
In Vitro Reactivity of the Glucose Degradation Product 3,4-Dideoxyglucosone-3-ene (3,4-DGE) towards Abundant Components of the Human Blood Circulatory System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
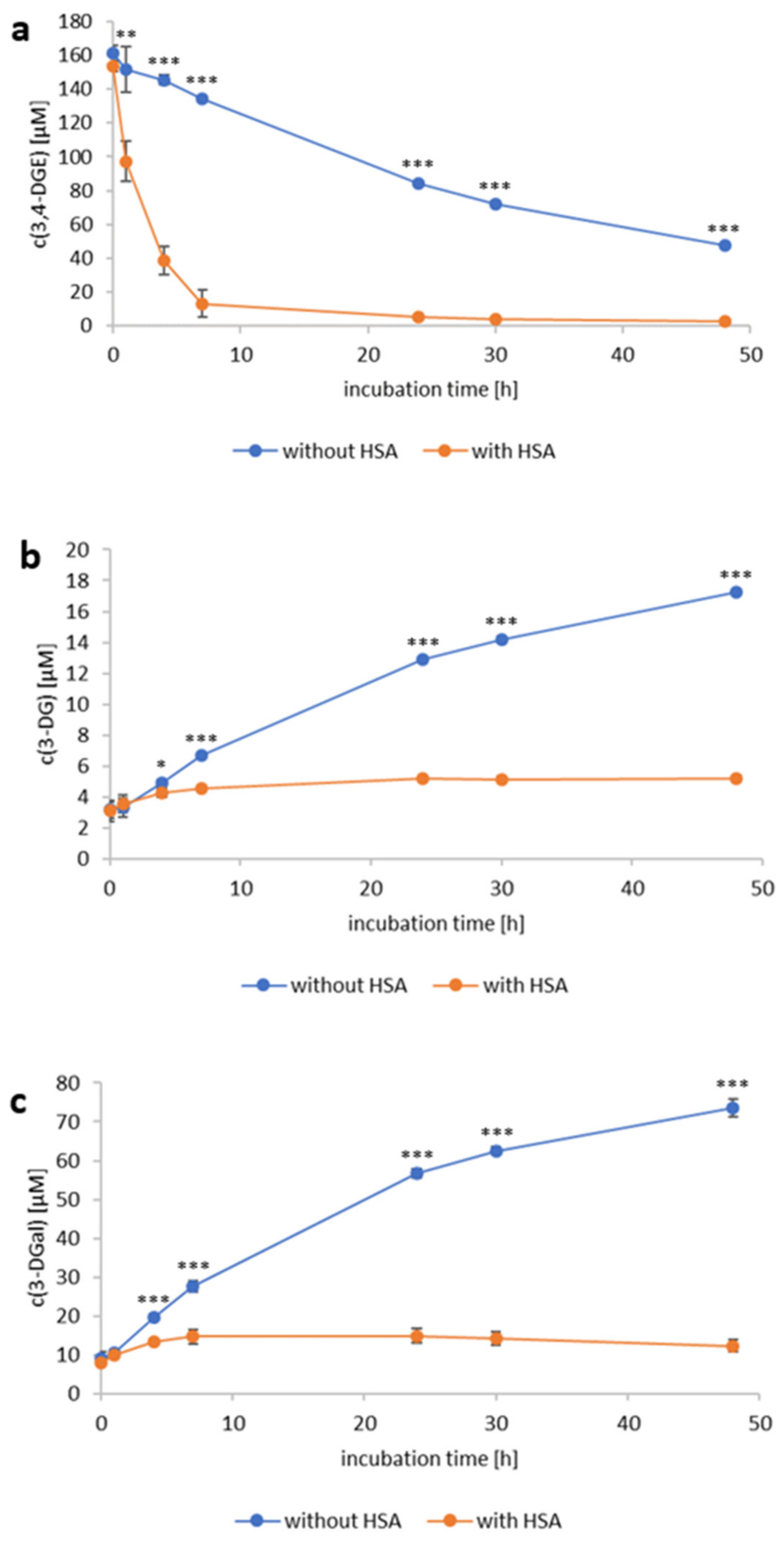
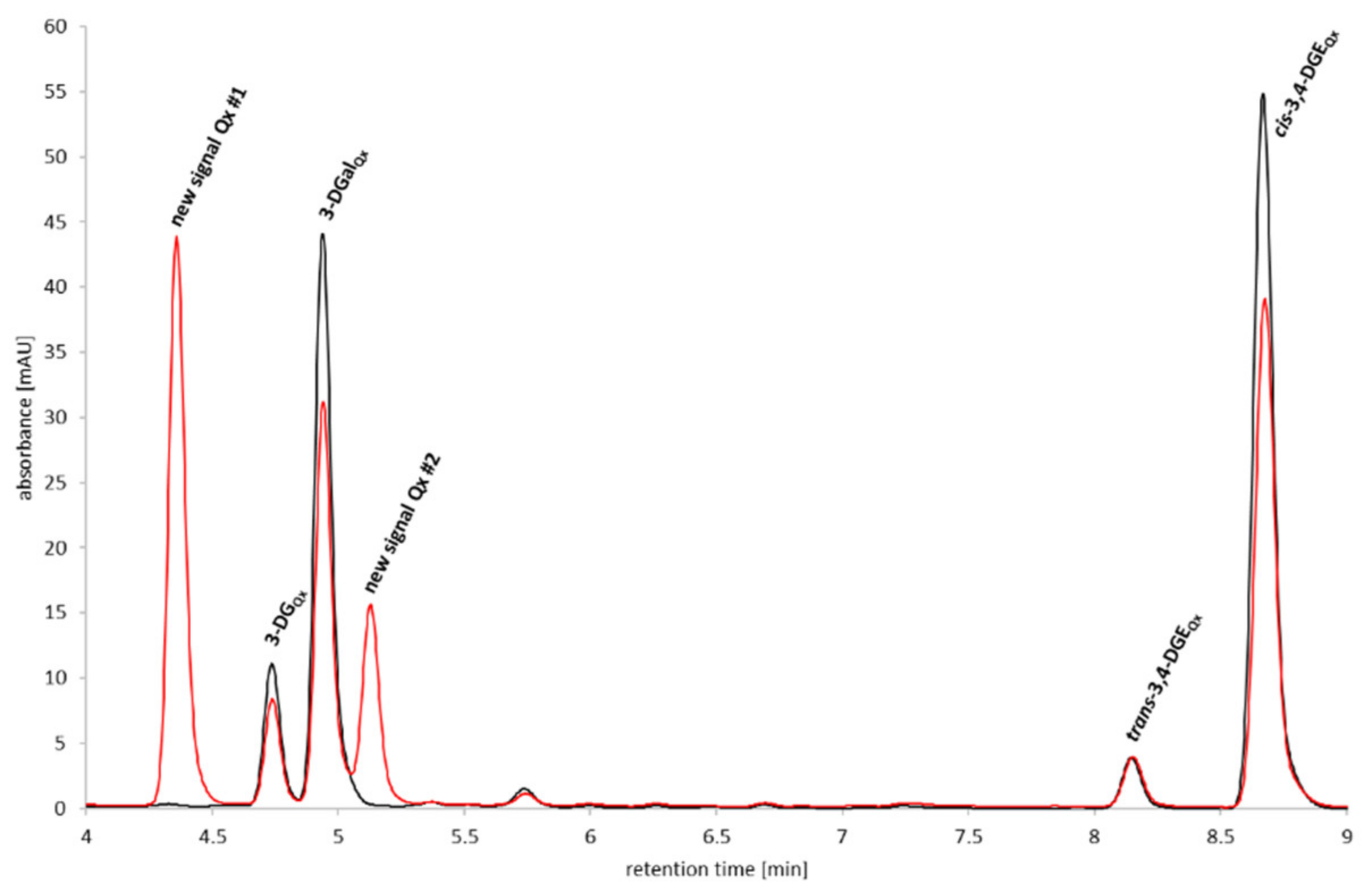
2.1. Reactivity of 3,4-Dideoxyglucosone-3-ene towards Human Serum Albumin
2.2. Effects of Glycation with 3,4-Dideoxyglucosone-3-ene on the Thiol Content, Ketoprofen-Binding, and Esterase-like Activity of Human Serum Albumin
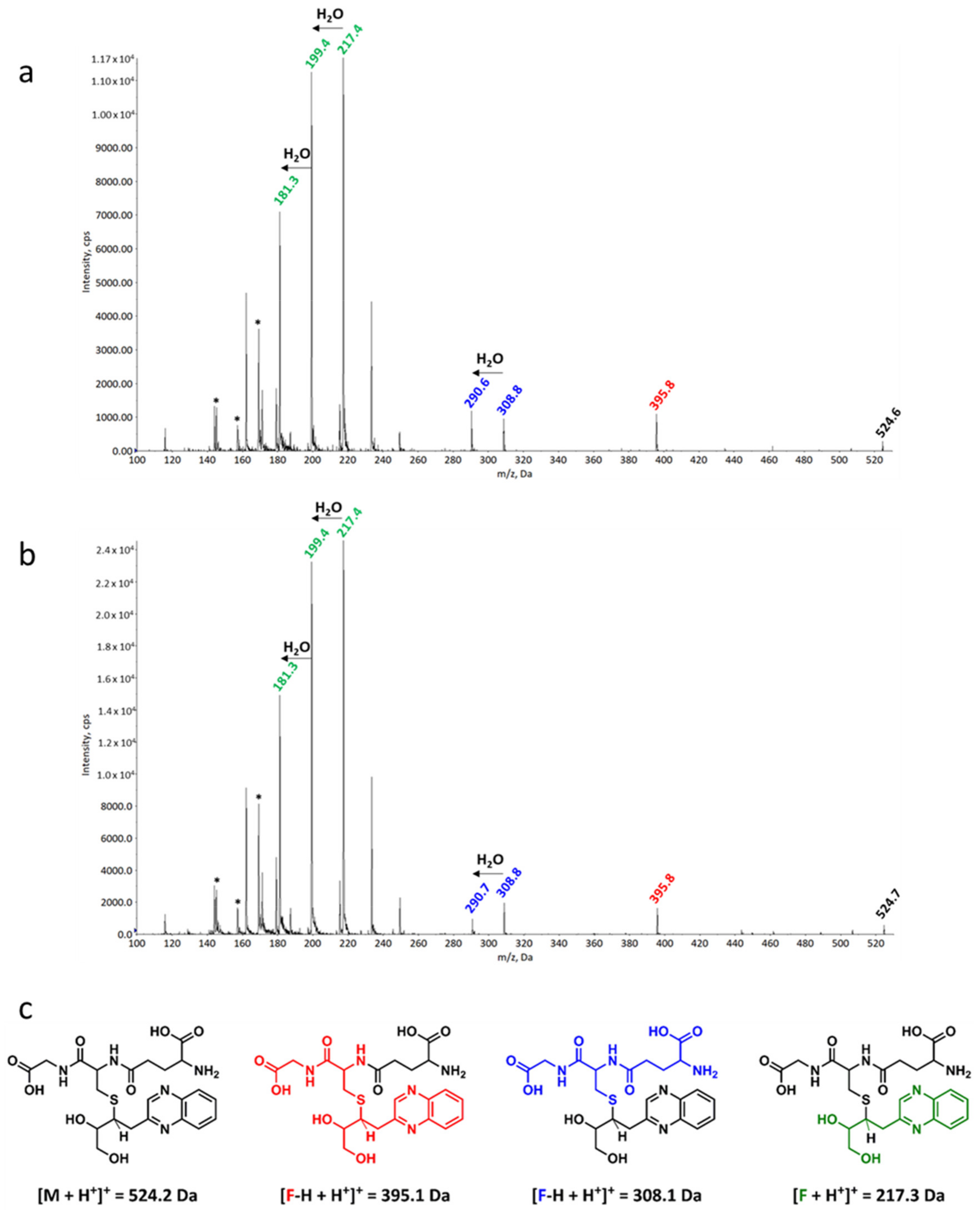
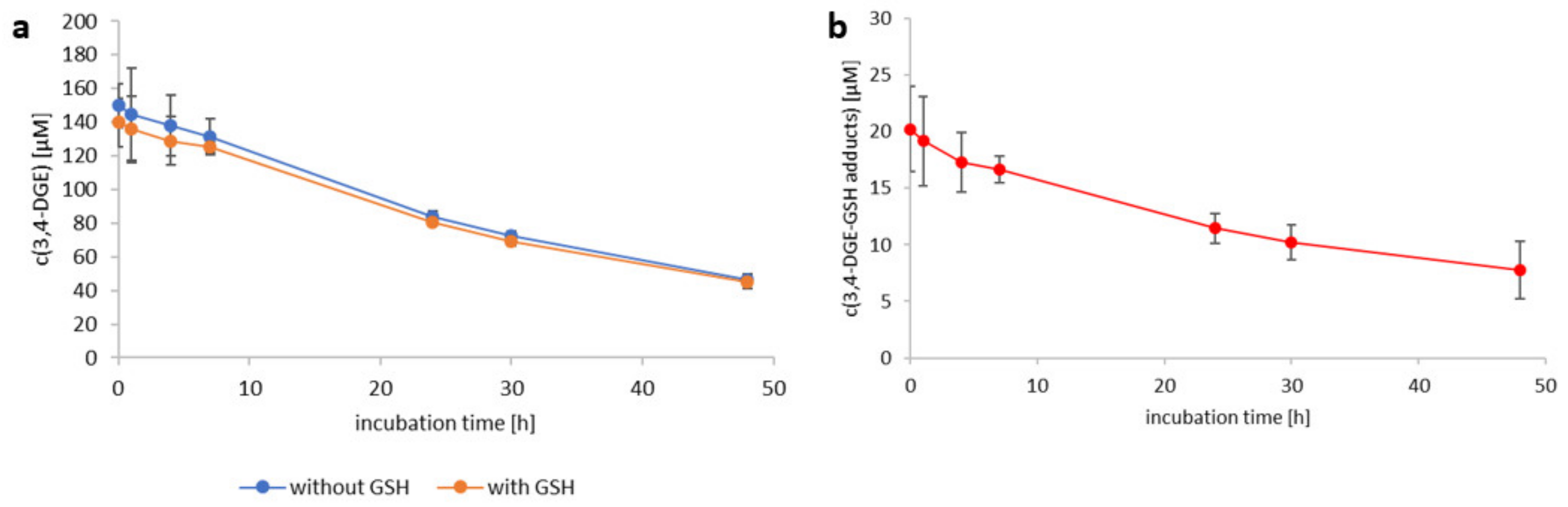
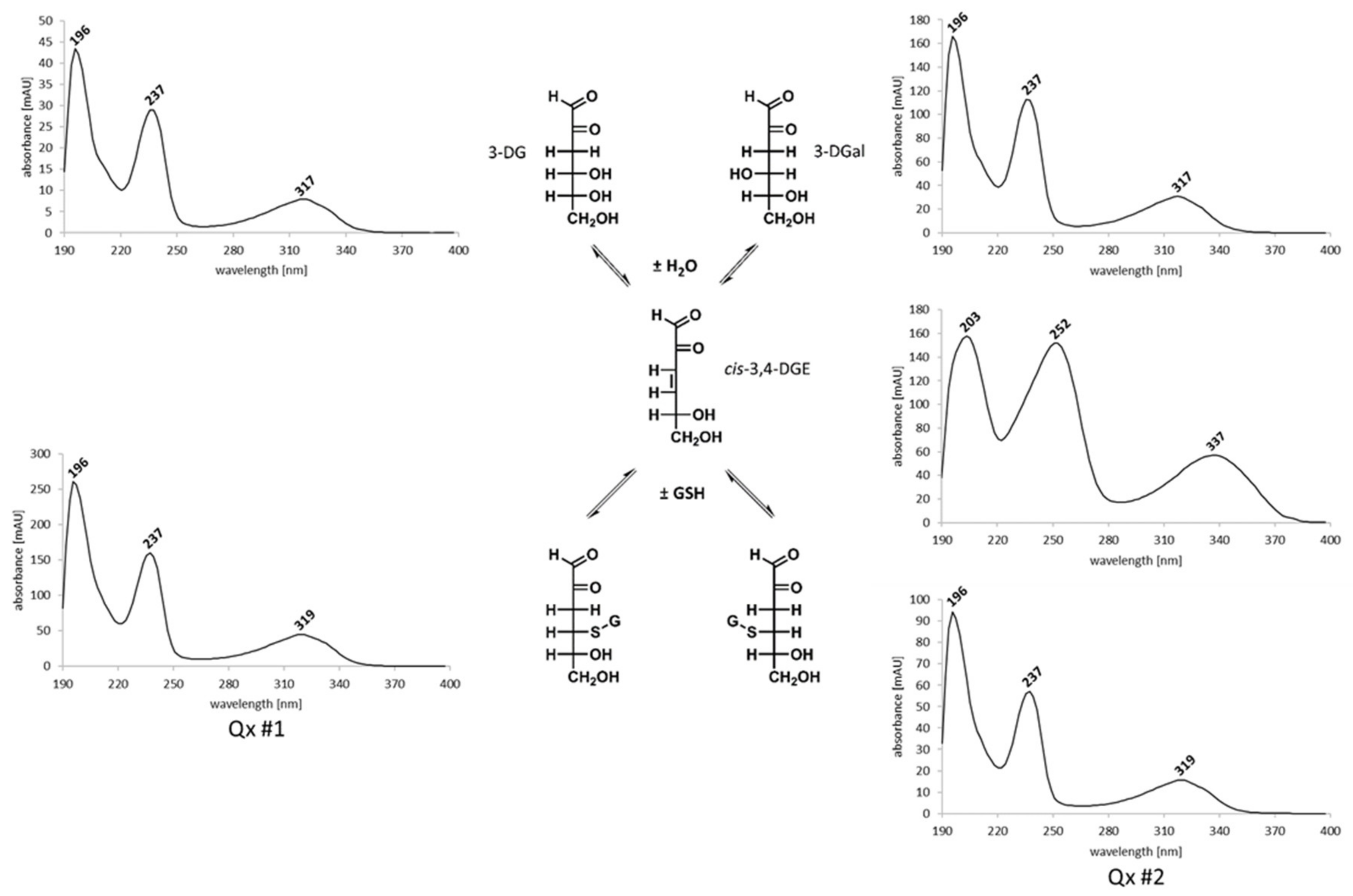
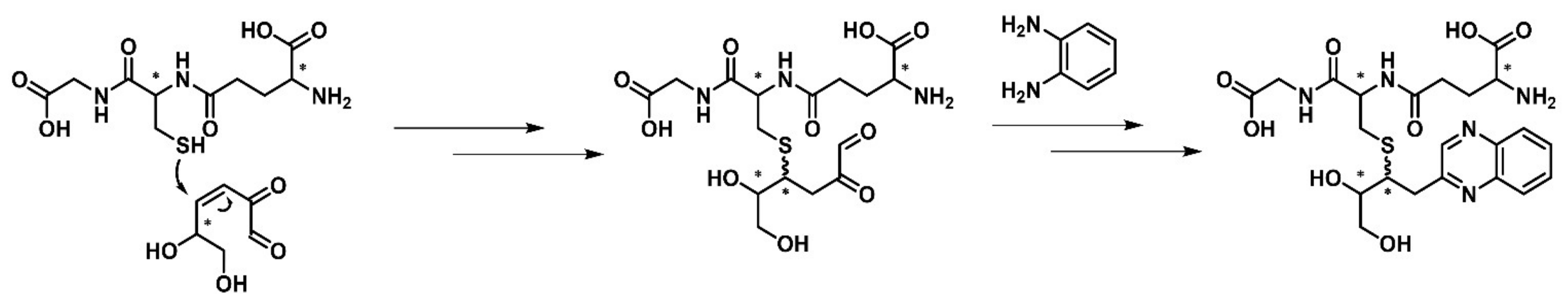
2.3. Reactivity of 3,4-Dideoxyglucosone-3-ene towards l-Glutathione
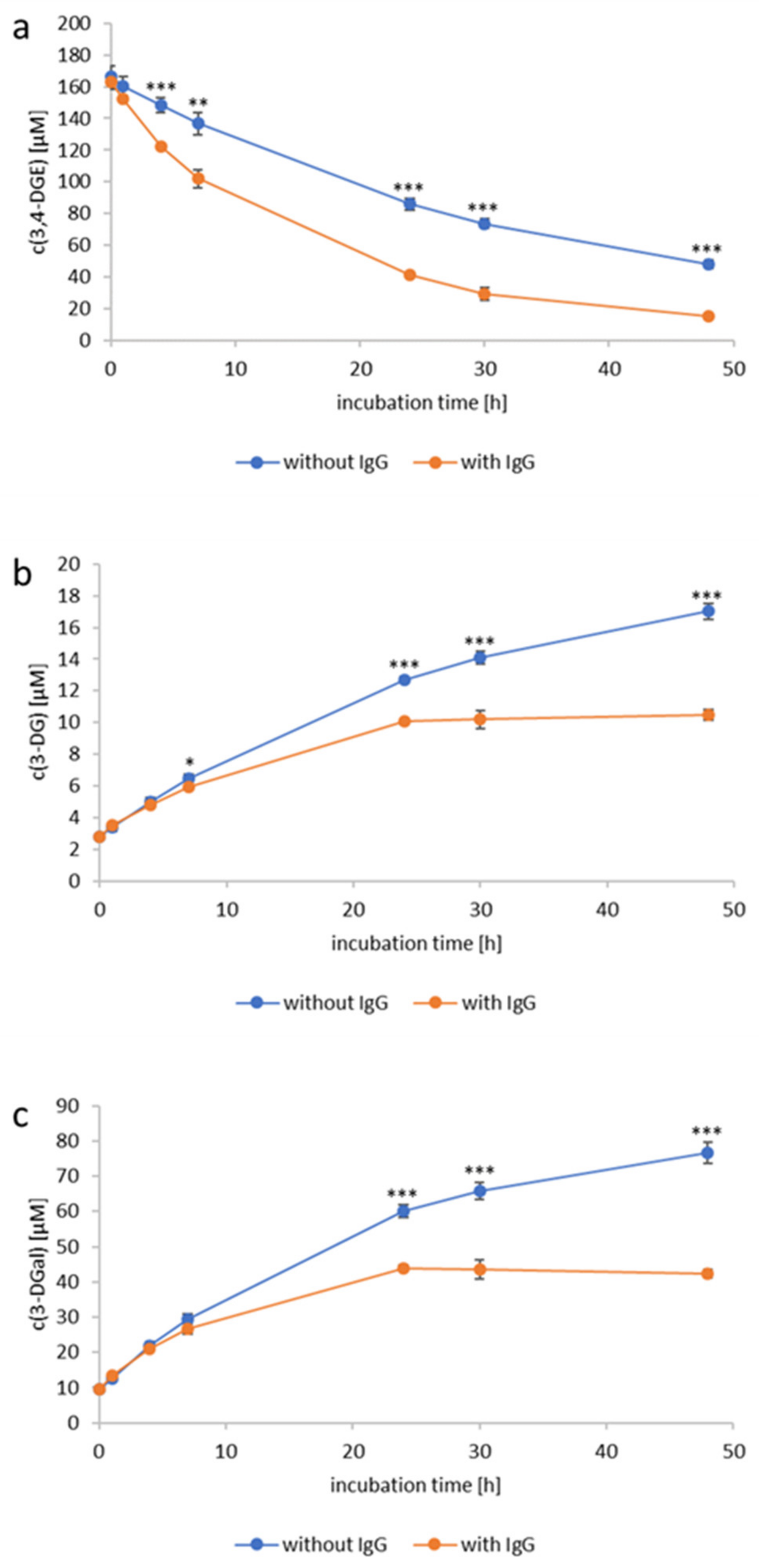
2.4. Reactivity of 3,4-Dideoxyglucosone-3-ene towards Human Immunoglobulin G
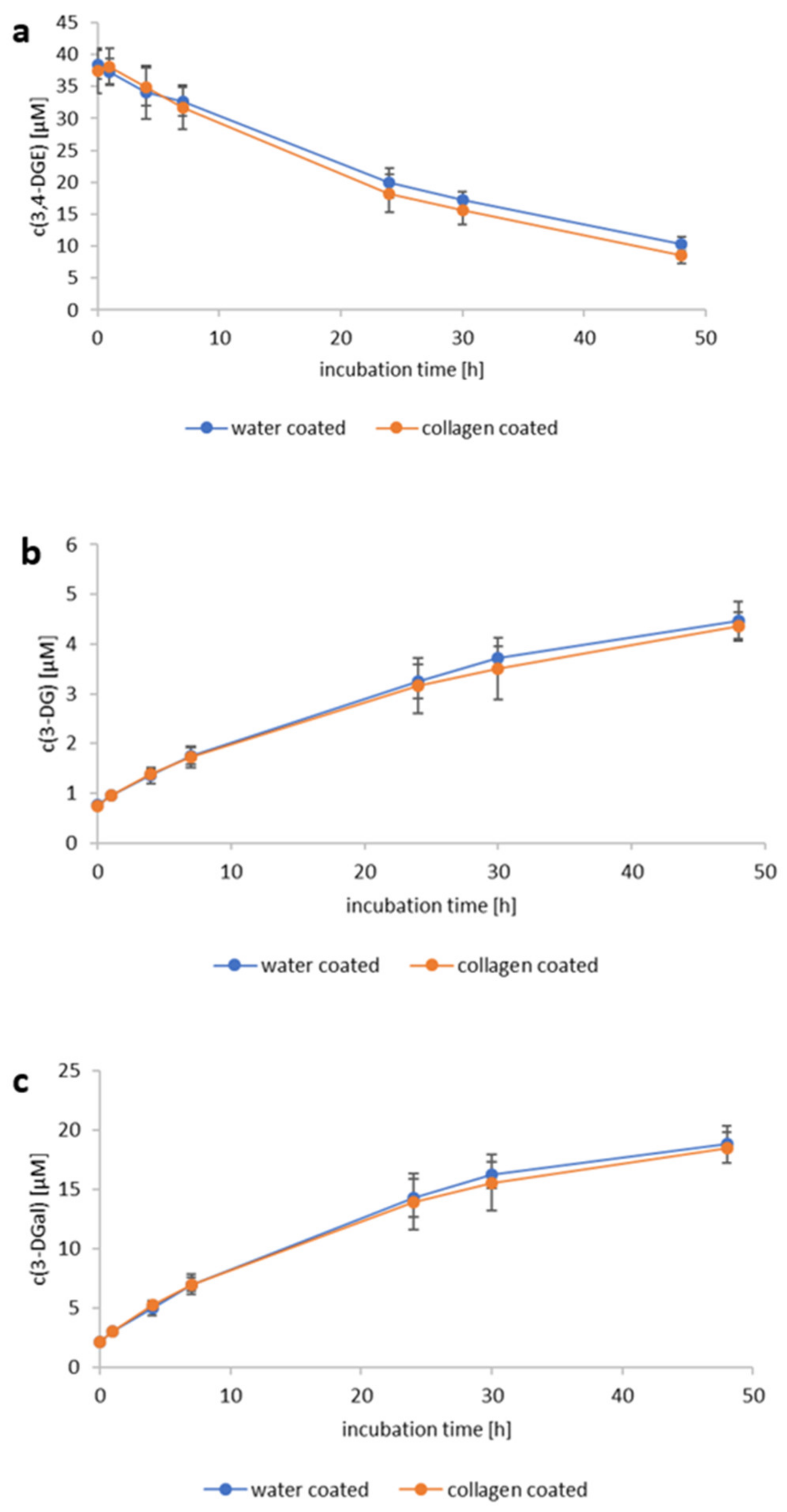
2.5. Reactivity of 3,4-Dideoxyglucosone-3-ene towards Human Collagen Type IV
3. Materials and Methods
3.1. Reagents
3.2. Coating of Eppendorf Tubes with Collagen
3.3. Incubations of 3,4-Dideoxyglucosone-3-ene with Components of the Human Blood Circulatory System
3.4. Quantitative UHPLC-DAD Analysis of Dicarbonyl Compounds and l-Glutathione Adducts
3.5. Qualitative UHPLC-DAD and -MS/MS Analysis of the 3,4-Dideoxyglucosone-3-ene-l-Glutathione Adducts
3.6. Reaction of 3,4-Dideoxyglucosone-3-ene with Human Serum Albumin for Functional Assays
3.7. Analysis of Thiol Content, Ketoprofen-Binding, and Esterase-like Ability of Modified Human Serum Albumin
3.8. Statistics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hellwig, M.; Gensberger-Reigl, S.; Henle, T.; Pischetsrieder, M. Food-derived 1,2-dicarbonyl compounds and their role in diseases. Semin. Cancer Biol. 2018, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Maasen, K.; Scheijen, J.; Opperhuizen, A.; Stehouwer, C.D.A.; Van Greevenbroek, M.M.; Schalkwijk, C.G. Quantification of dicarbonyl compounds in commonly consumed foods and drinks; presentation of a food composition database for dicarbonyls. Food Chem. 2021, 339, 128063. [Google Scholar] [CrossRef] [PubMed]
