
Development and Characterization of Indole-Responsive Whole-Cell Biosensor Based on the Inducible Gene Expression System from Pseudomonas putida KT2440

 , , and
, , and
Abstract
:1. Introduction
2. Results
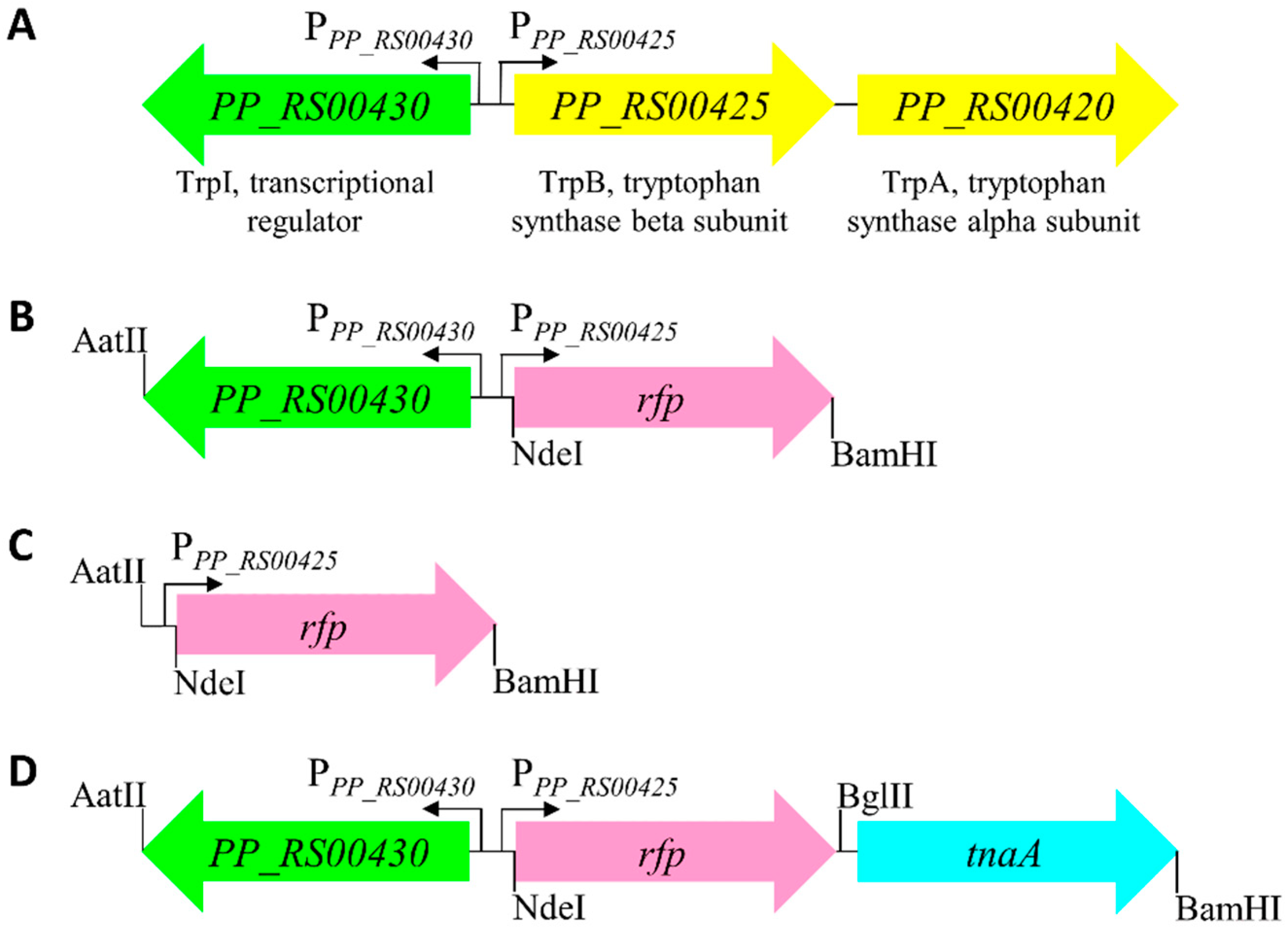
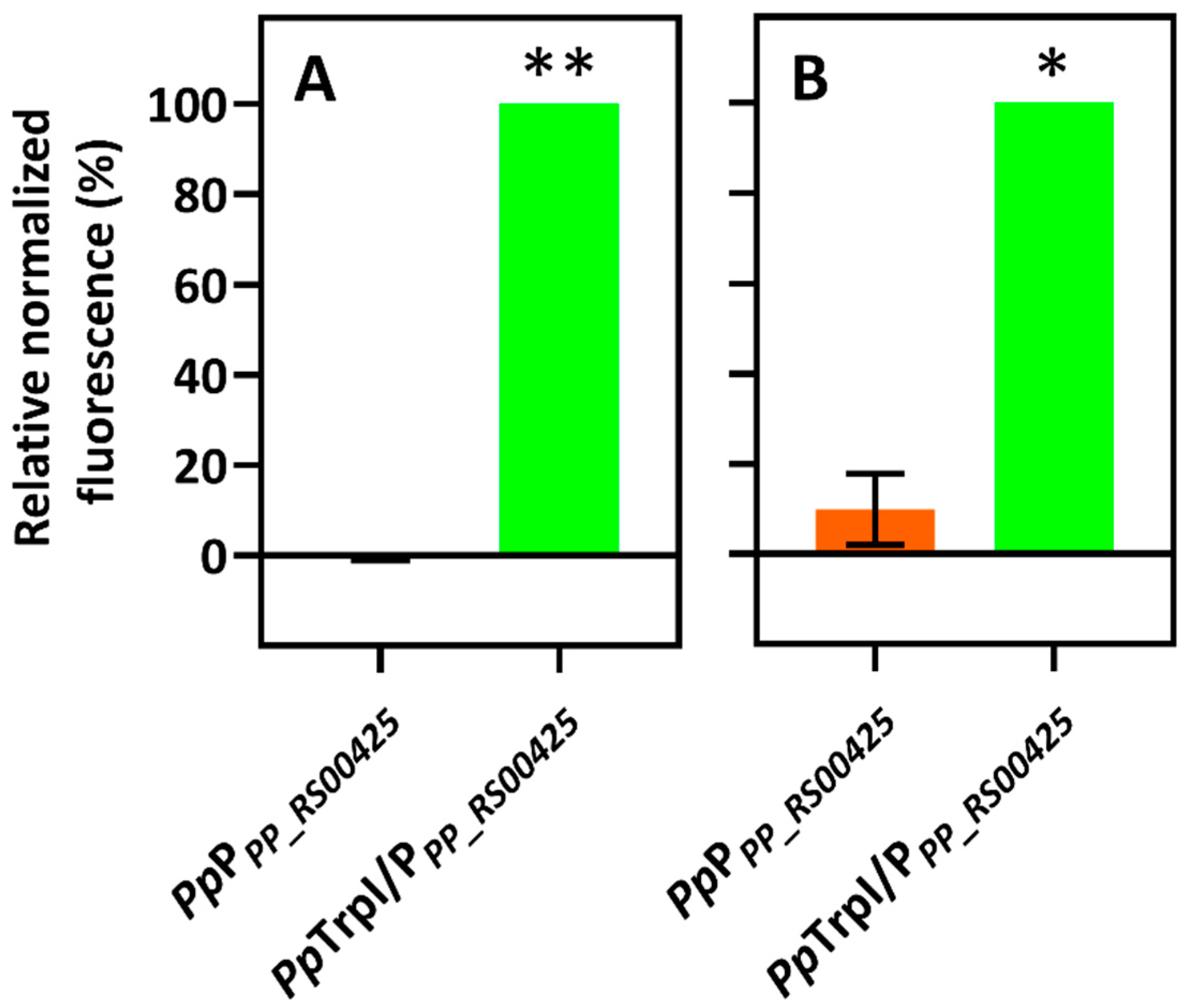
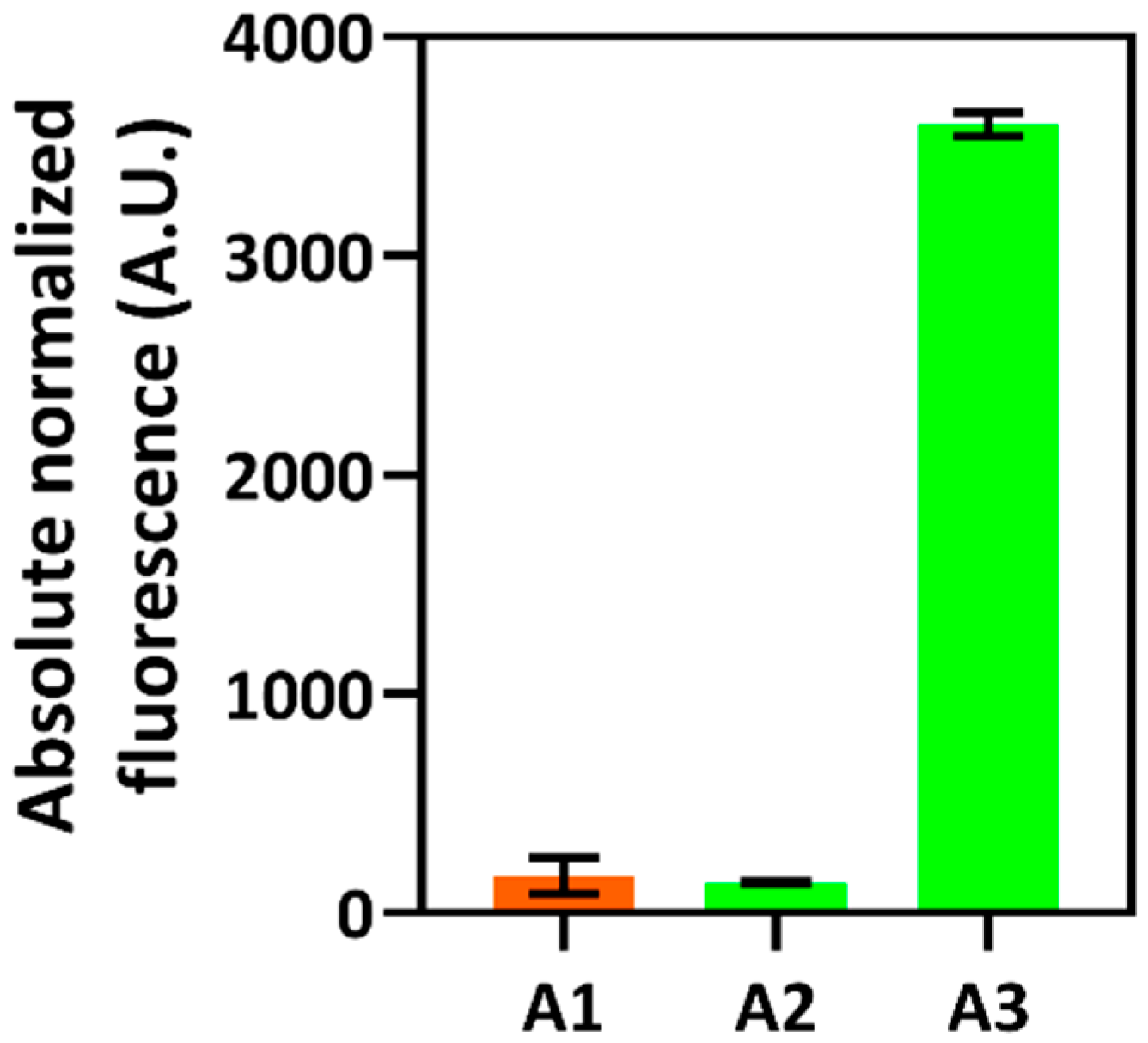
2.1. Identification of Indole-Inducible System and Biosensor Development
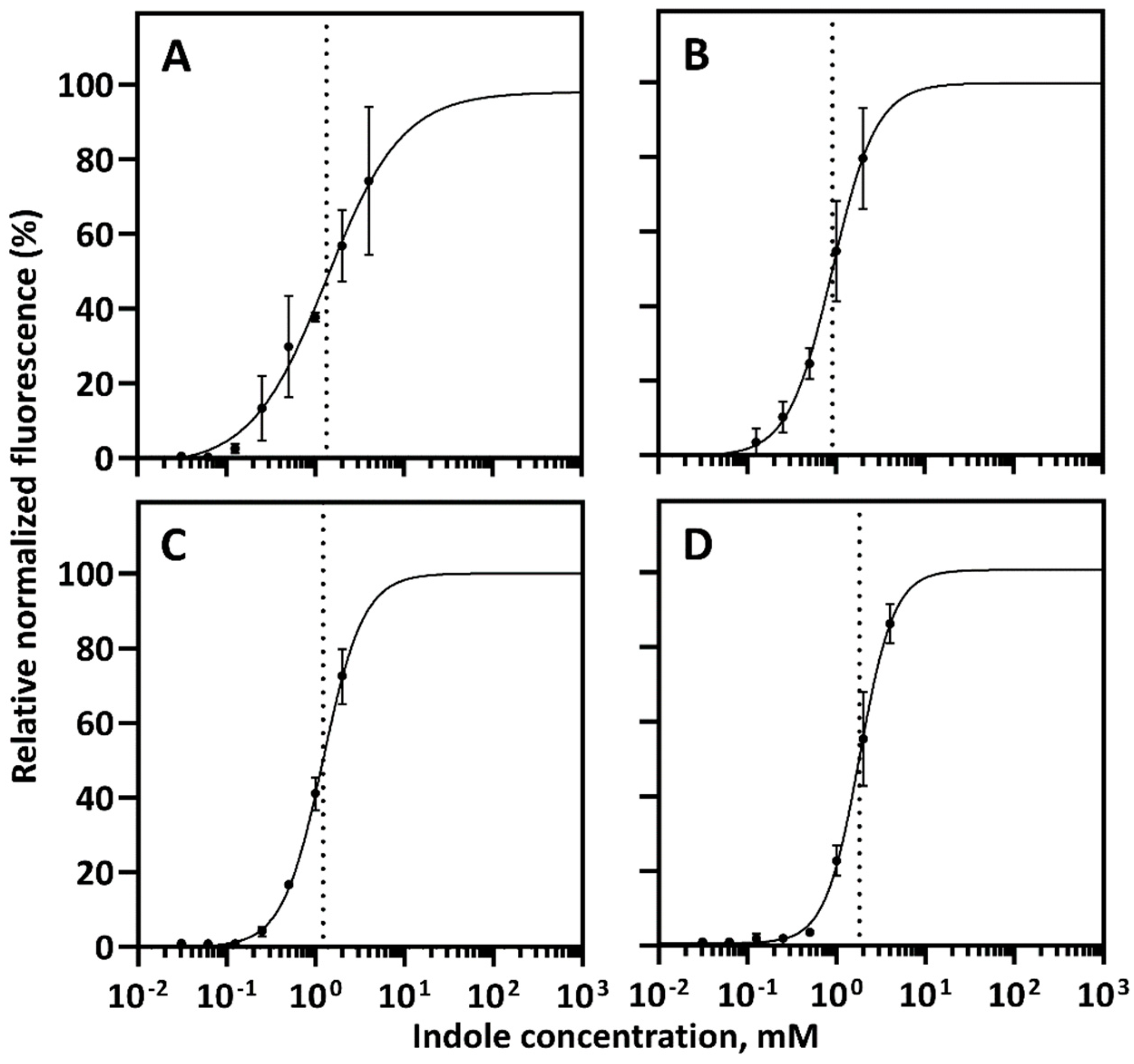
2.2. Characterization of Indole-Inducible Biosensors
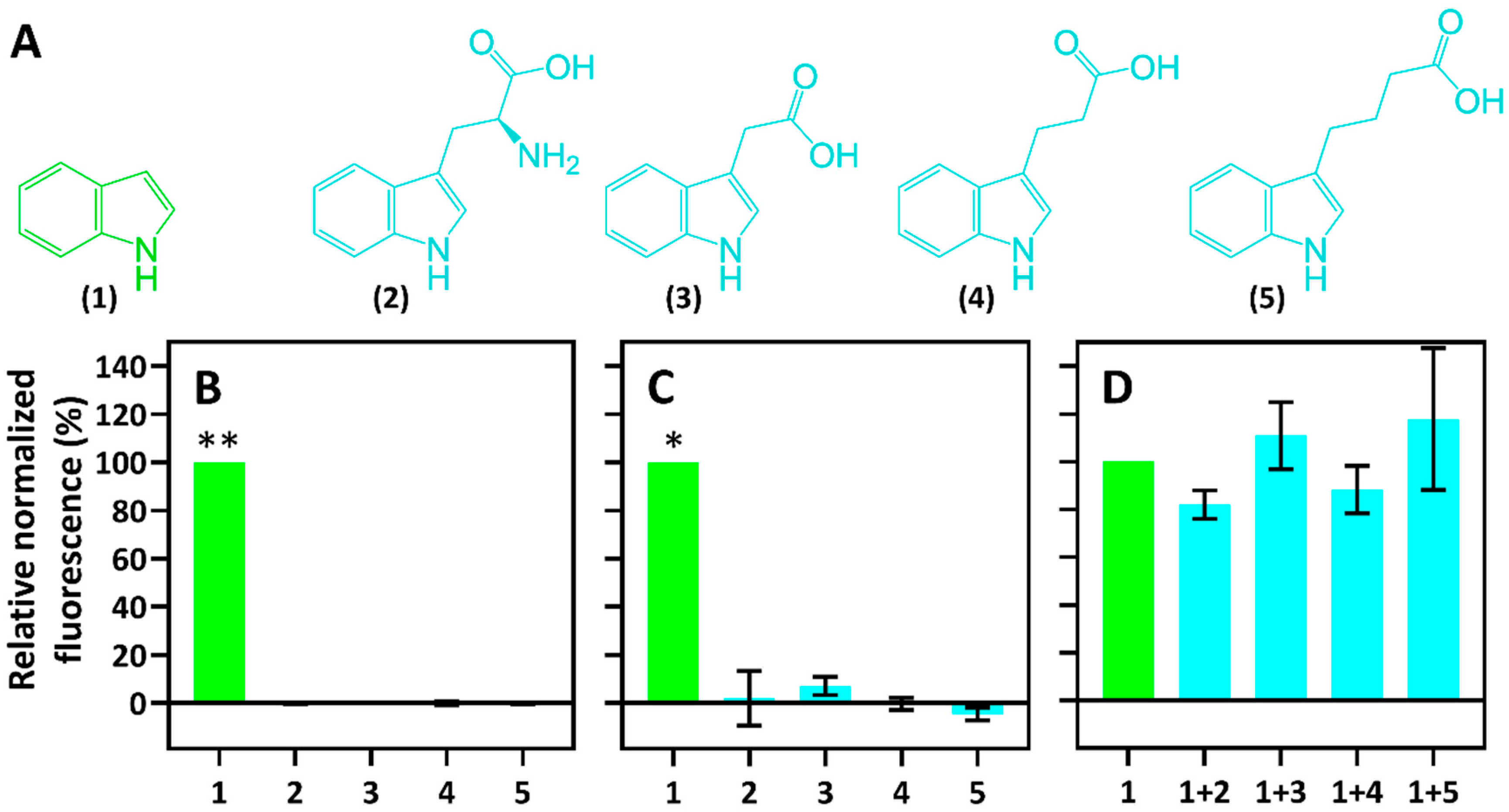
2.3. Specificity of Indole-Inducible Biosensors
2.4. Application of Whole-Cell Biosensor
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Bacterial Strains and Media
4.3. Cloning and Transformation
4.4. Plasmid Construction
4.5. Fluorescence and Absorbance Measurements
4.6. Non-Linear Least-Squares Fitting and Dynamic Range Calculation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zarkan, A.; Liu, J.; Matuszewska, M.; Gaimster, H.; Summers, D.K. Local and universal action: The paradoxes of indole signalling in bacteria. Trends Microbiol. 2020, 28, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zheng, X.; Ji, H.; Wang, T.L.; Xing, X.H.; Zhang, C. Dynamics of transcription–translation coordination tune bacterial indole signaling. Nat. Chem. Biol. 2020, 16, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sperandio, V. Indole signaling at the host-microbiota-pathogen interface. MBio 2019, 10, e01031-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, H.L.; Liu, J.; Fang, W.Y.; Ravindar, L.; Rakesh, K.P. Indole-based derivatives as potential antibacterial activity against methicillin-resistance Staphylococcus aureus (MRSA). Eur. J. Med. Chem. 2020, 194, 112245. