
Probiotics Enhance Bone Growth and Rescue BMP Inhibition: New Transgenic Zebrafish Lines to Study Bone Health




Abstract
:1. Introduction
2. Results
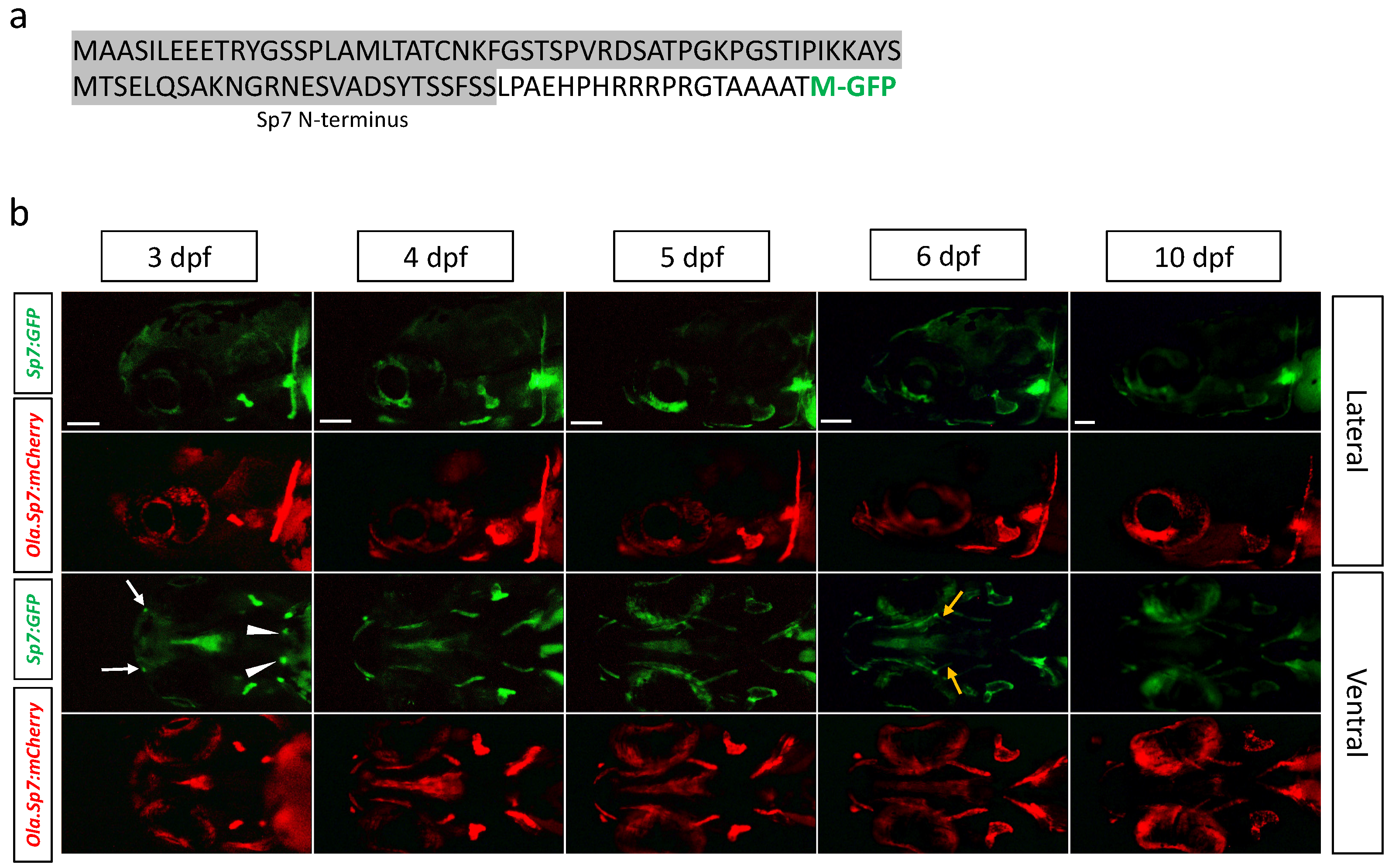
2.1. Generation and Characterization of New Transgenic Lines
2.1.1. Tg(col10a1a:col10a1a-GFP) Line
2.1.2. Tg(sp7:sp7-GFP) Line
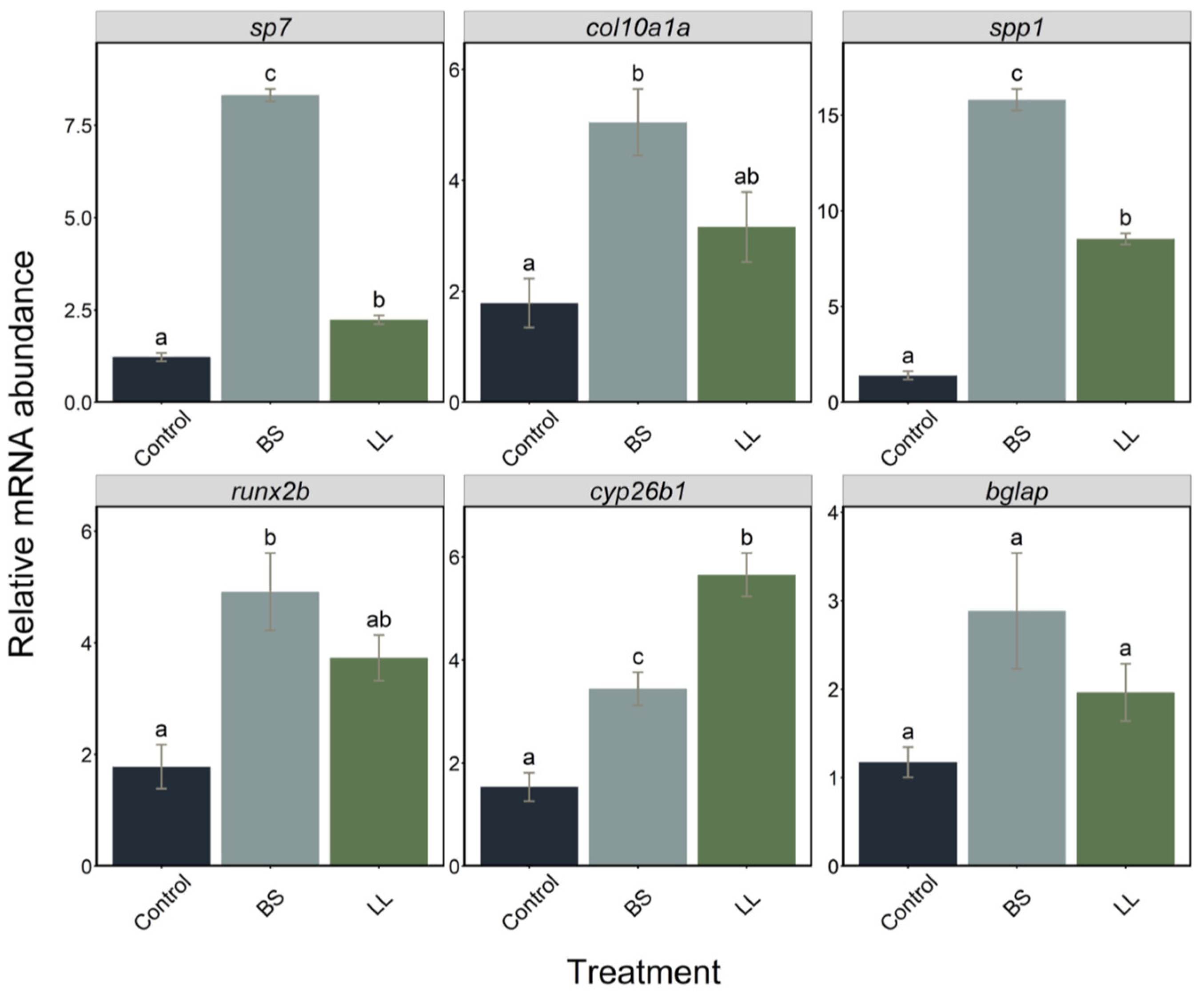
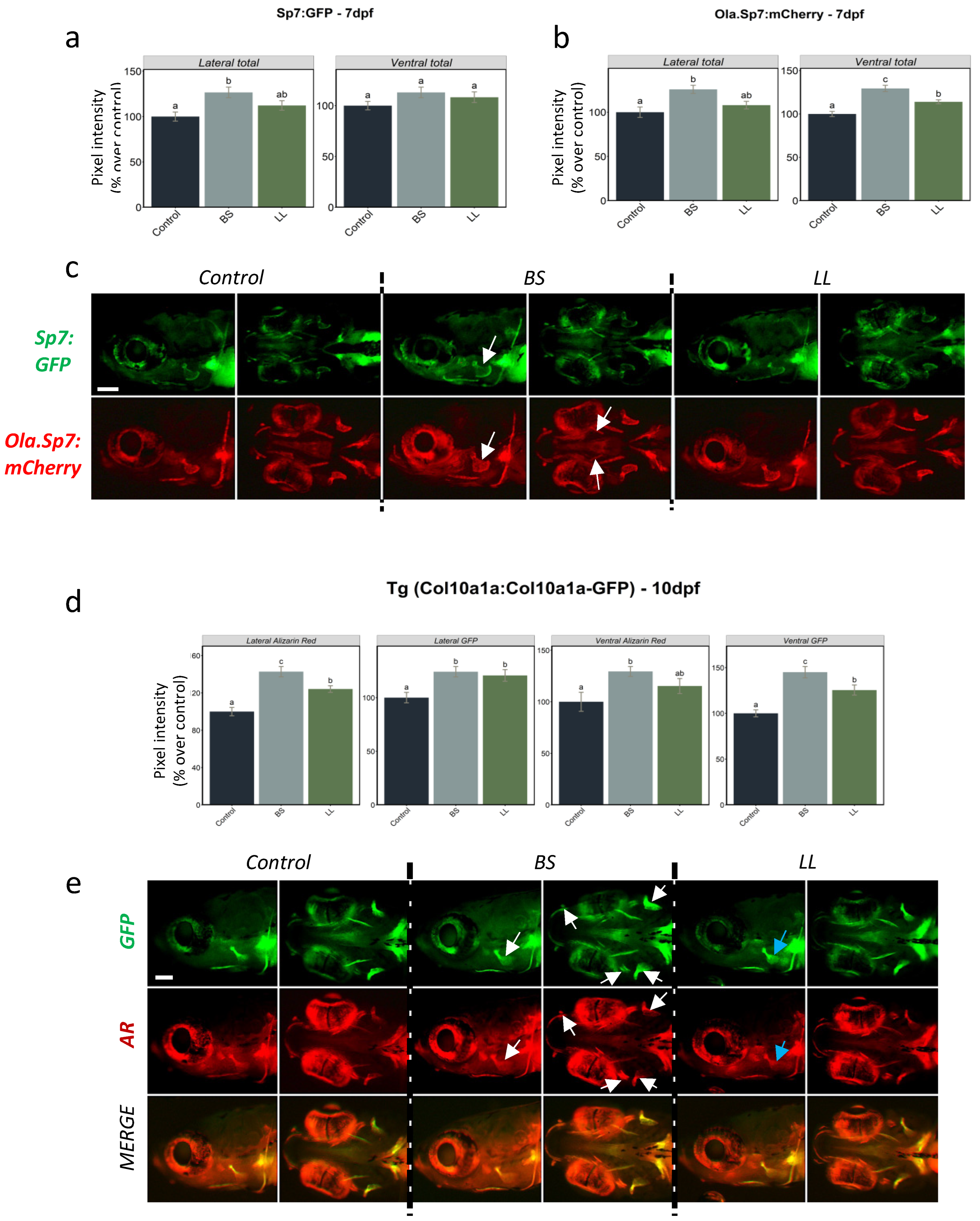
2.2. Effect of Probiotics
2.2.1. Effect of Probiotics on Bone Formation
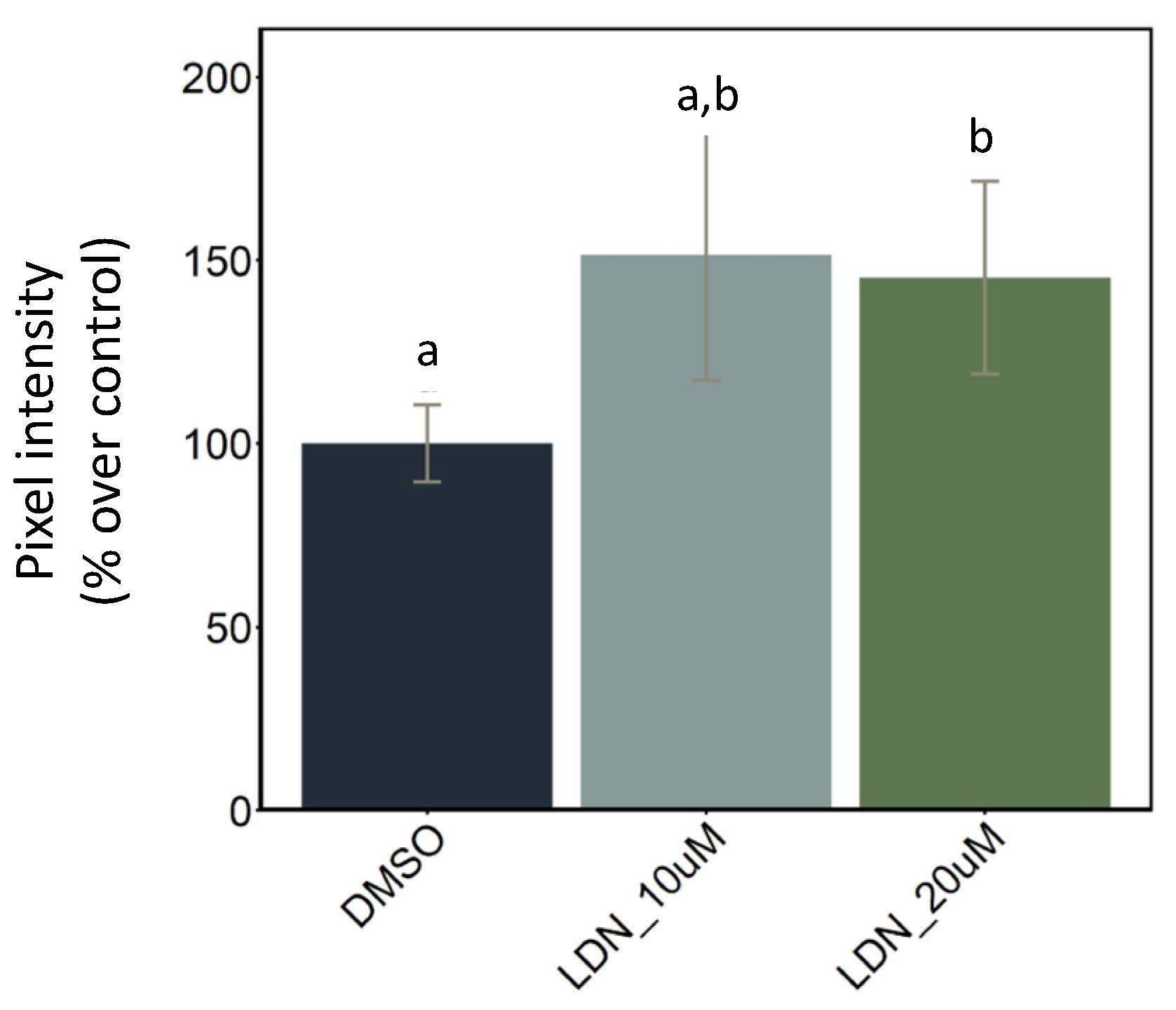
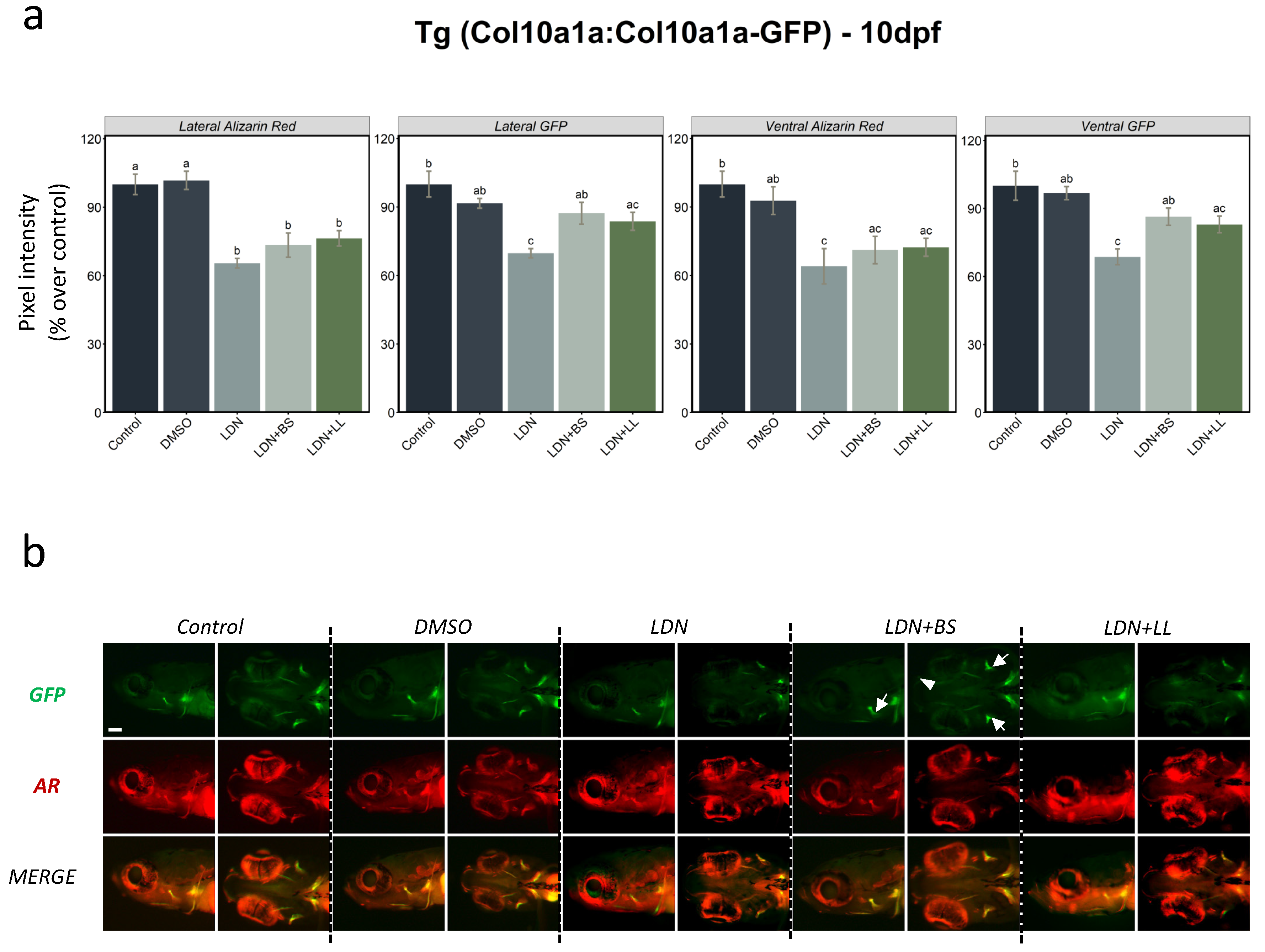
2.2.2. BMP Inhibitor Exposure Followed by Probiotic Treatment
3. Discussion
4. Materials and Methods
4.1. Generation of Transgenic Lines Using the CRISPR/Cas9 Method
- Mbait: gRNA1: GGCTGCTGCGGTTCCAGAGG
- col10a1a: gRNA2: GGAGTAAGGCTGGTACTGCG
- sp7: gRNA3: GGCTCATTCAGCTCAAGCGG
- GFP-rev: GGTCTTGTAGTTGCCGTCGT
- col10a1a-for: TTGTCAAGAAGGTGATGAAGG
- sp7-for: AAAAGGCCTACAGCATGACTTC
4.2. Morpholino Injection
- MOentpd5: AATTTAGTCTTACCTTTTCAGGC
- MOcon: random sequence
- MOp53: GACCTCCTCTCCACTAAACTACGAT
4.3. RNA Extraction and Quantification
4.4. Real Time PCR (RT-PCR)
4.5. Zebrafish Transgenic Lines Maintenance
4.6. Exposure to LDN212854 and Probiotics
4.7. Alizarin Red (AR) Staining
4.8. Image Acquisition and Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCabe, L.; Britton, R.A.; Parameswaran, N. Prebiotic and Probiotic Regulation of Bone Health: Role of the Intestine and Its Microbiome. Curr. Osteoporos. Rep. 2015, 13, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Ohlsson, C.; Sjögren, K. Osteomicrobiology: A New Cross-Disciplinary Research Field. Calcif. Tissue Int. 2018, 102, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R.; Biver, E. Are Probiotics the New Calcium and Vitamin D for Bone Health? Curr. Osteoporos. Rep. 2020, 18, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Cosme-Silva, L.; Dal-Fabbro, R.; Cintra, L.T.A.; Ervolino, E.; Plazza, F.; Mogami Bomfim, S.; Duarte, P.C.T.; Junior, V.E.D.S.; Gomes-Filho, J.E. Reduced Bone Resorption and Inflammation in Apical Periodontitis Evoked by Dietary Supplementation with Probiotics in Rats. Int. Endod. J. 2020, 53, 1084–1092. [Google Scholar] [CrossRef] [PubMed]
