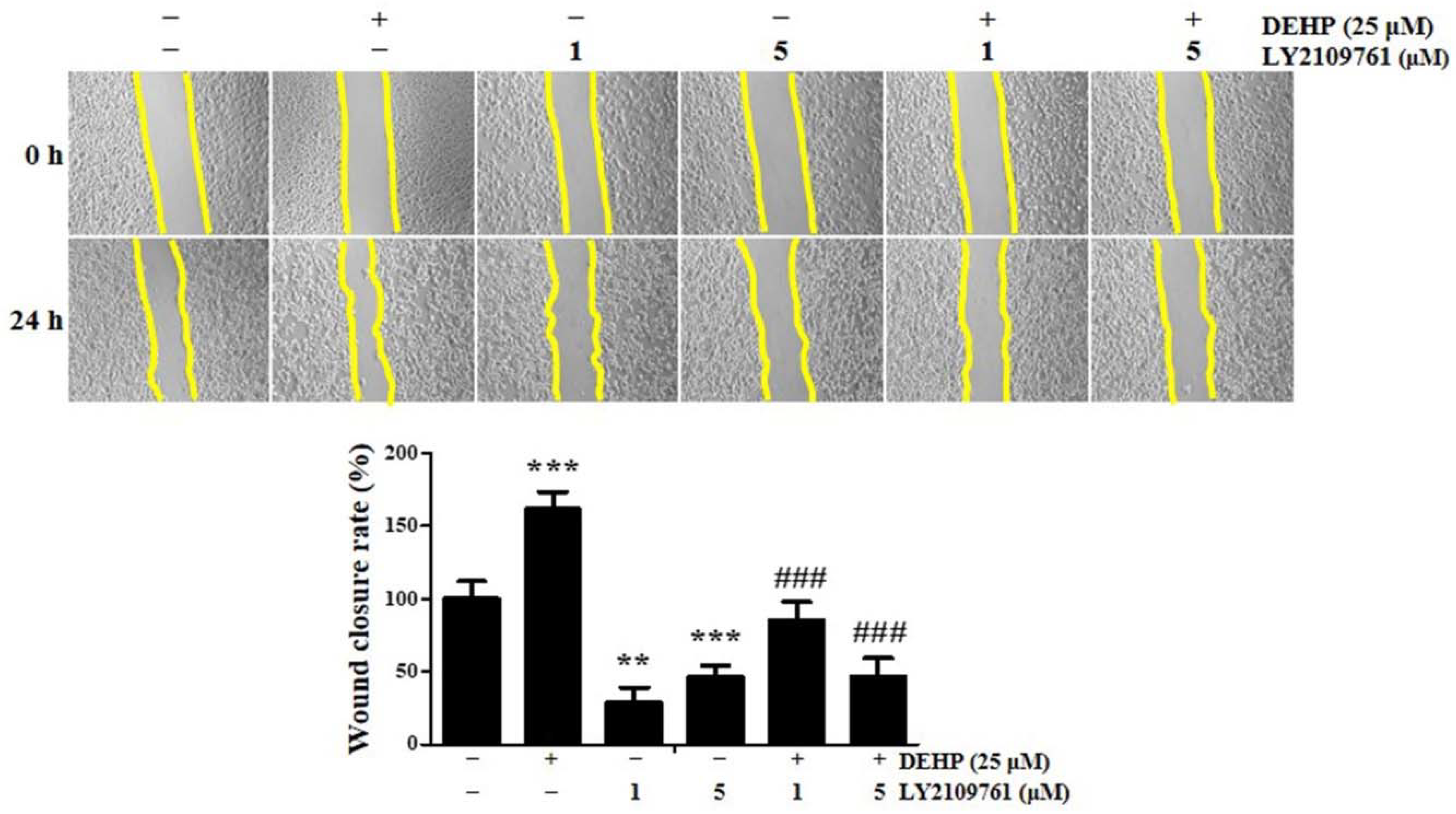
2.5. TGF-β Signaling Is Essential in DEHP-Induced Human Endometrial Epithelial Cell Migration
2.6. DEHP Augments Human Endometrial Epithelial Cell Stemness Traits through TGF-β Signaling Pathway
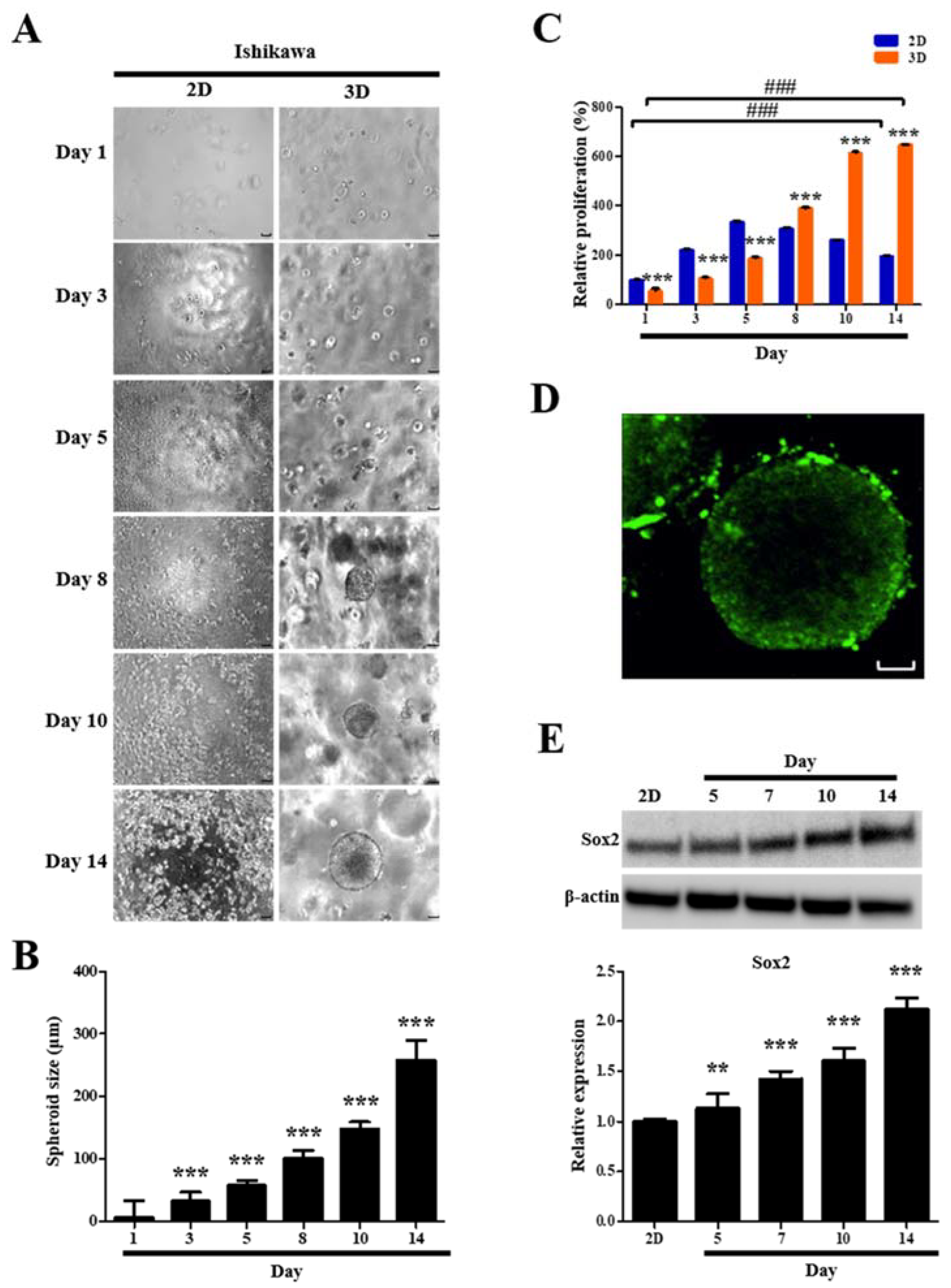
2.7. Formation and Growth of Human Endometrial Epithelial Cell Spheroids
2.8. Proliferation and Viability of Human Endometrial Epithelial Cells Are Facilitated after 3D Culture
2.9. Stemness Marker Expression of Human Endometrial Epithelial Cells Is Upregulated after 3D Culture
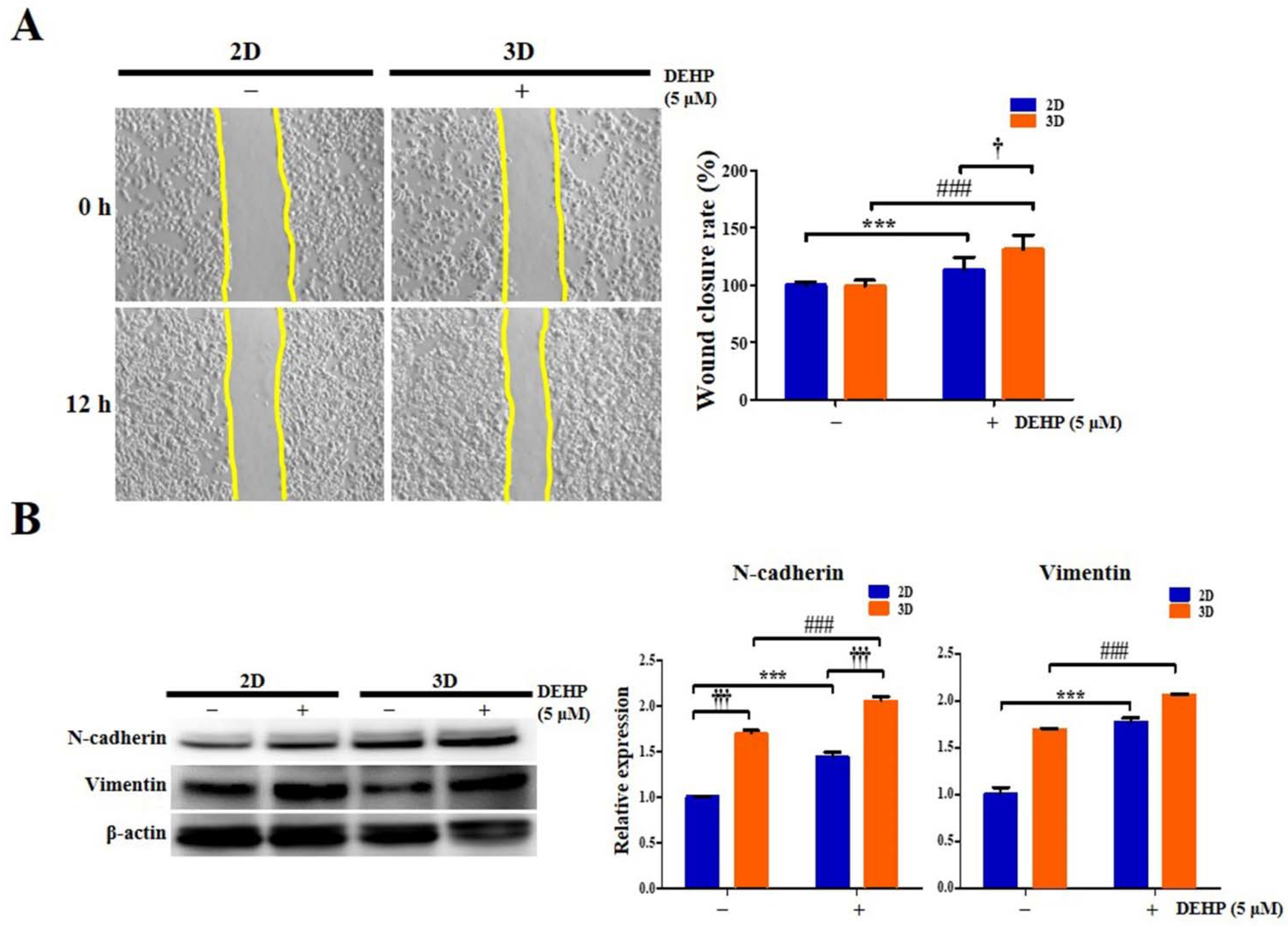
2.10. Migratory Potential of Human Endometrial Epithelial Cells Is Elevated after DEHP Treatment in 3D Culture
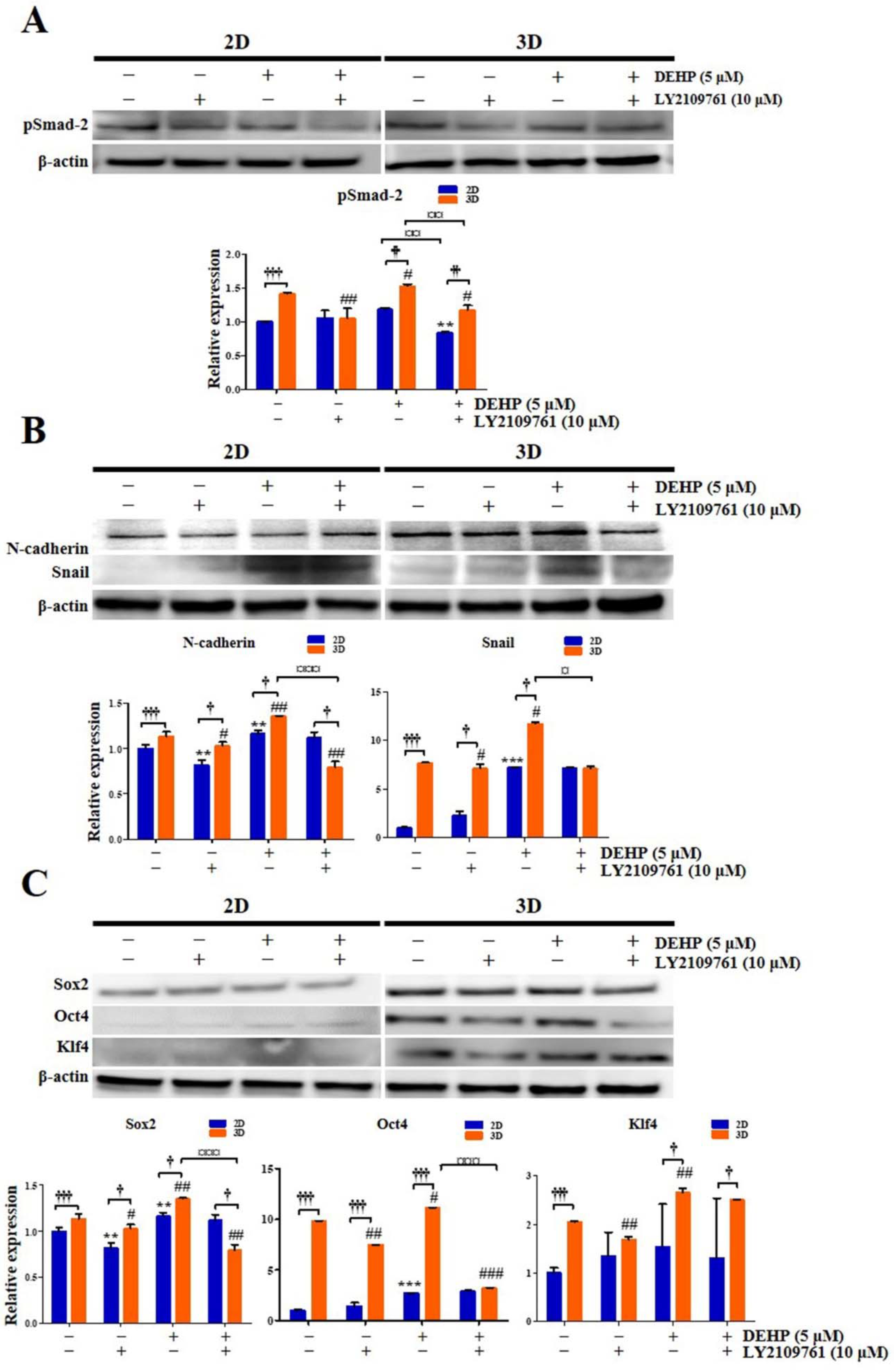
2.11. DEHP Augments EMT and Stemness through TGF-β Signaling Pathway in 3D-Cultured Endometrial Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Hydrogel Synthesis for 3D Cell Culture
4.3. Cell Proliferation Assay
4.4. Spheroid Growth Assay
4.5. Evaluation of Immunofluorescence Using Confocal Microscopy
4.6. Wound Healing Assay
4.7. RNA Isolation and cDNA Synthesis
4.8. Quantitative Real-Time PCR (qRT-PCR)
4.9. Western Blot Analysis
4.10. Flow Cytometry
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DEHP | di-(2-ethylhexyl) phthalate |
| EDCs | endocrine-disrupting chemicals |
| EMT | epithelial–mesenchymal transition |
| PAEs | phthalate esters |
| TGF-β | transforming growth factor-β |
| EEECs | endometrial and endometriotic epithelial cells |
References
- Lyche, J.L.; Gutleb, A.C.; Bergman, A.; Eriksen, G.S.; Murk, A.J.; Ropstad, E.; Saunders, M.; Skaare, J.U. Reproductive and developmental toxicity of phthalates. J. Toxicol. Environ. Health B Crit. Rev. 2009, 12, 225–249. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, H.; Kannan, K. A review of biomonitoring of phthalate exposures. Toxics 2019, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Rowdhwal, S.S.S.; Chen, J. Toxic effects of di-2-ethylhexyl phthalate: An overview. BioMed Res. Int. 2018, 2018, 1750368. [Google Scholar] [CrossRef]
