
Molecular Research on Oral Diseases and Related Biomaterials: A Journey from Oral Cell Models to Advanced Regenerative Perspectives



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
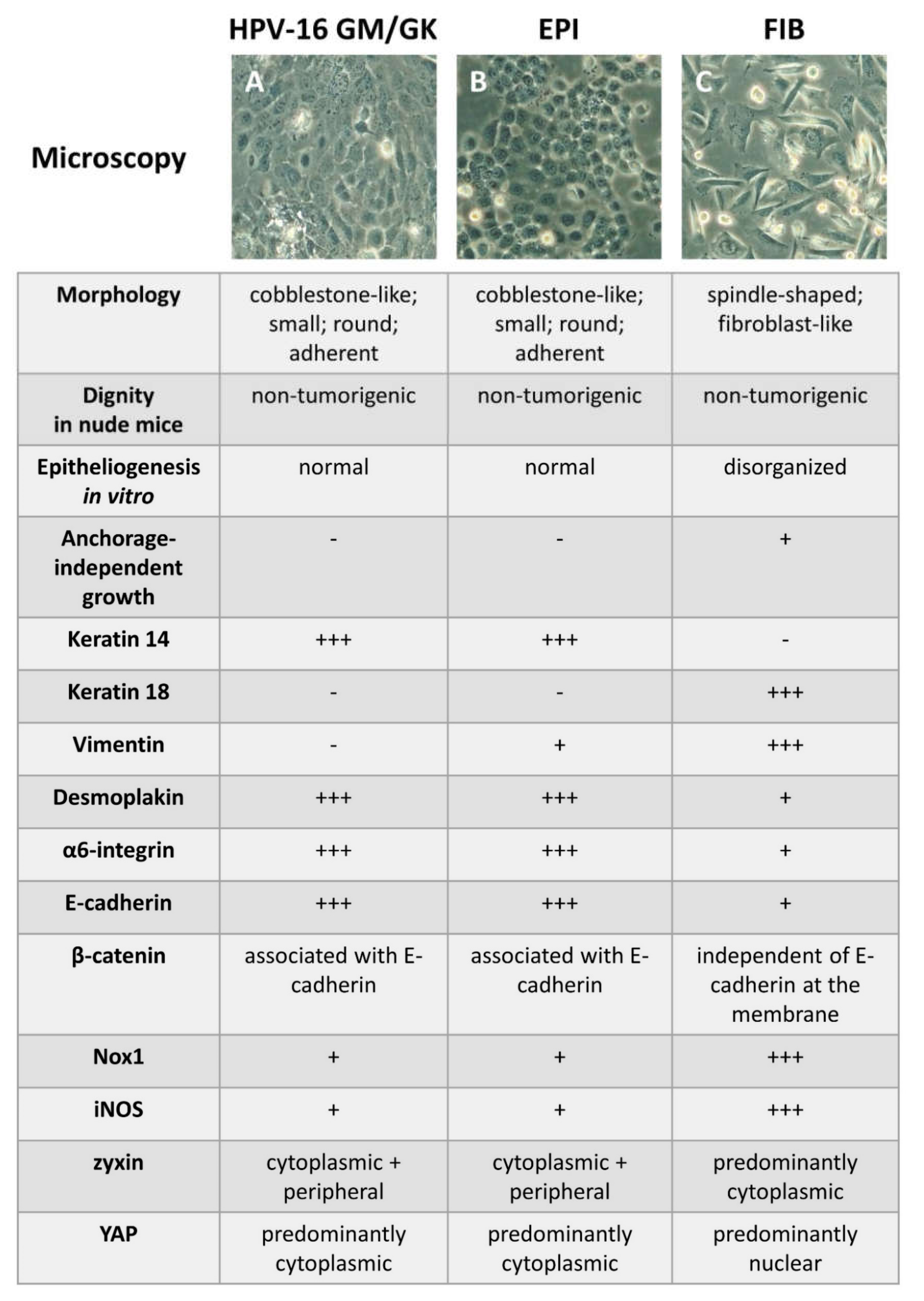
2.1. In Vitro Cell Models to Study Oral Epithelial Tissue Homeostasis and Carcinogenesis
2.2. Biomaterial-Based Approaches in the Context of Oral Diseases
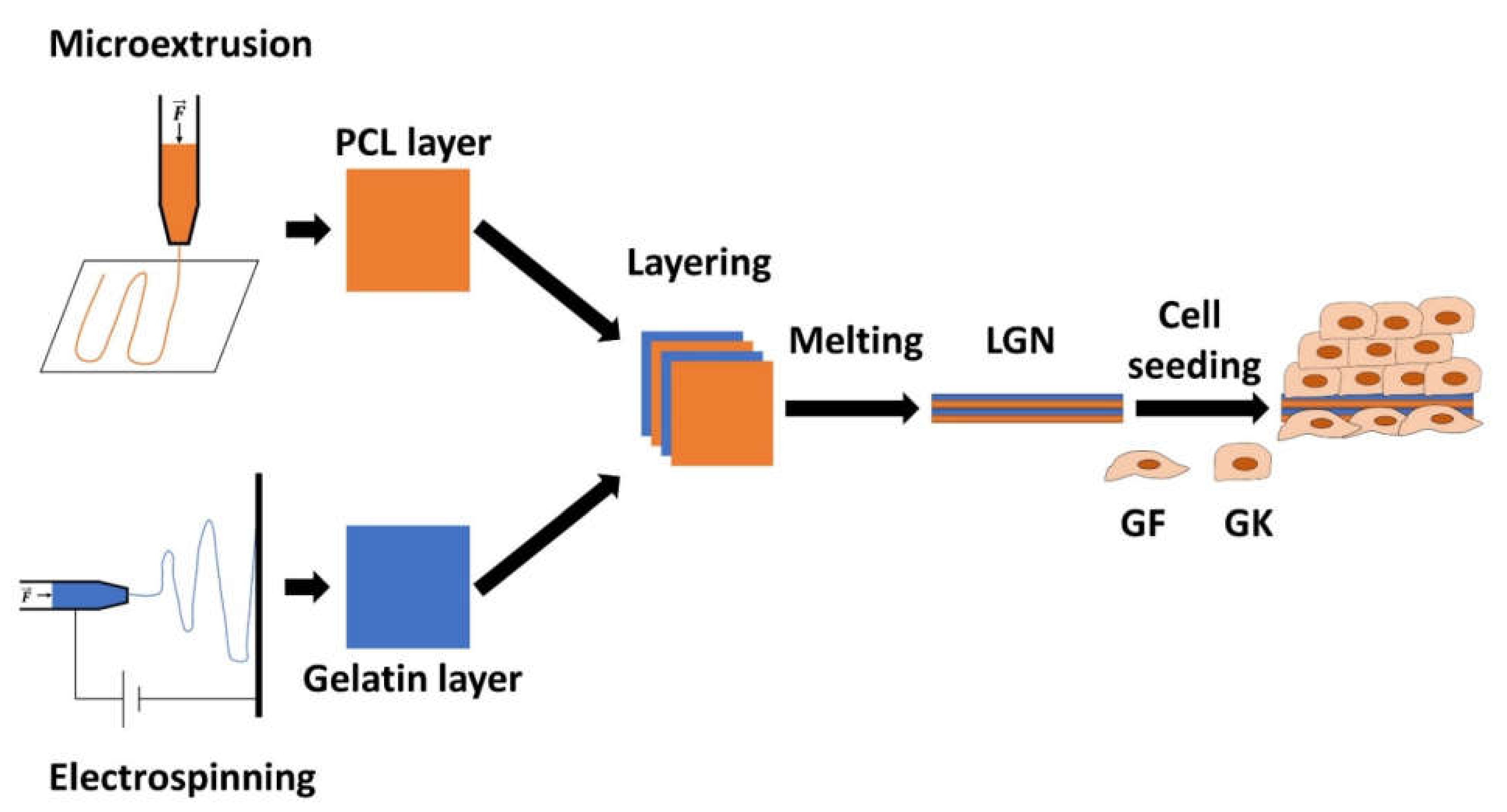
2.2.1. Nonwoven Biomaterial Design for Oral Epithelial and Soft Tissue Regeneration
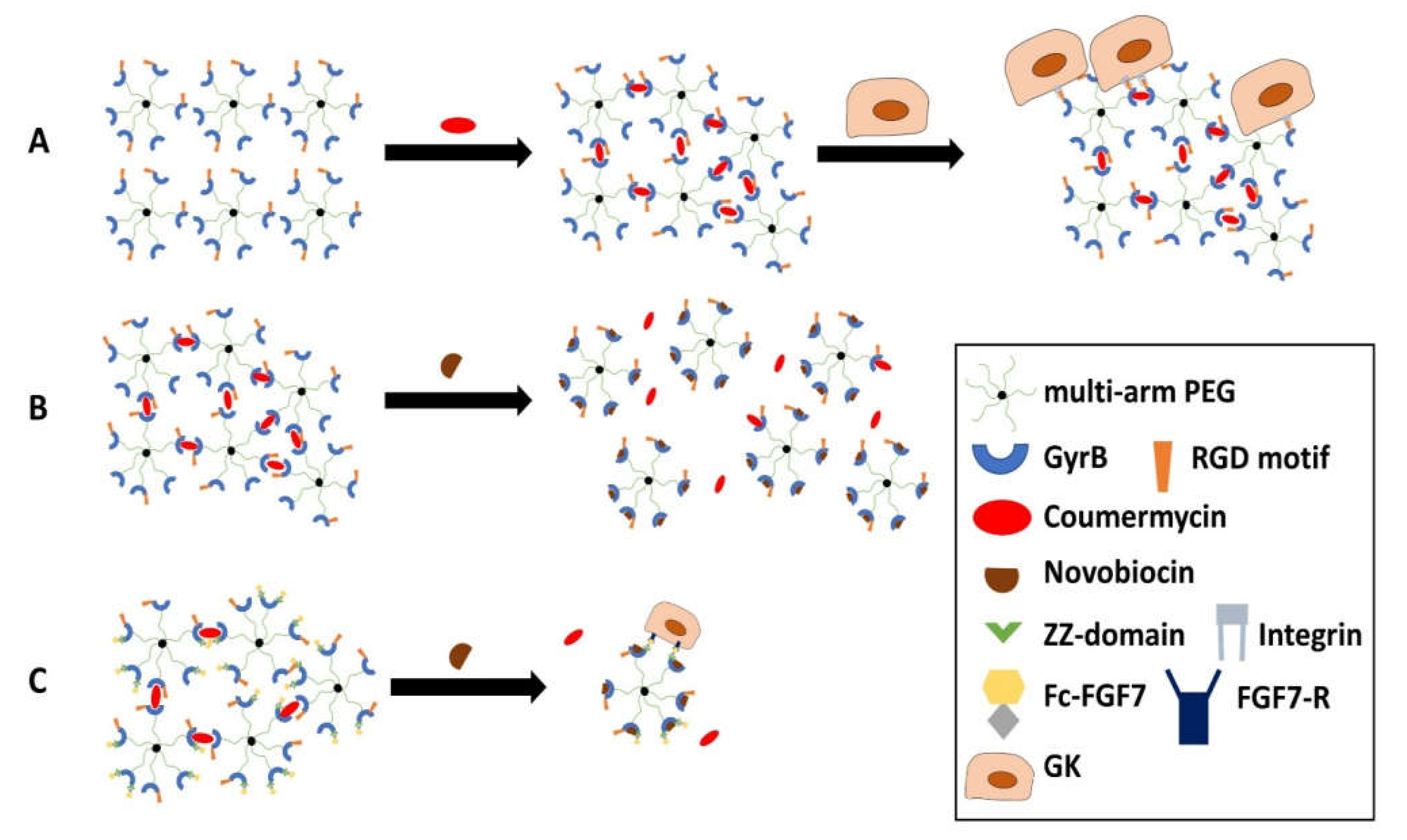
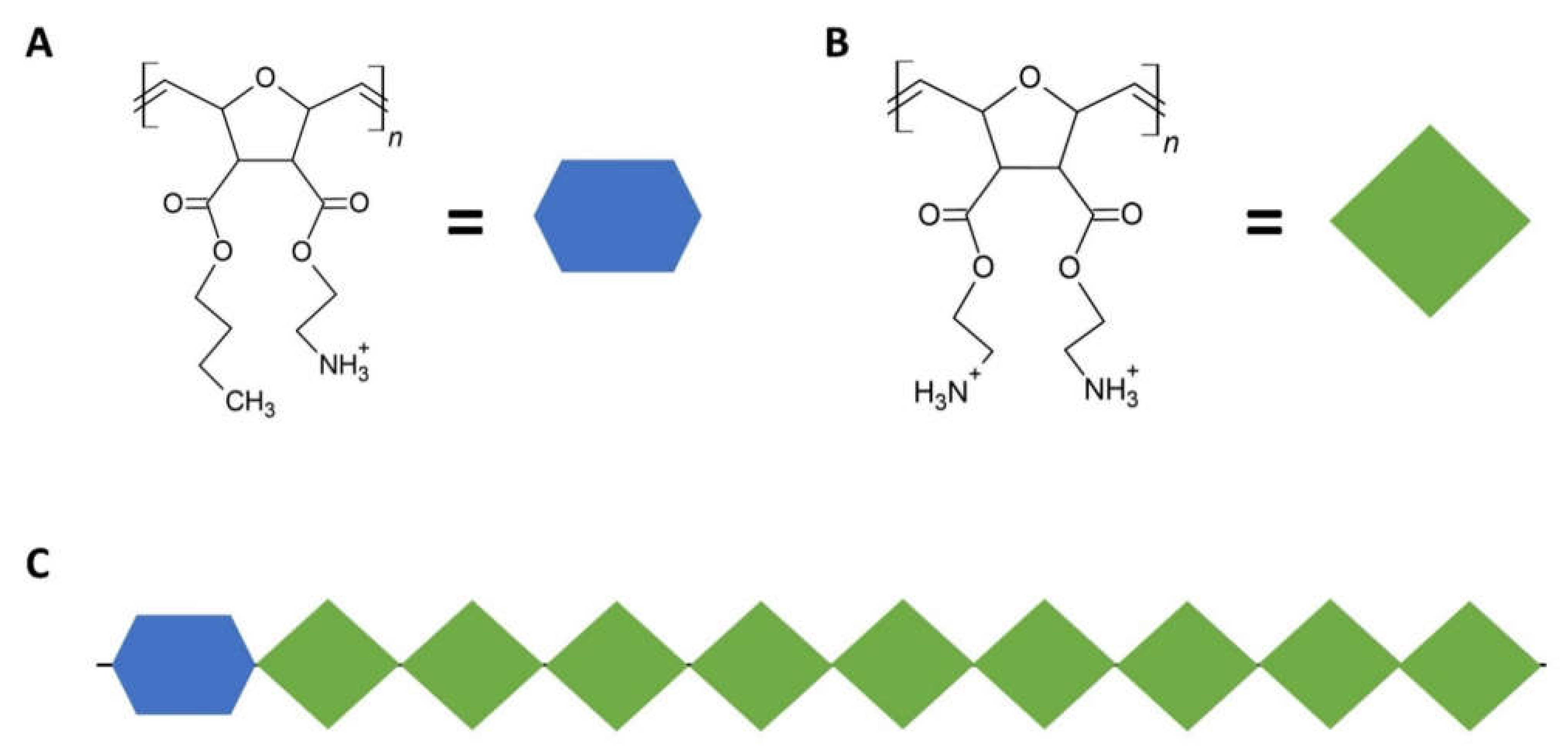
2.2.2. A Smart Hydrogel for Controlled Drug Release in Periodontal Regeneration
2.2.3. Synthetic Mimics of Antimicrobial Peptides (SMAMPS) as a Strategy to Combat Oral Pathogens
2.3. Cell-Based and Optogenetic Approaches: Perspectives in the Context of Oral Diseases
2.3.1. Cell-Based Approaches
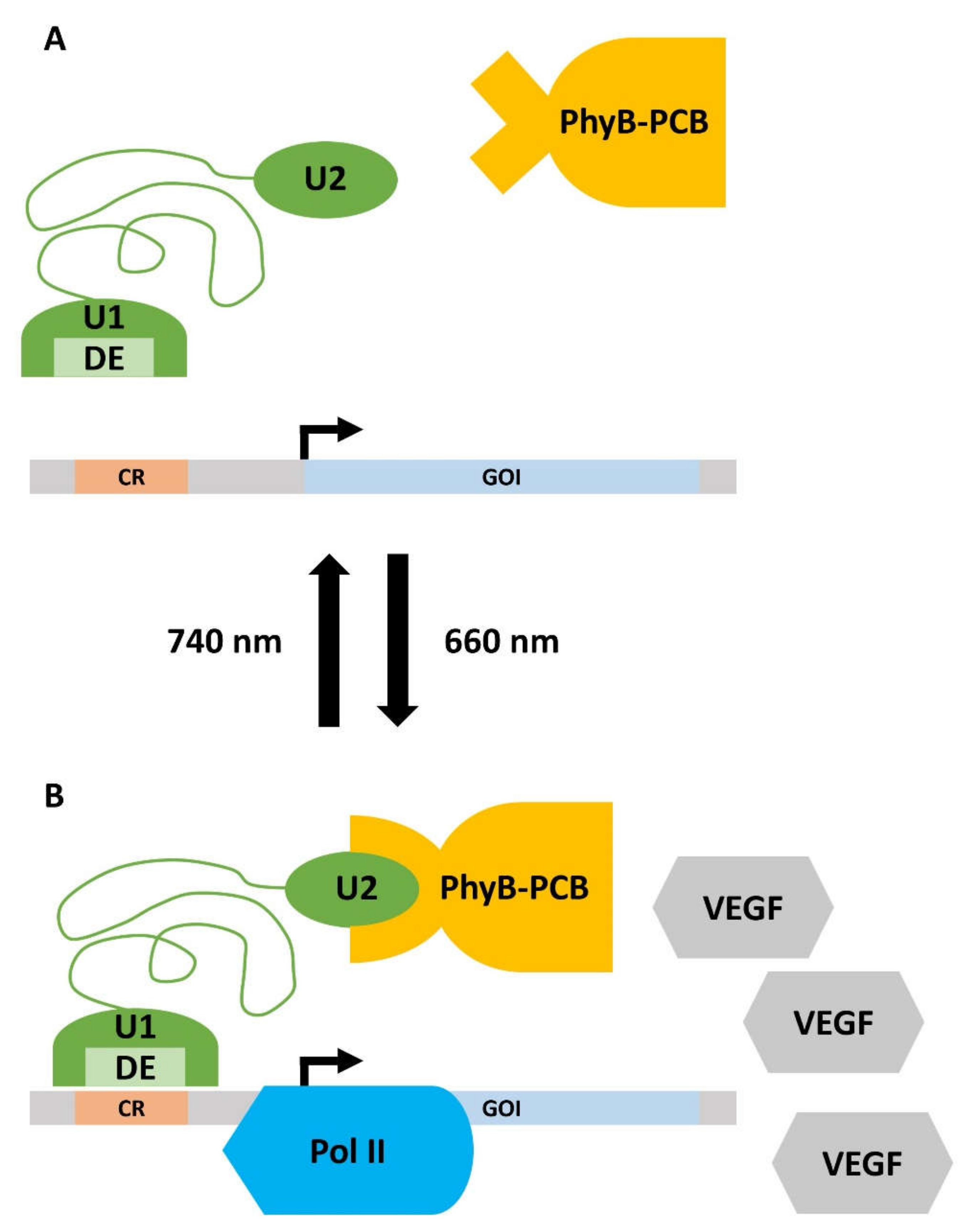
2.3.2. Optogenetic Strategies
3. Conclusions
4. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmad, P.; Rubbia Nawaz, M.Q.; Shaikh, G.M.; Mohamed, R.N.; Nagarajappa, A.K.; Asif, J.A.; Alam, M.K. Risk factors associated with the mortality rate of oral squamous cell carcinoma patients: A 10-year retrospective study. Medicine 2021, 100, e27127. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, S.Q.; Zhao, L.; Ren, Z.H.; Hu, C.Y. Global, regional, and national burden of periodontitis from 1990 to 2019: Results from the Global Burden of Disease study 2019. J. Periodontol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, S.; Caton, J.G.; Albandar, J.M.; Bissada, N.F.; Bouchard, P.; Cortellini, P.; Demirel, K.; de Sanctis, M.; Ercoli, C.; Fan, J. Periodontal manifestations of systemic diseases and developmental and acquired conditions: Consensus report of workgroup 3 of the 2017 World Workshop on the Classification of Periodontal and Peri-Implant Diseases and Conditions. J. Clin. Periodontol. 2018, 45, S219–S229. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Tilakaratne, W.M. Update from the 5th Edition of the World Health Organization Classification of Head and Neck Tumors: Tumours of the Oral Cavity and Mobile Tongue. Head Neck Pathol. 2022, 16, 54–62. [Google Scholar] [CrossRef]
- Saccucci, M.; Di Carlo, G.; Bossù, M.; Giovarruscio, F.; Salucci, A.; Polimeni, A. Autoimmune diseases and their manifestations on oral cavity: Diagnosis and clinical management. J. Immunol. Res. 2018, 2018. [Google Scholar] [CrossRef]
- Schmalz, G.; Garbade, J.; Kollmar, O.; Ziebolz, D. Does oral health-related quality of life of patients after solid organ transplantation indicate a response shift? Results of a systematic review. BMC Oral Health 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Warnakulasuriya, S.; Kujan, O.; Aguirre-Urizar, J.M.; Bagan, J.V.; González-Moles, M.Á.; Kerr, A.R.; Lodi, G.; Mello, F.W.; Monteiro, L.; Ogden, G.R. Oral potentially malignant disorders: A consensus report from an international seminar on nomenclature and classification, convened by the WHO Collaborating Centre for Oral Cancer. Oral Dis. 2021, 27, 1862–1880. [Google Scholar] [CrossRef]
- Wenig, B.M.; Childers, E.L.; Richardson, M.S.; Seethala, R.R.; Thompson, L.D. Non-Neoplastic Diseases of the Head and Neck; American Registry of Pathology in collaboration with the Armed Forces: Washington, DC, USA, 2017. [Google Scholar]
- Nazir, M.A. Prevalence of periodontal disease, its association with systemic diseases and prevention. Int. J. Health Sci. 2017, 11, 72–80. [Google Scholar]
- Ghantous, Y.; Elnaaj, A. Global incidence and risk factors of oral cancer. Harefuah 2017, 156, 645–649. [Google Scholar]
- Romano, A.; Di Stasio, D.; Petruzzi, M.; Fiori, F.; Lajolo, C.; Santarelli, A.; Lucchese, A.; Serpico, R.; Contaldo, M. Noninvasive Imaging Methods to Improve the Diagnosis of Oral Carcinoma and Its Precursors: State of the Art and Proposal of a Three-Step Diagnostic Process. Cancers 2021, 13, 2864. [Google Scholar] [CrossRef]
- Bouaoud, J.; Bossi, P.; Elkabets, M.; Schmitz, S.; van Kempen, L.C.; Martinez, P.; Jagadeeshan, S.; Breuskin, I.; Puppels, G.J.; Hoffmann, C. Unmet Needs and Perspectives in Oral Cancer Prevention. Cancers 2022, 14, 1815. [Google Scholar] [CrossRef] [PubMed]
- Nocini, R.; Capocasale, G.; Marchioni, D.; Zotti, F. A snapshot of knowledge about oral cancer in Italy: A 505 person survey. Int. J. Environ. Res. Public Health 2020, 17, 4889. [Google Scholar] [CrossRef] [PubMed]
- Räisänen, I.T.; Umeizudike, K.A.; Pärnänen, P.; Heikkilä, P.; Tervahartiala, T.; Nwhator, S.O.; Grigoriadis, A.; Sakellari, D.; Sorsa, T. Periodontal disease and targeted prevention using aMMP-8 point-of-care oral fluid analytics in the COVID-19 era. Med. Hypotheses 2020, 144, 110276. [Google Scholar] [CrossRef] [PubMed]
- Schmalz, G.; Ziebolz, D. Changing the focus to the whole patient instead of one oral disease: The concept of individualized prevention. Adv. Prev. Med. 2020, 2020, 6752342. [Google Scholar] [CrossRef] [PubMed]
- Algadi, H.A.H.; Abou-Bakr, A.A.-E.; Jamali, O.M.; Fathy, L.M. Toluidine blue versus frozen section for assessment of mucosal tumor margins in oral squamous cell carcinoma. BMC Cancer 2020, 20, 1147. [Google Scholar] [CrossRef]
- Shavlokhova, V.; Sandhu, S.; Flechtenmacher, C.; Koveshazi, I.; Neumeier, F.; Padrón-Laso, V.; Jonke, Ž.; Saravi, B.; Vollmer, M.; Vollmer, A. Deep Learning on Oral Squamous Cell Carcinoma Ex Vivo Fluorescent Confocal Microscopy Data: A Feasibility Study. J. Clin. Med. 2021, 10, 5326. [Google Scholar] [CrossRef]
- Sun, L.-F.; Wang, C.-X.; Cao, Z.-Y.; Han, W.; Guo, S.-S.; Wang, Y.-Z.; Meng, Y.; Hou, C.-X.; Zhu, Q.-H.; Tang, Y.-T. Evaluation of autofluorescence visualization system in the delineation of oral squamous cell carcinoma surgical margins. Photodiagnosis Photodyn. Ther. 2021, 36, 102487. [Google Scholar] [CrossRef]
- Yang, Z.; Shang, J.; Liu, C.; Zhang, J.; Liang, Y. Identification of oral precancerous and cancerous tissue by swept source optical coherence tomography. Lasers Surg. Med. 2022, 54, 320–328. [Google Scholar] [CrossRef]
- Chaicharoenaudomrung, N.; Kunhorm, P.; Noisa, P. Three-dimensional cell culture systems as an in vitro platform for cancer and stem cell modeling. World J. Stem Cells 2019, 11, 1065–1083. [Google Scholar] [CrossRef]
- Al-Dabbagh, R.; Al-Hazmi, N.; Alhazzazi, T.Y.; Barrett, A.; Speight, P.M. Human papillomavirus and head and neck squamous cell carcinoma in a UK population: Is there an association. Indian J. Cancer 2021. [Google Scholar] [CrossRef]
- Duan, Y.; Huang, X.; Qiao, B.; Ma, R.; Li, J. Eugenol inhibits the biological activities of an oral squamous cell carcinoma cell line SCC9 via targeting MIF. Anti-Cancer Agents Med. Chem. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-C.; Huang, M.-Y.; Chen, Y.-K.; Lan, W.-C.; Shieh, T.-M.; Shih, Y.-H. Salivary Exosome Proteomics and Bioinformatics Analysis in 7, 12-Dimethylbenz [a] anthracene-Induced Oral Cancer with Radiation Therapy—A Syrian Golden Hamster Model. Diagnostics 2021, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Truchard, E.; Bertolus, C.; Martinez, P.; Thomas, E.; Saintigny, P.; Foy, J.-P. Identification of a Gene-Expression-Based Surrogate of Genomic Instability during Oral Carcinogenesis. Cancers 2022, 14, 834. [Google Scholar] [CrossRef] [PubMed]
- Fleetwood, A.J.; Lee, M.K.; Singleton, W.; Achuthan, A.; Lee, M.-C.; O’Brien-Simpson, N.M.; Cook, A.D.; Murphy, A.J.; Dashper, S.G.; Reynolds, E.C. Metabolic remodeling, inflammasome activation, and pyroptosis in macrophages stimulated by Porphyromonas gingivalis and its outer membrane vesicles. Front. Cell. Infect. Microbiol. 2017, 7, 351. [Google Scholar] [CrossRef]
- Ram-Mohan, N.; Meyer, M.M. Comparative metatranscriptomics of periodontitis supports a common polymicrobial shift in metabolic function and identifies novel putative disease-associated ncRNAs. Front. Microbiol. 2020, 11, 482. [Google Scholar] [CrossRef] [Green Version]
