
Expression of the AHPND Toxins PirAvp and PirBvp Is Regulated by Components of the Vibrio parahaemolyticus Quorum Sensing (QS) System

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Results
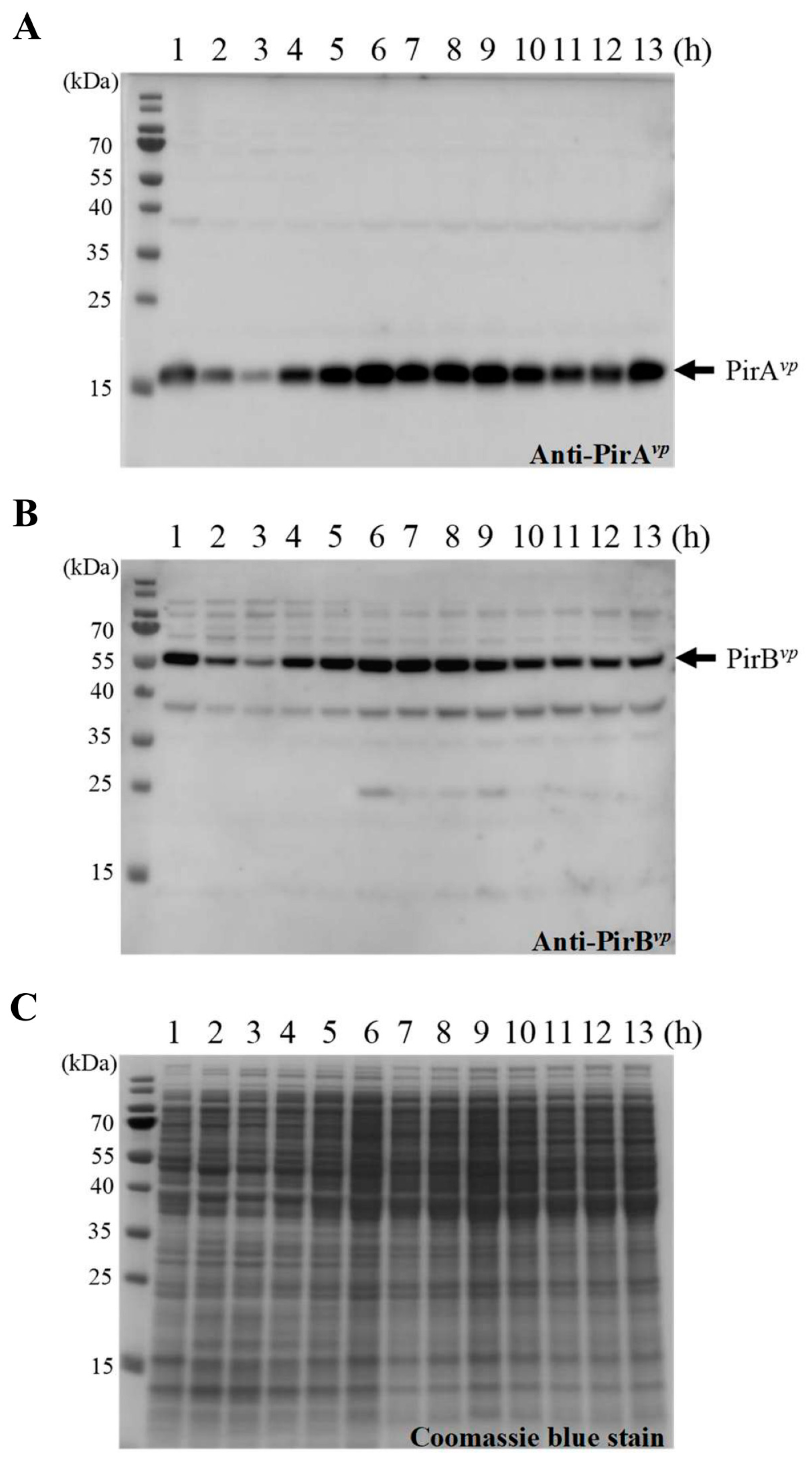
2.1. The Expression Levels of pirAvp/pirBvp during Different Growth Phases of V. parahaemolyticus
2.2. LuxO Is a Negative Regulator for the Expression of pirAvp and pirBvp
2.3. The QS Transcription Factor AphBvp Is a Possible Regulator for pirAvp and pirBvp
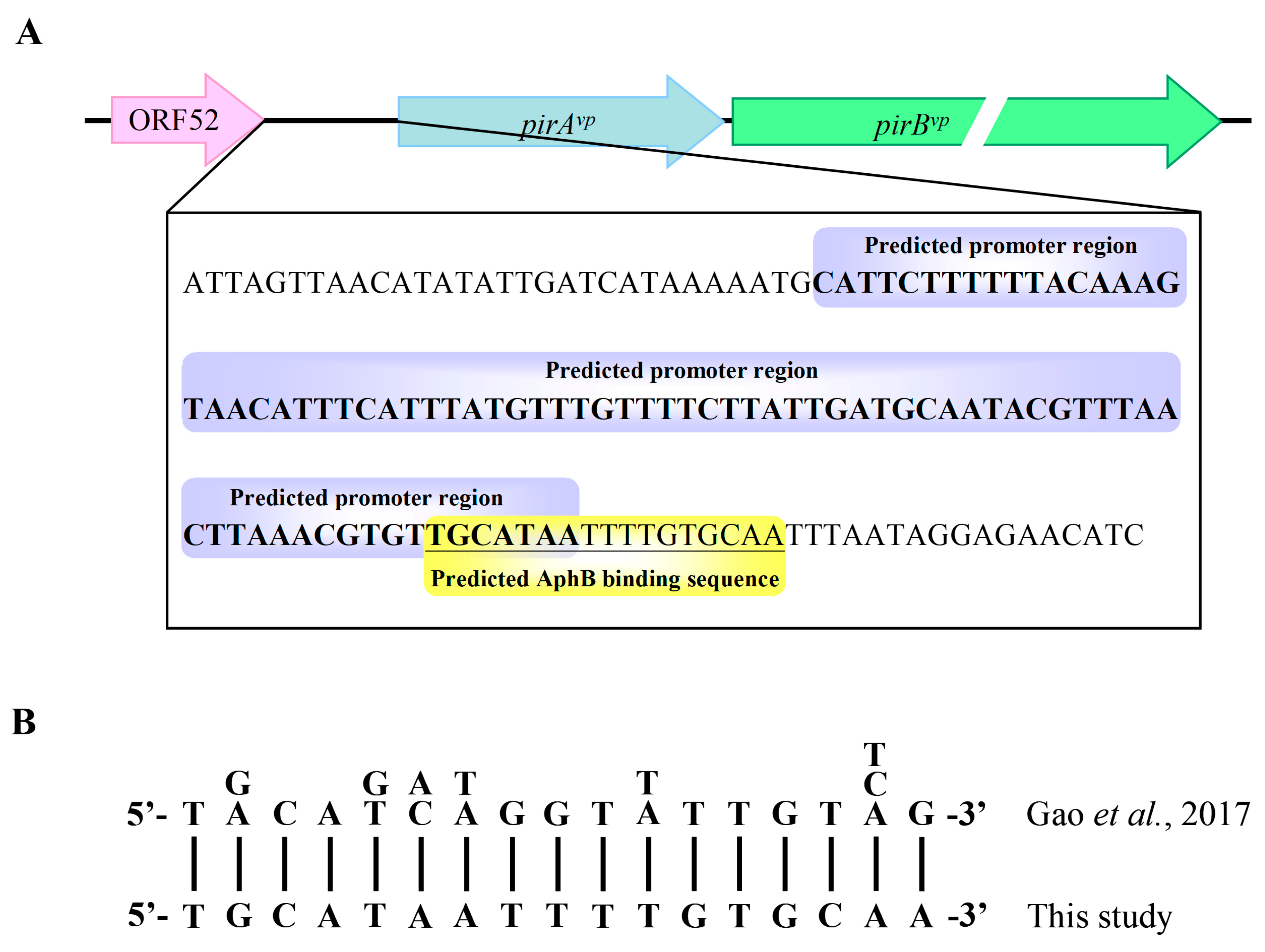
2.4. His-AphBvp Binds with the Predicted Promoter Region of pirAvp/pirBvp
3. Discussion
4. Materials and Methods
4.1. Growth Curve Measurement and Sample Collection
4.2. RNA Extraction and Real-Time PCR
4.3. Protein Extraction and Western Blots
4.4. Construction of the ΔopaR, ΔluxO, ΔopaRΔluxO, and luxOD47E Mutants
4.5. Confirmation of the ΔopaR, luxO, ΔopaRΔluxO and luxOD47E Mutants by PCR
4.6. Plasmid Construction for Recombinant Protein Expression
4.7. Expression and Purification of Recombinant His-AphA and His-AphB
4.8. Electrophoretic Mobility Shift Assay (EMSA)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera vibrios: The microbial barometer of climate change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [Green Version]
- McCarter, L. The multiple identities of Vibrio parahaemolyticus. J. Mol. Microbiol. Biotechnol. 1999, 1, 51–57. [Google Scholar]
- Newton, A.; Kendall, M.; Vugia, D.J.; Henao, O.L.; Mahon, B.E. Increasing rates of vibriosis in the United States, 1996–2010: Review of surveillance data from 2 systems. Clin. Infect. Dis. 2012, 54, S391–S395. [Google Scholar] [CrossRef]
- Broberg, C.A.; Calder, T.J.; Orth, K. Vibrio parahaemolyticus cell biology and pathogenicity determinants. Microbes Infect. 2011, 13, 992–1001. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Ono, T.; Rokuda, M.; Jang, M.H.; Okada, K.; Iida, T.; Honda, T. Functional characterization of two type III secretion systems of Vibrio parahaemolyticus. Infect. Immun. 2004, 72, 6659–6665. [Google Scholar] [CrossRef] [Green Version]
- Salomon, D.; Gonzalez, H.; Updegraff, B.L.; Orth, K. Vibrio parahaemolyticus type VI secretion system 1 is activated in marine conditions to target bacteria, and is differentially regulated from system 2. PLoS ONE 2013, 8, e61086. [Google Scholar] [CrossRef]
