
The Landscape of Gene Expression during Hyperfilamentous Biofilm Development in Oral Candida albicans Isolated from a Lung Cancer Patient

 , ,
, , 
Abstract
:1. Introduction
2. Results
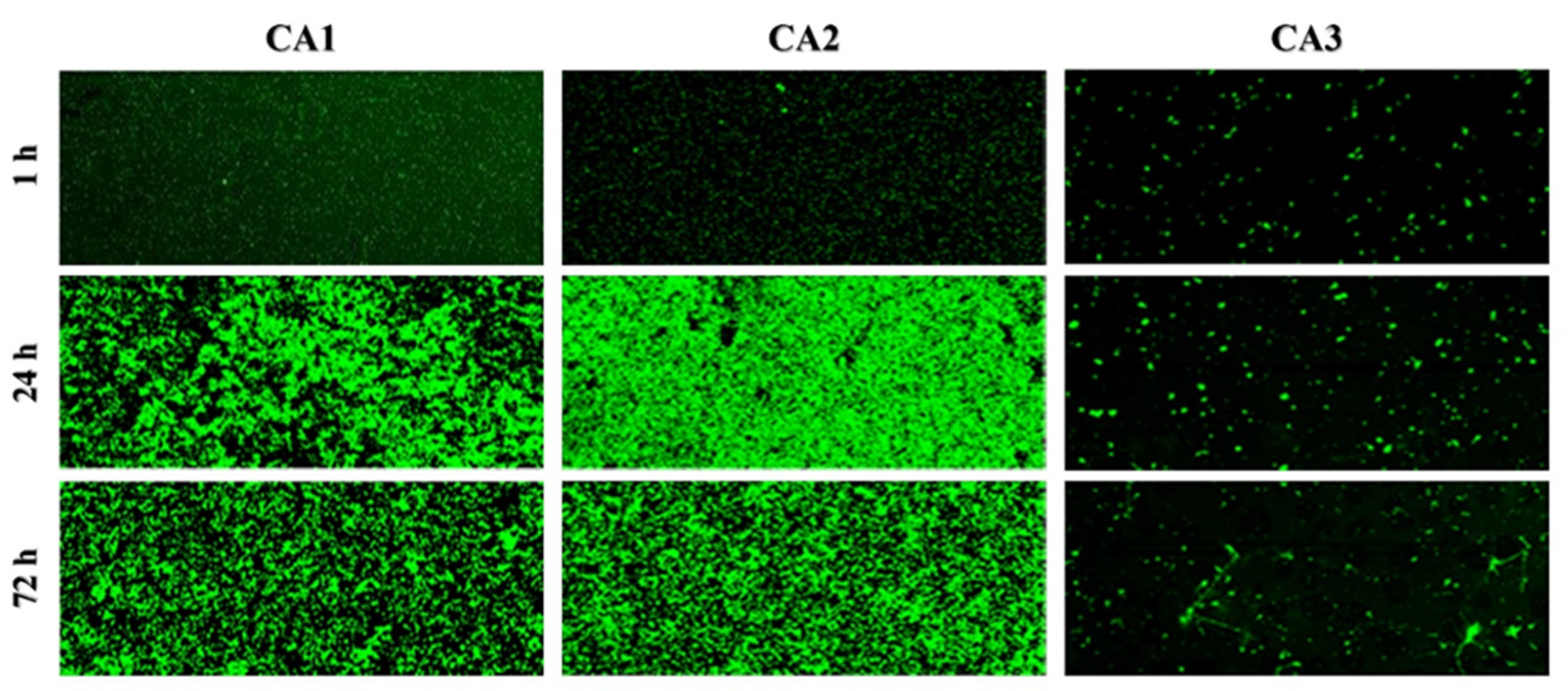
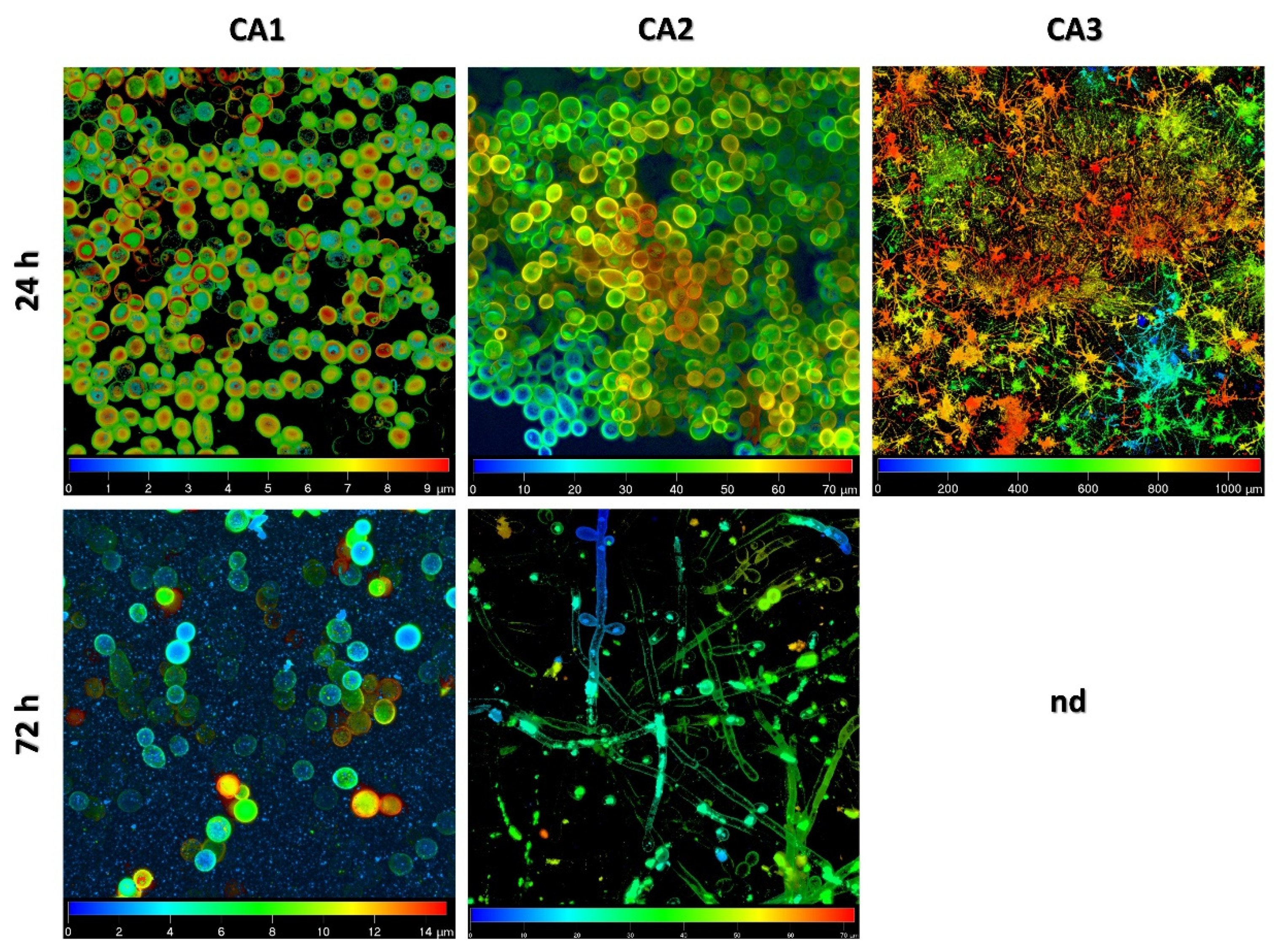
2.1. Biofilm Structure and Development Analysis by Confocal Scanning Laser Microscopy
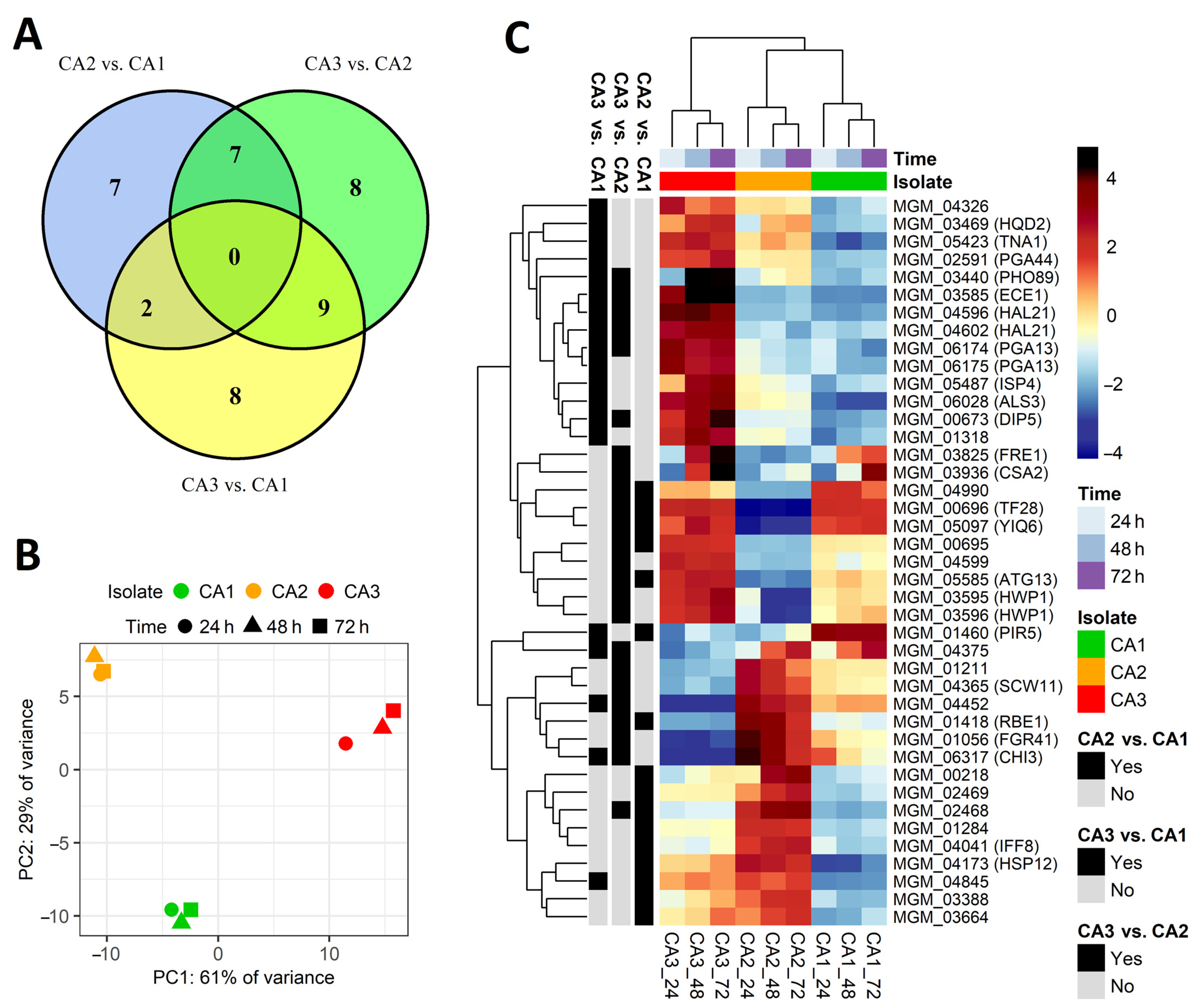
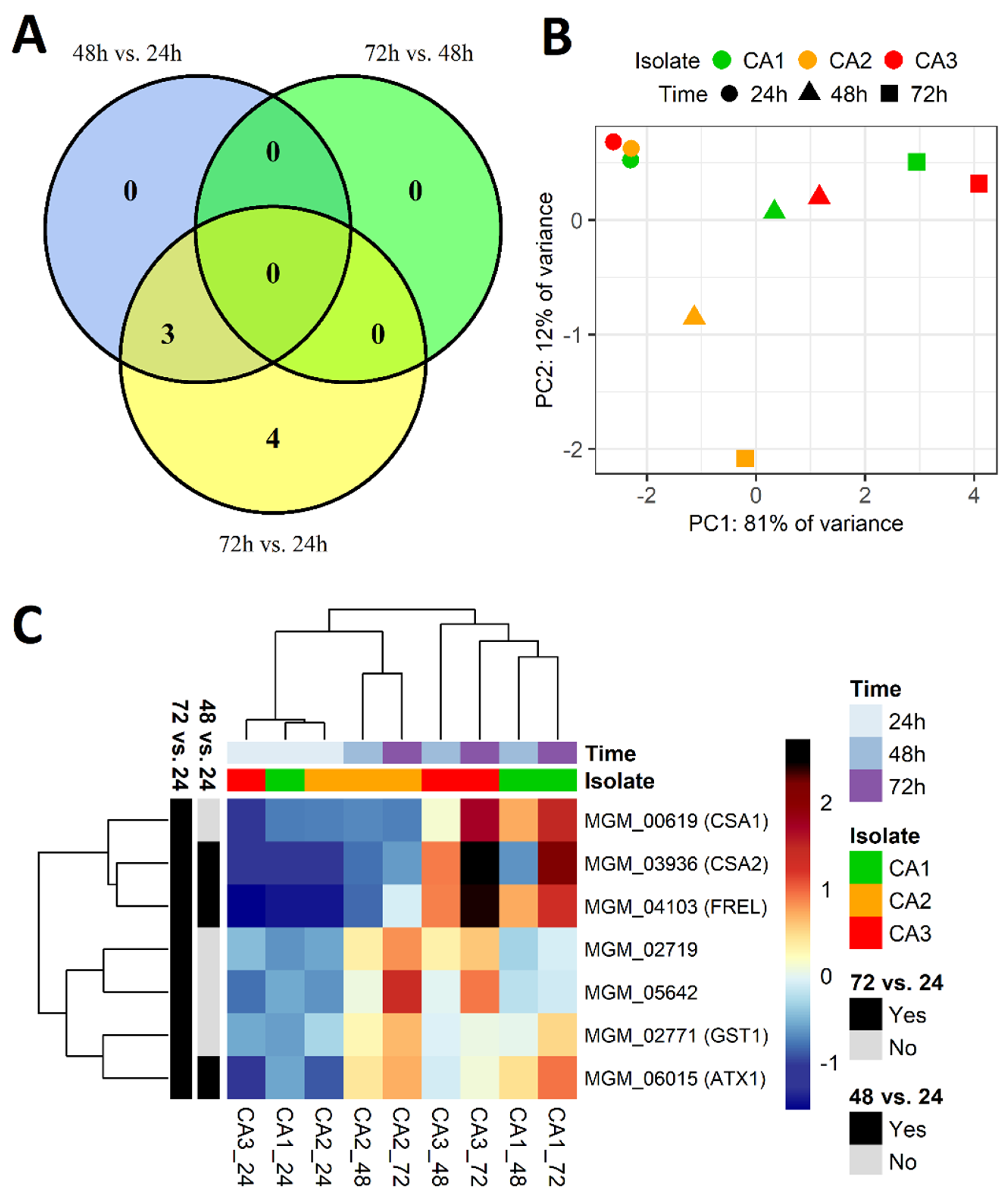
2.2. Differentially Expressed Genes between Candida albicans Isolates
2.3. Differentially Expressed Genes between Incubation Time Points
3. Discussion
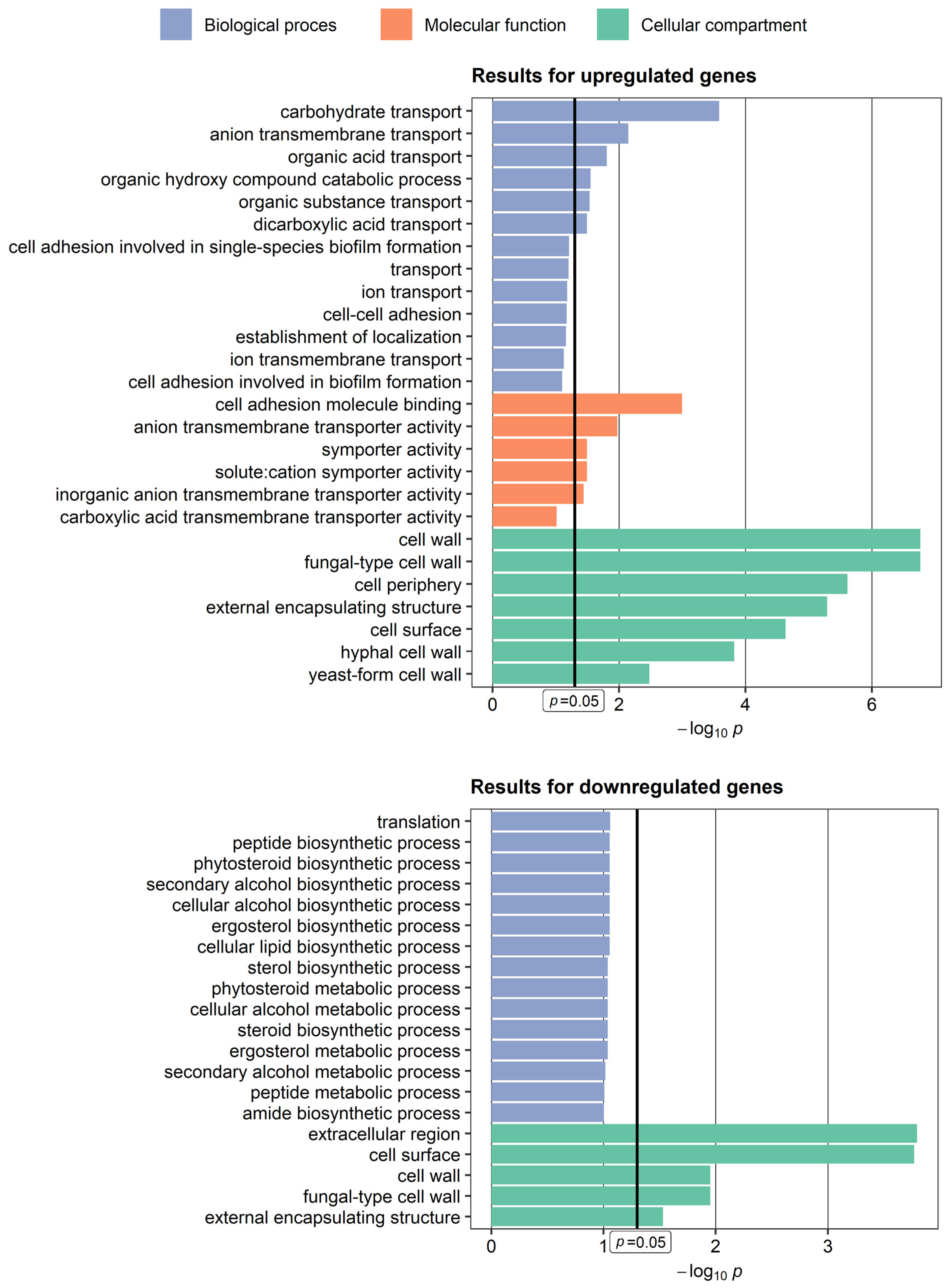
3.1. Genes Upregulated in Candida albicans Isolate Exhibiting Hyperfilamentation
3.2. Genes Downregulated in Candida albicans Isolate Exhibiting Hyperfilamentation
3.3. Iron Metabolism in Candida albicans Is Involved in the Process of Hypheal Morphogenesis
3.4. Limitations of the Study
4. Materials and Methods
4.1. Candida albicans Strains and Culture Conditions
4.2. Confocal Scanning Laser Microscopy Analysis
4.3. Total RNA Preparation
4.4. RNA Sequencing
4.5. Sequencing Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atriwal, T.; Azeem, K.; Husain, F.M.; Hussain, A.; Khan, M.N.; Alajmi, M.F.; Abid, M. Mechanistic understanding of Candida albicans biofilm formation and approaches for its inhibition. Front. Microbiol. 2021, 12, 638609. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.; dos Santos Fontenelle, R.O.; de Brito, E.H.S.; de Morais, S.M. Biofilm of Candida albicans: Formation, regulation and resistance. J. Appl. Microbiol. 2021, 131, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Rao, R.S.; Majumdar, B.; Anil, S. Clinical appearance of oral Candida infection and therapeutic strategies. Front. Microbiol. 2015, 6, 1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, L.J. Medical importance of biofilms in Candida infections. Rev. Iberoam. Micol. 2002, 19, 139–143. [Google Scholar] [PubMed]