- Nakagaki, T.; Tamura, M.; Kobashi, K.; Koyama, R.; Fukushima, H.; Ohashi, T.; Idogawa, M.; Ogi, K.; Hiratsuka, H.; Tokino, T.; et al. Profiling cancer-related gene mutations in oral squamous cell carcinoma from Japanese patients by targeted amplicon sequencing. Oncotarget 2017, 8, 59113–59122. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Ji, X.; Zhang, X.; Tang, D.; Feng, Q. Persistent exposure to Fusobacterium nucleatum triggers chemokine/cytokine release and inhibits the proliferation and osteogenic differentiation capabilities of human gingiva-derived mesenchymal stem cells. Front. Cell. Infect. Microbiol. 2019, 9, 429. [Google Scholar] [CrossRef]
- Keong, J.Y.; Low, L.W.; Chong, J.M.; Ong, Y.Y.; Pulikkotil, S.J.; Singh, G.; Nagendrababu, V.; Banavar, S.R.; Khoo, S.P. Effect of lipopolysaccharide on cell proliferation and vascular endothelial growth factor secretion of periodontal ligament stem cells. Saudi Dent. J. 2020, 32, 148–154. [Google Scholar] [CrossRef]
- Listyarifah, D.; Al-Samadi, A.; Salem, A.; Syaify, A.; Salo, T.; Tervahartiala, T.; Grenier, D.; Nordström, D.C.; Sorsa, T.; Ainola, M. Infection and apoptosis associated with inflammation in periodontitis: An immunohistologic study. Oral Dis. 2017, 23, 1144–1154. [Google Scholar] [CrossRef] [Green Version]
- Ramenzoni, L.L.; Russo, G.; Moccia, M.D.; Attin, T.; Schmidlin, P.R. Periodontal bacterial supernatants modify differentiation, migration and inflammatory cytokine expression in human periodontal ligament stem cells. PLoS ONE 2019, 14, e0219181. [Google Scholar] [CrossRef]
- Mountcastle, S.E.; Cox, S.C.; Sammons, R.L.; Jabbari, S.; Shelton, R.M.; Kuehne, S.A. A review of co-culture models to study the oral microenvironment and disease. J. Oral Microbiol. 2020, 12, 1773122. [Google Scholar] [CrossRef] [PubMed]
- Müssig, E.; Steinberg, T.; Kohl, A.; Chamulitrat, W.; Komposch, G.; Tomakidi, P. Discrimination of epithelium-like and fibroblast-like phenotypes derived from ethanol-treated immortalised human gingival keratinocytes in epithelial equivalents. Cell Tissue Res. 2008, 332, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Yang, H.; Wang, F.; Zhao, S. Oct4 downregulation-induced inflammation increases the migration and invasion rate of oral squamous cell carcinoma. Acta Biochim. Biophys. Sin. 2021, 53, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Teh, M.-T.; Ma, H.; Liang, Y.-Y.; Solomon, M.C.; Chaurasia, A.; Patil, R.; Tekade, S.A.; Mishra, D.; Qadir, F.; Yeung, J.-Y.S. Molecular Signatures of Tumour and Its Microenvironment for Precise Quantitative Diagnosis of Oral Squamous Cell Carcinoma: An International Multi-Cohort Diagnostic Validation Study. Cancers 2022, 14, 1389. [Google Scholar] [CrossRef]
- Yen, W.-C.; Chang, I.Y.-F.; Chang, K.-P.; Ouyang, C.N.; Liu, C.-R.; Tsai, T.-L.; Zhang, Y.-C.; Wang, C.-I.; Wang, Y.-H.; Yu, A.L. Genomic and molecular signatures of successful patient-derived xenografts for oral cavity squamous cell carcinoma. Front. Oncol. 2022, 12, 792297. [Google Scholar] [CrossRef]
- Han, J.; Menicanin, D.; Gronthos, S.; Bartold, P. Stem cells, tissue engineering and periodontal regeneration. Aust. Dent. J. 2014, 59 (Suppl. S1), 117–130. [Google Scholar] [CrossRef]
- Lam, V.K.L.; Wong, J.Y.H.; Chew, S.Y.; Chan, B.P. Rac1-GTPase regulates compression-induced actin protrusions (CAPs) of mesenchymal stem cells in 3D collagen micro-tissues. Biomaterials 2021, 274, 120829. [Google Scholar] [CrossRef]
- Bjørge, I.M.; de Sousa, B.M.; Patrício, S.G.; Silva, A.S.; Nogueira, L.P.; Santos, L.F.; Vieira, S.I.; Haugen, H.J.; Correia, C.R.; Mano, J.F. Bioengineered Hierarchical Bonelike Compartmentalized Microconstructs Using Nanogrooved Microdiscs. ACS Appl. Mater. Interfaces 2022, 14, 19116–19128. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Wu, S.; Fan, Y.; Li, X. Influence of the mechanical properties of biomaterials on degradability, cell behaviors and signaling pathways: Current progress and challenges. Biomater. Sci. 2020, 8, 2714–2733. [Google Scholar] [CrossRef]
- Vasconcelos e Cruz, J.; Delgado, A.H.; Félix, S.; Brito, J.; Gonçalves, L.; Polido, M. Improving Properties of an Experimental Universal Adhesive by Adding a Multifunctional Dendrimer (G-IEMA): Bond Strength and Nanoleakage Evaluation. Polymers 2022, 14, 1462. [Google Scholar] [CrossRef]
- Na, S.; Collin, O.; Chowdhury, F.; Tay, B.; Ouyang, M.; Wang, Y.; Wang, N. Rapid signal transduction in living cells is a unique feature of mechanotransduction. Proc. Natl. Acad. Sci. USA 2008, 105, 6626–6631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Tytell, J.D.; Ingber, D.E. Mechanotransduction at a distance: Mechanically coupling the extracellular matrix with the nucleus. Nat. Rev. Mol. Cell Biol. 2009, 10, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kanemoto, Y.; Miyaji, H.; Nishida, E.; Miyata, S.; Mayumi, K.; Yoshino, Y.; Kato, A.; Sugaya, T.; Akasaka, T.; Nathanael, A.J. Periodontal tissue engineering using an apatite/collagen scaffold obtained by a plasma-and precursor-assisted biomimetic process. J. Periodontal Res. 2022, 57, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Kwack, K.H.; Ji, J.Y.; Park, B.; Heo, J.S. Fucoidan (Undaria pinnatifida)/Polydopamine Composite-Modified Surface Promotes Osteogenic Potential of Periodontal Ligament Stem Cells. Mar. Drugs 2022, 20, 181. [Google Scholar] [CrossRef]
- Tryba, A.M.; Krok-Borkowicz, M.; Kula, M.; Piergies, N.; Marzec, M.; Wegener, E.; Frączyk, J.; Jordan, R.; Kolesińska, B.; Scharnweber, D. Surface Functionalization of Poly (L-lactide-co-glycolide) Membranes with RGD-Grafted Poly (2-oxazoline) for Periodontal Tissue Engineering. J. Funct. Biomater. 2022, 13, 4. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Li, J.; Chen, Z.; Fan, M.; Lin, F.; Xie, Y. Electrospun Polysaccharides for Periodontal Tissue Engineering: A Review of Recent Advances and Future Perspectives. Ann. Biomed. Eng. 2022, 1–25. [Google Scholar] [CrossRef]
- Amano, A. Molecular Interaction of Porphyromonas gingivalis with Host Cells: Implication for the Microbial Pathogenesis of Periodontal Disease. J. Periodontol. 2003, 74, 90–96. [Google Scholar] [CrossRef]
- Sela, M.N. Role of Treponema denticola in periodontal diseases. Crit. Rev. Oral Biol. Med. 2001, 12, 399–413. [Google Scholar] [CrossRef]
- Sharma, A. Virulence mechanisms of Tannerella forsythia. Periodontol. 2000 2010, 54, 106. [Google Scholar] [CrossRef] [Green Version]
- Kesic, L.; Petrovic, M.; Obradovic, R.; Pejcic, A. The importance of aggregatibacter actinomycetemcomitans in etiology of periodontal disease—Mini review. Acta Med. Median. 2009, 48, 35–37. [Google Scholar]
- Lagosz-Cwik, K.B.; Wielento, A.; Lipska, W.; Kantorowicz, M.; Darczuk, D.; Kaczmarzyk, T.; Gibbs, S.; Potempa, J.; Grabiec, A.M. hTERT-immortalized gingival fibroblasts respond to cytokines but fail to mimic primary cell responses to Porphyromonas gingivalis. Sci. Rep. 2021, 11, 10770. [Google Scholar] [CrossRef] [PubMed]
- Rocha, F.G.; Ottenberg, G.; Eure, Z.G.; Davey, M.E.; Gibson III, F.C. Sphingolipid-containing outer membrane vesicles serve as a delivery vehicle to limit macrophage immune response to Porphyromonas gingivalis. Infect. Immun. 2021, 89, e00614-20. [Google Scholar] [CrossRef] [PubMed]
- Hammami, C.; Nasri, W. Antibiotics in the Treatment of Periodontitis: A Systematic Review of the Literature. Int. J. Dent. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; He, L.; Jin, D.; Zhu, Y.; Meng, H. Effect of adjunctive systemic antibiotics on microbial populations compared with scaling and root planing alone for the treatment of periodontitis: A pilot randomized clinical trial. J. Periodontol. 2021, 93, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Povšič, K.; Čuk, K.; Milavec, S.; Erčulj, V.; Seme, K.; Gašperšič, R. Systemic azithromycin as an adjunct to scaling and root planing in patients with stage III/IV periodontitis: 12-month results of a randomized controlled clinical trial. Clin. Oral Investig. 2021, 25, 5997–6006. [Google Scholar] [CrossRef] [PubMed]
- Rubio, F.; Wienecke, F.; Arnabat-Domínguez, J.; Betancourt, P. Photobiomodulation therapy and endodontic treatment of teeth with apical periodontitis using 940-nm diode laser. Report of two cases. J. Clin. Exp. Dent. 2022, 14, e298. [Google Scholar] [CrossRef]