- Gholami, A.; Dabbaghmanesh, M.H.; Ghasemi, Y.; Talezadeh, P.; Koohpeyma, F.; Montazeri-Najafabady, N. Probiotics Ameliorate Pioglitazone-Associated Bone Loss in Diabetic Rats. Diabetol. Metab. Syndr. 2020, 12, 78. [Google Scholar] [CrossRef]
- Huidrom, S.; Beg, M.A.; Masood, T. Post-Menopausal Osteoporosis and Probiotics. Curr. Drug Targets 2021, 22, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Tu, Y.; Jia, X.; Du, Q.; Zheng, X.; Yuan, Q.; Zheng, L.; Zhou, X.; Xu, X. Probiotics Ameliorate Alveolar Bone Loss by Regulating Gut Microbiota. Cell Prolif. 2021, 54, e13075. [Google Scholar] [CrossRef]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R.; Probiotic, L. Reuteri Treatment Prevents Bone Loss in a Menopausal Ovariectomized Mouse Model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.-S.; Pan, T.-M. Antiosteoporotic Effects of Lactobacillus-Fermented Soy Skim Milk on Bone Mineral Density and the Microstructure of Femoral Bone in Ovariectomized Mice. J. Agric. Food Chem. 2011, 59, 7734–7742. [Google Scholar] [CrossRef]
- Ohlsson, C.; Engdahl, C.; Fåk, F.; Andersson, A.; Windahl, S.H.; Farman, H.H.; Movérare-Skrtic, S.; Islander, U.; Sjögren, K. Probiotics Protect Mice from Ovariectomy-Induced Cortical Bone Loss. PLoS ONE 2014, 9, e92368. [Google Scholar] [CrossRef]
- Parvaneh, K.; Ebrahimi, M.; Sabran, M.R.; Karimi, G.; Hwei, A.N.M.; Abdul-Majeed, S.; Ahmad, Z.; Ibrahim, Z.; Jamaluddin, R. Probiotics (Bifidobacterium Longum) Increase Bone Mass Density and Upregulate Sparc and Bmp-2 Genes in Rats with Bone Loss Resulting from Ovariectomy. BioMed. Res. Int. 2015, 2015, e897639. [Google Scholar] [CrossRef] [Green Version]
- Foureaux, R.d.C.; Messora, M.R.; de Oliveira, L.F.F.; Napimoga, M.H.; Pereira, A.N.J.; Ferreira, M.S.; Pereira, L.J. Effects of Probiotic Therapy on Metabolic and Inflammatory Parameters of Rats with Ligature-Induced Periodontitis Associated with Restraint Stress. J. Periodontol. 