
Hyperosmotic Stress Induces Phosphorylation of CERT and Enhances Its Tethering throughout the Endoplasmic Reticulum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
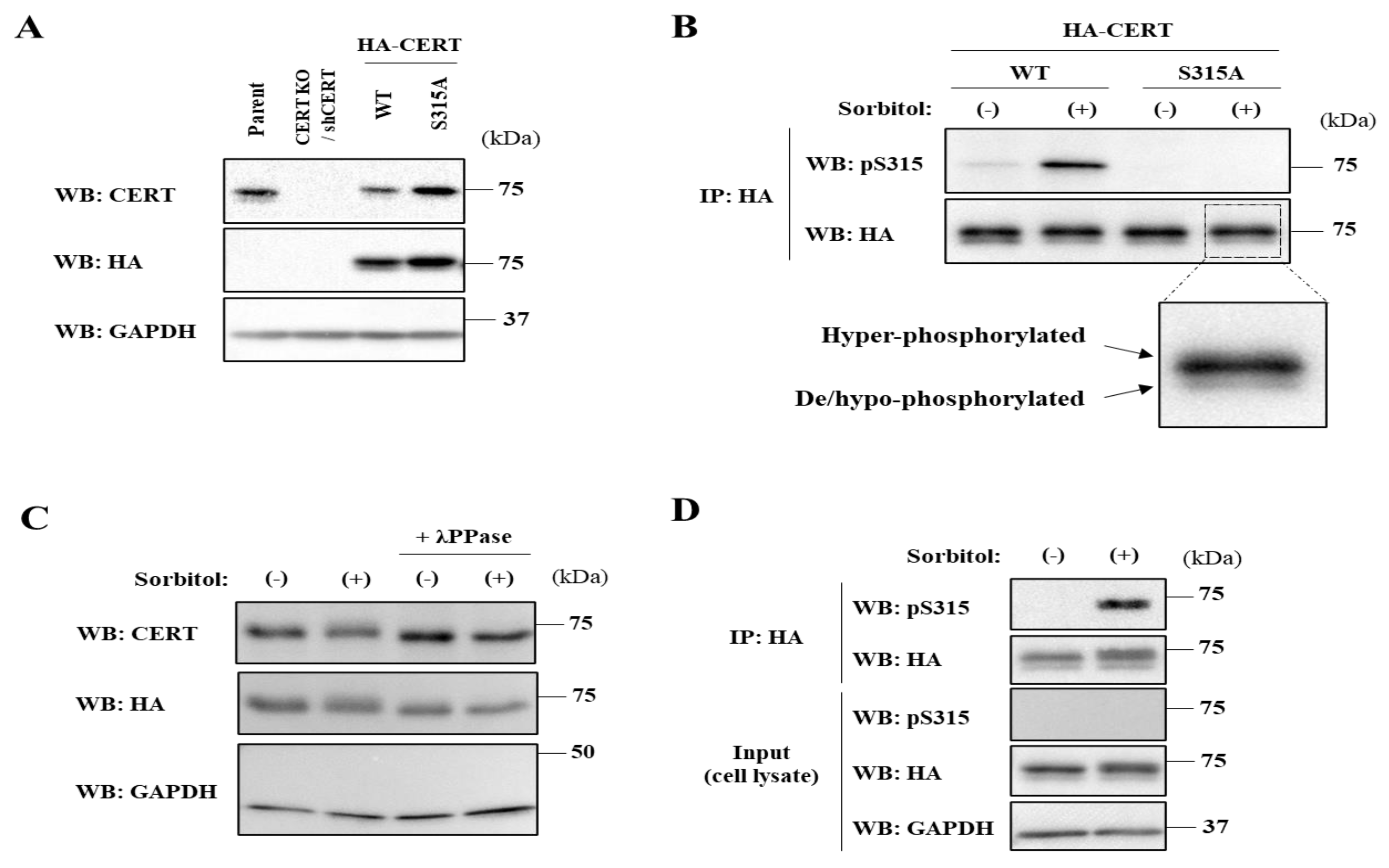
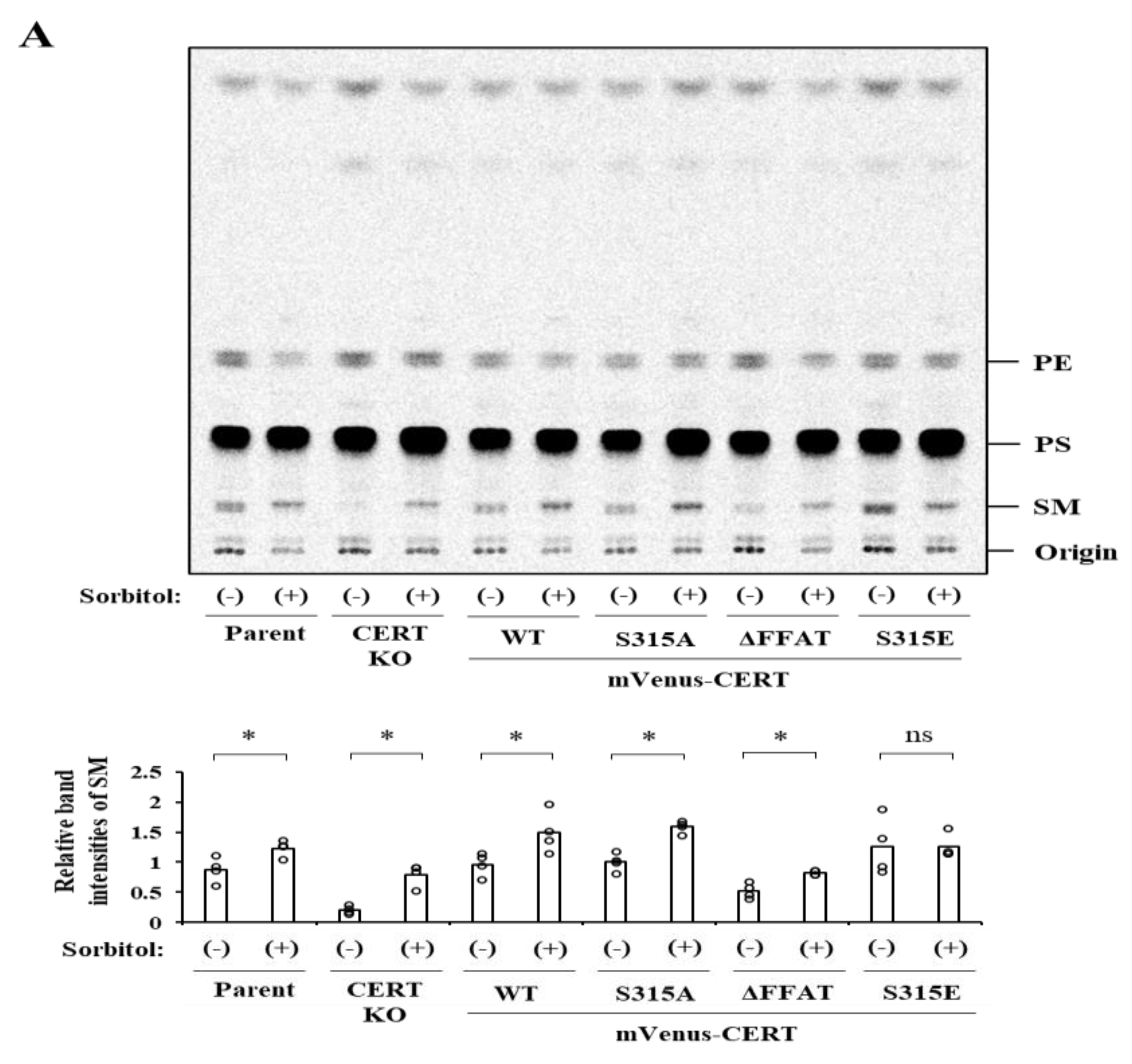
2.1. Phosphorylation State of S315 but Not SRM in CERT Is Affected by Hyperosmotic Stress
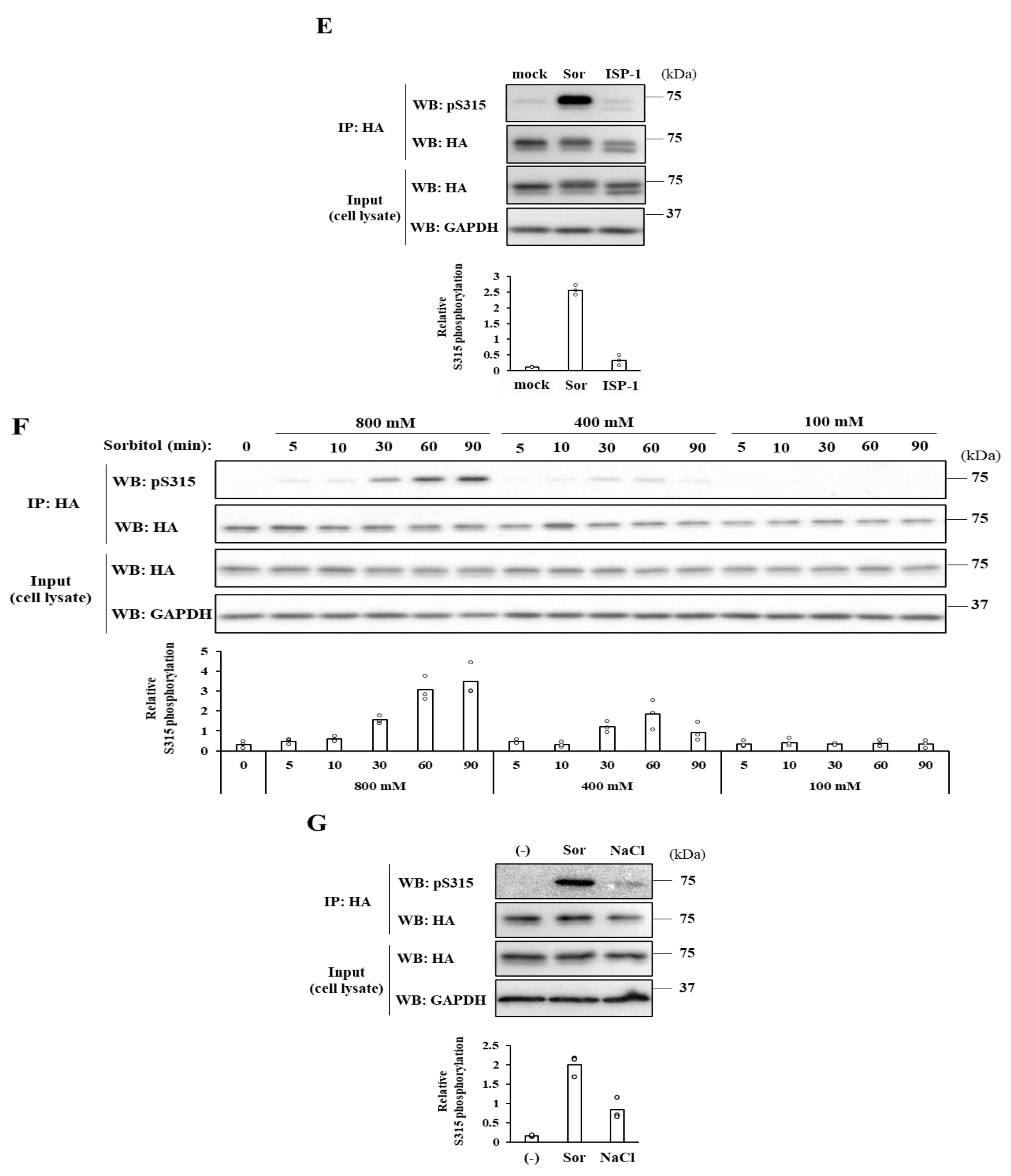
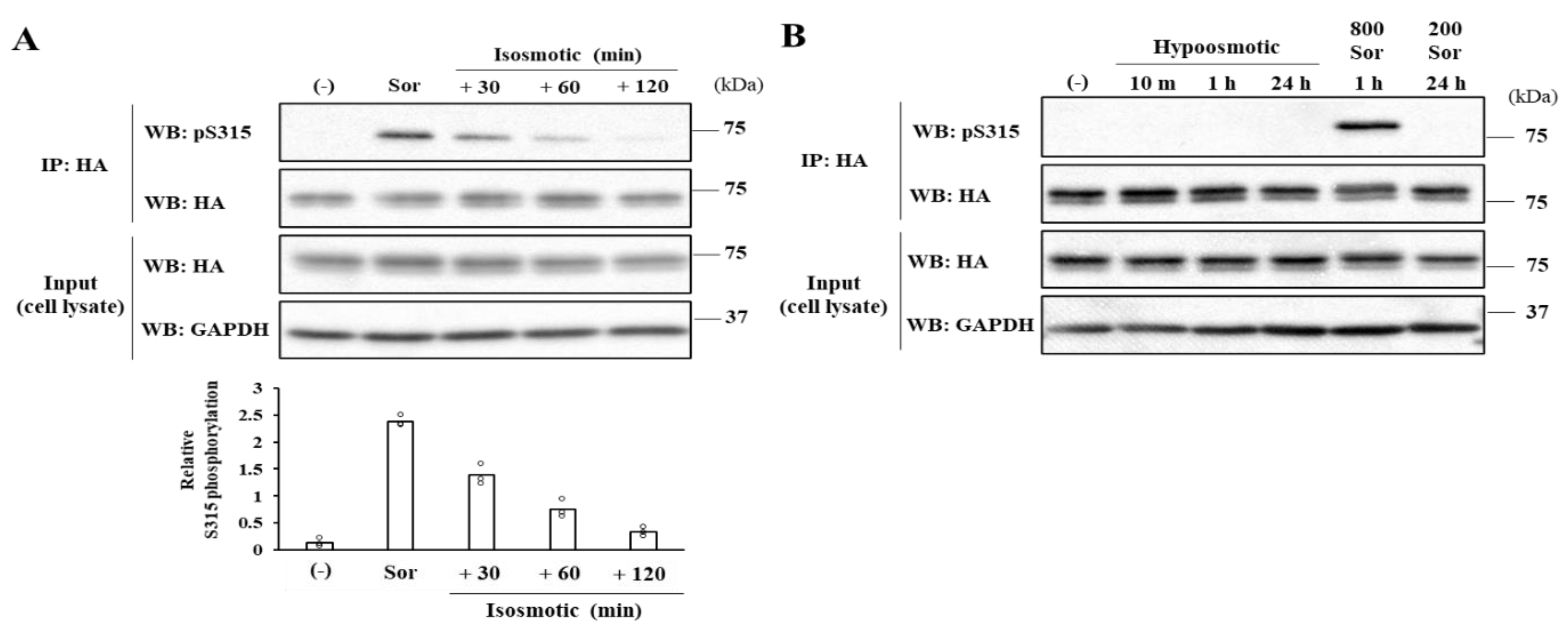
2.2. Severe Hyperosmotic Stress, but Not Mild Hyperosmotic nor Hypoosmotic Stress, Induces Reversible CERT S315 Phosphorylation
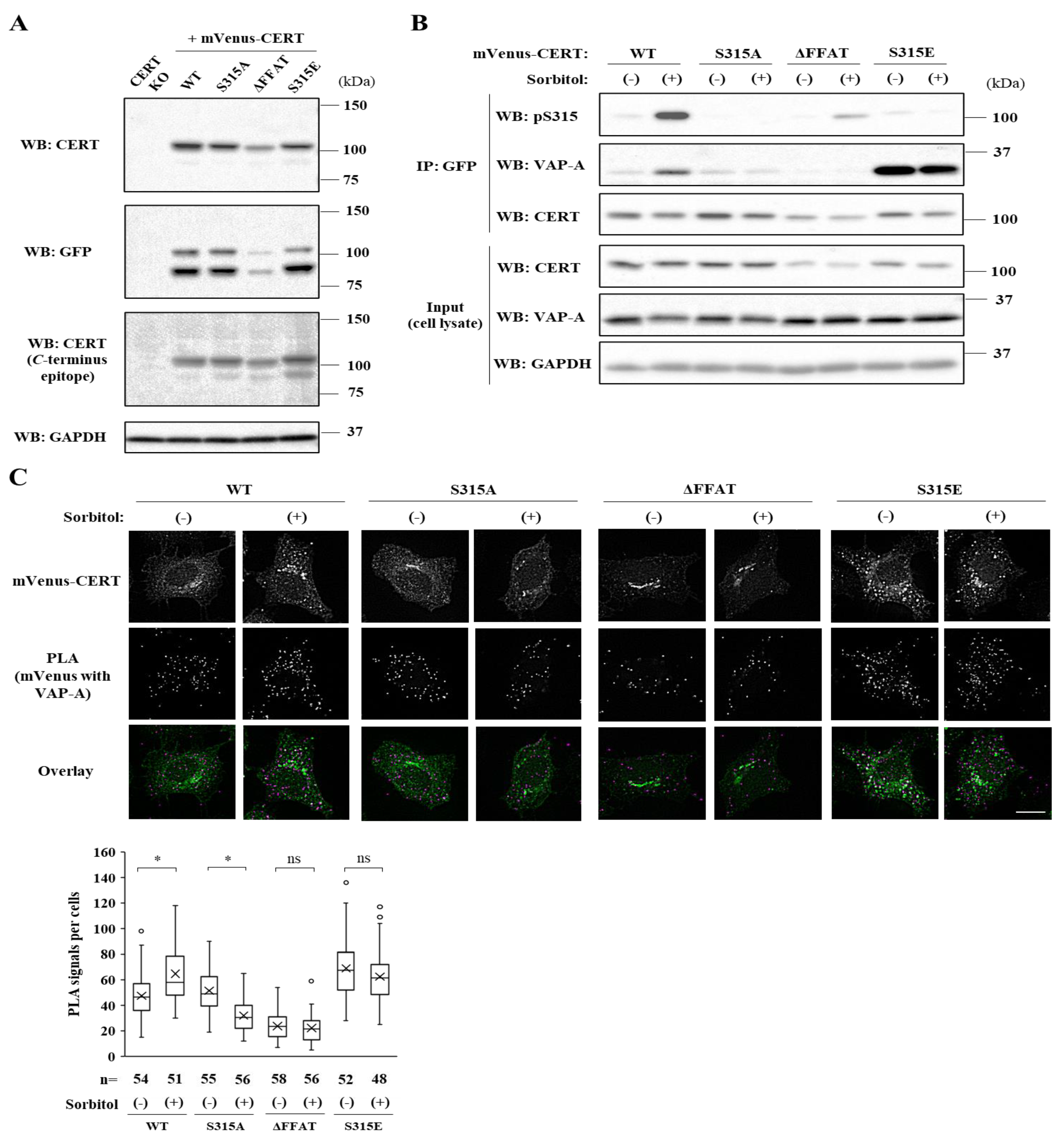
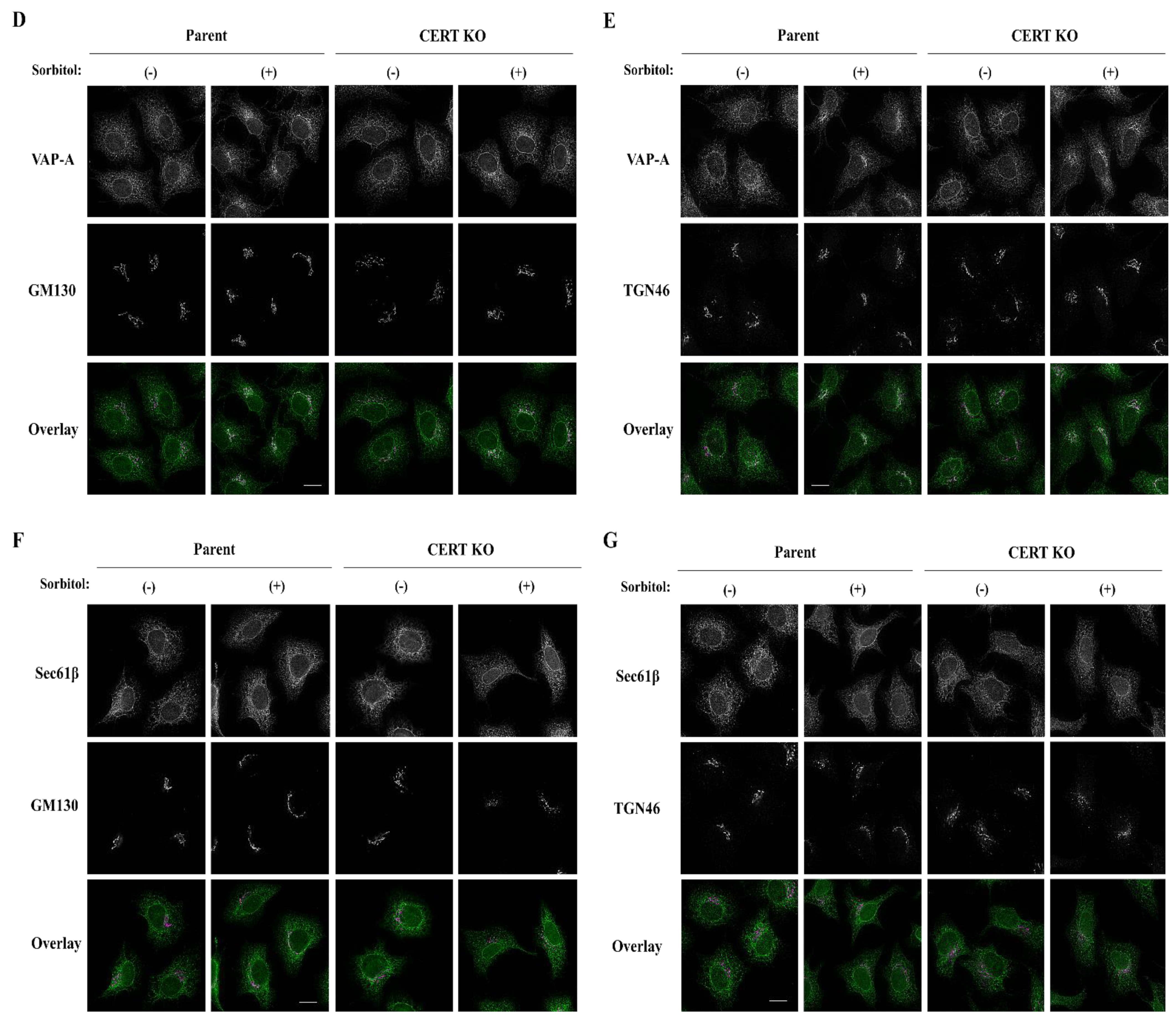
2.3. The FFAT Motif-Dependent Interaction of CERT with VAP-A Is Enhanced under Hyperosmotic Conditions
2.4. Effects of Hyperosmotic Stress on the Synthesis of SM
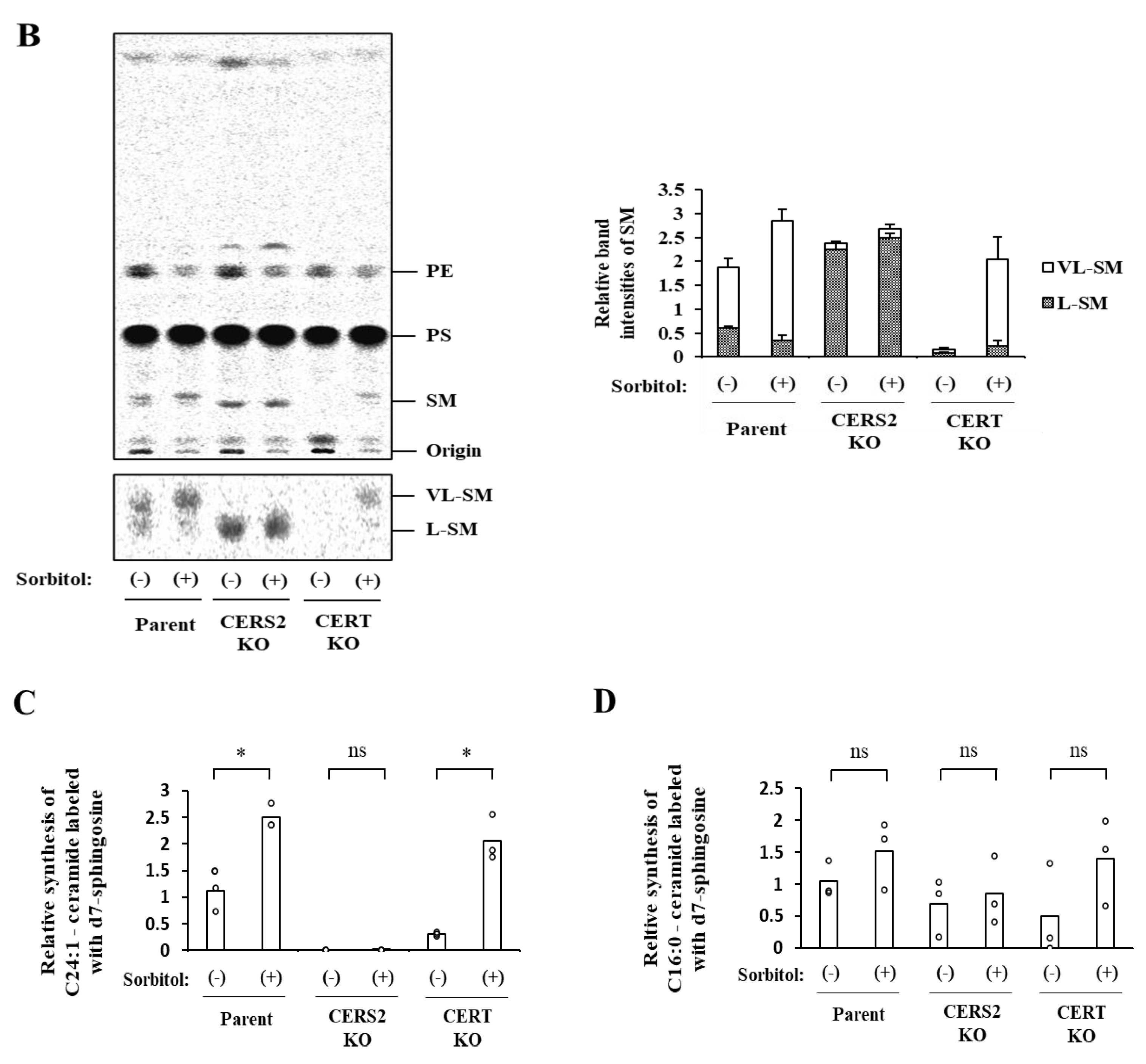
2.5. Hyperosmotic Stress Induces the Preferential Synthesis of the Very-Long-Chain SM via a CERS2-Dependent and CERT-Independent Manner
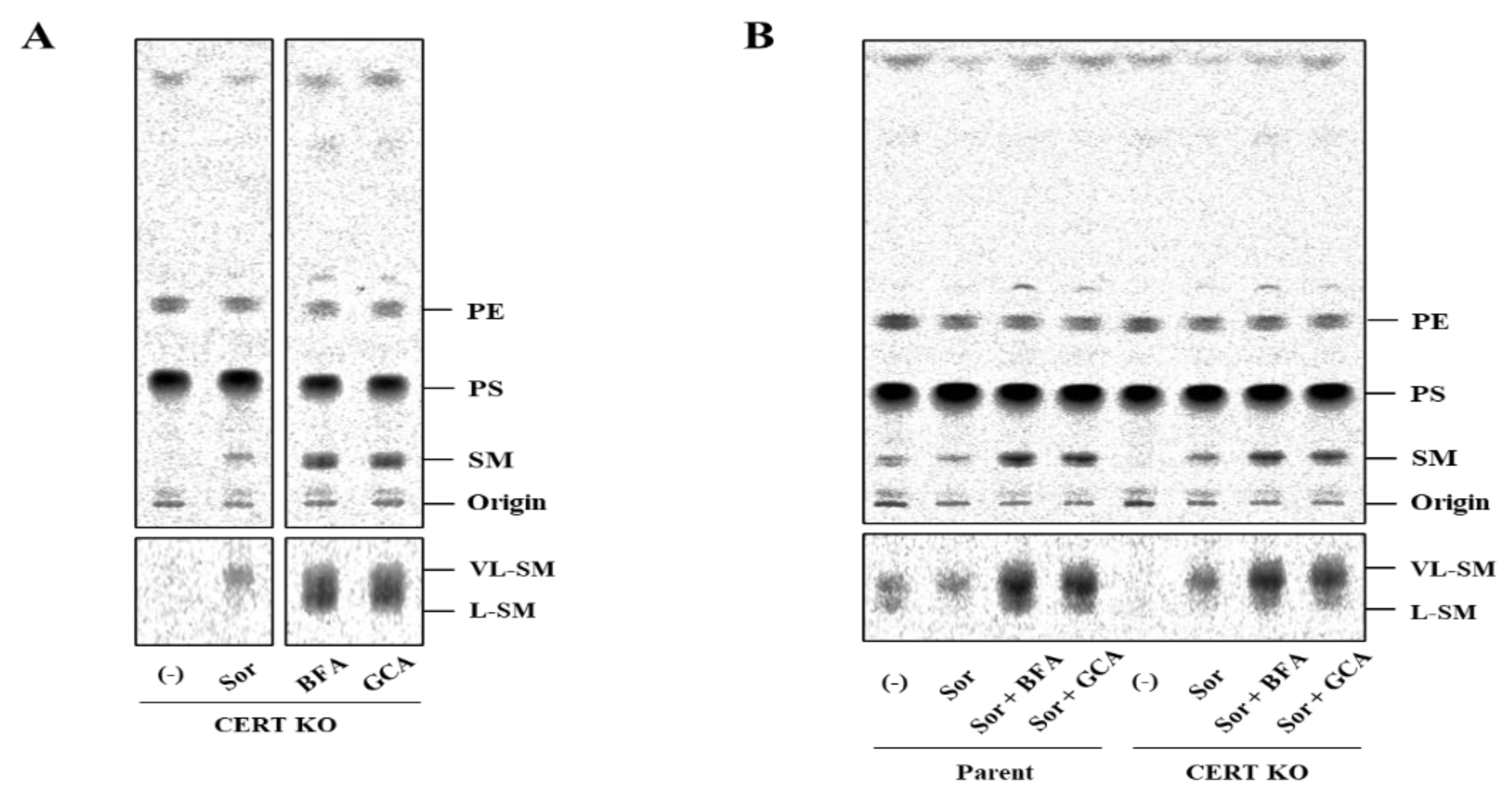
2.6. Pharmacologically Induced Merging of the Golgi Apparatus with the ER Does Not Abrogate the Hyperosmotic Stress-Induced Preferential Synthesis of VL-SM
3. Discussion
4. Materials and Methods
4.1. HeLa Cell Lines and Cell Culture
4.2. Plasmids
4.3. Retroviral Transfection and Establishment of Stable Cell Lines