- Ramage, G.; Saville, S.P.; Thomas, D.P.; López-Ribot, J.L. Candida biofilms: An update. Eukaryot. Cell 2005, 4, 633–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannanusorn, S.; Ramírez-Zavala, B.; Lünsdorf, H.; Agerberth, B.; Morschhäuser, J.; Römling, U. Characterization of biofilm formation and the role of BCR1 in clinical isolates of Candida parapsilosis. Eukaryot. Cell 2014, 13, 438–451. [Google Scholar] [CrossRef] [Green Version]
- Chong, P.P.; Chin, V.K.; Wong, W.F.; Madhavan, P.; Yong, V.C.; Looi, C.Y. Transcriptomic and genomic approaches for unravelling Candida albicans biofilm formation and drug resistance—An update. Genes 2018, 9, 540. [Google Scholar] [CrossRef] [Green Version]
- Alim, D.; Sircaik, S.; Panwar, S.L. The significance of lipids to biofilm formation in Candida albicans: An emerging perspective. J. Fungi 2018, 4, 140. [Google Scholar] [CrossRef] [Green Version]
- Chudzik-Rząd, B.; Malm, A.; Piersiak, T.; Gawron, A. Quantitative analysis of biofilm formation by oropharyngeal Candida albicans isolates under static conditions by confocal scanning laser microscopy. Curr. Issues Pharm. Med. Sci. 2013, 26, 54–56. [Google Scholar]
- Eix, E.F.; Nett, J.E. How biofilm growth affects Candida-host interactions. Front. Microbiol. 2020, 11, 1437. [Google Scholar] [CrossRef] [PubMed]
- de Barros, P.P.; Rossoni, R.D.; de Souza, C.M.; Scorzoni, L.; Fenley, J.D.C.; Junqueira, J.C. Candida biofilms: An update on developmental mechanisms and therapeutic challenges. Mycopathologia 2020, 185, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Romo, J.A.; Zhang, H.; Cai, H.; Kadosh, D.; Koehler, J.R.; Saville, S.P.; Wang, Y.; Lopez-Ribot, J.L. Global transcriptomic analysis of the Candida albicans response to treatment with a novel inhibitor of filamentation. mSphere 2019, 4, e00620-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthew, W.M.; Walsh, T.J. Candidemia in the cancer patient: Diagnosis, treatment, and future directions. Expert. Rev. Anti. Infect. Ther. 2018, 16, 849–854. [Google Scholar]