- Kahn, L.G.; Philippat, C.; Nakayama, S.F.; Slama, R.; Trasande, L. Endocrine-disrupting chemicals: Implications for human health. Lancet Diabetes Endocrinol. 2020, 8, 703–718. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, S.H.; Lee, H.W.; Chae, H.D.; Kim, C.H.; Kang, B.M. Increased viability of endometrial cells by in vitro treatment with di-(2-ethylhexyl) phthalate. Fertil. Steril. 2010, 94, 2413–2416. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.H. Exposure to phthalate esters and the risk of endometriosis. Dev. Reprod. 2020, 24, 71–78. [Google Scholar] [CrossRef]
- Liu, X.; He, D.W.; Zhang, D.Y.; Lin, T.; Wei, G.H. Di(2-ethylhexyl) phthalate (DEHP) increases transforming growth factor-beta1 expression in fetal mouse genital tubercles. J. Toxicol. Environ. Health A 2008, 71, 1289–1294. [Google Scholar] [CrossRef]
- Zhao, Z.B.; Ji, K.; Shen, X.Y.; Zhang, W.W.; Wang, R.; Xu, W.P.; Wei, W. Di(2-ethylhexyl) phthalate promotes hepatic fibrosis by regulation of oxidative stress and inflammation responses in rats. Environ. Toxicol. Pharmacol. 2019, 68, 109–119. [Google Scholar] [CrossRef]
- Fan, P.; Li, Z.; Zuo, C.; Fang, M. Promotion effects of mono-2-ethyhexyl phthalate (MEHP) on migration and invasion of human melanoma cells via activation of TGF-β signals. Cell Biochem. Funct. 2020, 38, 38–46. [Google Scholar] [CrossRef]
- Arafah, M.; Rashid, S.; Akhtar, M. Endometriosis: A Comprehensive Review. Adv. Anat. Pathol. 2021, 28, 30–43. [Google Scholar] [CrossRef]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The endometrial immune environment of women with endometriosis. Hum. Reprod. Update 2019, 25, 565–592. [Google Scholar] [CrossRef]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Young, V.J.; Brown, J.K.; Saunders, P.T.; Horne, A.W. The role of the peritoneum in the pathogenesis of endometriosis. Hum. Reprod. Update 2013, 19, 558–569. [Google Scholar] [CrossRef]
- Koninckx, P.; Ussia, A.; Adamyan, L.; Wattiez, A.; Gomel, V.; Martin, D. Pathogenesis of endometriosis: The genetic/epigenetic theory. Fertil. Steril. 2019, 111, 327–340. [Google Scholar] [CrossRef]
- Gargett, C.; Schwab, K.E.; Brosens, J.J.; Puttemans, P.; Benagiano, G.; Brosens, I. Potential role of endometrial stem/progenitor cells in the pathogenesis of early-onset endometriosis. Mol. Hum. Reprod. 2014, 20, 591–598. [Google Scholar] [CrossRef]
- Leyendecker, G.; Kunz, G.; Herbertz, M.; Beil, D.; Huppert, P.; Mall, G.; Kissler, S.; Noe, M.; Wildt, L. Uterine peristaltic activity and the development of endometriosis. Ann. N. Y. Acad. Sci. 2004, 1034, 338–355. [Google Scholar] [CrossRef]