4.4. Western Blotting
4.5. Protein Phosphatase Treatment
4.6. Immunoprecipitation
4.7. Immunocytochemistry
4.8. In situ Proximity Ligation Assay (PLA)
4.9. Metabolic Labeling of Lipids with Radioactive Serine
4.10. In Vitro Ceramide Synthesis Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanada, K. Lipid Transfer Proteins Rectify Inter-Organelle Flux and Accurately Deliver Lipids at Membrane Contact Sites. J. Lipid Res. 2018, 59, 1341–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, L.H.; Gatta, A.T.; Levine, T.P. Lipid Transfer Proteins: The Lipid Commute via Shuttles, Bridges and Tubes. Nat. Rev. Mol. Cell Biol. 2019, 20, 85–101. [Google Scholar] [CrossRef]
- Hanada, K.; Kumagai, K.; Yasuda, S.; Miura, Y.; Kawano, M.; Fukasawa, M.; Nishijima, M. Molecular Machinery for Non-Vesicular Trafficking of Ceramide. Nature 2003, 426, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Kawano, M.; Kumagai, K.; Nishijima, M.; Hanada, K. Efficient Trafficking of Ceramide from the Endoplasmic Reticulum to the Golgi Apparatus Requires a VAMP-Associated Protein-Interacting FFAT Motif of CERT. J. Biol. Chem. 2006, 281, 30279–30288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, T.; Hanada, K. Establishment of HeLa Cell Mutants Deficient in Sphingolipid-Related Genes Using TALENs. PLoS ONE 2014, 9, e88124. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, T.; Horie, A.; Tachida, Y.; Sakuma, C.; Suzuki, Y.; Kushi, Y.; Hanada, K. Role of Intracellular Lipid Logistics in the Preferential Usage of Very Long Chain-Ceramides in Glucosylceramide. Int. J. Mol. Sci. 2016, 17, 1761. [Google Scholar] [CrossRef] [Green Version]
- Fugmann, T.; Hausser, A.; Schöffler, P.; Schmid, S.; Pfizenmaier, K.; Olayioye, M.A. Regulation of Secretory Transport by Protein Kinase D-Mediated Phosphorylation of the Ceramide Transfer Protein. J. Cell Biol. 2007, 178, 15–22. [Google Scholar] [CrossRef]
- Tomishige, N.; Kumagai, K.; Kusuda, J.; Nishijima, M.; Hanada, K. Casein Kinase Iγ2 Down-Regulates Trafficking of Ceramide in the Synthesis of Sphingomyelin. Mol. Biol. Cell 2009, 20, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, K.; Kawano, M.; Shinkai-Ouchi, F.; Nishijima, M.; Hanada, K. Interorganelle Trafficking of Ceramide Is Regulated by Phosphorylation–Dependent Cooperativity between the PH and START Domains of CERT. J. Biol. Chem. 2007, 282, 17758–17766. [Google Scholar] [CrossRef] [Green Version]
- Kumagai, K.; Kawano-Kawada, M.; Hanada, K. Phosphoregulation of the Ceramide Transport Protein CERT at Serine 315 in the Interaction with VAMP-Associated Protein (VAP) for Inter-Organelle Trafficking of Ceramide in Mammalian Cells. J. Biol. Chem. 2014, 289, 10748–10760. [Google Scholar] [CrossRef] [Green Version]