- Ruhnke, M.; Cornely, O.A.; Schmidt-Hieber, M.; Alakel, N.; Boell, B.; Buchheidt, D.; Christopeit, M.; Hasenkamp, J.; Jeinz, W.J.; Hentrich, M.; et al. Treatment of invasive fungal diseases in cancer patients—Revised 2019. Recommendations of the Infectious Diseases Working Party (AGIHO) of the German Society of Hematology and Oncology (DGHO). Mycoses 2020, 63, 653–682. [Google Scholar]
- Wall, G.; Lopez-Ribot, J.L. Current antimycotics, new prospects, and future approaches to antifungal therapy. Antibiotics 2020, 9, 445. [Google Scholar] [CrossRef]
- Silva, S.; Rodrigues, C.F.; Araújo, D.; Rodrigues, M.E.; Henriques, M. Candida Species Biofilms’ Antifungal Resistance. J. Fungi 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, P.K.; Chandra, J.; Kuhn, D.M.; Ghannoum, M.A. Mechanism of fluconazole resistance in Candida albicans biofilms: Phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 2003, 71, 4333–4340. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Filler, S.G. Candida albicans Als3, a multifunctional adhesin and invasin. Eukaryot. Cell 2011, 10, 168–173. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Daniels, K.J.; Oh, S.-H.; Green, C.B.; Yeater, K.M.; Soll, D.R.; Hoyer, L.L. Candida albicans Als3p is required for wild-type biofilm formation on silicone elastomer surfaces. Microbiology 2006, 152, 2287–2299. [Google Scholar] [CrossRef] [Green Version]
- Deng, K.; Jiang, W.; Jiang, Y.; Deng, Q.; Cao, J.; Yang, W.; Zhao, X. ALS3 expression as an indicator for Candida albicans biofilm formation and drug resistance. Front. Microbiol. 2021, 12, 655242. [Google Scholar] [CrossRef] [PubMed]
- Short, B.; Delaney, C.; McKloud, E.; Brown, J.L.; Kean, R.; Litherland, G.J.; Williams, C.; Martin, S.L.; MacKay, W.G.; Ramage, G. Investigating the transcriptome of Candida albicans in a dual-species Staphylococcus aureus biofilm model. Front. Cell. Infect. Microbiol. 2021, 11, 791523. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Willems, H.M.; Sansevere, E.A.; Allert, S.; Barker, K.S.; Lowes, D.J.; Dixson, A.C.; Xu, Z.; Miao, J.; De Jarnette, C.; et al. A variant ECE1 allele contributes to reduced pathogenicity of Candida albicans during vulvovaginal candidiasis. PLoS Pathog. 2021, 17, e1009884. [Google Scholar] [CrossRef] [PubMed]