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhang, X.; Qu, Y. Biodegradation and biotransformation of indole: Advances and perspectives. Front. Microbiol. 2018, 9, 2625. [Google Scholar] [CrossRef] [PubMed]
- Barden, T.C. Indoles: Industrial, agricultural and over-the-counter uses. Heterocycl. Chem. 2010, 31–46. [Google Scholar]
- Gupta, N.; Goyal, D. Synthesis of indole and its derivatives in water. Chem. Heterocycl. Compd. 2015, 51, 4–16. [Google Scholar] [CrossRef]
- Dhuguru, J.; Skouta, R. Role of indole scaffolds as pharmacophores in the development of anti-lung cancer agents. Molecules 2020, 25, 1615. [Google Scholar] [CrossRef] [Green Version]
- De Klerk, G.J.; Van Der Krieken, W.; De Jong, J.C. The formation of adventitious roots: New concepts, new possibilities. In Vitro Cell. Dev. Biol.-Plant 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Colella, M.; Degennaro, L.; Luisi, R. Continuous flow synthesis of heterocycles: A recent update on the flow synthesis of indoles. Molecules 2020, 25, 3242. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; López-Martínez, L.X.; Contreras-Angulo, L.A.; Elizalde-Romero, C.A.; Heredia, J.B. Plant Alkaloids: Structures and Bioactive Properties. In Plant-derived Bioactives; Swamy, M.K., Ed.; Springer: Singapore, 2020; pp. 85–117. [Google Scholar]