- Viganó, D.; Zara, F.; Pinto, S.; Loddo, E.; Casula, L.; Soru, M.B.; D’Ancona, G.; D’Alterio, M.N.; Giuliani, C.; Angioni, S.; et al. How is small bowel permeability in endometriosis patients? a case control pilot study. Gynecol. Endocrinol. 2020, 36, 1010–1014. [Google Scholar] [CrossRef]
- D’Alterio, M.N.; Giuliani, C.; Scicchitano, F.; Laganà, A.S.; Oltolina, N.M.; Sorrentino, F.; Nappi, L.; Orru, G.; Angioni, S. Possible role of microbiome in the pathogenesis of endometriosis. Minerva Obstet. Gynecol. 2021, 73, 193–214. [Google Scholar] [CrossRef]
- Matsuzaki, S.; Darcha, C. Epithelial to mesenchymal transition-like and mesenchymal to epithelial transition-like processes might be involved in the pathogenesis of pelvic endometriosis. Hum. Reprod. 2012, 27, 712–721. [Google Scholar] [CrossRef]
- Konrad, L.; Dietze, R.; Riaz, M.A.; Scheiner-Bobis, G.; Behnke, J.; Horné, F.; Hoerscher, A.; Reising, C.; Meinhold-Heerlein, I. Epithelial–mesenchymal transition in endometriosis—when does it happen? J. Clin. Med. 2020, 9, 1915. [Google Scholar] [CrossRef]
- Pradella, D.; Naro, C.; Sette, C.; Ghigna, C. EMT and stemness: Flexible processes tuned by alternative splicing in development and cancer progression. Mol. Cancer 2017, 16, 8. [Google Scholar] [CrossRef]
- Owusu-Akyaw, A.; Krishnamoorthy, K.; Goldsmith, L.T.; Morelli, S.S. The role of mesenchymal-epithelial transition in endometrial function. Hum. Reprod. Update 2019, 25, 114–133. [Google Scholar] [CrossRef]
- Pei, D.; Shu, X.; Gassama-Diagne, A.; Thiery, J.P. Mesenchymal–epithelial transition in development and reprogramming. Nat. Cell. Biol. 2019, 21, 44–53. [Google Scholar] [CrossRef]
- Bich, L.; Pradeu, T.; Moreau, J.F. Understanding multicellularity: The functional organization of the intercellular space. Front. Physiol. 2019, 10, 1170. [Google Scholar] [CrossRef]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef]
- Brueggmann, D.; Templeman, C.; Starzinski-Powitz, A.; Rao, N.P.; Gayther, S.A.; Lawrenson, K. Novel three-dimensional in vitro models of ovarian endometriosis. J. Ovarian Res. 2014, 7, 17. [Google Scholar] [CrossRef]
- Wendel, J.R.H.; Wang, X.; Smith, L.J.; Hawkins, S.M. Three-dimensional biofabrication models of endometriosis and the endometriotic microenvironment. Biomedicines 2020, 8, 525. [Google Scholar] [CrossRef]