- Engku, N.S.E.A.F.; Ahmad, H.; Ramzi, A.B.; Wahab, A.R.; Kaderi, M.A.; Wan, W.H.A.W.; Dashper, S.; McCullough, M.; Arzmi, M.H. The role of Candida albicans candidalysin ECE1 gene in oral carcinogenesis. J. Oral Pathol. Med. 2020, 49, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Orsi, C.F.; Borghi, E.; Colombari, B.; Neglia, R.G.; Quaglino, D.; Ardizzoni, A.; Morace, G.; Blasi, E. Impact of Candida albicans Hyphal Wall Protein 1 (HWP1) genotype on biofilm production and fungal susceptibility to microglial cells. Microb. Pathog. 2014, 69, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobile, C.J.; Schneider, H.A.; Nett, J.E.; Sheppard, D.C.; Filler, S.G.; Andes, D.R.; Mitchell, A.P. Complementary adhesin function in C. albicans biofilm formation. Curr. Biol. 2008, 18, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nett, J.E.; Lepak, A.J.; Marchillo, K.; Andes, D.R. Time course global gene expression analysis of an in vivo Candida biofilm. J. Infect. Dis. 2009, 200, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Torbensen, R.; Møller, H.D.; Gresham, D.; Alizadeh, S.; Ochmann, D.; Boles, E.; Regenberg, B. Amino acid transporter genes are essential for FLO11-dependent and FLO11-independent biofilm formation and invasive growth in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e41272. [Google Scholar] [CrossRef] [Green Version]
- Murillo, L.A.; Newport, G.; Lan, C.Y.; Habelitz, S.; Dungan, J.; Agabian, N.M. Genome-wide transcription profiling of the early phase of biofilm formation by Candida albicans. Eukaryot. Cell 2005, 4, 1562–1573. [Google Scholar] [CrossRef] [Green Version]