2014, 85, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.G.; Sundh, D.; Bäckhed, F.; Lorentzon, M. Lactobacillus Reuteri Reduces Bone Loss in Older Women with Low Bone Mineral Density: A Randomized, Placebo-Controlled, Double-Blind, Clinical Trial. J. Intern. Med. 2018, 284, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, T.; Hatanaka, M.; Hoshino, T.; Takara, T.; Tanaka, K.; Shimizu, A.; Morita, H.; Nakamura, T. Effect of Bacillus Subtilis C-3102 on Bone Mineral Density in Healthy Postmenopausal Japanese Women: A Randomized, Placebo-Controlled, Double-Blind Clinical Trial. Biosci. Microbiota Food Health 2018, 37, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maradonna, F.; Gioacchini, G.; Falcinelli, S.; Bertotto, D.; Radaelli, G.; Olivotto, I.; Carnevali, O. Probiotic Supplementation Promotes Calcification in Danio Rerio Larvae: A Molecular Study. PLoS ONE 2013, 8, e83155. [Google Scholar] [CrossRef] [Green Version]
- Terashima, A.; Takayanagi, H. Overview of Osteoimmunology. Calcif. Tissue Int. 2018, 102, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yoon, S.; Zhang, Y.-G.; Lu, R.; Xia, Y.; Wan, J.; Petrof, E.O.; Claud, E.C.; Chen, D.; Sun, J. Vitamin D Receptor Pathway Is Required for Probiotic Protection in Colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G341–G349. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-Y.; Chassaing, B.; Tyagi, A.M.; Vaccaro, C.; Luo, T.; Adams, J.; Darby, T.M.; Weitzmann, M.N.; Mulle, J.G.; Gewirtz, A.T.; et al. Sex Steroid Deficiency–Associated Bone Loss Is Microbiota Dependent and Prevented by Probiotics. J. Clin. Investig. 2016, 126, 2049–2063. [Google Scholar] [CrossRef] [Green Version]
- Chaplin, A.; Parra, P.; Laraichi, S.; Serra, F.; Palou, A. Calcium Supplementation Modulates Gut Microbiota in a Prebiotic Manner in Dietary Obese Mice. Mol. Nutr. Food Res. 2016, 60, 468–480. [Google Scholar] [CrossRef]
- Atkins, G.J.; Welldon, K.J.; Wijenayaka, A.R.; Bonewald, L.F.; Findlay, D.M. Vitamin K Promotes Mineralization, Osteoblast-to-Osteocyte Transition, and an Anticatabolic Phenotype by γ-Carboxylation-Dependent and -Independent Mechanisms. Am. J. Physiol. Cell Physiol. 2009, 297, C1358–C1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, S.L. Roles for Vitamin K Beyond Coagulation. Annu. Rev. Nutr. 2009, 29, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, M.; Strong, J.M.; Alabi, D.A.; Hernandez, C.J. The Gut Microbiome and Bone Strength. Curr. Osteoporos. Rep. 2020, 18, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Lleras-Forero, L.; Winkler, C.; Schulte-Merker, S. Zebrafish