- Pischetsrieder, M.; Gensberger-Reigl, S.; Atzenbeck, L.; Weigel, I. Chemistry and clinical relevance of carbohydrate degradation in drugs. Drug Discov. Today 2016, 21, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T. 3-Deoxyglucosone: Metabolism, analysis, biological activity, and clinical implication. J. Chromatogr. B Biomed. Sci. Appl. 1999, 731, 23–36. [Google Scholar] [CrossRef]
- Thornalley, P.J. Dicarbonyl intermediates in the Maillard reaction. Ann. N. Y. Acad. Sci. 2005, 1043, 111–117. [Google Scholar] [CrossRef]
- Nursten, H.E. Recent developments in studies of the maillard reaction. Food Chem. 1981, 6, 263–277. [Google Scholar] [CrossRef]
- Mittelmaier, S.; Pischetsrieder, M. Multistep ultrahigh performance liquid chromatography/tandem mass spectrometry analysis for untargeted quantification of glycating activity and identification of most relevant glycation products. Anal. Chem. 2011, 83, 9660–9668. [Google Scholar] [CrossRef]
- Gensberger-Reigl, S.; Atzenbeck, L.; Gottler, A.; Pischetsrieder, M. Identification of [6-hydroxy-2-(hydroxymethyl)-5-oxo-5,6-dihydro-2 H-pyran-3-yl]-cysteine (HHPC) as a cysteine-specific modification formed from 3,4-dideoxyglucosone-3-ene (3,4-DGE). Chem. Res. Toxicol. 2019, 32, 304–311. [Google Scholar] [CrossRef]
- Sanchez-Nino, M.D.; Poveda, J.; Sanz, A.B.; Carrasco, S.; Ruiz-Ortega, M.; Selgas, R.; Egido, J.; Ortiz, A. 3,4-DGE is cytotoxic and decreases HSP27/HSPB1 in podocytes. Arch. Toxicol. 2014, 88, 597–608. [Google Scholar] [CrossRef]
- Catalan, M.P.; Santamaria, B.; Reyero, A.; Ortiz, A.; Egido, J.; Ortiz, A. 3,4-di-deoxyglucosone-3-ene promotes leukocyte apoptosis. Kidney Int. 2005, 68, 1303–1311. [Google Scholar] [CrossRef] [Green Version]