- Röhm, M.; Lindemann, E.; Hiller, E.; Ermert, D.; Lemuth, K.; Trkulja, D.; Sogukpinar, O.; Brunner, H.; Rupp, S.; Urban, C.F.; et al. A family of secreted pathogenesis-related proteins in Candida albicans. Mol. Microbiol. 2013, 87, 132–151. [Google Scholar] [CrossRef]
- Sohn, K.; Urban, C.; Brunner, H.; Rupp, S. EFG1 is a major regulator of cell wall dynamics in Candida albicans as revealed by DNA microarrays. Mol. Microbiol. 2003, 47, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Glazier, V.E. EFG1, everyone’s favorite gene in Candida albicans: A comprehensive literature review. Front. Cell. Infect. Microbiol. 2022, 12, 855229. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, W.L. Candida albicans cell wall proteins. Microbiol. Mol. Biol. Rev. 2008, 72, 495–544. [Google Scholar] [CrossRef] [PubMed]
- Mulhern, S.M.; Logue, M.E.; Butler, G. Candida albicans transcription factor Ace2 regulates metabolism and is required for filamentation in hypoxic conditions. Eukaryot. Cell 2006, 5, 2001–2013. [Google Scholar] [CrossRef] [Green Version]
- Dünkler, A.; Walther, A.; Specht, C.A.; Wendland, J. Candida albicans CHT3 encodes the functional homolog of the Cts1 chitinase of Saccharomyces cerevisiae. Fungal Genet. Biol. 2005, 42, 935–947. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Bourbon-Melo, N.; Sá-Correia, I. The cell wall and the response and tolerance to stresses of biotechnological relevance in yeasts. Front. Microbiol. 2002, 13, 953479. [Google Scholar] [CrossRef]
- Fourie, R.; Kuloyo, O.O.; Mochochoko, B.M.; Albertyn, J.; Pohl, C.H. Iron at the centre of Candida albicans interactions. Front. Cell. Infect. Microbiol. 2018, 8, 185. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Lin, C.N.; Wang, Y.C.; Liu, F.Y.; Chien, Y.W.; Chuang, Y.J.; Lan, C.Y.; Hsieh, W.P.; Chen, B.S. Robustness analysis on interspecies interaction network for iron and glucose competition between Candida albicans and zebrafish during infection. BMC Syst. Biol. 2014, 8, S6. [Google Scholar] [CrossRef] [Green Version]