- Zhu, R.; Nasu, K.; Aoyagi, Y.; Hirakawa, T.; Takebayashi, K.; Narahara, H. Chemokine expression profiles of ovarian endometriotic stromal cells in three-dimensional culture. J. Reprod. Immunol. 2020, 138, 103100. [Google Scholar] [CrossRef]
- Trepat, X.; Chen, Z.; Jacobson, K. Cell migration. Compr. Physiol. 2012, 2, 2369–2392. [Google Scholar] [CrossRef]
- Chang, W.H.; Herianto, S.; Lee, C.C.; Hung, H.; Chen, H.L. The effects of phthalate ester exposure on human health: A review. Sci. Total Environ. 2021, 786, 147371. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Zhang, H.; Chen, Y.J.; Chi, Y.L.; Dong, S. The inflammation response to DEHP through PPARγ in endometrial cells. Int. J. Environ. Res. Public Health 2016, 13, 318. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Yan, Y.; Liu, Z.; Wang, Y. Inflammation and endometriosis. Front. Biosci. 2016, 21, 941–948. [Google Scholar] [CrossRef]
- Ahn, S.H.; Monsanto, S.P.; Miller, C.; Singh, S.S.; Thomas, R.; Tayade, C. Pathophysiology and immune dysfunction in endometriosis. Biomed Res. Int. 2015, 2015, 795976. [Google Scholar] [CrossRef]
- Kotlyar, A.; Taylor, H.S.; D’Hooghe, T.M. Use of immunomodulators to treat endometriosis. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 60, 56–65. [Google Scholar] [CrossRef]
- Stadnyk, A.W. Cytokine production by epithelial cells. FASEB J. 1994, 8, 1041–1047. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Abrao, M.S.; Mechsner, S. Can chemokines be used as biomarkers for endometriosis? A systematic review. Hum. Reprod. 2014, 29, 253–266. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, E.N.; Park, J.K.; Lee, J.R.; Kim, J.H.; Choi, H.J.; Kim, B.S.; Lee, H.W.; Yoon, S. Curcumin attenuates TNF-alpha-induced expression of intercellular adhesion molecule-1, vascular cell adhesion molecule-1 and proinflammatory cytokines in human endometriotic stromal cells. Phytother. Res. 2012, 26, 1037–1047. [Google Scholar] [CrossRef]
- Laganà, A.S.; Garzon, S.; Gotte, M.; Vigano, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The pathogenesis of endometriosis: Molecular and cell biology insights. Int. J. Mol. Sci. 2019, 20, 5615. [Google Scholar] [CrossRef]