- Favale, N.O.; Santacreu, B.J.; Pescio, L.G.; Marquez, M.G.; Sterin-Speziale, N.B. Sphingomyelin Metabolism Is Involved in the Differentiation of MDCK Cells Induced by Environmental Hypertonicity. J. Lipid Res. 2015, 56, 786–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fredriksson, S.; Gullberg, M.; Jarvius, J.; Olsson, C.; Pietras, K.; Gústafsdóttir, S.M.; Östman, A.; Landegren, U. Protein Detection Using Proximity-Dependent DNA Ligation Assays. Nat. Biotechnol. 2002, 20, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Söderberg, O.; Gullberg, M.; Jarvius, M.; Ridderstråle, K.; Leuchowius, K.-J.; Jonas, J.; Wester, K.; Hydbring, P.; Bahram, F.; Larsson, L.-G.; et al. Direct Observation of Individual Endogenous Protein Complexes in Situ by Proximity Ligation. Nat. Methods 2006, 12, 995–1000. [Google Scholar] [CrossRef]
- Krokowski, D.; Guan, B.J.; Wu, J.; Zheng, Y.; Pattabiraman, P.P.; Jobava, R.; Gao, X.H.; Di, X.J.; Snider, M.D.; Mu, T.W.; et al. GADD34 Function in Protein Trafficking Promotes Adaptation to Hyperosmotic Stress in Human Corneal Cells. Cell Rep. 2017, 21, 2895–2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizutani, Y.; Kihara, A.; Chiba, H.; Tojo, H.; Igarashi, Y. 