- Distler, L.; Georgieva, A.; Kenkel, I.; Huppert, J.; Pischetsrieder, M. Structure- and concentration-specific assessment of the physiological reactivity of α-dicarbonyl glucose degradation products in peritoneal dialysis fluids. Chem. Res. Toxicol. 2014, 27, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Tomo, T.; Okabe, E.; Namoto, S.; Suzuki, K.; Hirao, Y. Glutathione depletion as a mechanism of 3,4-dideoxyglucosone-3-ene-induced cytotoxicity in human peritoneal mesothelial cells: Role in biocompatibility of peritoneal dialysis fluids. Nephrol. Dial. Transplant. 2009, 24, 1436–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepas, V.; Manig, F.; Mayo, J.C.; Hellwig, M.; Collotta, D.; Sanmartino, V.; Carrocera-Pumarino, R.; Collino, M.; Henle, T.; Sainz, R.M. In vitro evaluation of the toxicological profile and oxidative stress of relevant diet-related advanced glycation end products and related 1,2-dicarbonyls. Oxid. Med. Cell. Longev. 2021, 2021, 9912240. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.K.; Auditore, A.; Pischetsrieder, M.; Messlinger, K.; Fleming, T.; Reeh, P.W.; Sauer, S.K. Reactive dicarbonyl compounds cause Calcitonin Gene-Related Peptide release and synergize with inflammatory conditions in mouse skin and peritoneum. J. Biol. Chem. 2020, 295, 6330–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, B.; Ucero, A.C.; Reyero, A.; Selgas, R.; Ruiz-Ortega, M.; Catalan, M.; Egido, J.; Ortiz, A. 3,4-Dideoxyglucosone-3-ene as a mediator of peritoneal demesothelization. Nephrol. Dial. Transplant. 2008, 23, 3307–3315. [Google Scholar] [CrossRef] [Green Version]
- Kato, F.; Mizukoshi, S.; Aoyama, Y.; Matsuoka, H.; Tanaka, H.; Nakamura, K.; Tsukamoto, M. Immunosuppressive effects of 3, 4-dideoxyglucosone-3-ene, an intermediate in the Maillard reaction. J. Agric. Food Chem. 1994, 42, 2068–2073. [Google Scholar] [CrossRef]
- Hellwig, M.; Henle, T. Baking, ageing, diabetes: A short history of the Maillard reaction. Angew. Chem. Int. Ed. Engl. 2014, 53, 10316–10329. [Google Scholar] [CrossRef]