- Hamid, H.A.; Ramli, A.N.M.; Yusoff, M.M. Indole alkaloids from plants as potential leads for antidepressant drugs: A mini review. Front. Pharmacol. 2017, 8, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonaci, F.; Ghiotto, N.; Wu, S.; Pucci, E.; Costa, A. recent advances in migraine therapy. Springerplus 2016, 5, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, G.R.; Kuethe, J.T. Practical methodologies for the synthesis of indoles. Chem. Rev. 2006, 106, 2875–2911. [Google Scholar] [CrossRef] [PubMed]
- Inman, M.; Moody, C.J. Indole synthesis-something old, something new. Chem. Sci. 2013, 4, 29–41. [Google Scholar] [CrossRef]
- Hopkins, F.G.; Cole, S.W. A contribution to the chemistry of proteids: Part II. The constitution of tryptophane, and the action of bacteria upon It. J. Physiol. 1903, 29, 451–466. [Google Scholar] [CrossRef]
- Lee, J.H.; Wood, T.K.; Lee, J. Roles of indole as an interspecies and interkingdom signaling molecule. Trends Microbiol. 2015, 23, 707–718. [Google Scholar] [CrossRef]
- Han, T.H.; Lee, J.H.; Cho, M.H.; Wood, T.K.; Lee, J. Environmental factors affecting indole production in Escherichia coli. Res. Microbiol. 2011, 162, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Hua, Y.; Zhang, B.; Sun, L.; Li, W.; Kong, X.; Hong, J. Metabolic engineering of indole pyruvic acid biosynthesis in Escherichia coli with TdiD. Microb. Cell Fact. 2017, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Berry, A.; Dodge, T.; Pepsin, M.; Weyler, W. Application of metabolic engineering to improve both the production and use of biotech indigo. J. Ind. Microbiol. Biotechnol. 2002, 28, 127–133. [Google Scholar] [CrossRef]
- Yanofsky, C. Tryptophan synthetase from Neurospora: Indole+L-serine→L-tryptophan. Methods Enzymol. 1955, 2, 233–238. [Google Scholar]