- Okamoto-Shibayama, K.; Kikuchi, Y.; Kokubu, E.; Ishihara, K. Possible Involvement of surface antigen protein 2 in the morphological transition and biofilm formation of Candida albicans. Med. Mycol. J. 2017, 58, E139–E143. [Google Scholar] [CrossRef] [Green Version]
- Almeida, R.S.; Wilson, D.; Hube, B. Candida albicans iron acquisition within the host. FEMS Yeast Res. 2009, 9, 1000–1012. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, T.; Tóth, Z.; Tóth, R.; Vágvölgyi, C.; Gácser, A. Iron metabolism, pseudohypha production, and biofilm formation through a multicopper oxidase in the human-pathogenic fungus Candida parapsilosis. mSphere 2020, 5, e00227-20. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-Seq experiments with TopHat and cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessau, R.B.; Pipper, C.B. ’’R"-project for statistical computing. Ugeskr. Laeg. 2008, 170, 328–330. [Google Scholar] [PubMed]
- Skrzypek, M.S.; Binkley, J.; Binkley, G.; Miyasato, S.R.; Simison, M.; Sherlock, G. The Candida Genome Database (CGD): Incorporation of assembly 22, systematic identifiers and visualization of high throughput sequencing data. Nucleic Acids Res. 2017, 45, D592–D596. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID 1 | Gene Name 2 | Description 3 | Mean Expression 4 | Log2(Fold Change) | p |
|---|---|---|---|---|---|
| CA2vs. CA1 upregulated genes | |||||
| MGM_04173 | HSP12 | Heat shock protein | 7359.3 | 6.38 | 2.47 × 10-76 |
| MGM_02468 | Uncharacterized | 1306.1 | 6.44 | 1.54 × 10-68 | |
| MGM_01284 | Uncharacterized | 123.7 | 5.83 | 3.42 × 10-35 | |
| MGM_01418 | RBE1 | Pry family cell wall protein | 1370.4 | 5.03 | 2.14 × 10-30 |
| MGM_04041 | IFF8 | Putative GPI-anchored adhesin-like protein | 141.5 | 5.50 | 5.07 × 10-23 |
| MGM_03388 | Uncharacterized | 171.4 | 5.39 | 5.02 × 10-19 | |
| MGM_02469 | Uncharacterized | 110.6 | 6.10 | 5.46 × 10-18 | |
| MGM_04845 | Uncharacterized | 182.6 | 9.96 | 1.25 × 10-14 | |
| MGM_03664 | Uncharacterized | 150.3 | 5.72 | 2.21 × 10-14 | |
| MGM_00218 | Uncharacterized | 220.2 | 5.65 | 8 × 10-7 | |