- Marceau, E.; Yaylayan, V.A. Profiling of alpha-dicarbonyl content of commercial honeys from different botanical origins: Identification of 3,4-dideoxyglucoson-3-ene (3,4-DGE) and related compounds. J. Agric. Food Chem. 2009, 57, 10837–10844. [Google Scholar] [CrossRef]
- Hellwig, M.; Nobis, A.; Witte, S.; Henle, T. Occurrence of (Z)-3,4-dideoxyglucoson-3-ene in different types of beer and malt beer as a result of 3-deoxyhexosone interconversion. J. Agric. Food Chem. 2016, 64, 2746–2753. [Google Scholar] [CrossRef]
- Gensberger, S.; Mittelmaier, S.; Glomb, M.A.; Pischetsrieder, M. Identification and quantification of six major alpha-dicarbonyl process contaminants in high-fructose corn syrup. Anal. Bioanal. Chem. 2012, 403, 2923–2931. [Google Scholar] [CrossRef]
- Gensberger, S.; Glomb, M.A.; Pischetsrieder, M. Analysis of sugar degradation products with α-dicarbonyl structure in carbonated soft drinks by UHPLC-DAD-MS/MS. J. Agric. Food Chem. 2013, 61, 10238–10245. [Google Scholar] [CrossRef] [PubMed]
- Erixon, M.; Linden, T.; Kjellstrand, P.; Carlsson, O.; Ernebrant, M.; Forsback, G.; Wieslander, A.; Jonsson, J.A. PD fluids contain high concentrations of cytotoxic GDPs directly after sterilization. Perit. Dial. Int. 2004, 24, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Bryland, A.; Broman, M.; Erixon, M.; Klarin, B.; Linden, T.; Friberg, H.; Wieslander, A.; Kjellstrand, P.; Ronco, C.; Carlsson, O.; et al. Infusion fluids contain harmful glucose degradation products. Intensive Care Med. 2010, 36, 1213–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erixon, M.; Wieslander, A.; Lindén, T.; Carlsson, O.; Jönsson, J.Å.; Simonsen, O.; Kjellstrand, P. 3, 4-DGE in peritoneal dialysis fluids cannot be found in plasma after infusion into the peritoneal cavity. Perit. Dial. Int. 2008, 28, 277–282. [Google Scholar] [CrossRef]
- Scheijen, J.L.; Schalkwijk, C.G. Quantification of glyoxal, methylglyoxal and 3-deoxyglucosone in blood and plasma by ultra performance liquid chromatography tandem mass spectrometry: Evaluation of blood specimen. Clin. Chem. Lab. Med. 2014, 52, 85–91. [Google Scholar] [CrossRef]