- Tran, L.; Nunan, L.; Redman, R.M.; Mohney, L.L.; Pantoja, C.R.; Fitzsimmons, K.; Lightner, D.V. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Dis. Aquat. Organ. 2013, 105, 45–55. [Google Scholar] [CrossRef]
- NACA-FAO. Quarterly Aquatic Animal Disease Report (Asia and Pacific Region), April–June 2011; NACA: Bangkok, Thailand, 2011. [Google Scholar]
- Lightner, D.V.; Redman, R.M.; Pantoja, C.R.; Noble, B.L.; Tran, L. Early mortality syndrome affects shrimp in Asia. Glob. Aquac. Advocate 2012, 15, 40. [Google Scholar]
- Nunan, L.; Lightner, D.; Pantoja, C.; Gomez-Jimenez, S. Detection of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Dis. Aquat. Organ. 2014, 111, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Lee, C.T.; Chen, I.T.; Yang, Y.T.; Ko, T.P.; Huang, Y.T.; Huang, J.Y.; Huang, M.F.; Lin, S.J.; Chen, C.Y.; Lin, S.S.; et al. The opportunistic marine pathogen Vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proc. Natl. Acad. Sci. USA 2015, 112, 10798–10803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.J.; Hsu, K.C.; Wang, H.C. Structural insights into the cytotoxic mechanism of Vibrio parahaemolyticus PirAvp and PirBvp toxins. Mar. Drugs 2017, 15, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.J.; Chen, Y.F.; Hsu, K.C.; Chen, Y.L.; Ko, T.P.; Lo, C.F.; Wang, H.C.; Wang, H.C. Structural insights to the heterotetrameric interaction between the Vibrio parahaemolyticus PirAvp and PirBvp toxins and activation of the Cry-like pore-forming domain. Toxins 2019, 11, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, S.T.; van Kessel, J.C.; Shao, Y.; Bassler, B.L. AphA and LuxR/HapR reciprocally control quorum sensing in vibrios. Genes Dev. 2011, 25, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Kalburge, S.S.; Carpenter, M.R.; Rozovsky, S.; Boyd, E.F. Quorum sensing regulators are required for metabolic fitness in Vibrio parahaemolyticus. Infect. Immun. 2017, 85, e00930-16. [Google Scholar] [CrossRef] [Green Version]
- Paopradit, P.; Tansila, N.; Surachat, K.; Mittraparp-Arthorn, P. Vibrio alginolyticus influences quorum sensing-controlled phenotypes of acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus. PeerJ 2021, 9, e11567. [Google Scholar] [CrossRef]