- Michalska, K.; Gale, J.; Joachimiak, G.; Chang, C.; Hatzos-Skintges, C.; Nocek, B.; Johnston, S.E.; Bigelow, L.; Bajrami, B.; Jedrzejczak, R.P.; et al. Conservation of the structure and function of bacterial tryptophan synthases. IUCrJ 2019, 6, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Merino, E.; Jensen, R.A.; Yanofsky, C. Evolution of bacterial Trp operons and their regulation. Curr. Opin. Microbiol. 2008, 11, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.; Crawford, I.P. The roles of indoleglycerol phosphate and the TrpI protein in the expression of trpBA from Pseudomonas aeruginosa. Nucleic Acids Res. 1990, 18, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Ehmann, A.; Pathology, P.; Lansing, E.; Mich, U.S.A. The van URK-Salkowski reagent—A sensitive and specific chromogenic reagent for silica gel thin-layer chromatographic detection and identification of indole derivatives. J. Chromatogr. 1977, 132, 267–276. [Google Scholar] [CrossRef]
- Darkoh, C.; Chappell, C.; Gonzales, C.; Okhuysen, P. A rapid and specific method for the detection of indole in complex biological samples. Appl. Environ. Microbiol. 2015, 81, 8093–8097. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, S.; Xu, J.; Acosta, K.; Poulev, A.; Lebeis, S.; Lam, E. Bacterial production of indole related compounds reveals their role in association between duckweeds and endophytes. Front. Chem. 2018, 6, 265. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.A.; Ho, J.M.L.; Bennett, M.R. Strategies for improving small-molecule biosensors in bacteria. Biosensors 2022, 12, 64. [Google Scholar] [CrossRef]