- Tietz, N.W.; Andresen, B.D. Textbook of Clinical Chemistry, 1st ed.; W.B. Saunders: Philadelphia, PA, USA, 1985. [Google Scholar]
- Oettl, K.; Stauber, R.E. Physiological and pathological changes in the redox state of human serum albumin critically influence its binding properties. Br. J. Pharmacol. 2007, 151, 580–590. [Google Scholar] [CrossRef]
- Anguizola, J.; Matsuda, R.; Barnaby, O.S.; Hoy, K.S.; Wa, C.; DeBolt, E.; Koke, M.; Hage, D.S. Review: Glycation of human serum albumin. Clin. Chim. Acta 2013, 425, 64–76. [Google Scholar] [CrossRef] [Green Version]
- Sudlow, G.; Birkett, D.J.; Wade, D.N. The characterization of two specific drug binding sites on human serum albumin. Mol. Pharmacol. 1975, 11, 824–832. [Google Scholar]
- Kurono, Y.; Maki, T.; Yotsuyanagi, T.; Ikeda, K. Esterase-like activity of human serum albumin: Structure-Activity relationships for the reactions with phenyl acetates and p-nitrophenyl esters. Chem. Pharm. Bull. 1979, 27, 2781–2786. [Google Scholar] [CrossRef] [Green Version]
- Mittelmaier, S.; Funfrocken, M.; Fenn, D.; Pischetsrieder, M. 3-Deoxygalactosone, a new glucose degradation product in peritoneal dialysis fluids: Identification, quantification by HPLC/DAD/MSMS and its pathway of formation. Anal. Bioanal. Chem. 2011, 399, 1689–1697. [Google Scholar] [CrossRef]
- Tauer, A.; Bender, T.O.; Fleischmann, E.H.; Niwa, T.; Jorres, A.; Pischetsrieder, M. Fate of the glucose degradation products 3-deoxyglucosone and glyoxal during peritoneal dialysis. Mol. Nutr. Food Res. 2005, 49, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, R.; Becker, M.; Behlke, J.; Billwitz, H.; Böhm, S.; Ebert, B.; Hamann, H.; Krumbiegel, J.; Lassmann, G. Temperature behaviour of human serum albumin. Eur. J. Biochem. 1980, 104, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Baraka-Vidot, J.; Planesse, C.; Meilhac, O.; Militello, V.; van den Elsen, J.; Bourdon, E.; Rondeau, P. Glycation alters ligand binding, enzymatic, and pharmacological properties of human albumin. Biochemistry 2015, 54, 3051–3062. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Tanase, S.; Nakajou, K.; Maruyama, T.; Kragh-Hansen, U.; Otagiri, M. Role of Arg-410 and Tyr-411 in human serum albumin for ligand binding and esterase-like activity. Biochem. J. 2000, 349, 813–819. [Google Scholar] [CrossRef]
- Ahmed, N.; Dobler, D.; Dean, M.; Thornalley, P.J. Peptide mapping identifies hotspot site of modification in human serum albumin by methylglyoxal involved in ligand binding and esterase activity. J. Biol. Chem. 2005, 280, 5724–5732. [Google Scholar] [CrossRef] [Green Version]
- Look, M.P.; Rockstroh, J.K.; Rao, G.S.; Kreuzer, K.A.; Barton, S.; Lemoch, H.; Sudhop, T.; Hoch, J.; Stockinger, K.; Spengler, U.; et al. Serum selenium, plasma glutathione (GSH) and erythrocyte glutathione peroxidase (GSH-Px)-levels in asymptomatic versus symptomatic human immunodeficiency virus-1 (HIV-1)-infection. Eur. J. Clin. Nutr. 1997, 51, 266–272. [Google Scholar] [CrossRef] [Green Version]
- Baillie, T.A.; Davis, M.R. Mass spectrometry in the analysis of glutathione conjugates. Biol. Mass Spectrom. 1993, 22, 319–325. [Google Scholar] [CrossRef]
- Baillie, T.A.; Slatter, J.G. Glutathione—A vehicle for the transport of chemically reactive metabolites in vivo. Acc. Chem. Res. 1991, 24, 264–270. [Google Scholar] [CrossRef]
- Meister, A.; Anderson, M.E. Glutathione. Annu. Rev. Biochem. 1983, 52, 711–760. [Google Scholar] [CrossRef]
- Silverman, R.B.; Holladay, M.W. Drug Metabolism. In The Organic Chemistry of Drug Design and Drug Action, 3rd ed.; Academic Press: San Diego, CA, USA, 2014; pp. 357–422. [Google Scholar]
- Thornalley, P.J. The glyoxalase system: New developments towards functional characterization of a metabolic pathway fundamental to biological life. Biochem. J. 1990, 269, 1–11. [Google Scholar] [CrossRef]