- Kovacikova, G.; Skorupski, K. Binding site requirements of the virulence gene regulator AphB: Differential affinities for the Vibrio cholerae classical and El Tor tcpPH promoters. Mol. Microbiol. 2002, 44, 533–547. [Google Scholar] [CrossRef]
- Gao, X.; Liu, Y.; Liu, H.; Yang, Z.; Liu, Q.; Zhang, Y.; Wang, Q. Identification of the regulon of AphB and its essential roles in LuxR and exotoxin Asp expression in the pathogen Vibrio alginolyticus. J. Bacteriol. 2017, 199, e00252-17. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.T.; Amaro, C.; Wu, K.M.; Valiente, E.; Chang, Y.F.; Tsai, S.F.; Chang, C.H.; Hor, L.I. A common virulence plasmid in biotype 2 Vibrio vulnificus and its dissemination aided by a conjugal plasmid. J. Bacteriol. 2008, 190, 1638–1648. [Google Scholar] [CrossRef] [Green Version]
- Luangtrakul, W.; Boonchuen, P.; Jaree, P.; Kumar, R.; Wang, H.C.; Somboonwiwat, K. Cytotoxicity of Vibrio parahaemolyticus AHPND toxin on shrimp hemocytes, a newly identified target tissue, involves binding of toxin to aminopeptidase N1 receptor. PLoS Pathog. 2021, 17, e1009463. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Li, R.; Aweya, J.J.; Yao, D.; Wang, F.; Li, S.; Tuan, T.N.; Zhang, Y. The PirB toxin protein from Vibrio parahaemolyticus induces apoptosis in hemocytes of Penaeus vannamei. Virulence 2021, 12, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Henke, J.M.; Bassler, B.L. Quorum sensing regulates type III secretion in Vibrio harveyi and Vibrio parahaemolyticus. J. Bacteriol. 2004, 186, 3794–3805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhou, D.; Mao, P.; Zhang, Y.; Hou, J.; Hu, Y.; Li, J.; Hou, S.; Yang, R.; Wang, R.; et al. Cell density- and quorum sensing-dependent expression of type VI secretion system 2 in Vibrio parahaemolyticus. PLoS ONE 2013, 8, e73363. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Hu, L.; Qiu, Y.; Osei-Adjei, G.; Tang, H.; Zhang, Y.; Zhang, R.; Sheng, X.; Xu, S.; Yang, W.; et al. QsvR integrates into quorum sensing circuit to control Vibrio parahaemolyticus virulence. Environ. Microbiol. 2019, 21, 1054–1067. [Google Scholar] [CrossRef]