- Lai, Z.Z.; Yang, H.L.; Ha, S.Y.; Chang, K.K.; Mei, J.; Zhou, W.J.; Qiu, X.M.; Wang, X.Q.; Zhu, R.; Li, D.J.; et al. Cyclooxygenase-2 in endometriosis. Int. J. Biol. Sci. 2019, 15, 2783–2797. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, R.; Jiang, Q.; Li, Z.; Wu, R. Expression of cellular adherent and invasive molecules in recurrent ovarian endometriosis. J. Int. Med. Res. 2020, 48, 300060520971993. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, J.; Iwahara, C.; Kawasaki, M.; Yoshizaki, F.; Nakayama, H.; Takamori, K.; Ogawa, H.; Iwabuchi, K. Di-(2-ethylhexyl) phthalate induces production of inflammatory molecules in human macrophages. Inflamm. Res. 2012, 61, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Ma, K.; Na, X. Rosmarinic acid alleviates di-2-ethylhexyl phthalate (DEHP) -induced thyroid dysfunction via multiple inflammasomes activation. J. Toxicol. Sci. 2020, 45, 373–390. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H. Glucagon-like peptide-1 receptor agonist reduces di(2-ethylhexyl) phthalate-induced atherosclerotic processes in vascular smooth muscle cells. Physiol. Res. 2020, 69, 1095–1102. [Google Scholar] [CrossRef]
- Proestling, K.; Birner, P.; Gamperl, S.; Nirtl, N.; Marton, E.; Yerlikaya, G.; Wenzl, R.; Streubel, B.; Husslein, H. Enhanced epithelial to mesenchymal transition (EMT) and upregulated MYC in ectopic lesions contribute independently to endometriosis. Reprod. Biol. Endocrinol. 2015, 13, 75. [Google Scholar] [CrossRef]
- López-Novoa, J.M.; Nieto, M.A. Inflammation and EMT: An alliance towards organ fibrosis and cancer progression. EMBO Mol. Med. 2009, 1, 303–314. [Google Scholar] [CrossRef]
- Katsuno, Y.; Lamouille, S.; Derynck, R. TGF-beta signaling and epithelial-mesenchymal transition in cancer progression. Curr. Opin. Oncol. 2013, 25, 76–84. [Google Scholar] [CrossRef]
- Audebert, A.; Petousis, S.; Margioula-Siarkou, C.; Ravanos, K.; Prapas, N.; Prapas, Y. Anatomic distribution of endometriosis: A reappraisal based on series of 1101 patients. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 230, 36–40. [Google Scholar] [CrossRef]
- Xu, X.; Zheng, L.; Yuan, Q.; Zhen, G.; Crane, J.L.; Zhou, X.; Cao, X. Transforming growth factor-β in stem cells and tissue homeostasis. Bone Res. 2018, 6, 2. [Google Scholar] [CrossRef]