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG subclasses and allotypes: From structure to effector functions. Front. Immunol. 2014, 5, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schauenstein, E.; Sorger, S.; Reiter, M.; Dachs, F. Free thiol groups and labile disulfide bonds in the IgG fraction of human serum. J. Immunol. Methods 1982, 50, 51–56. [Google Scholar] [CrossRef]
- Danze, P.M.; Tarjoman, A.; Rousseaux, J.; Fossati, P.; Dautrevaux, M. Evidence for an increased glycation of IgG in diabetic patients. Clin. Chim. Acta 1987, 166, 143–153. [Google Scholar] [CrossRef]
- Kennedy, D.M.; Skillen, A.W.; Self, C.H. Glycation of monoclonal antibodies impairs their ability to bind antigen. Clin. Exp. Immunol. 1994, 98, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.K.; Hambly, D.M.; Kerwin, B.A.; Treuheit, M.J.; Gadgil, H.S. Characterization of site-specific glycation during process development of a human therapeutic monoclonal antibody. J. Pharm. Sci. 2011, 100, 2543–2550. [Google Scholar] [CrossRef]
- Ligier, S.; Fortin, P.R.; Newkirk, M.M. A new antibody in rheumatoid arthritis targeting glycated IgG: IgM anti-IgG-AGE. Br. J. Rheumatol. 1998, 37, 1307–1314. [Google Scholar] [CrossRef] [Green Version]
- Kühn, K. Basement membrane (type IV) collagen. Matrix Biol. 1995, 14, 439–445. [Google Scholar] [CrossRef]
- Monnier, V.M.; Kohn, R.R.; Cerami, A. Accelerated age-related browning of human collagen in diabetes mellitus. Proc. Natl. Acad. Sci. USA 1984, 81, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.G.; Avery, N.C.; Slatter, D.A.; Sims, T.J.; Bailey, A.J. Isolation and characterization of advanced glycation end products derived from the in vitro reaction of ribose and collagen. Biochem. J. 1998, 330, 1241–1248. [Google Scholar] [CrossRef] [Green Version]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [Green Version]