- Tu, K.C.; Bassler, B.L. Multiple small RNAs act additively to integrate sensory information and control quorum sensing in Vibrio harveyi. Genes Dev. 2007, 21, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ling, Y.; Jiang, H.; Qiu, Y.; Qiu, J.; Chen, H.; Yang, R.; Zhou, D. AphA is required for biofilm formation, motility, and virulence in pandemic Vibrio parahaemolyticus. Int. J. Food Microbiol. 2013, 160, 245–251. [Google Scholar] [CrossRef]
- Haycocks, J.R.J.; Warren, G.Z.L.; Walker, L.M.; Chlebek, J.L.; Dalia, T.N.; Dalia, A.B.; Grainger, D.C. The quorum sensing transcription factor AphA directly regulates natural competence in Vibrio cholerae. PLoS Genet. 2019, 15, e1008362. [Google Scholar] [CrossRef] [Green Version]
- Kovacikova, G.; Lin, W.; Skorupski, K. The LysR-type virulence activator AphB regulates the expression of genes in Vibrio cholerae in response to low pH and anaerobiosis. J. Bacteriol. 2010, 192, 4181–4191. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Taboada, B.; Ciria, R.; Martinez-Guerrer, C.E.; Merino, E. ProOpDB: Prokaryotic Operon DataBase. Nucleic Acids Res. 2021, 40, D627–D631. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′–3′) | Usage |
|---|---|---|
| opaR-1 | GAGACCGTTGAAGCATCG | Mutant construction |
| opaR-2 | CAGGTACCGAGTCCATATCCATTT | Mutant construction |
| opaR-3 | CAGGTACCCGAACACTAAAGCTCA | Mutant construction |
| opaR-4 | CAGAGCTCGGGTACGGTTTACCAC | Mutant construction |
| opaR-5 | GTTCTAGAGTGGGTTGAGGTAGGT | Mutant selection |
| opaR-6 | GGTCTAGAGTTGGTACTAACGGTG | Mutant selection |
| luxO-1 | GAGAGCTCCGTATTCGTGCCGCCAAAG | Mutant construction |
| luxO-2 | CTGGTACCGCTGTATCCTCAACCATC | Mutant construction |
| luxO-3 | GAGGTACCAGAAGAGCGGCAGAAGGTG | Mutant construction |
| luxO-4 | CTGGTACCCGACCGCTGGATGCAATC | Mutant construction |
| luxO-5 | GCTCTAGACGGCTGAGAAGCGTGATG | Mutant selection |
| luxO-6 | GGTCTAGAGAGTCCAAGAGCGATACG | Mutant selection |
| pirAQF | TTAGCCACTTTCCAGCCGC | qPCR |
| pirAQR | CCGGAAGTCGGTCGTAGTGT | qPCR |
| pirBQF | TCGTTATCAGCCCACGCAG | qPCR |
| pirBQR | TTTCACCGATTCTGATGTGCA | qPCR |
| aphAQF | GAAACTTATGGCTTGTGCTG | qPCR |
| aphAQR | GCGGCTTCAATTTCTTTGTA | qPCR |
| aphBQF | TGGGATGTTATTTTCCGTGT | qPCR |
| aphBQR | CTGCTAGATAGTCTTGGCTG | qPCR |