and Medaka as Models for Biomedical Research of Bone Diseases. Dev. Biol. 2020, 457, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Bergen, D.J.M.; Kague, E.; Hammond, C.L. Zebrafish as an Emerging Model for Osteoporosis: A Primary Testing Platform for Screening New Osteo-Active Compounds. Front. Endocrinol. 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhang, R.; Chen, H.; Du, X.; Chen, S.; Huang, J.; Liu, M.; Xu, M.; Luo, F.; Jin, M.; et al. Fgfr3 Mutation Disrupts Chondrogenesis and Bone Ossification in Zebrafish Model Mimicking CATSHL Syndrome Partially via Enhanced Wnt/β-Catenin Signaling. Theranostics 2020, 10, 7111–7130. [Google Scholar] [CrossRef]
- Jacobs, C.T.; Huang, P. Notch Signalling Maintains Hedgehog Responsiveness via a Gli-Dependent Mechanism during Spinal Cord Patterning in Zebrafish. eLife 2019, 8, e49252. [Google Scholar] [CrossRef]
- Alhazmi, N.; Carroll, S.H.; Kawasaki, K.; Woronowicz, K.C.; Hallett, S.A.; Macias Trevino, C.; Li, E.B.; Baron, R.; Gori, F.; Yelick, P.C.; et al. Synergistic Roles of Wnt Modulators R-Spondin2 and R-Spondin3 in Craniofacial Morphogenesis and Dental Development. Sci. Rep. 2021, 11, 5871. [Google Scholar] [CrossRef]
- Lovely, C.B.; Swartz, M.E.; McCarthy, N.; Norrie, J.L.; Eberhart, J.K. Bmp Signaling Mediates Endoderm Pouch Morphogenesis by Regulating Fgf Signaling in Zebrafish. Development 2016, 143, 2000–2011. [Google Scholar] [CrossRef] [Green Version]
- Schiavone, M.; Rampazzo, E.; Casari, A.; Battilana, G.; Persano, L.; Moro, E.; Liu, S.; Leach, S.D.; Tiso, N.; Argenton, F. Zebrafish Reporter Lines Reveal in Vivo Signaling Pathway Activities Involved in Pancreatic Cancer. Dis. Model Mech. 2014, 7, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Westphal, M.; Panza, P.; Kastenhuber, E.; Wehrle, J.; Driever, W. Wnt/β-Catenin Signaling Promotes Neurogenesis in the Diencephalospinal Dopaminergic System of Embryonic Zebrafish. Sci. Rep. 2022, 12, 1030. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Shibata, E.; Hans, S.; Brand, M.; Kawakami, A. Osteoblast Production by Reserved Progenitor Cells in Zebrafish Bone Regeneration and Maintenance. Dev. Cell 2017, 43, 643–650.e3. [Google Scholar] [CrossRef] [Green Version]