2-Hydroxy-Ceramide Synthesis by Ceramide Synthase Family: Enzymatic Basis for the Preference of FA Chain Length. J. Lipid Res. 2008, 49, 2356–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, Y.; Kohyama-Koganeya, A.; Hirabayashi, Y. New Insights on Glucosylated Lipids: Metabolism and Functions. Biochim. Biophys. Acta–Mol. Cell Biol. Lipids 2013, 1831, 1475–1485. [Google Scholar] [CrossRef] [Green Version]
- Donaldson, J.G.; Finazzi, D.; Klausner, R.D. Brefeldin a Inhibits Golgi Membrane-Catalysed Exchange of Guanine Nucleotide onto ARF Protein. Nature 1992, 360, 350–352. [Google Scholar] [CrossRef] [PubMed]
- Sciaky, N.; Presley, J.; Smith, C.; Zaal, K.J.M.; Cole, N.; Moreira, J.E.; Terasaki, M.; Siggia, E.; Lippincott-Schwartz, J. Golgi Tubule Traffic and the Effects of Brefeldin A Visualized in Living Cells. J. Cell Biol. 1997, 139, 1137–1155. [Google Scholar] [CrossRef] [PubMed]
- Saenz, J.B.; Sun, W.J.; Chang, J.W.; Li, J.; Bursulaya, B.; Gray, N.S.; Haslam, D.B. Golgicide A Reveals Essential Roles for GBF1 in Golgi Assembly and Function. Nat. Chem. Biol. 2009, 5, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Mogelsvang, S.; Marsh, B.J.; Ladinsky, M.S.; Howell, K.E. Predicting Function from Structure: 3D Structure Studies of the Mammalian Golgi Complex. Traffic 