| CA2 vs. CA1 downregulated genes | |||||
| MGM_00696 | TF28 | Transposon Tf2-8 polyprotein | 6275.0 | −8.75 | 1.73 × 10-219 |
| MGM_05097 | YIQ6 | Uncharacterized transporter YIL166C | 4152.0 | −7.12 | 5.38 × 10-62 |
| MGM_04990 | Uncharacterized | 134.6 | −10.65 | 6.99 × 10-16 | |
| MGM_05585 | ATG13 | Autophagy-related protein 13 | 295.8 | −6.41 | 1.01 × 10-14 |
| MGM_01460 | PIR5 | Cell wall Pir protein | 552.2 | −5.55 | 3.1 × 10-9 |
| MGM_00695 | Uncharacterized | 110.6 | −7.63 | 7.41 × 10-9 | |
| CA3 vs. CA1 upregulated genes | |||||
| MGM_05423 | TNA1 | Putative nicotinic acid transporter | 1693.7 | 6.34 | 8.64 × 10-83 |
| MGM_06028 | ALS3 | Cell wall adhesin | 1232.6 | 8.50 | 2.75 × 10-81 |
| MGM_04596 | HAL21 | 3’-phosphoadenosine 5’-phosphosulfate phosphatase | 542.9 | 9.46 | 7.5 × 10-53 |
| MGM_01318 | Uncharacterized | 9938.0 | 5.99 | 1.55 × 10-51 | |
| MGM_04602 | HAL21 | 3’-phosphoadenosine 5’-phosphosulfate phosphatase | 391.3 | 6.04 | 2.75 × 10-48 |
| MGM_00673 | DIP5 | Dicarboxylic amino acid permease | 13,615.4 | 6.68 | 3.33 × 10-43 |
| MGM_06175 | PGA13 | GPI-anchored cell wall protein | 11,790.7 | 5.44 | 1.02 × 10-41 |
| MGM_03585 | ECE1 | Candidalysin | 910.7 | 10.70 | 1.31 × 10-40 |
| MGM_06174 | PGA13 | GPI-anchored cell wall protein | 11,682.1 | 5.84 | 3.27 × 10-36 |
| MGM_02591 | PGA44 | Putative GPI-anchored protein | 200.6 | 5.71 | 8.22 × 10-32 |
| MGM_04326 | Uncharacterized | 186.4 | 5.23 | 2.78 × 10-16 | |
| MGM_03469 | HQD2 | Catechol 1,2-dioxygenase | 256.3 | 5.26 | 1.46 × 10-15 |
| MGM_04845 | Uncharacterized | 182.6 | 9.12 | 1.62 × 10-12 | |
| MGM_05487 | ISP4 | Sexual differentiation process protein isp4 | 490.5 | 5.82 | 6.45 × 10-10 |
| MGM_03440 | PHO89 | Putative phosphate permease | 1088.7 | 7.31 | 4.2 × 10-7 |
| CA3vs. CA1 downregulated genes | |||||
| MGM_04452 | Uncharacterized | 15,450.6 | −5.13 | 2.03 × 10-57 | |
| MGM_01460 | PIR5 | Cell wall Pir protein | 552.2 | −6.24 | 2.17 × 10-11 |
| MGM_06317 | CHI3 | Chitinase | 11,032.2 | −5.42 | 6.29 × 10-11 |