- Sterczała, B.; Grzech-Leśniak, K.; Michel, O.; Trzeciakowski, W.; Dominiak, M.; Jurczyszyn, K. Assessment of human gingival fibroblast proliferation after laser stimulation in vitro using different laser types and wavelengths (1064, 980, 635, 450, and 405 nm)—Preliminary report. J. Pers. Med. 2021, 11, 98. [Google Scholar] [CrossRef]
- Almoudi, M.M.; Hussein, A.S.; Abu-Hassan, M.I.; Saripudin, B.; Mohamad, M.S.F. The Association of Early Childhood Caries with Salivary Antimicrobial Peptide LL37 and Mutans Streptococci. J. Clin. Pediatric Dent. 2021, 45, 330–336. [Google Scholar] [CrossRef]
- Khabbaz, H.; Karimi-Jafari, M.H.; Saboury, A.A.; BabaAli, B. Prediction of antimicrobial peptides toxicity based on their physico-chemical properties using machine learning techniques. BMC Bioinform. 2021, 22, 549. [Google Scholar] [CrossRef]
- Zou, P.; Laird, D.; Riga, E.K.; Deng, Z.; Dorner, F.; Perez-Hernandez, H.-R.; Guevara-Solarte, D.L.; Steinberg, T.; Al-Ahmad, A.; Lienkamp, K. Antimicrobial and cell-compatible surface-attached polymer networks—How the correlation of chemical structure to physical and biological data leads to a modified mechanism of action. J. Mater. Chem. B 2015, 3, 6224–6238. [Google Scholar] [CrossRef] [Green Version]
- Moreno Sancho, F.; Leira, Y.; Orlandi, M.; Buti, J.; Giannobile, W.V.; D’Aiuto, F. Cell-based therapies for alveolar bone and periodontal regeneration: Concise review. Stem Cells Transl. Med. 2019, 8, 1286–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assis, R.I.; Racca, F.; Ferreira, R.S.; Ruiz, K.G.; da Silva, R.A.; Clokie, S.J.; Wiench, M.; Andia, D.C. Osteogenic commitment of human periodontal ligament cells is predetermined by methylation, chromatin accessibility and expression of key transcription factors. Cells 2022, 11, 1126. [Google Scholar] [CrossRef] [PubMed]
- Antarianto, R.D.; Pragiwaksana, A.; Septiana, W.L.; Mazfufah, N.F.; Mahmood, A. Hepatocyte Differentiation from iPSCs or MSCs in Decellularized Liver Scaffold: Cell–ECM Adhesion, Spatial Distribution, and Hepatocyte Maturation Profile. Organogenesis 2022, 18, 2061263. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xie, T.; Sun, Y. Towards organogenesis and morphogenesis in vitro: Harnessing engineered microenvironment and autonomous behaviors of pluripotent stem cells. Integr. Biol. 2018, 10, 574–586. [Google Scholar]
- Huang, D.; Li, R.; Ren, J.; Luo, H.; Wang, W.; Zhou, C. Temporal induction of Lhx8 by optogenetic control system for efficient bone regeneration. Stem Cell Res. Ther. 2021, 12, 339. [Google Scholar] [CrossRef]
- Spagnuolo, G.; Genovese, F.; Fortunato, L.; Simeone, M.; Rengo, C.; Tatullo, M. The impact of optogenetics on regenerative medicine. Appl. Sci. 2019, 10, 173. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.S.; Chang, Y.H.; Chen, Y.Y.; Xu, J.; Yu, C.S.; Chang, S.J.; Chen, P.S.; Tsai, S.J.; Chiu, W.T. Ca2+-regulated cell migration revealed by optogenetically engineered Ca2+ oscillations. J. Cell. Physiol. 2021, 236, 4681–4693. [Google Scholar] [CrossRef]
- Wong, C.-W.; Ko, L.-N.; Huang, H.-J.; Yang, C.-S.; Hsu, S.-H. Engineered Bacteriorhodopsin May Induce Lung Cancer Cell Cycle Arrest and Suppress Their Proliferation and Migration. Molecules 2021, 26, 7344. [Google Scholar] [CrossRef]
- McDowell, J.D. An overview of epidemiology and common risk factors for oral squamous cell carcinoma. Otolaryngol. Clin. N. Am. 2006, 39, 277–294. [Google Scholar] [CrossRef]
- Espinosa-Cotton, M.; Rodman, S.N., III; Ross, K.A.; Jensen, I.J.; Sangodeyi-Miller, K.; McLaren, A.J.; Dahl, R.A.; Gibson-Corley, K.N.; Koch, A.T.; Fu, Y.-X. Interleukin-1 alpha increases anti-tumor efficacy of cetuximab in head and neck squamous cell carcinoma. J. Immunother. Cancer 2019, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Olmos, M.; Glajzer, J.; Büntemeyer, T.-O.; Frohwitter, G.; Ries, J.; Eckstein, M.; Hecht, M.; Lutz, R.; Kesting, M.R.; Weber, M. Neoadjuvant Immunotherapy of Oral Squamous Cell Carcinoma: Case Report and Assessment of Histological Response. Front. Oncol. 2021, 11, 3421–3432. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Cortes, J.; Pusztai, L.; McArthur, H.; Kümmel, S.; Bergh, J.; Denkert, C.; Park, Y.H.; Hui, R.; Harbeck, N. Pembrolizumab for early triple-negative breast cancer. N. Engl. J. Med. 2020, 382, 810–821. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, M.R.; Frazon, T.F.; Inacio, K.K.; Smiderle, F.R.; Amôr, N.G.; Dionísio, T.J.; Santos, C.F.; Rodini, C.O.; Lara, V.S. Ganoderma lucidum polysaccharides inhibit in vitro tumorigenesis, cancer stem cell properties and epithelial-mesenchymal transition in oral squamous cell carcinoma. J. Ethnopharmacol. 2022, 286, 114891. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, J.; Bandara, B.; Tilakaratne, W.M.; Kim, D. Leaf extract of Osbeckia octandra induces apoptosis in oral squamous cell carcinoma cells. BMC Complementary Med. Ther. 2022, 22, 20. [Google Scholar] [CrossRef]
- Shahbaz, K.; Asif, J.A.; Liszen, T.; Nurul, A.A.; Alam, M.K. Cytotoxic and Antioxidant Effects of Phoenix dactylifera L. (Ajwa Date Extract) on Oral Squamous Cell Carcinoma Cell Line. BioMed Res. Int. 2022, 2022, 5792830. [Google Scholar] [CrossRef]
- Ibrahim, R.S.; El-Banna, A.A. Network pharmacology-based analysis for unraveling potential cancer-related molecular targets of Egyptian propolis phytoconstituents accompanied with molecular docking and in vitro studies. RSC Adv. 2021, 11, 11610–11626. [Google Scholar]
- Wezgowiec, J.; Wieczynska, A.; Wieckiewicz, W.; Kulbacka, J.; Saczko, J.; Pachura, N.; Wieckiewicz, M.; Gancarz, R.; Wilk, K.A. Polish propolis—Chemical composition and biological effects in tongue cancer cells and macrophages. Molecules 2020, 25, 2426. [Google Scholar] [CrossRef]
- Chun, K.-S.; Kundu, J.K.; Park, K.-K.; Chung, W.-Y.; Surh, Y.-J. Inhibition of phorbol ester-induced mouse skin tumor promotion and COX-2 expression by celecoxib: C/EBP as a potential molecular target. Cancer Res. Treat. Off. J. Korean Cancer Assoc. 2006, 38, 152. [Google Scholar] [CrossRef] [Green Version]
- Tomakidi, P.; Stark, H.J.; Herold-Mende, C.; Bosch, F.X.; Steinbauer, H.; Fusenig, N.E.; Breitkreutz, D. Discriminating expression of differentiation markers evolves in transplants of benign and malignant human skin keratinocytes through stromal interactions. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2003, 200, 298–307. [Google Scholar] [CrossRef]
- Mery, B.; Rancoule, C.; Guy, J.-B.; Espenel, S.; Wozny, A.-S.; Battiston-Montagne, P.; Ardail, D.; Beuve, M.; Alphonse, G.; Rodriguez-Lafrasse, C. Preclinical models in HNSCC: A comprehensive review. Oral Oncol. 2017, 65, 51–56. [Google Scholar]
- Gawas, N.P.; Navarange, S.S.; Chovatiya, G.L.; Chaturvedi, P.; Waghmare, S.K. Establishment and characterization of novel human oral squamous cell carcinoma cell lines from advancedstage tumors of buccal mucosa. Oncol. Rep. 2019, 41, 2289–2298. [Google Scholar] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamarin, D.; Holmgaard, R.B.; Subudhi, S.K.; Park, J.S.; Mansour, M.; Palese, P.; Merghoub, T.; Wolchok, J.D.; Allison, J.P. Localized oncolytic virotherapy overcomes systemic tumor resistance to immune checkpoint blockade immunotherapy. Sci. Transl. Med. 2014, 6, 226ra32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröger, S.; Michel, J.; Meyle, J. Establishment and characterization of immortalized human gingival keratinocyte cell lines. J. Periodontal Res. 2008, 43, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, L.; Chen, H.; Jing, X.; Wang, M.; Ge, L.; Yang, J.; Zhang, M.; Tang, X. Peroxiredoxin1 Knockdown Inhibits Oral Carcinogenesis via Inducing Cell Senescence Dependent on Mitophagy. OncoTargets Ther. 2021, 14, 239. [Google Scholar] [CrossRef]
- Fischer, M.; Uxa, S.; Stanko, C.; Magin, T.M.; Engeland, K. Human papilloma virus E7 oncoprotein abrogates the p53-p21-DREAM pathway. Sci. Rep. 2017, 7, 2603. [Google Scholar]
- Hallikeri, K.; Burde, K.; Anehosur, V.; Kulkarni, B.B.; Hiremath, S.V. p53 polymorphism and association of human papillomavirus in oral submucous fibrosis and oral squamous cell carcinoma: A case–control study. J. Oral Maxillofac. Pathol. JOMFP 2019, 23, 97. [Google Scholar]