2004, 5, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Tafesse, F.G.; Vacaru, A.M.; Bosma, E.F.; Hermansson, M.; Jain, A.; Hilderink, A.; Somerharju, P.; Holthuis, J.C.M. Sphingomyelin Synthase-Related Protein SMSr Is a Suppressor of Ceramide-Induced Mitochondrial Apoptosis. J. Cell Sci. 2014, 127, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, A.; Beutel, O.; Ebell, K.; Korneev, S.; Holthuis, J.C.M. Diverting CERT-Mediated Ceramide Transport to Mitochondria Triggers Bax-Dependent Apoptosis. J. Cell Sci. 2017, 130, 360–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.H.; Linstedt, A.D. Osmotically Induced Cell Volume Changes Alter Anterograde and Retrograde Transport, Golgi Structure, and COPI Dissociation. Mol. Biol. Cell 1999, 10, 1445–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, P.; Roth, I.; Meda, P.; Féraille, E.; Brown, D.; Hasler, U. Ionic Imbalance, in Addition to Molecular Crowding, Abates Cytoskeletal Dynamics and Vesicle Motility during Hypertonic Stress. Proc. Natl. Acad. Sci. USA 2015, 112, E3104–E3113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knepper, M.A. Measurement of Osmolality in Kidney Slices Using Vapor Pressure Osmometry. Kidney Int. 1982, 21, 653–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.; Wakabayashi, M.; Shimizu, Y.; Yoshioka, T.; Higashino, K.; Numata, Y.; Okuda, T.; Zhao, S.; Sakai, S.; Igarashi, Y.; et