- Cooney, O.D.; Nagareddy, P.R.; Murphy, A.J.; Lee, M.K.S. Healthy Gut, Healthy Bones: Targeting the Gut Microbiome to Promote Bone Health. Front. Endocrinol. 2021, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Renn, J.; Pruvot, B.; Muller, M. Detection of Nitric Oxide by Diaminofluorescein Visualizes the Skeleton in Living Zebrafish. J. Appl. Ichthyol. 2014, 30, 701–706. [Google Scholar] [CrossRef]
- Debiais-Thibaud, M.; Simion, P.; Ventéo, S.; Muñoz, D.; Marcellini, S.; Mazan, S.; Haitina, T. Skeletal Mineralization in Association with Type X Collagen Expression Is an Ancestral Feature for Jawed Vertebrates. Mol. Biol. Evol. 2019, 36, 2265–2276. [Google Scholar] [CrossRef] [Green Version]
- Huitema, L.F.A.; Apschner, A.; Logister, I.; Spoorendonk, K.M.; Bussmann, J.; Hammond, C.L.; Schulte-Merker, S. Entpd5 Is Essential for Skeletal Mineralization and Regulates Phosphate Homeostasis in Zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 21372–21377. [Google Scholar] [CrossRef] [Green Version]
- Zinck, N.W.; Jeradi, S.; Franz-Odendaal, T.A. Elucidating the Early Signaling Cues Involved in Zebrafish Chondrogenesis and Cartilage Morphology. J. Exp. Zool. B Mol. Dev. Evol. 2021, 336, 18–31. [Google Scholar] [CrossRef]
- Windhausen, T.; Squifflet, S.; Renn, J.; Muller, M. BMP Signaling Regulates Bone Morphogenesis in Zebrafish through Promoting Osteoblast Function as Assessed by Their Nitric Oxide Production. Molecules 2015, 20, 7586–7601. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Felber, K.; Elks, P.; Croucher, P.; Roehl, H.H. Tracking Gene Expression during Zebrafish Osteoblast Differentiation. Dev. Dyn. 2009, 238, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Azetsu, Y.; Inohaya, K.; Takano, Y.; Kinoshita, M.; Tasaki, M.; Kudo, A. The Sp7 Gene Is Required for Maturation of Osteoblast-Lineage Cells in Medaka (Oryzias Latipes) Vertebral Column Development. Dev. Biol. 2017, 431, 252–262. [Google Scholar] [CrossRef]