- Moraskie, M.; Roshid, M.H.O.; O’Connor, G.; Dikici, E.; Zingg, J.-M.; Deo, S.; Daunert, S. Microbial whole-cell biosensors: Current applications, challenges, and future perspectives. Biosens. Bioelectron. 2021, 191, 113359. [Google Scholar] [CrossRef]
- 30. Kim, N.M.; Sinnott, R.W.; Sandoval, N.R. Transcription factor-based biosensors and inducible systems in non-model bacteria: Current progress and future directions. Curr. Opin. Biotechnol. 2020, 64, 39–46. [Google Scholar] [CrossRef]
- Mitchler, M.M.; Garcia, J.M.; Montero, N.E.; Williams, G.J. Transcription factor-based biosensors: A molecular-guided approach for natural product engineering. Curr. Opin. Biotechnol. 2021, 69, 172–181. [Google Scholar] [CrossRef]
- 32. Ding, N.; Zhou, S.; Deng, Y. Transcription-factor-based biosensor engineering for applications in synthetic biology. ACS Synth. Biol. 2021, 10, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.K.; Church, G.M. Genetically encoded sensors enable real-time observation of metabolite production. Proc. Natl. Acad. Sci. USA 2016, 113, 2388–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, I.P.; Gunsalus, I.C. Inducibility of tryptophan synthetase in Pseudomonas putida. Proc. Natl. Acad. Sci. USA 1966, 56, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, J.I.; Nogales, J.; García, J.L.; Díaz, E. A genomic view of the catabolism of aromatic compounds in Pseudomonas. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1297–1325. [Google Scholar]