- Zhang, J.; Yu, G.; Yang, Y.; Wang, Y.; Guo, M.; Yin, Q.; Yan, C.; Tian, J.; Fu, F.; Wang, H. A small-molecule inhibitor of MDMX suppresses cervical cancer cells via the inhibition of E6-E6AP-p53 axis. Pharmacol. Res. 2022, 177, 106128. [Google Scholar] [CrossRef]
- Rodolico, V.; Arancio, W.; Amato, M.C.; Aragona, F.; Cappello, F.; Di Fede, O.; Pannone, G.; Campisi, G. Hypoxia inducible factor-1 alpha expression is increased in infected positive HPV16 DNA oral squamous cell carcinoma and positively associated with HPV16 E7 oncoprotein. Infect. Agents Cancer 2011, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Berdugo, J.; Rooper, L.M.; Chiosea, S.I. RB1, p16, and Human Papillomavirus in Oropharyngeal Squamous Cell Carcinoma. Head Neck Pathol. 2021, 15, 1109–1118. [Google Scholar] [CrossRef]
- Sudhakaran, A.; Hallikeri, K.; Babu, B. p16 as an independent marker for detection of high-risk HPV in oral submucous fibrosis and oral squamous cell carcinoma. Indian J. Pathol. Microbiol. 2019, 62, 523. [Google Scholar] [PubMed]
- Pannone, G.; Santoro, A.; Carinci, F.; Bufo, P.; Papagerakis, S.; Rubini, C.; Campisi, G.; Giovannelli, L.; Contaldo, M.; Serpico, R. Double demonstration of oncogenic high risk human papilloma virus DNA and HPV-E7 protein in oral cancers. Int. J. Immunopathol. Pharmacol. 2011, 24 (Suppl. S2), 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Søland, T.M.; Bjerkli, I.H.; Georgsen, J.B.; Schreurs, O.; Jebsen, P.; Laurvik, H.; Sapkota, D. High-risk human papilloma virus was not detected in a Norwegian cohort of oral squamous cell carcinoma of the mobile tongue. Clin. Exp. Dent. Res. 2021, 7, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Vipparthi, K.; Patel, A.K.; Ghosh, S.; Das, S.; Das, C.; Das, K.; Sarkar, A.; Thatikonda, V.; Pal, B.; Arora, N. Two novel cell culture models of buccal mucosal oral cancer from patients with no risk-habits of tobacco smoking or chewing. Oral Oncol. 2021, 113, 105131. [Google Scholar] [CrossRef] [PubMed]
- Chitturi Suryaprakash, R.T.; Kujan, O.; Shearston, K.; Farah, C.S. Three-dimensional cell culture models to investigate oral carcinogenesis: A scoping review. Int. J. Mol. Sci. 2020, 21, 9520. [Google Scholar] [CrossRef]
- Chamulitrat, W.; Schmidt, R.; Tomakidi, P.; Stremmel, W.; Chunglok, W.; Kawahara, T.; Rokutan, K. Association of gp91phox homolog Nox1 with anchorage-independent growth and MAP kinase-activation of transformed human keratinocytes. Oncogene 2003, 22, 6045–6053. [Google Scholar] [CrossRef] [Green Version]
- Roesch-Ely, M.; Steinberg, T.; Bosch, F.X.; Mussig, E.; Whitaker, N.; Wiest, T.; Kohl, A.; Komposch, G.; Tomakidi, P. Organotypic co-cultures allow for immortalized human gingival keratinocytes to reconstitute a gingival epithelial phenotype in vitro. Differentiation 2006, 74, 622–637. [Google Scholar] [CrossRef]
- Cheng, H.; Yang, X.; Si, H.; Saleh, A.D.; Xiao, W.; Coupar, J.; Gollin, S.M.; Ferris, R.L.; Issaeva, N.; Yarbrough, W.G. Genomic and transcriptomic characterization links cell lines with aggressive head and neck cancers. Cell Rep. 2018, 25, 1332–1345.e5. [Google Scholar] [CrossRef] [Green Version]
- Das, D.; Ghosh, S.; Maitra, A.; Biswas, N.K.; Panda, C.K.; Roy, B.; Sarin, R.; Majumder, P.P. Epigenomic dysregulation-mediated alterations of key biological pathways and tumor immune evasion are hallmarks of gingivo-buccal oral cancer. Clin. Epigenet. 2019, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Heim, J.B.; McDonald, C.A.; Wyles, S.P.; Sominidi-Damodaran, S.; Squirewell, E.J.; Li, M.; Motsonelidze, C.; Böttcher, R.T.; van Deursen, J.; Meves, A. FAK auto-phosphorylation site tyrosine 397 is required for development but dispensable for normal skin homeostasis. PLoS ONE 2018, 13, e0200558. [Google Scholar] [CrossRef]
- Wang, X.; Steinberg, T.; Dieterle, M.P.; Ramminger, I.; Husari, A.; Tomakidi, P. FAK Shutdown: Consequences on Epithelial Morphogenesis and Biomarker Expression Involving an Innovative Biomaterial for Tissue Regeneration. Int. J. Mol. Sci. 2021, 22, 9774. [Google Scholar] [CrossRef] [PubMed]
- Husari, A.; Hülter-Hassler, D.; Steinberg, T.; Schulz, S.D.; Tomakidi, P. Disruption of adherens junction and alterations in YAP-related proliferation behavior as part of the underlying cell transformation process of alcohol-induced oral carcinogenesis. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2018, 1865, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Piemonte, E.; Lazos, J.; Belardinelli, P.; Secchi, D.; Brunotto, M.; Lanfranchi-Tizeira, H. Oral cancer associated with chronic mechanical irritation of the oral mucosa. Med. Oral Patol. Oral Cirugía Bucal 2018, 23, e151. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.A.; Kheur, S.; Varadarajan, S.; Parveen, S.; Dewan, H.; Alhazmi, Y.A.; Raj, T.A.; Testarelli, L.; Patil, S. Chronic mechanical irritation and oral squamous cell carcinoma: A systematic review and meta-analysis. Bosn. J. Basic Med. Sci. 2021, 21, 647. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-S.; Lee, K.-W.; Huang, J.-L.; Liu, Y.-S.; Juo, S.-H.H.; Kuo, W.-R.; Chang, J.-G.; Lin, C.-S.; Jong, Y.-J. Arecoline, a major alkaloid of areca nut, inhibits p53, represses DNA repair, and triggers DNA damage response in human epithelial cells. Toxicology 2008, 249, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Qin, N.; Song, X.; Jiang, C.; Li, T.; Ji, P.; Li, Y.; Ding, D.; Wang, C.; Dai, J. Genomic Signature of Mismatch Repair Deficiency in Areca Nut–Related Oral Cancer. J. Dent. Res. 2020, 99, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Batta, N.; Pandey, M. Mutational spectrum of tobacco associated oral squamous carcinoma and its therapeutic significance. World J. Surg. Oncol. 2019, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- López-Verdín, S.; de la Luz Martínez-Fierro, M.; Garza-Veloz, I.; Zamora-Perez, A.; Grajeda-Cruz, J.; González-González, R.; Molina-Frechero, N.; Arocena, M.; Bologna-Molina, R. E-Cadherin gene expression in oral cancer: Clinical and prospective data. Med. Oral Patol. Oral Cir. Bucal 2019, 24, e444. [Google Scholar] [CrossRef]
- Li, M.; Rao, X.; Cui, Y.; Zhang, L.; Li, X.; Wang, B.; Zheng, Y.; Teng, L.; Zhou, T.; Zhuo, W. The keratin 17/YAP/IL6 axis contributes to E-cadherin loss and aggressiveness of diffuse gastric cancer. Oncogene 2022, 41, 770–781. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Advani, J.; Rajagopalan, P.; Patel, K.; Nanjappa, V.; Solanki, H.S.; Patil, A.H.; Bhat, F.A.; Mathur, P.P.; Nair, B. Cigarette smoke and chewing tobacco alter expression of different sets of miRNAs in oral keratinocytes. Sci. Rep. 2018, 8, 7040. [Google Scholar]
- Priya, N.; Nayak, R.; Bhat, K.; Kugaji, M.; Vijayalakshmi, K.; Rao, K. Site specificity and expression profile of miR-21 in oral squamous cell carcinoma. J. Oral Maxillofac. Pathol. JOMFP 2021, 25, 154. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, I.; Anderson, A.; O’Sullivan, J. The interplay of the oral microbiome and alcohol consumption in oral squamous cell carcinomas. Oral Oncol. 2020, 110, 105011. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, F.D.; Coveñas, R. Biochemical Mechanisms Associating Alcohol Use Disorders with Cancers. Cancers 2021, 13, 3548. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Okuyama, A.; Hoshino, M.; Nishimura, M.; Kikuchi, K. Influence of Acetaldehyde on Oral Epithelial Cells. J. Oral Cancer Res. 2021, 4, 44–48. [Google Scholar]
- Chamulitrat, W.; Schmidt, R.; Chunglok, W.; Kohl, A.; Tomakidi, P. Epithelium and fibroblast-like phenotypes derived from HPV16 E6/E7-immortalized human gingival keratinocytes following chronic ethanol treatment. Eur. J. Cell Biol. 2003, 82, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, P.F.; Rich, A.; Coates, D.E. Effects of galectin-1 inhibitor OTX008 on oral squamous cell carcinoma cells in vitro and the role of AP-1 and the MAPK/ERK pathway. Arch. Oral Biol. 2022, 134, 105335. [Google Scholar] [CrossRef]
