
Intraperitoneally Delivered Umbilical Cord Lining Mesenchymal Stromal Cells Improve Survival and Kidney Function in Murine Lupus via Myeloid Pathway Targeting

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. CL-MSCs Were Able to Improve Mice Survival and Reduce Disease Activity through IP Administration
2.2. Kidney Function Was Improved by CL-MSCs Treatment
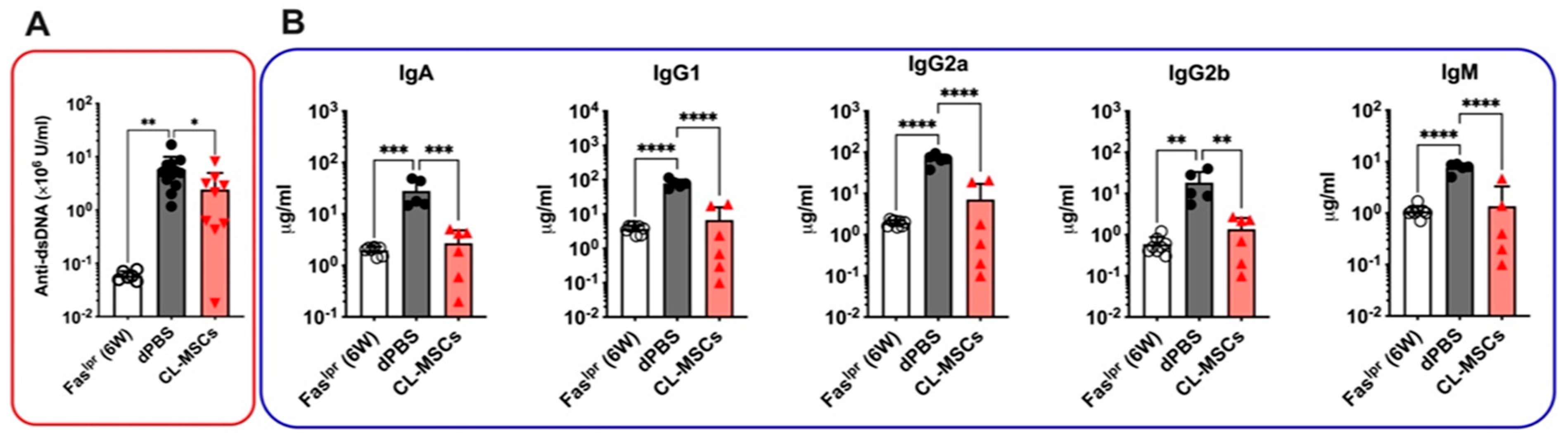
2.3. Both Anti-dsDNA and Isotype Antibodies Were Reduced by CL-MSCs Treatment
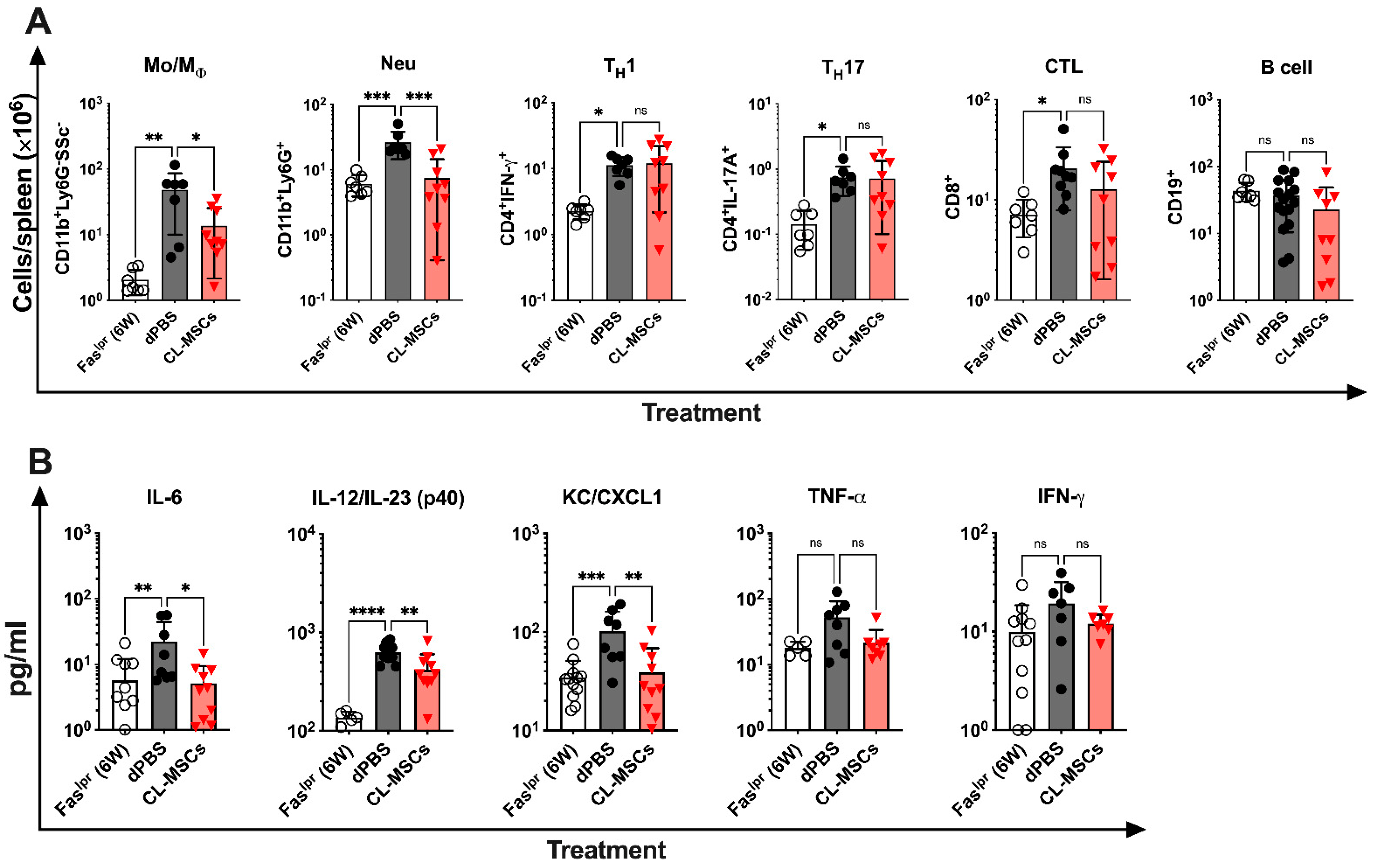
2.4. CL-MSCs Mediated Immunomodulation by Targeting Pro-Inflammatory Cytokine Secreting Monocytes/Macrophages and Neutrophils
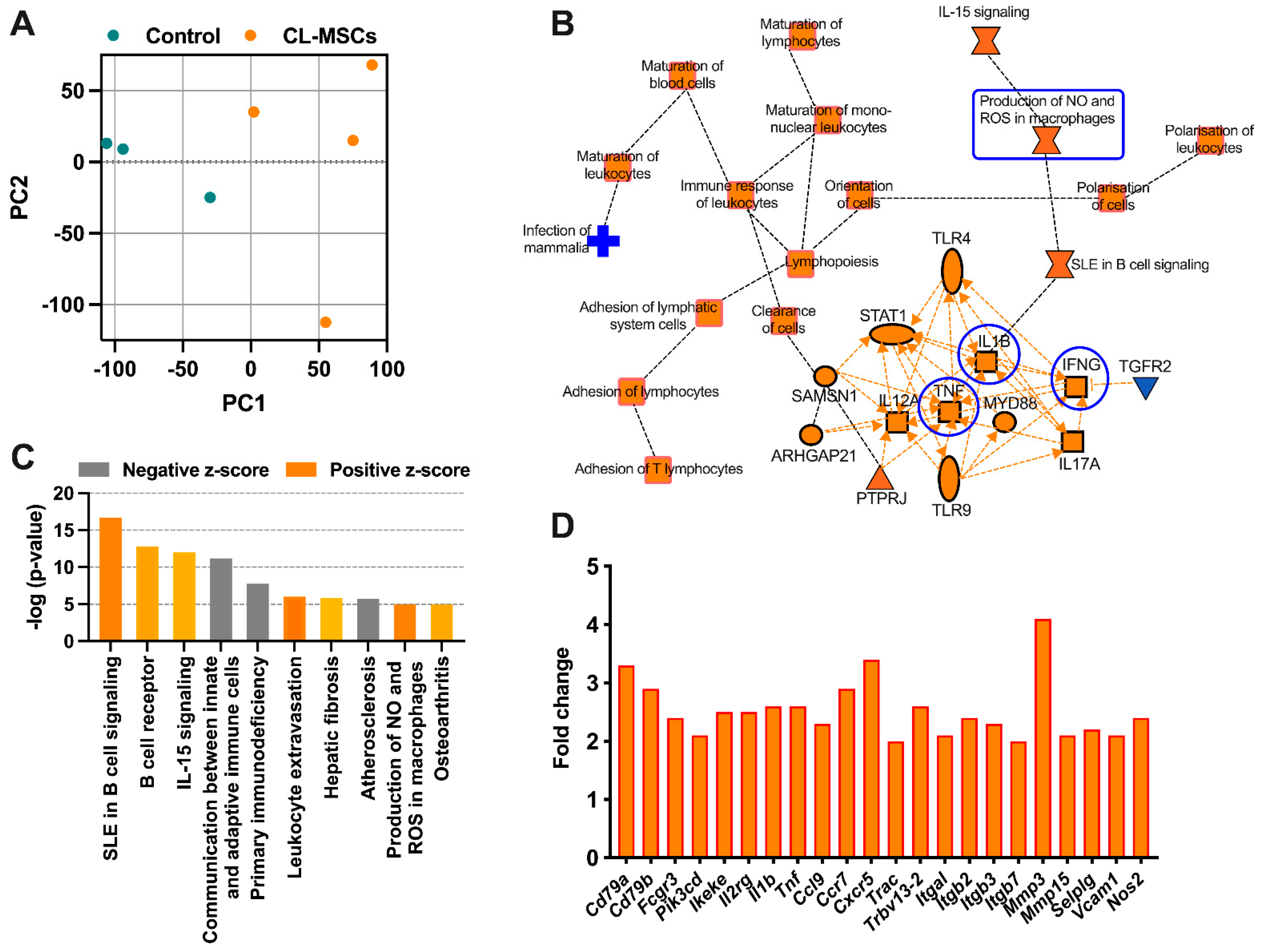
2.5. RNA Sequencing Confirmed the Immunomodulatory Effect of CL-MSCs in Murine SLE
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Mice Treatment
4.3. NIH Lupus Nephritis (LN) Activity and Chronicity Indices
4.4. Renal Lymphocyte Infiltration
4.5. Albumin-to-Creatinine Ratio (ACR) Measurement
4.6. Renal Complement C3 Deposition Staining
4.7. Anti-dsDNA Antibody Analysis
4.8. Flow Cytometric Assay
4.9. Luminex Assay
4.10. RNA Sequencing (RNA-Seq) and Data Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nehar-Belaid, D.; Hong, S.; Marches, R.; Chen, G.; Bolisetty, M.; Baisch, J.; Walters, L.; Punaro, M.; Rossi, R.J.; Chung, C.-H.; et al. Mapping systemic lupus erythematosus heterogeneity at the single-cell level. Nat. Immunol. 2020, 21, 1094–1106. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, R.; Hong, S.; Cantarel, B.; Baldwin, N.; Baisch, J.; Edens, M.; Cepika, A.-M.; Acs, P.; Turner, J.; Anguiano, E.; et al. Personalized Immunomonitoring Uncovers Molecular Networks that Stratify Lupus Patients. Cell 2016, 165, 551–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting Neutrophils Are Major Inducers of Type I IFN Production in Pediatric Systemic Lupus Erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caielli, S.; Veiga, D.T.; Balasubramanian, P.; Athale, S.; Domic, B.; Murat, E.; Banchereau, R.; Xu, Z.; Chandra, M.; Chung, C.-H.; et al. A CD4+ T cell population expanded in lupus blood provides B cell help through interleukin-10 and succinate. Nat. Med. 2019, 25, 75–81. [Google Scholar] [CrossRef]