- Hanko, E.K.R.; Minton, N.P.; Malys, N. Design, Cloning and characterization of transcription factor-based inducible gene expression systems. Methods Enzymol. 2019, 621, 153–169. [Google Scholar]
- Hanko, E.K.R.; Paiva, A.C.; Jonczyk, M.; Abbott, M.; Minton, N.P.; Malys, N. A genome-wide approach for identification and characterisation of metabolite-inducible systems. Nat. Commun. 2020, 11, 1213. [Google Scholar] [CrossRef] [Green Version]
- Alagesan, S.; Hanko, E.K.R.; Malys, N.; Ehsaan, M.; Winzer, K.; Minton, N.P. Functional genetic elements for controlling gene expression in Cupriavidus necator H16. Appl. Environ. Microbiol. 2018, 84, e00878-18. [Google Scholar] [CrossRef] [Green Version]
- Sadauskas, M.; Vaitekunas, J.; Gasparavičiute, R.; Meškys, R. Indole biodegradation in Acinetobacter sp. strain O153: Genetic and biochemical characterization. Appl. Environ. Microbiol. 2017, 83, e01453-17. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Ma, Q.; Liu, Z.; Wang, W.; Tang, H.; Zhou, J.; Xu, P. Unveiling the biotransformation mechanism of indole in a Cupriavidus sp. strain. Mol. Microbiol. 2017, 106, 905–918. [Google Scholar] [CrossRef] [Green Version]
- Newton, W.A.; Morino, Y.; Snell, E.E. Properties of crystalline tryptophanase. J. Biol. Chem. 1965, 240, 1211–1218. [Google Scholar] [CrossRef]
- Shen, T.; Liu, Q.; Xie, X.; Xu, Q.; Chen, N. Improved production of tryptophan in genetically engineered Escherichia coli with TktA and PpsA overexpression. J. Biomed. Biotechnol. 2012, 2012, 605219. [Google Scholar] [CrossRef] [Green Version]