- Mittelmaier, S.; Funfrocken, M.; Fenn, D.; Berlich, R.; Pischetsrieder, M. Quantification of the six major alpha-dicarbonyl contaminants in peritoneal dialysis fluids by UHPLC/DAD/MSMS. Anal. Bioanal. Chem. 2011, 401, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Danaei, G.; Finucane, M.M.; Lu, Y.; Singh, G.M.; Cowan, M.J.; Paciorek, C.J.; Lin, J.K.; Farzadfar, F.; Khang, Y.-H.; Stevens, G.A.; et al. National, regional, and global trends in fasting plasma glucose and diabetes prevalence since 1980: Systematic analysis of health examination surveys and epidemiological studies with 370 country-years and 2.7 million participants. Lancet 2011, 378, 31–40. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Eyer, P.; Worek, F.; Kiderlen, D.; Sinko, G.; Stuglin, A.; Simeon-Rudolf, V.; Reiner, E. Molar absorption coefficients for the reduced Ellman reagent: Reassessment. Anal. Biochem. 2003, 312, 224–227. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Auditore, A.; Gensberger-Reigl, S.; Pischetsrieder, M. In Vitro Reactivity of the Glucose Degradation Product 3,4-Dideoxyglucosone-3-ene (3,4-DGE) towards Abundant Components of the Human Blood Circulatory System. Int. J. Mol. Sci. 2022, 23, 4557. https://doi.org/10.3390/ijms23094557
Auditore A, Gensberger-Reigl S, Pischetsrieder M. In Vitro Reactivity of the Glucose Degradation Product 3,4-Dideoxyglucosone-3-ene (3,4-DGE) towards Abundant Components of the Human Blood Circulatory System. International Journal of Molecular Sciences. 2022; 23(9):4557. https://doi.org/10.3390/ijms23094557
Chicago/Turabian StyleAuditore, Andrea, Sabrina Gensberger-Reigl, and Monika Pischetsrieder. 2022. "In Vitro Reactivity of the Glucose Degradation Product 3,4-Dideoxyglucosone-3-ene (3,4-DGE) towards Abundant Components of the Human Blood Circulatory System" International Journal of Molecular Sciences 23, no. 9: 4557. https://doi.org/10.3390/ijms23094557
APA StyleAuditore, A., Gensberger-Reigl, S., & Pischetsrieder, M. (2022). In Vitro Reactivity of the Glucose Degradation Product 3,4-Dideoxyglucosone-3-ene (3,4-DGE) towards Abundant Components of the Human Blood Circulatory System. International Journal of Molecular Sciences, 23(9), 4557. https://doi.org/10.3390/ijms23094557