- Caielli, S.; Athale, S.; Domic, B.; Murat, E.; Chandra, M.; Banchereau, R.; Baisch, J.; Phelps, K.; Clayton, S.; Gong, M.; et al. Oxidized mitochondrial nucleoids released by neutrophils drive type I interferon production in human lupus. J. Exp. Med. 2016, 213, 697–713. [Google Scholar] [CrossRef] [Green Version]
- Chatham, W.W.; Kimberly, R.P. Treatment of lupus with corticosteroids. Lupus 2001, 10, 140–147. [Google Scholar] [CrossRef]
- Sattwika, P.D.; Mustafa, R.; Paramaiswari, A.; Herningtyas, E.H. Stem cells for lupus nephritis: A concise review of current knowledge. Lupus 2018, 27, 1881–1897. [Google Scholar] [CrossRef]
- Bultink, I.E.M.; de Vries, F.; van Vollenhoven, R.F.; Lalmohamed, A. Mortality, causes of death and influence of medication use in patients with systemic lupus erythematosus vs. matched controls. Rheumatology 2021, 60, 207–216. [Google Scholar] [CrossRef]
- Lim, C.C.; Liu, P.Y.; Tan, H.Z.; Lee, P.; Chin, Y.M.; Mok, I.Y.; Chan, C.M.; Choo, J. Severe infections in patients with lupus nephritis treated with immunosuppressants: A retrospective cohort study. Nephrology 2017, 22, 478–484. [Google Scholar] [CrossRef]
- Kanatlı, U.; Ataoğlu, B.; Özer, M.; Şenköylü, A.; Cetinkaya, M. Kyphoplasty for Intractable Pain Due to Glucocorticosteroid-induced Osteoporotic Vertebra Fracture of a 9-Year-Old Patient With Systemic Lupus Erythematosus: 8-Year Follow-up. J. Pediatr. Orthop. 2015, 35, e55–e59. [Google Scholar] [CrossRef]
- Nauta, A.J.; Fibbe, W.E. Immunomodulatory properties of mesenchymal stromal cells. Blood 2007, 110, 3499–3506. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Chen, W.; Huang, S.; Yao, G.; Tang, X.; Sun, L. Mesenchymal stem cells prevent overwhelming inflammation and reduce infection severity via recruiting CXCR3 + regulatory T cells. Clin. Transl. Immunol. 2020, 9, e1181. [Google Scholar] [CrossRef]
- Sun, W.; Yan, S.; Yang, C.; Yang, J.; Wang, H.; Li, C.; Zhang, L.; Zhao, L.; Zhang, J.; Cheng, M.; et al. Mesenchymal Stem Cells-derived Exosomes Ameliorate Lupus by Inducing M2 Macrophage Polarization and Regulatory T Cell Expansion in MRL/lpr Mice. Immunol. Investig. 2022, 51, 1785–1803. [Google Scholar] [CrossRef]