- Shariati, F.; Favaedi, R.; Ramazanali, F.; Ghoraeian, P.; Afsharian, P.; Aflatoonian, B.; Aflatoonian, R.; Shahhoseini, M. Increased expression of stemness genes REX-1, OCT-4, NANOG, and SOX-2 in women with ovarian endometriosis versus normal endometrium: A case-control study. Int. J. Reprod. Biomed. 2018, 16, 783–790. [Google Scholar] [CrossRef]
- Kong, Y.; Shao, Y.; Ren, C.; Yang, G. Endometrial stem/progenitor cells and their roles in immunity, clinical application, and endometriosis. Stem Cell Res. Ther. 2021, 12, 474. [Google Scholar] [CrossRef] [PubMed]
- Verdi, J.; Tan, A.; Shoae-Hassani, A.; Seifalian, A.M. Endometrial stem cells in regenerative medicine. J. Biol. Eng. 2014, 8, 20. [Google Scholar] [CrossRef]
- Wilson, M.M.; Weinberg, R.A.; Lees, J.A.; Guen, V.J. Emerging mechanisms by which EMT programs control stemness. Trends Cancer 2020, 6, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.P.T.; Xiao, L.; Deane, J.A.; Tan, K.S.; Cousins, F.L.; Masuda, H.; Sprung, C.N.; Rosamilia, A.; Gargett, C.E. N-cadherin identifies human endometrial epithelial progenitor cells by in vitro stem cell assays. Hum. Reprod. 2017, 32, 2254–2268. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, F. Stem cell and endometriosis: New knowledge may be producing novel therapies. Int. J. Clin. Exp. Med. 2014, 7, 3853–3858. [Google Scholar] [PubMed]
- Baranov, V.; Malysheva, O.; Yarmolinskaya, M. Pathogenomics of endometriosis development. Int. J. Mol. Sci. 2018, 19, 1852. [Google Scholar] [CrossRef]
- Au, H.K.; Chang, J.H.; Wu, Y.C.; Kuo, Y.C.; Chen, Y.H.; Lee, W.C.; Chang, T.S.; Lan, P.C.; Kuo, H.C.; Lee, K.L.; et al. TGF-βI regulates cell migration through pluripotent transcription factor OCT4 in endometriosis. PLoS ONE 2015, 10, e0145256. [Google Scholar] [CrossRef]
- Abdanipour, A.; Noori-Zadeh, A.; Mesbah-Namin, S.A.; Bakhtiyari, S.; Nejatbakhsh, R.; Anarkooli, I.J. Di-(2-ethylhexyl) phthalate-induced hippocampus-derived neural stem cells proliferation. Cell J. 2017, 19, 166–172. [Google Scholar] [CrossRef]
- Chen, H.P.; Lee, Y.K.; Huang, S.Y.; Shi, P.C.; Hsu, P.C.; Chang, C.F. Phthalate exposure promotes chemotherapeutic drug resistance in colon cancer cells. Oncotarget 2017, 9, 13167–13180. [Google Scholar] [CrossRef]