- Chunglok, W.; Ittarat, W.; Tomakidi, P.; Schmidt, R.; Stremmel, W.; Chamulitrat, W. Human gingival mucosal keratinocytes exhibiting anchorage-independent growth express increased inducible nitric oxide synthase: Regulation by MAP kinases. Nitric Oxide 2004, 11, 237–246. [Google Scholar] [CrossRef]
- Xue, Q.; Yan, Y.; Zhang, R.; Xiong, H. Regulation of iNOS on Immune Cells and Its Role in Diseases. Int. J. Mol. Sci. 2018, 19, 3805. [Google Scholar] [CrossRef] [Green Version]
- Varghese, S.S.; Sunil, P.; Madhavan, R.N. Expression of inducible nitric oxide synthase (iNOS) in oral precancer and oral squamous cell carcinoma: An immunohistochemical study. Cancer Biomark. 2011, 8, 155–160. [Google Scholar] [CrossRef]
- Silva Servato, J.P.; Ueira Vieira, C.; de Faria, P.R.; Cardoso, S.V.; Loyola, A.M. The importance of inducible nitric oxide synthase and nitrotyrosine as prognostic markers for oral squamous cell carcinoma. J. Oral Pathol. Med. 2019, 48, 967–975. [Google Scholar] [CrossRef]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Rathinam, M.K.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The soft agar colony formation assay. JoVE J. Vis. Exp. 2014, 92, e51998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Wang, L.; Jin, Y.; Lu, P.; Wang, L.-L.; Bai, N.; Li, G.; Zhu, P.; Wang, Y.; Zhang, J.; et al. Tuning the Rigidity of Silk Fibroin for the Transfer of Highly Stretchable Electronics. Adv. Funct. Mater. 2020, 30, 2001518. [Google Scholar] [CrossRef]
- Guo, M.; Mu, Y.; Yu, D.; Li, J.; Chen, F.; Wei, B.; Bi, S.; Yu, J.; Liang, F. Comparison of the expression of TGF-β1, E-cadherin, N-cadherin, TP53, RB1CC1 and HIF-1α in oral squamous cell carcinoma and lymph node metastases of humans and mice. Oncol. Lett. 2018, 15, 1639–1645. [Google Scholar] [CrossRef]
- Kato, K.; Shimasaki, M.; Kato, T.; Segami, N.; Ueda, Y. Expression of sphingosine kinase-1 is associated with invasiveness and poor prognosis of oral squamous cell carcinoma. Anticancer Res. 2018, 38, 1361–1368. [Google Scholar] [PubMed]
- Wangmo, C.; Charoen, N.; Jantharapattana, K.; Dechaphunkul, A.; Thongsuksai, P. Epithelial–mesenchymal transition predicts survival in oral squamous cell carcinoma. Pathol. Oncol. Res. 2020, 26, 1511–1518. [Google Scholar] [CrossRef]
- Burassakarn, A.; Pientong, C.; Sunthamala, N.; Chuerduangphui, J.; Vatanasapt, P.; Patarapadungkit, N.; Kongyingyoes, B.; Ekalaksananan, T. Aberrant gene promoter methylation of E-cadherin, p16 INK4a, p14 ARF, and MGMT in Epstein–Barr virus-associated oral squamous cell carcinomas. Med. Oncol. 2017, 34, 128. [Google Scholar] [CrossRef]
- Pannone, G.; Santoro, A.; Feola, A.; Bufo, P.; Papagerakis, P.; Lo Muzio, L.; Staibano, S.; Ionna, F.; Longo, F.; Franco, R. The role of E-cadherin down-regulation in oral cancer: CDH1 gene expression and epigenetic blockage. Curr. Cancer Drug Targets 2014, 14, 115–127. [Google Scholar] [CrossRef]
- Muzio, L.L.; Campisi, G.; Farina, A.; Rubini, C.; Pannone, G.; Serpico, R.; Laino, G.; De Lillo, A.; Carinci, F. P-cadherin expression and survival rate in oral squamous cell carcinoma: An immunohistochemical study. BMC Cancer 2005, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Seppälä, M.; Jauhiainen, L.; Tervo, S.; Al-Samadi, A.; Rautiainen, M.; Salo, T.; Lehti, K.; Monni, O.; Hautaniemi, S.; Tynninen, O. The expression and prognostic relevance of CDH3 in tongue squamous cell carcinoma. Apmis 2021, 129, 717–728. [Google Scholar] [CrossRef]
- Kumar, V.; Panda, A.; Dash, K.C.; Bhuyan, L.; Mahapatra, N.; Mishra, P. Immunohistochemical Expression of the Epithelial to Mesenchymal Transition Proteins E-cadherin and β-catenin in Grades of Oral Squamous Cell Carcinoma. J. Pharm. Bioallied Sci. 2021, 13 (Suppl. S1), S555–S560. [Google Scholar]
- Mis, M.; O’Brien, S.; Steinhart, Z.; Lin, S.; Hart, T.; Moffat, J.; Angers, S. IPO11 mediates βcatenin nuclear import in a subset of colorectal cancers. J. Cell Biol. 2020, 219, e201903017. [Google Scholar] [CrossRef] [PubMed]
- Reyes, M.; Peña-Oyarzún, D.; Silva, P.; Venegas, S.; Criollo, A.; Torres, V.A. Nuclear accumulation of β-catenin is associated with endosomal sequestration of the destruction complex and increased activation of Rab5 in oral dysplasia. FASEB J. 2020, 34, 4009–4025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belgardt, E.; Steinberg, T.; Husari, A.; Dieterle, M.P.; Hülter-Hassler, D.; Jung, B.; Tomakidi, P. Force-responsive Zyxin modulation in periodontal ligament cells is regulated by YAP rather than TAZ. Cell. Signal. 2020, 72, 109662. [Google Scholar] [CrossRef] [PubMed]
- Barcelona-Estaje, E.; Dalby, M.J.; Cantini, M.; Salmeron-Sanchez, M. You Talking to Me? Cadherin and Integrin Crosstalk in Biomaterial Design. Adv. Healthc. Mater. 2021, 10, 2002048. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Cheng, H.; Gao, R.; Mu, C.; Chen, L.; Wu, S.; Chen, Q.; Zhu, Y. Zyxin-Siah2–Lats2 axis mediates cooperation between Hippo and TGF-β signalling pathways. Nat. Commun. 2016, 7, 11123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, M.; Noguchi, K.; Nakano, Y.; Segawa, E.; Zushi, Y.; Takaoka, K.; Kishimoto, H.; Hashimoto-Tamaoki, T.; Urade, M. [Corrigendum] Functional analysis of Zyxin in cell migration and invasive potential of oral squamous cell carcinoma cells. Int. J. Oncol. 2016, 49, 2188. [Google Scholar] [PubMed]
- Omori, H.; Nishio, M.; Masuda, M.; Miyachi, Y.; Ueda, F.; Nakano, T.; Sato, K.; Mimori, K.; Taguchi, K.; Hikasa, H. YAP1 is a potent driver of the onset and progression of oral squamous cell carcinoma. Sci. Adv. 2020, 6, eaay3324. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, K.; Fujii, S.; Matsumoto, S.; Tajiri, Y.; Kikuchi, A.; Kiyoshima, T. YAP signaling induces PIEZO1 to promote oral squamous cell carcinoma cell proliferation. J. Pathol. 2021, 253, 80–93. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, T.; Wu, Y.; Xu, H.; Xie, C.; Dong, Y.; Zhong, L.; Wang, Z.; Zhao, H.; Zhou, Y. GPR39 overexpression in OSCC promotes YAP-sustained malignant progression. J. Dent. Res. 2020, 99, 949–958. [Google Scholar] [CrossRef]
- Meng, Z.; Qiu, Y.; Lin, K.C.; Kumar, A.; Placone, J.K.; Fang, C.; Wang, K.-C.; Lu, S.; Pan, M.; Hong, A.W. RAP2 mediates mechanoresponses of the Hippo pathway. Nature 2018, 560, 655–660. [Google Scholar] [CrossRef] [Green Version]
- Paradowska-Stolarz, A.; Wieckiewicz, M.; Owczarek, A.; Wezgowiec, J. Natural Polymers for the Maintenance of Oral Health: Review of Recent Advances and Perspectives. Int. J. Mol. Sci. 2021, 22, 10337. [Google Scholar] [CrossRef]
- Samiei, M.; Alipour, M.; Khezri, K.; Saadat, Y.R.; Forouhandeh, H.; Abdolahinia, E.D.; Vahed, S.Z.; Sharifi, S.; Dizaj, S.M. Application of collagen and mesenchymal stem cells in regenerative dentistry. Curr. Stem Cell Res. Ther. 2021. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Shanbhag, S.; Mustafa, K. Scaffolds in Periodontal Regenerative Treatment. Dent. Clin. 2022, 66, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Dehghan-Baniani, D.; Mehrjou, B.; Chu, P.K.; Wu, H. A Biomimetic Nano-Engineered Platform for Functional Tissue Engineering of Cartilage Superficial Zone. Adv. Healthc. Mater. 2021, 10, 2001018. [Google Scholar] [CrossRef]
- Lin, H.-H.; Chao, P.-H.G.; Tai, W.-C.; Chang, P.-C. 3D-Printed Collagen-Based Waveform Microfibrous Scaffold for Periodontal Ligament Reconstruction. Int. J. Mol. Sci. 2021, 22, 7725. [Google Scholar] [CrossRef]
- Zheng, L.; Shi, Q.; Na, J.; Liu, N.; Guo, Y.; Fan, Y. Platelet-Derived Growth Factor Receptor-α and β are Involved in Fluid Shear Stress Regulated Cell Migration in Human Periodontal Ligament Cells. Cell. Mol. Bioeng. 2019, 12, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Asparuhova, M.B.; Stähli, A.; Guldener, K.; Sculean, A. A Novel Volume-Stable Collagen Matrix Induces Changes in the Behavior of Primary Human Oral Fibroblasts, Periodontal Ligament, and Endothelial Cells. Int. J. Mol. Sci. 2021, 22, 4051. [Google Scholar] [CrossRef]
- Guarnieri, R.; Reda, R.; Di Nardo, D.; Miccoli, G.; Zanza, A.; Testarelli, L. In Vitro Direct and Indirect Cytotoxicity Comparative Analysis of One Pre-Hydrated versus One Dried Acellular Porcine Dermal Matrix. Materials 2022, 15, 1937. [Google Scholar] [CrossRef]