- Ren, G.; Zhang, L.; Zhao, X.; Xu, G.; Zhang, Y.; Roberts, A.I.; Zhao, R.C.; Shi, Y. Mesenchymal Stem Cell-Mediated Immunosuppression Occurs via Concerted Action of Chemokines and Nitric Oxide. Cell Stem Cell 2008, 2, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Walsh, K.P.; Brady, M.T.; Finlay, C.M.; Boon, L.; Mills, K.H. Infection with a helminth parasite attenuates autoimmunity through TGF-beta-mediated suppression of Th17 and Th1 responses. J. Immunol. 2009, 183, 1577–1586. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Di Nicola, M.; Carlo-Stella, C.; Magni, M.; Milanesi, M.; Longoni, P.D.; Matteucci, P.; Grisanti, S.; Gianni, A.M. Human bone marrow stromal cells suppress T-lymphocyte proliferation induced by cellular or nonspecific mitogenic stimuli. Blood 2002, 99, 3838–3843. [Google Scholar] [CrossRef]
- Meisel, R.; Zibert, A.; Laryea, M.; Göbel, U.; Däubener, W.; Dilloo, D. Human bone marrow stromal cells inhibit allogeneic T-cell responses by indoleamine 2,3-dioxygenase–mediated tryptophan degradation. Blood 2004, 103, 4619–4621. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Chen, X.; Huang, Y.; Li, W.; Li, J.; Cao, K.; Cao, G.; Zhang, L.; Li, F.; Roberts, A.I.; et al. Phylogenetic distinction of iNOS and IDO function in mesenchymal stem cell-mediated immunosuppression in mammalian species. Cell Death Differ. 2014, 21, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Németh, K.; Leelahavanichkul, A.; Yuen, P.S.; Mayer, B.; Parmelee, A.; Doi, K.; Robey, P.G.; Leelahavanichkul, K.; Koller, B.H.; Brown, J.M.; et al. Bone marrow stromal cells attenuate sepsis via prostaglandin E2–dependent reprogramming of host macrophages to increase their interleukin-10 production. Nat. Med. 2009, 15, 42–49. [Google Scholar] [CrossRef]
- Oh, J.Y.; Lee, R.H.; Yu, J.M.; Ko, J.H.; Lee, H.J.; Ko, A.Y.; Roddy, G.W.; Prockop, D.J. Intravenous Mesenchymal Stem Cells Prevented Rejection of Allogeneic Corneal Transplants by Aborting the Early Inflammatory Response. Mol. Ther. J. Am. Soc. Gene Ther. 2012, 20, 2143–2152. [Google Scholar] [CrossRef] [Green Version]