- Fang, H.; Fang, W.; Cao, H.; Luo, S.; Cong, J.; Liu, S.; Pan, F.; Jia, X. Di-(2-ethylhexyl)-phthalate induces apoptosis via the PPARγ/PTEN/AKT pathway in differentiated human embryonic stem cells. Food Chem. Toxicol. 2019, 131, 110552. [Google Scholar] [CrossRef]
- Gutierrez-Garcia, A.K.; Flores-Kelly, J.M.; Ortiz-Rodriguez, T.; Kalixto-Sanchez, M.A.; De Leon-Rodríguez, A. Phthalates affect the in vitro expansion of human hematopoietic stem cell. Cytotechnology 2019, 71, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.; Teng, Y. Is it time to start transitioning from 2D to 3D cell culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Ikram, M.; Subhan, F.; Kang, H.Y.; Lim, Y.; Lee, R.; Jin, S.; Jeong, Y.H.; Kwak, J.Y.; Na, Y.J.; et al. Alginate–marine collagen–agarose composite hydrogels as matrices for biomimetic 3D cell spheroid formation. RSC Adv. 2016, 6, 46952–46965. [Google Scholar] [CrossRef]
- Moon, S.; Ok, Y.; Hwang, S.; Lim, Y.S.; Kim, H.Y.; Na, Y.J.; Yoon, S. A marine collagen-based biomimetic hydrogel recapitulates cancer stem cell niche and enhances progression and chemoresistance in human ovarian cancer. Mar. Drugs 2020, 18, 498. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5ʹ-3ʹ) | Reverse (5ʹ-3ʹ) |
|---|---|---|
| CD44 | CTGCCGCTTTGCAGGTGTA | CATTGTGGGCAAGGTGCTATT |
| COX-2 | GCCTGAATGTGCCATAAGACTGAC | AAACCCACAGTGCTTGACACAGA |
| E-cadherin | ATTTTTCCCTCGACACCCGAT | TCCCAGGCGTAGACCAAGA |
| ICAM-1 | CCGGAAGGTGTATGAACTGA | GGCAGCGTAGGGTAAGGTT |
| IFN-γ | TGGCTTTTCAGCTCTGCATC | CCGCTACATCTGAATGACCTG |
| IL-1β | CCTGTCCTGCGTGTTGAAAGA | GGGAACTGGGCAGACTCAAA |
| IL-6 | GGTACATCCTCGACGGCATCT | GTGCCTCTTTGCTGCTTTCAC |
| IL-8 | GCATAAAGACATACTCCAAACC | ACTTCTCCACAACCCTCTG |
| Klf4 | CAGCTTCACCTATCCGATCCG | GACTCCCTGCCATAGAGGAGG |
| MCP-1 | CCGAGAGGCTGAGACTAACC | CTTTCATGCTGGAGGCGAGA |
| MMP-2 | TGACGGTAAGGACGGACTC | ATACTTCACACGGACCACTTG |
| MMP-9 | TTGACAGCGACAAGAAGTGG | GCCATTCACGTCGTCCTTAT |
| NANOG | CAAAGCAGGAGTCCACTGAG | TAAGGGCATCCACTTCACAG |
| N-cadherin | AGCCAACCTTAACTGAGGAGT | GGCAAGTTGATTGGAGGGATG |
| OCT4 | CTTGAATCCCGAATGGAAAGGG | GTGTATATCCCAGGGTGATCCTC |
| RANTES | TCTGCGCTCCTGCATCTG | AGTGGGCGGGCAATGTAG |
| SNAIL | ACTGCAACAAGGAATACCTCAG | GCACTGGTACTTCTTGACATCTG |
| SLUG | TGTGACAAGGAATATGTGAGCC | TGTGACAAGGAATATGTGAGCC |
| SOX2 | TACAGCATGTCCTACTCGCAG | GAGGAAGAGGTAACCACAGGG |