- Schmitt, C.M.; Brückbauer, P.; Schlegel, K.A.; Buchbender, M.; Adler, W.; Matta, R.E. Volumetric soft tissue alterations in the early healing phase after peri-implant soft tissue contour augmentation with a porcine collagen matrix versus the autologous connective tissue graft: A controlled clinical trial. J. Clin. Periodontol. 2021, 48, 146–163. [Google Scholar] [CrossRef]
- Angarano, M.; Schulz, S.; Fabritius, M.; Vogt, R.; Steinberg, T.; Tomakidi, P.; Friedrich, C.; Mülhaupt, R. Layered gradient nonwovens of in situ crosslinked electrospun collagenous nanofibers used as modular scaffold systems for soft tissue regeneration. Adv. Funct. Mater. 2013, 23, 3277–3285. [Google Scholar] [CrossRef]
- Schulz, S.; Angarano, M.; Fabritius, M.; Mülhaupt, R.; Dard, M.; Obrecht, M.; Tomakidi, P.; Steinberg, T. Nonwoven-based gelatin/polycaprolactone membrane proves suitability in a preclinical assessment for treatment of soft tissue defects. Tissue Eng. A 2014, 20, 1935–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, C.; Hipler, U.-C.; Elsner, P.; Tittelbach, J. Keratinocyte and Fibroblast Wound Healing In Vitro Is Repressed by Non-Optimal Conditions but the Reparative Potential Can Be Improved by Water-Filtered Infrared A. Biomedicines 2021, 9, 1802. [Google Scholar] [CrossRef] [PubMed]
- Khorolsuren, Z.; Lang, O.; Pallinger, E.; Foldes, A.; Szabolcs, G.G.; Varga, G.; Mezo, G.; Vag, J.; Kohidai, L. Functional and cell surface characteristics of periodontal ligament cells (PDLCs) on RGD-synthetic polypeptide conjugate coatings. J. Periodontal Res. 2020, 55, 713–723. [Google Scholar] [CrossRef]
- Tribst, J.P.M.; Dal Piva, A.M.D.O.; Ausiello, P.; De Benedictis, A.; Bottino, M.A.; Borges, A.L.S. Biomechanical analysis of a custom-made mouthguard reinforced with different Elastic Modulus laminates during a simulated maxillofacial trauma. Craniomaxillofac. Trauma Reconstr. 2021, 14, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Hunter-Featherstone, E.; Young, N.; Chamberlain, K.; Cubillas, P.; Hulette, B.; Wei, X.; Tiesman, J.P.; Bascom, C.C.; Benham, A.M.; Goldberg, M.W. Culturing Keratinocytes on Biomimetic Substrates Facilitates Improved Epidermal Assembly In Vitro. Cells 2021, 10, 1177. [Google Scholar] [CrossRef] [PubMed]
- Kirschneck, C.; Thuy, M.; Leikam, A.; Memmert, S.; Deschner, J.; Damanaki, A.; Spanier, G.; Proff, P.; Jantsch, J.; Schröder, A. Role and Regulation of Mechanotransductive HIF-1α Stabilisation in Periodontal Ligament Fibroblasts. Int. J. Mol. Sci. 2020, 21, 9530. [Google Scholar] [CrossRef]
- Sun, C.; Rankovic, M.J.; Folwaczny, M.; Otto, S.; Wichelhaus, A.; Baumert, U. Effect of Tension on Human Periodontal Ligament Cells: Systematic Review and Network Analysis. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef]
- Steinberg, T.; Schulz, S.; Spatz, J.P.; Grabe, N.; Mussig, E.; Kohl, A.; Komposch, G.; Tomakidi, P. Early keratinocyte differentiation on micropillar interfaces. Nano Lett. 2007, 7, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Mussig, E.; Schulz, S.; Spatz, J.P.; Ziegler, N.; Tomakidi, P.; Steinberg, T. Soft micropillar interfaces of distinct biomechanics govern behaviour of periodontal cells. Eur. J. Cell Biol. 2010, 89, 315–325. [Google Scholar] [CrossRef]
- Barua, N.; Huang, L.; Li, C.; Yang, Y.; Luo, M.; Wei, W.I.; Wong, K.T.; Lo, N.W.S.; Kwok, K.O.; Ip, M. Comparative Study of Two-Dimensional (2D) vs. Three-Dimensional (3D) Organotypic Kertatinocyte-Fibroblast Skin Models for Staphylococcus aureus (MRSA) Infection. Int. J. Mol. Sci. 2021, 23, 299. [Google Scholar] [CrossRef]
- Stark, H.-J.; Baur, M.; Breitkreutz, D.; Mirancea, N.; Fusenig, N.E. Organotypic keratinocyte cocultures in defined medium with regular epidermal morphogenesis and differentiation. J. Investig. Dermatol. 1999, 112, 681–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jedrusik, N.; Meyen, C.; Finkenzeller, G.; Stark, G.B.; Meskath, S.; Schulz, S.D.; Steinberg, T.; Eberwein, P.; Strassburg, S.; Tomakidi, P. Nanofibered gelatin-based nonwoven elasticity promotes epithelial Histogenesis. Adv. Healthc. Mater. 2018, 7, 1700895. [Google Scholar] [CrossRef] [PubMed]
- Strassburg, S.; Caduc, M.; Stark, G.B.; Jedrusik, N.; Tomakidi, P.; Steinberg, T.; Simunovic, F.; Finkenzeller, G. In vivo evaluation of an electrospun gelatin nonwoven mat for regeneration of epithelial tissues. J. Biomed. Mater. Res. A 2019, 107, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Jedrusik, N.; Steinberg, T.; Husari, A.; Volk, L.; Wang, X.; Finkenzeller, G.; Strassburg, S.; Tomakidi, P. Gelatin nonwovens-based epithelial morphogenesis involves a signaling axis comprising EGF-receptor, MAP kinases ERK 1/2, and β1 integrin. J. Biomed. Mater. Res. A 2019, 107, 663–677. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Hu, F.; Rausch-Fan, X.; Steinberg, T.; Lan, Z.; Zhang, X. The effect of modifying the nanostructure of gelatin fiber scaffolds on early angiogenesis in vitro and in vivo. Biomed. Mater. 2021, 17, 015010. [Google Scholar] [CrossRef]
- Lee, J.B.; Ko, Y.-G.; Cho, D.; Park, W.H.; Kim, B.N.; Lee, B.C.; Kang, I.-K.; Kwon, O.H. Modification of PLGA nanofibrous mats by electron beam irradiation for soft tissue regeneration. J. Nanomater. 2015, 16, 136. [Google Scholar] [CrossRef]
- Saveleva, M.S.; Ivanov, A.N.; Chibrikova, J.A.; Abalymov, A.A.; Surmeneva, M.A.; Surmenev, R.A.; Parakhonskiy, B.V.; Lomova, M.V.; Skirtach, A.G.; Norkin, I.A. Osteogenic Capability of Vaterite-Coated Nonwoven Polycaprolactone Scaffolds for In Vivo Bone Tissue Regeneration. Macromol. Biosci. 2021, 21, 2100266. [Google Scholar] [CrossRef]
- Abalymov, A.; Van der Meeren, L.; Skirtach, A.G.; Parakhonskiy, B.V. Identification and analysis of key parameters for the ossification on particle functionalized composites hydrogel materials. ACS Appl. Mater. Interfaces 2020, 12, 38862–38872. [Google Scholar] [CrossRef]
- Green, D.W.; Bolland, B.J.; Kanczler, J.M.; Lanham, S.A.; Walsh, D.; Mann, S.; Oreffo, R.O. Augmentation of skeletal tissue formation in impaction bone grafting using vaterite microsphere biocomposites. Biomaterials 2009, 30, 1918–1927. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, Y.J.; Chao, J.R.; Kim, D.Y.; Sultan, M.T.; Lee, H.J.; Lee, J.M.; Lee, J.S.; Lee, O.J.; Hong, H.; et al. Rapidly photocurable silk fibroin sealant for clinical applications. NPG Asia Mater. 2020, 12, 46. [Google Scholar] [CrossRef]
- Yang, C.; Shang, S.; Shou, D.; Ran, L.; Lan, G.; Hu, E. Transforming natural silk nonwovens into robust bioadhesives for in vivo tissue amendment. J. Clean. Prod. 2021, 314, 127996. [Google Scholar] [CrossRef]
- Mikołajczyk, T.; Boguń, M.; Błażewicz, M.; Piekarczyk, I. Effect of spinning conditions on the structure and properties of PAN fibers containing nano-hydroxyapatite. J. Appl. Polym. Sci. 2006, 100, 2881–2888. [Google Scholar] [CrossRef]
- Frączyk, J.; Magdziarz, S.; Stodolak-Zych, E.; Dzierzkowska, E.; Puchowicz, D.; Kamińska, I.; Giełdowska, M.; Boguń, M. Chemical Modification as a Method of Improving Biocompatibility of Carbon Nonwovens. Materials 2021, 14, 3198. [Google Scholar] [CrossRef] [PubMed]
- Mertgen, A.-S.; Trossmann, V.T.; Guex, A.G.; Maniura-Weber, K.; Scheibel, T.; Rottmar, M. Multifunctional Biomaterials: Combining Material Modification Strategies for Engineering of Cell-Contacting Surfaces. ACS Appl. Mater. Interfaces 2020, 12, 21342–21367. [Google Scholar] [CrossRef] [PubMed]
- Gübeli, R.J.; Hövermann, D.; Seitz, H.; Rebmann, B.; Schoenmakers, R.G.; Ehrbar, M.; Charpin-El Hamri, G.; Daoud-El Baba, M.; Werner, M.; Müller, M. Remote-Controlled Hydrogel Depots for Time-Scheduled Vaccination. Adv. Funct. Mater. 2013, 23, 5355–5362. [Google Scholar] [CrossRef]
- Laird, D.; El-Baba, M.D.; Charpin-El Hamri, G.; Eberwein, P.; Nelson, K.; Tomakidi, P.; Steinberg, T. In vitro and in vivo biocompatibility evaluation of a novobiocin stimulus-responsive poly (ethylene glycol)-based hydrogel designed for soft tissue regeneration. J. Bioact. Compat. Polym. 2015, 30, 319–339. [Google Scholar] [CrossRef]