- Mougiakakos, D.; Jitschin, R.; Johansson, C.C.; Okita, R.; Kiessling, R.; Le Blanc, K.; Fos, J.; Pabst, T.; Petkovic, V.; Ratschiller, D.; et al. The impact of inflammatory licensing on heme oxygenase-1–mediated induction of regulatory T cells by human mesenchymal stem cells. Blood 2011, 117, 4826–4835. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.; Vanella, L.; Bellner, L.; Schwartzman, M.L.; Abraham, N.G. Epoxyeicosatrienoic acids and heme oxygenase-1 interaction attenuates diabetes and metabolic syndrome complications. Prostaglandins Other Lipid Mediat. 2012, 97, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Gieseke, F.; Böhringer, J.; Bussolari, R.; Dominici, M.; Handgretinger, R.; Müller, I. Human multipotent mesenchymal stromal cells use galectin-1 to inhibit immune effector cells. Blood 2010, 116, 3770–3779. [Google Scholar] [CrossRef]
- Yang, H.M.; Sung, J.H.; Choi, Y.S.; Lee, H.J.; Roh, C.R.; Kim, J.; Shin, M.; Song, S.; Kwon, C.H.; Joh, J.W.; et al. Enhancement of the immunosuppressive effect of human adipose tissue-derived mesenchymal stromal cells through HLA-G1 expression. J. Immunol. Res. 2012, 14, 70–79. [Google Scholar] [CrossRef]
- Bukulmez, H.; Kumar, G. Clinical Use of Mesenchymal Stem Cells in Treatment of Systemic Lupus Erythematosus. In Lupus—Need to Know; IntechOpen: London, UK, 2021. [Google Scholar]
- Zhou, Y.; Hu, Q.; Chen, F.; Zhang, J.; Guo, J.; Wang, H.; Gu, J.; Ma, L.; Ho, G. Human umbilical cord matrix-derived stem cells exert trophic effects on β-cell survival in diabetic rats and isolated islets. Dis. Model. Mech. 2015, 8, 1625–1633. [Google Scholar] [CrossRef] [Green Version]
- Hayward, C.J.; Fradette, J.; Martin, P.M.; Guignard, R.; Germain, L.; Auger, F.A. Using human umbilical cord cells for tissue engineering: A comparison with skin cells; research in biological diversity. Differentiation 2014, 87, 172–181. [Google Scholar] [CrossRef]
- Nagamura-Inoue, H.H.T. Umbilical cord-derived mesenchymal stem cells: Their advantages and potential clinical utility. World J. Stem Cells 2014, 6, 195–202. [Google Scholar] [CrossRef]
- Batsali, A.K.; Kastrinaki, M.-C.; Papadaki, H.A.; Pontikoglou, C. Mesenchymal Stem Cells Derived from Wharton’s Jelly of the Umbilical Cord: Biological Properties and Emerging Clinical Applications. Curr. Stem Cell Res. Ther. 2013, 8, 144–155. [Google Scholar] [CrossRef]
- Lim, I.J.; Phan, T.T. Epithelial and Mesenchymal Stem Cells from the Umbilical Cord Lining Membrane. Cell Transplant. 2014, 23, 497–503. [Google Scholar] [CrossRef]