| gyrB-1 | GAAGGTGGTATTCAAGCGTTCG | qPCR |
| gyrB-2 | GAGATGCCGTCTTCACGTTCT | qPCR |
| AphA-F-NdeI | AATGCCCCATATGAGCCTGCCGCACGTG | Protein expression |
| AphA-R-XhoI | CCGCTCGAGTTAGCCAATAACTTCCAGCTCG | Protein expression |
| AphB-F-NdeI | AAGGCCCCATATGAAGCTGGACGATCTGAACC | Protein expression |
| AphB-R-XhoI | GCCGCTCGAGTTAGTGGATGTTATACGCAATAACAAAG | Protein expression |
| pirAB promoter-F1-NdeI | AGGCTTCCATATGAGTGGAAATGGTGAACTTGCGGAAG | EMSA |
| pirAB promoter-R1-XhoI | AAGCTCGAGGTCTACTTCTGTGACGCCTCCG | EMSA |
| pirAB promoter-F2-NdeI | AGGCTTCCATATGATTGATCATAAAAATGCATTCTTTTTTACAAAG | EMSA |
| pirAB promoter-R3-XhoI | AAGCTCGAGTATTAAATTGCACAAAATTATGCAACACG | EMSA |
| pirAB promoter-F7 | TTTGTGCAATTTAATAGGAGAACATCATGAG | EMSA |
| pirAB promoter-R-XhoI | AAGCTCGAGGTCTACTTCTGTGACGCCTCCG | EMSA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-J.; Huang, J.-Y.; Le, P.-T.; Lee, C.-T.; Chang, C.-C.; Yang, Y.-Y.; Su, E.C.-Y.; Lo, C.-F.; Wang, H.-C. Expression of the AHPND Toxins PirAvp and PirBvp Is Regulated by Components of the Vibrio parahaemolyticus Quorum Sensing (QS) System. Int. J. Mol. Sci. 2022, 23, 2889. https://doi.org/10.3390/ijms23052889
Lin S-J, Huang J-Y, Le P-T, Lee C-T, Chang C-C, Yang Y-Y, Su EC-Y, Lo C-F, Wang H-C. Expression of the AHPND Toxins PirAvp and PirBvp Is Regulated by Components of the Vibrio parahaemolyticus Quorum Sensing (QS) System. International Journal of Molecular Sciences. 2022; 23(5):2889. https://doi.org/10.3390/ijms23052889
Chicago/Turabian StyleLin, Shin-Jen, Jiun-Yan Huang, Phuoc-Thien Le, Chung-Te Lee, Che-Chang Chang, Yi-Yuan Yang, Emily Chia-Yu Su, Chu-Fang Lo, and Hao-Ching Wang. 2022. "Expression of the AHPND Toxins PirAvp and PirBvp Is Regulated by Components of the Vibrio parahaemolyticus Quorum Sensing (QS) System" International Journal of Molecular Sciences 23, no. 5: 2889. https://doi.org/10.3390/ijms23052889
APA StyleLin, S.-J., Huang, J.-Y., Le, P.-T., Lee, C.-T., Chang, C.-C., Yang, Y.-Y., Su, E. C.-Y., Lo, C.-F., & Wang, H.-C. (2022). Expression of the AHPND Toxins PirAvp and PirBvp Is Regulated by Components of the Vibrio parahaemolyticus Quorum Sensing (QS) System. International Journal of Molecular Sciences, 23(5), 2889. https://doi.org/10.3390/ijms23052889