- Denzinger, M.; Link, A.; Kurz, J.; Krauss, S.; Thoma, R.; Schlensak, C.; Wendel, H.P.; Krajewski, S. Keratinocyte growth factor modified messenger RNA accelerating cell proliferation and migration of keratinocytes. Nucleic Acid Ther. 2018, 28, 335–347. [Google Scholar] [CrossRef]
- Kapp, T.G.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.V.; Cavalcanti-Adam, E.A.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.-J.; Mas-Moruno, C. A comprehensive evaluation of the activity and selectivity profile of ligands for RGD-binding integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Li, Z.; Ji, Q.; Sun, H.; Liang, Y.; Yang, P. Carboxymethyl chitin or chitosan for osteoinduction effect on the human periodontal ligament stem cells. Dent. Mater. J. 2022, 2021–2250. [Google Scholar] [CrossRef]
- Hsieh, H.-Y.; Yao, C.-C.; Hsu, L.-F.; Tsai, L.-H.; Jeng, J.-H.; Young, T.-H.; Chen, Y.-J. Biological properties of human periodontal ligament cell spheroids cultivated on chitosan and polyvinyl alcohol membranes. J. Formos. Med. Assoc. 2022, in press. [CrossRef]
- Yan, X.-Z.; van den Beucken, J.J.; Cai, X.; Yu, N.; Jansen, J.A.; Yang, F. Periodontal tissue regeneration using enzymatically solidified chitosan hydrogels with or without cell loading. Tissue Eng. A 2015, 21, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Duan, X.; Chen, Y.; Liu, B.; Chen, G. Dental stem cell-derived extracellular vesicles as promising therapeutic agents in the treatment of diseases. Int. J. Oral Sci. 2022, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Baranov, N.; Popa, M.; Atanase, L.I.; Ichim, D.L. Polysaccharide-based drug delivery systems for the treatment of periodontitis. Molecules 2021, 26, 2735. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuang, S.; Zhang, Y.; Yang, M.; Qin, W.; Shi, X.; Lin, Z. Chitosan hydrogel incorporated with dental pulp stem cell-derived exosomes alleviates periodontitis in mice via a macrophage-dependent mechanism. Bioact. Mater. 2020, 5, 1113–1126. [Google Scholar] [CrossRef]
- Lin, J.; He, Z.; Liu, F.; Feng, J.; Huang, C.; Sun, X.; Deng, H. Hybrid hydrogels for synergistic periodontal antibacterial treatment with sustained drug release and NIR-responsive photothermal effect. Int. J. Nanomed. 2020, 15, 5377. [Google Scholar] [CrossRef] [PubMed]
- Abusleme, L.; Dupuy, A.K.; Dutzan, N.; Silva, N.; Burleson, J.A.; Strausbaugh, L.D.; Gamonal, J.; Diaz, P.I. The subgingival microbiome in health and periodontitis and its relationship with community biomass and inflammation. ISME J. 2013, 7, 1016–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kida, D.; Karolewicz, B.; Junka, A.; Sender-Janeczek, A.; Duś, I.; Marciniak, D.; Szulc, M. Metronidazole-loaded porous matrices for local periodontitis treatment: In vitro evaluation and in vivo pilot study. Appl. Sci. 2019, 9, 4545. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Kantorski, K.Z.; Dubey, N.; Daghrery, A.; Fenno, J.C.; Mishina, Y.; Chan, H.-L.; Mendonça, G.; Bottino, M.C. Personalized and Defect-Specific Antibiotic-Laden Scaffolds for Periodontal Infection Ablation. ACS Appl. Mater. Interfaces 2021, 13, 49642–49657. [Google Scholar] [CrossRef]
- Li, L.; Jiang, H.; Chen, R.; Zhou, J.; Xiao, Y.; Zhang, Y.; Yan, F. Human β-defensin 3 gene modification promotes the osteogenic differentiation of human periodontal ligament cells and bone repair in periodontitis. Int. J. Oral Sci. 2020, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stulz, A.; Vogt, A.; Saar, J.S.; Akil, L.; Lienkamp, K.; Hoernke, M. Quantified membrane permeabilization indicates the lipid selectivity of membrane-active antimicrobials. Langmuir 2019, 35, 16366–16376. [Google Scholar] [CrossRef]
- Beckloff, N.; Laube, D.; Castro, T.; Furgang, D.; Park, S.; Perlin, D.; Clements, D.; Tang, H.; Scott, R.W.; Tew, G.N. Activity of an antimicrobial peptide mimetic against planktonic and biofilm cultures of oral pathogens. Antimicrob. Agents Chemother. 2007, 51, 4125–4132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, J.; Scott, R.; Diamond, G. Activity of antimicrobial peptide mimetics in the oral cavity: II. Activity against periopathogenic biofilms and anti-inflammatory activity. Mol. Oral Microbiol. 2010, 25, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Al-Ahmad, A.; Laird, D.; Zou, P.; Tomakidi, P.; Steinberg, T.; Lienkamp, K. Nature-inspired antimicrobial polymers–assessment of their potential for biomedical applications. PLoS ONE 2013, 8, e73812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Zhang, Y.; Li, L.; Fu, H.; Yang, W.; Yan, F. Human β-defensin 3-combined gold nanoparticles for enhancement of osteogenic differentiation of human periodontal ligament cells in inflammatory microenvironments. Int. J. Nanomed. 2018, 13, 555. [Google Scholar] [CrossRef] [Green Version]
- Chidambar, C.K.; Shankar, S.M.; Raghu, P.; Gururaj, S.B.; Bushan, K.S. Detection of Enterococcus faecalis in subgingival biofilms of healthy, gingivitis, and chronic periodontitis subjects. J. Indian Soc. Periodontol. 2019, 23, 416. [Google Scholar] [CrossRef]
- O’Connor, A.M.; McManus, B.A.; Kinnevey, P.M.; Brennan, G.I.; Fleming, T.E.; Cashin, P.J.; O’Sullivan, M.; Polyzois, I.; Coleman, D.C. Significant enrichment and diversity of the staphylococcal arginine catabolic mobile element ACME in Staphylococcus epidermidis isolates from subgingival peri-implantitis sites and periodontal pockets. Front. Microbiol. 2018, 9, 1558. [Google Scholar] [CrossRef]
- Kumar, P.; Takayesu, A.; Abbasi, U.; Kalathottukaren, M.T.; Abbina, S.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptide–polymer conjugates with high activity: Influence of polymer molecular weight and peptide sequence on antimicrobial activity, proteolysis, and biocompatibility. ACS Appl. Mater. Interfaces 2017, 9, 37575–37586. [Google Scholar] [CrossRef]
- Kuroda, K.; Caputo, G.A. Antimicrobial polymers as synthetic mimics of host-defense peptides. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2013, 5, 49–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scudiero, O.; Nigro, E.; Cantisani, M.; Colavita, I.; Leone, M.; Mercurio, F.A.; Galdiero, M.; Pessi, A.; Daniele, A.; Salvatore, F. Design and activity of a cyclic mini-β-defensin analog: A novel antimicrobial tool. Int. J. Nanomed. 2015, 10, 6523. [Google Scholar]
- Patil, P.C.; Tan, J.; Demuth, D.R.; Luzzio, F.A. 1, 2, 3-Triazole-based inhibitors of Porphyromonas gingivalis adherence to oral streptococci and biofilm formation. Bioorg. Med. Chem. 2016, 24, 5410–5417. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.; Patil, P.C.; Luzzio, F.A.; Demuth, D.R. In vitro and in vivo activity of peptidomimetic compounds that target the periodontal pathogen Porphyromonas gingivalis. Antimicrob. Agents Chemother. 2018, 62, e00400-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, T.; Dannewitz, B.; Tomakidi, P.; Hoheisel, J.; Müssig, E.; Kohl, A.; Nees, M. Analysis of interleukin-1β-modulated mRNA gene transcription in human gingival keratinocytes by epithelia-specific cDNA microarrays. J. Periodontal Res. 2006, 41, 426–446. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Yang, F.; Jansen, J.A.; Walboomers, X.F. Lipoxin suppresses inflammation via the TLR4/MyD88/NF-κB pathway in periodontal ligament cells. Oral Dis. 2020, 26, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.W.; Shin, Y.Y.; Seo, Y.; Kim, H.-S. Therapeutic functions of stem cells from oral cavity: An update. Int. J. Mol. Sci. 2020, 21, 4389. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.L.; Holanda-Afonso, R.C.; Moura-Neto, V.; Bolognese, A.M.; DosSantos, M.F.; Souza, M.M. Human dental follicle cells express embryonic, mesenchymal and neural stem cells markers. Arch. Oral Biol. 2017, 73, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulos, I.; Brizuela, C.; Khoury, M. Gingival mesenchymal stem cells outperform haploidentical dental pulp-derived mesenchymal stem cells in proliferation rate, migration ability, and angiogenic potential. Cell Transplant. 2018, 27, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Andrukhov, C.B.; Blufstein, A.; Rausch-Fan, X. Immunomodulatory properties of dental tissue-derived mesenchymal stem cells: Implication in disease and tissue regeneration. World J. Stem Cells 2019, 11, 604. [Google Scholar] [CrossRef]