- Stubbendorff, M.; Deuse, T.; Hua, X.; Phan, T.T.; Bieback, K.; Atkinson, K.; Eiermann, T.H.; Velden, J.; Schröder, C.; Reichenspurner, H.; et al. Immunological Properties of Extraembryonic Human Mesenchymal Stromal Cells Derived from Gestational Tissue. Stem Cells Dev. 2013, 22, 2619–2629. [Google Scholar] [CrossRef] [Green Version]
- Deuse, T.; Stubbendorff, M.; Tang-Quan, K.; Phillips, N.; Kay, M.A.; Eiermann, T.; Phan, T.T.; Volk, H.-D.; Reichenspurner, H.; Robbins, R.C.; et al. Immunogenicity and Immunomodulatory Properties of Umbilical Cord Lining Mesenchymal Stem Cells. Cell Transplant. 2011, 20, 655–667. [Google Scholar] [CrossRef] [Green Version]
- Nam, Y.; Jung, S.M.; Rim, Y.A.; Jung, H.; Lee, K.; Park, N.; Kim, J.; Jang, Y.; Park, Y.-B.; Park, S.-H.; et al. Intraperitoneal infusion of mesenchymal stem cell attenuates severity of collagen antibody induced arthritis. PLoS ONE 2018, 13, e0198740. [Google Scholar] [CrossRef]
- Wiafe, B.; Kadam, R.; Metcalfe, P.D. Intraperitoneal administration of mesenchymal stem cells is effective at mitigating detrusor deterioration after pBOO. Am. J. Physiol. Physiol. 2020, 318, F549–F556. [Google Scholar] [CrossRef]
- Adam, R.M. Recent Insights into the Cell Biology of Bladder Smooth Muscle. Nephron Exp. Nephrol. 2006, 102, e1–e7. [Google Scholar] [CrossRef]
- Al-Saikan, B.; Ding, J.; Tredget, E.E.; Metcalfe, P. Benefits of mesenchymal stem cells after partial bladder outlet obstruction. Can. Urol. Assoc. J. 2016, 10, E1–E6. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.W.; Kwon, M.; Choi, J.C.; Shin, J.W.; Park, I.W.; Choi, B.W.; Kim, J.Y. Familial Occurrence of Pulmonary Embolism after Intravenous, Adipose Tissue-Derived Stem Cell Therapy. Yonsei Med. J. 2013, 54, 1293–1296. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, F.; Ebtekar, M.; Soleimani, M.; Soudi, S.; Hashemi, S.M. Comparison of in vivo immunomodulatory effects of intravenous and intraperitoneal administration of adipose-tissue mesenchymal stem cells in experimental autoimmune encephalomyelitis (EAE). Int. Immunopharmacol. 2013, 17, 608–616. [Google Scholar] [CrossRef]