al. Imaging Mass Spectrometry Reveals Acyl-Chain- and Region-Specific Sphingolipid Metabolism in the Kidneys of Sphingomyelin Synthase 2-Deficient Mice. PLoS ONE 2016, 11, e0152191. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Yin, N.; Luo, Q.; Liu, J.; Chen, X.; Liu, L.; Wu, J. Enhancement of Sphingolipid Synthesis Improves Osmotic Tolerance of Saccharomyces Cerevisiae. Appl. Environ. Microbiol. 2020, 86, e02911-19. [Google Scholar] [CrossRef] [Green Version]
- Stith, J.L.; Velazquez, F.N.; Obeid, L.M. Advances in Determining Signaling Mechanisms of Ceramide and Role in Disease. J. Lipid Res. 2019, 60, 913–918. [Google Scholar] [CrossRef] [Green Version]
- Fekry, B.; Jeffries, K.A.; Esmaeilniakooshkghazi, A.; Szulc, Z.M.; Knagge, K.J.; Kirchner, D.R.; Horita, D.A.; Krupenko, S.A.; Krupenko, N.I. C16-Ceramide Is a Natural Regulatory Ligand of P53 in Cellular Stress Response. Nat. Commun. 2018, 9, 4149. [Google Scholar] [CrossRef]
- Thiemicke, A.; Neuert, G. Kinetics of Osmotic Stress Regulate a Cell Fate Switch of Cell Survival. Sci. Adv. 2021, 7, eabe1122. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Nakao, N.; Ueno, M.; Sakai, S.; Egawa, D.; Hanzawa, H.; Kawasaki, S.; Kumagai, K.; Suzuki, M.; Kobayashi, S.; Hanada, K. Natural Ligand-Nonmimetic Inhibitors of the Lipid-Transfer Protein CERT. Commun. Chem. 2019, 2, 1–11. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimasaki, K.; Kumagai, K.; Sakai, S.; Yamaji, T.; Hanada, K. Hyperosmotic Stress Induces Phosphorylation of CERT and Enhances Its Tethering throughout the Endoplasmic Reticulum. Int. J. Mol. Sci. 2022, 23, 4025. https://doi.org/10.3390/ijms23074025
Shimasaki K, Kumagai K, Sakai S, Yamaji T, Hanada K. Hyperosmotic Stress Induces Phosphorylation of CERT and Enhances Its Tethering throughout the Endoplasmic Reticulum. International Journal of Molecular Sciences. 2022; 23(7):4025. https://doi.org/10.3390/ijms23074025
Chicago/Turabian StyleShimasaki, Kentaro, Keigo Kumagai, Shota Sakai, Toshiyuki Yamaji, and Kentaro Hanada. 2022. "Hyperosmotic Stress Induces Phosphorylation of CERT and Enhances Its Tethering throughout the Endoplasmic Reticulum" International Journal of Molecular Sciences 23, no. 7: 4025. https://doi.org/10.3390/ijms23074025
APA StyleShimasaki, K., Kumagai, K., Sakai, S., Yamaji, T., & Hanada, K. (2022). Hyperosmotic Stress Induces Phosphorylation of CERT and Enhances Its Tethering throughout the Endoplasmic Reticulum. International Journal of Molecular Sciences, 23(7), 4025. https://doi.org/10.3390/ijms23074025