| TNF-α | CCCAGGGACCTCTCTCTAATC | ATGGGCTACAGGCTTGTCACT |
| Vimentin | CAAAGCAGGAGTCCACTGAG | TAAGGGCATCCACTTCACAG |
| VCAM-1 | ACACCTCCCCCAAGAATACAG | GCTCATCCTCAACACCCACAG |
| ZEB1 | TTACACCTTTGCATACAGAACCC | TTTACGATTACACCCAGACTGC |
| ZEB2 | GCGATGGTCATGCAGTCAG | CAGGTGGCAGGTCATTTTCTT |
| GAPDH | GGAGAAGGCTGGGGCTCAT | TGATGGCATGGACTGTGGTC |
| Antibody | Source | Antibody Type | Size (kDa) |
|---|---|---|---|
| E-cadherin | Abcam | Mouse monoclonal | 110 |
| KLF4 | Abcam | Rabbit polyoclonal | 55 |
| NANOG | Abcam | Rabbit polyclonal | 35 |
| N-cadherin | Abcam | Mouse monoclonal | 100 |
| OCT4 | Abcam | Rabbit polyclonal | 43 |
| pSMAD-2 | Cell Signaling Technology | Rabbit monoclonal | 60 |
| SLUG | Cell Signaling Technology | Rabbit monoclonal | 30 |
| SNAIL | Cell Signaling Technology | Rabbit monoclonal | 29 |
| SOX2 | Abcam | Rabbit polyclonal | 34 |
| TGF-βR2 | Abcam | Rabbit polyclonal | 65 |
| TWIST | Abcam | Mouse monoclonal | 21 |
| Vimentin | Abcam | Rabbit polyclonal | 54 |
| ZEB1 | Cell Signaling Technology | Rabbit monoclonal | 200 |
| β-actin | Santa Cruz Biotechnology | Mouse monoclonal | 42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.G.; Lim, Y.S.; Hwang, S.; Kim, H.-Y.; Moon, Y.; Song, Y.J.; Na, Y.-J.; Yoon, S. Di-(2-ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial–Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 3938. https://doi.org/10.3390/ijms23073938
Kim HG, Lim YS, Hwang S, Kim H-Y, Moon Y, Song YJ, Na Y-J, Yoon S. Di-(2-ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial–Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(7):3938. https://doi.org/10.3390/ijms23073938
Chicago/Turabian StyleKim, Hwi Gon, Ye Seon Lim, Seonyeong Hwang, Hye-Yoon Kim, Yuseok Moon, Yong Jung Song, Yong-Jin Na, and Sik Yoon. 2022. "Di-(2-ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial–Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway" International Journal of Molecular Sciences 23, no. 7: 3938. https://doi.org/10.3390/ijms23073938
APA StyleKim, H. G., Lim, Y. S., Hwang, S., Kim, H. -Y., Moon, Y., Song, Y. J., Na, Y. -J., & Yoon, S. (2022). Di-(2-ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial–Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway. International Journal of Molecular Sciences, 23(7), 3938. https://doi.org/10.3390/ijms23073938