- Wang, M.; Liang, C.; Hu, H.; Zhou, L.; Xu, B.; Wang, X.; Han, Y.; Nie, Y.; Jia, S.; Liang, J.; et al. Intraperitoneal injection (IP), Intravenous injection (IV) or anal injection (AI)? Best way for mesenchymal stem cells transplantation for colitis. Sci. Rep. 2016, 6, 30696. [Google Scholar] [CrossRef]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-inflammatory protein TSG-6 secreted by activated MSCs attenuates zymosan-induced mouse peritonitis by decreasing TLR2/NF-κB signaling in resident macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef]
- Müller-Ehmsen, J. The problem is obvious, the solution is not: Numbers do matter in cardiac cell therapy! Cardiovasc. Res. 2012, 96, 208–209. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Ng, C.T.; Guo, D.; Lim, F.; Tan, J.C.; Law, A.; Goh, L.H.; Poon, Z.Y.; Cheung, A.; Kong, S.L.; et al. CXCL5 administration dampens inflammation and improves survival in murine lupus via myeloid and neutrophil pathways. Arthritis Rheumatol. 2022. [Google Scholar] [CrossRef]
- Li, A.; Guo, F.; Pan, Q.; Chen, S.; Chen, J.; Liu, H.-F.; Pan, Q. Mesenchymal Stem Cell Therapy: Hope for Patients With Systemic Lupus Erythematosus. Front. Immunol. 2021, 12, 4062. [Google Scholar] [CrossRef]
- Anton, E. Delayed toxicity of cyclophosphamide in normal mice. Br. J. Exp. Pathol. 1987, 68, 237–249. [Google Scholar]
- Abumaree, M.H.; Abomaray, F.M.; Alshabibi, M.A.; AlAskar, A.S.; Kalionis, B. Immunomodulatory properties of human placental mesenchymal stem/stromal cells. Placenta 2017, 59, 87–95. [Google Scholar] [CrossRef]
- Phan, T.T.; Freed, B.M. A Method of Transporting Mesenchymal Stem Cells by Means of a Transporting Solution and a Method of Administering Stem Cells to Wounds. WO2019199229A1, 17 October 2019. [Google Scholar]
- Kita, K.; Gauglitz, G.G.; Phan, T.T.; Herndon, D.N.; Jeschke, M.G. Isolation and Characterization of Mesenchymal Stem Cells from the Sub-Amniotic Human Umbilical Cord Lining Membrane. Stem Cells Dev. 2010, 19, 491–502. [Google Scholar] [CrossRef]
- Ma, X.; Che, N.; Gu, Z.; Huang, J.; Wang, D.; Liang, J.; Hou, Y.; Gilkeson, G.; Lu, L.; Sun, L. Allogenic Mesenchymal Stem Cell Transplantation Ameliorates Nephritis in Lupus Mice via Inhibition of B-Cell Activation. Cell Transplant. 2013, 22, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yang, J.; Li, X.; Ma, W.; Zou, H. Bone marrow-derived mesenchymal stem cells inhibit T follicular helper cell in lupus-prone mice. Lupus 2018, 27, 49–59. [Google Scholar] [CrossRef]