- Arza, C.R.; Zhang, B. Synthesis, thermal properties, and rheological characteristics of indole-based aromatic polyesters. ACS Omega 2019, 4, 15012–15021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herud-Sikimić, O.; Stiel, A.C.; Kolb, M.; Shanmugaratnam, S.; Berendzen, K.W.; Feldhaus, C.; Höcker, B.; Jürgens, G. A Biosensor for the direct visualization of auxin. Nature 2021, 592, 768–772. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Childers, W.S. A Biosensor for detection of indole metabolites. ACS Synth. Biol. 2021, 10, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Loughrin, J.H.; Lovanh, N.; Mahmood, R. Equilibrium sampling used to monitor malodors in a swine waste lagoon. J. Environ. Qual. 2008, 37, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tesso, T.A.; Liu, G. Indole degradation in a model system and in poultry manure by Acinetobacter spp. Appl. Sci. 2019, 9, 1622. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W.; Irwin, C.A.; Laboratory, C.S.H.; Fund, R.E.C.M.; Janssen, K.A. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Ausubel, F.M. Current Protocols in Molecular Biology; John Wiley & Sons Inc.: New York, NY, USA, 2003. [Google Scholar]
- Hanko, E.K.R.; Minton, N.P.; Malys, N. Characterisation of a 3-hydroxypropionic acid-inducible system from Pseudomonas putida for orthogonal gene expression control in Escherichia coli and Cupriavidus necator. Sci. Rep. 2017, 7, 1724. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whole-Cell Biosensor | Growth Conditions | Dynamic Range, fold | Km, mM |
|---|---|---|---|
| E. coli/PpTrpI/PPP_RS00425 | LB medium, 30 °C | 373.5 | 1.207 |
| E. coli/PpTrpI/PPP_RS00425 | Minimal medium, 30 °C | 639.6 | 1.347 |