- Hammond, C.L.; Moro, E. Using Transgenic Reporters to Visualize Bone and Cartilage Signaling during Development in Vivo. Front. Endocrinol. 2012, 3, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spoorendonk, K.M.; Peterson-Maduro, J.; Renn, J.; Trowe, T.; Kranenbarg, S.; Winkler, C.; Schulte-Merker, S. Retinoic Acid and Cyp26b1 Are Critical Regulators of Osteogenesis in the Axial Skeleton. Development 2008, 135, 3765–3774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.P.; Holdway, J.E.; Poss, K.D. Regeneration of Amputated Zebrafish Fin Rays from de Novo Osteoblasts. Dev. Cell 2012, 22, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renn, J.; Winkler, C. Osterix-MCherry Transgenic Medaka for in Vivo Imaging of Bone Formation. Dev. Dyn. 2009, 238, 241–248. [Google Scholar] [CrossRef]
- Wiweger, M.I.; Zhao, Z.; van Merkesteyn, R.J.P.; Roehl, H.H.; Hogendoorn, P.C.W. HSPG-Deficient Zebrafish Uncovers Dental Aspect of Multiple Osteochondromas. PLoS ONE 2012, 7, e29734. [Google Scholar] [CrossRef] [Green Version]
- Clément, A.; Wiweger, M.; von der Hardt, S.; Rusch, M.A.; Selleck, S.B.; Chien, C.-B.; Roehl, H.H. Regulation of Zebrafish Skeletogenesis by Ext2/Dackel and Papst1/Pinscher. PLoS Genet. 2008, 4, e1000136. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.-L.; Bhattacharya, P.; He, X.J.; Ponugoti, B.; Marquardt, B.; Layman, J.; Grunloh, M.; Postlethwait, J.H.; Rubin, D.A. Duplicated Zebrafish Co-Orthologs of Parathyroid Hormone-Related Peptide (PTHrP, Pthlh) Play Different Roles in Craniofacial Skeletogenesis. J. Endocrinol. 2012, 214, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Reijntjes, S.; Rodaway, A.; Maden, M. The Retinoic Acid Metabolising Gene, CYP26B1, Patterns the Cartilaginous Cranial Neural Crest in Zebrafish. Int. J. Dev. Biol. 2003, 51, 351–360. [Google Scholar] [CrossRef]
- Laue, K.; Jänicke, M.; Plaster, N.; Sonntag, C.; Hammerschmidt, M. Restriction of Retinoic Acid Activity by Cyp26b1 Is Required for Proper Timing and Patterning of Osteogenesis during Zebrafish Development. Development 2008, 135, 3775–3787. [Google Scholar] [CrossRef] [Green Version]
- Laue, K.; Pogoda, H.-M.; Daniel, P.B.; van Haeringen, A.; Alanay, Y.; von Ameln, S.; Rachwalski, M.; Morgan, T.; Gray, M.J.; Breuning, M.H.; et al. Craniosynostosis and Multiple Skeletal Anomalies in Humans and Zebrafish Result from a Defect in the Localized Degradation of Retinoic Acid. Am. J. Hum. Genet. 2011, 89, 595–606. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, Z.; Yoshida, C.A.; Furuichi, T.; Amizuka, N.; Ito, M.; Fukuyama, R.; Miyazaki, T.; Kitaura, H.; Nakamura, K.; Fujita, T.; et al. Runx2 Determines Bone Maturity and Turnover Rate in Postnatal Bone Development and Is Involved in Bone Loss in Estrogen Deficiency. Dev. Dyn. 2007, 236, 1876–1890. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of Bone Development and Extracellular Matrix Protein Genes by RUNX2. Cell Tissue Res. 2009, 339, 189. [Google Scholar] [CrossRef] [Green Version]
- Lowery, J.W.; Rosen, V. The BMP Pathway and Its Inhibitors in the Skeleton. Physiol. Rev. 2018, 98, 2431–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, Y.; Hisano, Y.; Kawahara, A.; Higashijima, S. Efficient Generation of Knock-in Transgenic Zebrafish Carrying Reporter/Driver Genes by CRISPR/Cas9-Mediated Genome Engineering. Sci. Rep. 2014, 4, 6545. [Google Scholar] [CrossRef] [Green Version]
- Dalcq, J.; Pasque, V.; Ghaye, A.; Larbuisson, A.; Motte, P.; Martial, J.A.; Muller, M. RUNX3, EGR1 and SOX9B Form a Regulatory Cascade Required to Modulate BMP-Signaling during Cranial Cartilage Development in Zebrafish. PLoS ONE 2012, 7, e50140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larbuisson, A.; Dalcq, J.; Martial, J.A.; Muller, M. Fgf Receptors Fgfr1a and Fgfr2 Control the Function of Pharyngeal Endoderm in Late Cranial Cartilage Development. Differentiation 2013, 86, 192–206. [Google Scholar] [CrossRef] [Green Version]
- Robu, M.E.; Larson, J.D.; Nasevicius, A.; Beiraghi, S.; Brenner, C.; Farber, S.A.; Ekker, S.C. P53 Activation by Knockdown Technologies. PLoS Genet. 2007, 3, e78. [Google Scholar] [CrossRef]
- Maradonna, F.; Nozzi, V.; Dalla Valle, L.; Traversi, I.; Gioacchini, G.; Benato, F.; Colletti, E.; Gallo, P.; Di Marco Pisciottano, I.; Mita, D.G.; et al. A Developmental Hepatotoxicity Study of Dietary Bisphenol A in Sparus Aurata Juveniles. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 166, 1–13. [Google Scholar] [CrossRef]
- Carnevali, O.; Notarstefano, V.; Olivotto, I.; Graziano, M.; Gallo, P.; Di Marco Pisciottano, I.; Vaccari, L.; Mandich, A.; Giorgini, E.; Maradonna, F. Dietary Administration of EDC Mixtures: A Focus on Fish Lipid Metabolism. Aquat. Toxicol. 2017, 185, 95–104. [Google Scholar] [CrossRef]