- Bajema, I.M.; Wilhelmus, S.; Alpers, C.E.; Bruijn, J.A.; Colvin, R.B.; Cook, H.T.; D’Agati, V.D.; Ferrario, F.; Haas, M.; Jennette, J.C.; et al. Revision of the International Society of Nephrology/Renal Pathology Society classification for lupus nephritis: Clarification of definitions and modified National Institutes of Health activity and chronicity indices. Kidney Int. 2018, 93, 789–796. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chua, A.W.C.; Guo, D.; Tan, J.C.; Lim, F.T.W.; Ong, C.T.; Masilamani, J.; Lim, T.K.H.; Hwang, W.Y.K.; Lim, I.J.; Chen, J.; et al. Intraperitoneally Delivered Umbilical Cord Lining Mesenchymal Stromal Cells Improve Survival and Kidney Function in Murine Lupus via Myeloid Pathway Targeting. Int. J. Mol. Sci. 2023, 24, 365. https://doi.org/10.3390/ijms24010365
Chua AWC, Guo D, Tan JC, Lim FTW, Ong CT, Masilamani J, Lim TKH, Hwang WYK, Lim IJ, Chen J, et al. Intraperitoneally Delivered Umbilical Cord Lining Mesenchymal Stromal Cells Improve Survival and Kidney Function in Murine Lupus via Myeloid Pathway Targeting. International Journal of Molecular Sciences. 2023; 24(1):365. https://doi.org/10.3390/ijms24010365
Chicago/Turabian StyleChua, Alvin Wen Choong, Dianyang Guo, Jia Chi Tan, Frances Ting Wei Lim, Chee Tian Ong, Jeyakumar Masilamani, Tony Kiat Hon Lim, William Ying Khee Hwang, Ivor Jiun Lim, Jinmiao Chen, and et al. 2023. "Intraperitoneally Delivered Umbilical Cord Lining Mesenchymal Stromal Cells Improve Survival and Kidney Function in Murine Lupus via Myeloid Pathway Targeting" International Journal of Molecular Sciences 24, no. 1: 365. https://doi.org/10.3390/ijms24010365
APA StyleChua, A. W. C., Guo, D., Tan, J. C., Lim, F. T. W., Ong, C. T., Masilamani, J., Lim, T. K. H., Hwang, W. Y. K., Lim, I. J., Chen, J., Phan, T. T., & Fan, X. (2023). Intraperitoneally Delivered Umbilical Cord Lining Mesenchymal Stromal Cells Improve Survival and Kidney Function in Murine Lupus via Myeloid Pathway Targeting. International Journal of Molecular Sciences, 24(1), 365. https://doi.org/10.3390/ijms24010365