| C. necator/PpTrpI/PPP_RS00425 | LB medium, 30 °C | 101.4 | 1.819 |
| C. necator/PpTrpI/PPP_RS00425 | Minimal medium, 30 °C | 11.9 | 0.9055 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matulis, P.; Kutraite, I.; Augustiniene, E.; Valanciene, E.; Jonuskiene, I.; Malys, N. Development and Characterization of Indole-Responsive Whole-Cell Biosensor Based on the Inducible Gene Expression System from Pseudomonas putida KT2440. Int. J. Mol. Sci. 2022, 23, 4649. https://doi.org/10.3390/ijms23094649
Matulis P, Kutraite I, Augustiniene E, Valanciene E, Jonuskiene I, Malys N. Development and Characterization of Indole-Responsive Whole-Cell Biosensor Based on the Inducible Gene Expression System from Pseudomonas putida KT2440. International Journal of Molecular Sciences. 2022; 23(9):4649. https://doi.org/10.3390/ijms23094649
Chicago/Turabian StyleMatulis, Paulius, Ingrida Kutraite, Ernesta Augustiniene, Egle Valanciene, Ilona Jonuskiene, and Naglis Malys. 2022. "Development and Characterization of Indole-Responsive Whole-Cell Biosensor Based on the Inducible Gene Expression System from Pseudomonas putida KT2440" International Journal of Molecular Sciences 23, no. 9: 4649. https://doi.org/10.3390/ijms23094649
APA StyleMatulis, P., Kutraite, I., Augustiniene, E., Valanciene, E., Jonuskiene, I., & Malys, N. (2022). Development and Characterization of Indole-Responsive Whole-Cell Biosensor Based on the Inducible Gene Expression System from Pseudomonas putida KT2440. International Journal of Molecular Sciences, 23(9), 4649. https://doi.org/10.3390/ijms23094649