- Bensimon-Brito, A.; Cardeira, J.; Dionísio, G.; Huysseune, A.; Cancela, M.L.; Witten, P.E. Revisiting in Vivo Staining with Alizarin Red S—A Valuable Approach to Analyse Zebrafish Skeletal Mineralization during Development and Regeneration. BMC Dev. Biol. 2016, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.yumpu.com/s/Fxit5xIdza5bKZeV (accessed on 24 March 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Acronym | NCBI Gene Accession No | Forward | Reverse |
|---|---|---|---|
| col10a1a | NM_001083827.1 | CCCATCCACATCACATCAAA | GCGTGCATTTCTCAGAACAA |
| runx2b | NM_212862.2 | GTGGCCACTTACCACAGAGC | TCGGAGAGTCATCCAGCTT |
| spp1 | NM_001002308.1 | GAGCCTACACAGACCACGCCAACAG | GGTAGCCCAAACTGTCTCCCCG |
| cyp26b1 | NM_212666.1 | GCTGTCAACCAGAACATTCCC | GGTTCTGATTGGAGTCGAGGC |
| sp7 | NM_212863.2 | AACCCAAGCCCGTCCCGACA | CCGTACACCTTCCCGCAGCC |
| bglap | NM_001083857.3 | GCCTGATGACTGTGTGTCTGAGCG | AGTTCCAGCCCTCTTCTGTCTCAT |
| rpl13a | NM_212784.1 | TCTGGAGGACTGTAAGAGGTATGC | AGACGCACAATCTTGAGAGCAG |
| rplp0 | NM_131580.2 | CTGAACATCTCGCCCTTCTC | TAGCCGATCTGCAGACACAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sojan, J.M.; Raman, R.; Muller, M.; Carnevali, O.; Renn, J. Probiotics Enhance Bone Growth and Rescue BMP Inhibition: New Transgenic Zebrafish Lines to Study Bone Health. Int. J. Mol. Sci. 2022, 23, 4748. https://doi.org/10.3390/ijms23094748
Sojan JM, Raman R, Muller M, Carnevali O, Renn J. Probiotics Enhance Bone Growth and Rescue BMP Inhibition: New Transgenic Zebrafish Lines to Study Bone Health. International Journal of Molecular Sciences. 2022; 23(9):4748. https://doi.org/10.3390/ijms23094748
Chicago/Turabian StyleSojan, Jerry Maria, Ratish Raman, Marc Muller, Oliana Carnevali, and Jörg Renn. 2022. "Probiotics Enhance Bone Growth and Rescue BMP Inhibition: New Transgenic Zebrafish Lines to Study Bone Health" International Journal of Molecular Sciences 23, no. 9: 4748. https://doi.org/10.3390/ijms23094748
APA StyleSojan, J. M., Raman, R., Muller, M., Carnevali, O., & Renn, J. (2022). Probiotics Enhance Bone Growth and Rescue BMP Inhibition: New Transgenic Zebrafish Lines to Study Bone Health. International Journal of Molecular Sciences, 23(9), 4748. https://doi.org/10.3390/ijms23094748