- Misawa, M.Y.O.; Silverio Ruiz, K.G.; Nociti, F.H., Jr.; Albiero, M.L.; Saito, M.T.; Nóbrega Stipp, R.; Condino-Neto, A.; Holzhausen, M.; Palombo, H.; Villar, C.C. Periodontal ligament-derived mesenchymal stem cells modulate neutrophil responses via paracrine mechanisms. J. Periodontol. 2019, 90, 747–755. [Google Scholar] [CrossRef]
- Hossein-Khannazer, N.; Hashemi, S.M.; Namaki, S.; Ghanbarian, H.; Sattari, M.; Khojasteh, A. Study of the immunomodulatory effects of osteogenic differentiated human dental pulp stem cells. Life Sci. 2019, 216, 111–118. [Google Scholar] [CrossRef]
- Liu, N.; Zhou, M.; Zhang, Q.; Yong, L.; Zhang, T.; Tian, T.; Ma, Q.; Lin, S.; Zhu, B.; Cai, X. Effect of substrate stiffness on proliferation and differentiation of periodontal ligament stem cells. Cell Prolif. 2018, 51, e12478. [Google Scholar] [CrossRef] [Green Version]
- Proksch, S.; Steinberg, T.; Schulz, S.; Sauerbier, S.; Hellwig, E.; Tomakidi, P. Environmental biomechanics substantiated by defined pillar micropatterns govern behavior of human mesenchymal stem cells. Cell Transplant. 2012, 21, 2455–2469. [Google Scholar] [CrossRef] [PubMed]
- Proksch, S.; Steinberg, T.; Stampf, S.; Schwarz, U.; Hellwig, E.; Tomakidi, P. Crosstalk on cell behavior in interactive cocultures of hMSCs with various oral cell types. Tissue Eng. A 2012, 18, 2601–2610. [Google Scholar] [CrossRef] [PubMed]
- Proksch, S.; Steinberg, T.; Vach, K.; Hellwig, E.; Tomakidi, P. Shaping oral cell plasticity to osteogenic differentiation by human mesenchymal stem cell coculture. Cell Tissue Res. 2014, 356, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Proksch, S.; Bittermann, G.; Vach, K.; Nitschke, R.; Tomakidi, P.; Hellwig, E. hMSC-Derived VEGF Release Triggers the Chemoattraction of Alveolar Osteoblasts. Stem Cells 2015, 33, 3114–3124. [Google Scholar] [CrossRef] [Green Version]
- Sumi, K.; Abe, T.; Kunimatsu, R.; Oki, N.; Tsuka, Y.; Awada, T.; Nakajima, K.; Ando, K.; Tanimoto, K. The effect of mesenchymal stem cells on chemotaxis of osteoclast precursor cells. J. Oral Sci. 2018, 60, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Chen, H.; Zhao, X.; Chen, Z.; Zhang, P.; Tian, Y.; Wang, Y.; Ding, T.; Wang, L.; Shen, Y. Advances in mesenchymal stem cell conditioned medium-mediated periodontal tissue regeneration. J. Transl. Med. 2021, 19, 456. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, X.; Zhou, H.; Zhang, C.; Wang, Y.; Huang, J.; Liu, M.; Yang, P.; Song, A. Enhancement of periodontal tissue regeneration by conditioned media from gingiva-derived or periodontal ligament-derived mesenchymal stem cells: A comparative study in rats. Stem Cell Res. Ther. 2020, 11, 42. [Google Scholar] [CrossRef]
- Nagata, M.; Iwasaki, K.; Akazawa, K.; Komaki, M.; Yokoyama, N.; Izumi, Y.; Morita, I. Conditioned medium from periodontal ligament stem cells enhances periodontal regeneration. Tissue Eng. A 2017, 23, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, W.; Kapila, Y.; Lotz, J.; Kapila, S. Multiple differentiation capacity of STRO-1+/CD146+ PDL mesenchymal progenitor cells. Stem Cells Dev. 2009, 18, 487–496. [Google Scholar] [CrossRef]
- Proksch, S.; Kirsch, K.; Vach, K.; Hellwig, E.; Tomakidi, P. Comparative differentiation analysis of distinct oral tissue-derived cells in response to osteogenic stimulation. Clin. Oral Investig. 2019, 23, 1077–1089. [Google Scholar] [CrossRef]
- Gottwald, E.; Giselbrecht, S.; Augspurger, C.; Lahni, B.; Dambrowsky, N.; Truckenmüller, R.; Piotter, V.; Gietzelt, T.; Wendt, O.; Pfleging, W. A chip-based platform for the in vitro generation of tissues in three-dimensional organization. Lab Chip 2007, 7, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Altmann, B.; Steinberg, T.; Giselbrecht, S.; Gottwald, E.; Tomakidi, P.; Bachle-Haas, M.; Kohal, R.J. Promotion of osteoblast differentiation in 3D biomaterial micro-chip arrays comprising fibronectin-coated poly(methyl methacrylate) polycarbonate. Biomaterials 2011, 32, 8947–8956. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, J.-E.; Seo, H.-J.; Jang, J.-H. Design of fibronectin type III domains fused to an elastin-like polypeptide for the osteogenic differentiation of human mesenchymal stem cells. Acta Biochim. Biophys. Sin. 2019, 51, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Escoda-Francolí, J.; Sánchez-Garcés, M.Á.; Gimeno-Sandig, Á.; Muñoz-Guzón, F.; Barbany-Cairó, J.R.; Badiella-Busquets, L.; Gay-Escoda, C. Guided bone regeneration using beta-tricalcium phosphate with and without fibronectin—An experimental study in rats. Clin. Oral Implants Res. 2018, 29, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; You, X.; Zhang, L.; Zhang, C.; Zou, W. Mechanical regulation of bone remodeling. Bone Res. 2022, 10, 16. [Google Scholar] [CrossRef]
- Wang, H.; Du, T.; Li, R.; Main, R.P.; Yang, H. Interactive effects of various loading parameters on the fluid dynamics within the lacunar-canalicular system for a single osteocyte. Bone 2022, 158, 116367. [Google Scholar] [CrossRef]
- Altmann, B.; Löchner, A.; Swain, M.; Kohal, R.-J.; Giselbrecht, S.; Gottwald, E.; Steinberg, T.; Tomakidi, P. Differences in morphogenesis of 3D cultured primary human osteoblasts under static and microfluidic growth conditions. Biomaterials 2014, 35, 3208–3219. [Google Scholar] [CrossRef]
- Radović, K.; Brković, B.; Roganović, J.; Ilić, J.; Milić Lemić, A.; Jovanović, B. Salivary VEGF and post-extraction wound healing in type 2 diabetic immediate denture wearers. Acta Odontol. Scand. 2022, 80, 9–14. [Google Scholar] [CrossRef]
- Müller, K.; Engesser, R.; Metzger, S.; Schulz, S.; Kämpf, M.M.; Busacker, M.; Steinberg, T.; Tomakidi, P.; Ehrbar, M.; Nagy, F. A red/far-red light-responsive bi-stable toggle switch to control gene expression in mammalian cells. Nucleic Acids Res. 2013, 41, e77. [Google Scholar] [CrossRef]
- Müller, K.; Engesser, R.; Schulz, S.; Steinberg, T.; Tomakidi, P.; Weber, C.C.; Ulm, R.; Timmer, J.; Zurbriggen, M.D.; Weber, W. Multi-chromatic control of mammalian gene expression and signaling. Nucleic Acids Res. 2013, 41, e124. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Yang, G.; Chen, M.; Wang, C.; He, L.; Xiang, L.; Chen, D.; Ling, J.; Mao, J.J. Lhx8 mediated Wnt and TGFβ pathways in tooth development and regeneration. Biomaterials 2015, 63, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-Y.; Choung, P.-H. USP1 inhibitor ML323 enhances osteogenic potential of human dental pulp stem cells. Biochem. Biophys. Res. Commun. 2020, 530, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Peng, Y.; Kanda, R.; Umeda, M.; Ishikawa, I. Stem Cell Transplantation and Cell-Free Treatment for Periodontal Regeneration. Int. J. Mol. Sci. 2022, 23, 1011. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinberg, T.; Dieterle, M.P.; Tomakidi, P. Molecular Research on Oral Diseases and Related Biomaterials: A Journey from Oral Cell Models to Advanced Regenerative Perspectives. Int. J. Mol. Sci. 2022, 23, 5288. https://doi.org/10.3390/ijms23095288
Steinberg T, Dieterle MP, Tomakidi P. Molecular Research on Oral Diseases and Related Biomaterials: A Journey from Oral Cell Models to Advanced Regenerative Perspectives. International Journal of Molecular Sciences. 2022; 23(9):5288. https://doi.org/10.3390/ijms23095288
Chicago/Turabian StyleSteinberg, Thorsten, Martin Philipp Dieterle, and Pascal Tomakidi. 2022. "Molecular Research on Oral Diseases and Related Biomaterials: A Journey from Oral Cell Models to Advanced Regenerative Perspectives" International Journal of Molecular Sciences 23, no. 9: 5288. https://doi.org/10.3390/ijms23095288
APA StyleSteinberg, T., Dieterle, M. P., & Tomakidi, P. (2022). Molecular Research on Oral Diseases and Related Biomaterials: A Journey from Oral Cell Models to Advanced Regenerative Perspectives. International Journal of Molecular Sciences, 23(9), 5288. https://doi.org/10.3390/ijms23095288