| MGM_04375 | Uncharacterized | 280.6 | −5.76 | 9.21 × 10-4 | |
| CA3 vs. CA2 upregulated genes | |||||
| MGM_00696 | TF28 | Transposon Tf2-8 polyprotein | 6275.0 | 9.10 | 1.33 × 10-237 |
| MGM_04596 | HAL21 | 3’(2’),5’-bisphosphate nucleotidase 1 | 542.9 | 7.72 | 8.31 × 10-81 |
| MGM_05097 | YIQ6 | Uncharacterized transporter YIL166C | 4152.0 | 7.57 | 2.82 × 10-70 |
| MGM_03585 | ECE1 | Candidalysin | 910.7 | 8.35 | 1.72 × 10-54 |
| MGM_04599 | Uncharacterized | 272.0 | 5.92 | 6.32 × 10-53 | |
| MGM_04602 | HAL21 | 3’-phosphoadenosine 5’-phosphosulfate phosphatase | 391.3 | 6.23 | 6.81 × 10-49 |
| MGM_05585 | ATG13 | Autophagy-related protein 13 | 295.8 | 9.10 | 1.43 × 10-29 |
| MGM_06174 | PGA13 | GPI-anchored cell wall protein | 11,682.1 | 5.17 | 1.57 × 10-28 |
| MGM_00673 | DIP5 | Dicarboxylic amino acid permease | 13,615.4 | 5.15 | 6.04 × 10-26 |
| MGM_00695 | Uncharacterized | 110.6 | 10.57 | 2.59 × 10-16 | |
| MGM_04990 | Uncharacterized | 134.6 | 8.91 | 2.65 × 10-11 | |
| MGM_03596 | HWP1 | Hyphal cell wall protein | 2223.7 | 5.38 | 2.2 × 10-7 |
| MGM_03595 | HWP1 | Hyphal cell wall protein | 6037.4 | 5.31 | 4.81 × 10-7 |
| MGM_03825 | FRE1 | Ferric reductase transmembrane component | 2044.7 | 6.87 | 1.81 × 10-6 |
| MGM_03440 | PHO89 | Putative phosphate permease | 1088.7 | 5.46 | 2.35 × 10-4 |
| CA3 vs. CA2—downregulated genes | |||||
| MGM_04452 | Uncharacterized | 15,450.6 | −7.61 | 1.28 × 10-127 | |
| MGM_01056 | FGR41 | Putative GPI-anchored adhesin-like protein | 7627.7 | −8.12 | 3 × 10-65 |
| MGM_01418 | RBE1 | Pry family cell wall protein | 1370.4 | −6.82 | 1.13 × 10-52 |
| MGM_02468 | Uncharacterized | 1306.1 | −5.06 | 5.69 × 10-46 | |
| MGM_04365 | SCW11 | Cell wall protein | 2554.2 | −5.08 | 6.42 × 10-41 |
| MGM_01211 | Uncharacterized | 1072.9 | −5.15 | 4.19 × 10-32 | |
| MGM_06317 | CHI3 | Chitinase | 11,032.2 | −8.77 | 2.24 × 10-27 |
| MGM_04375 | Uncharacterized | 280.6 | −5.57 | 1.75 × 10-3 | |
| Gene ID 1 | Gene Name 2 | Description 3 | Mean Expression 4 | Log2 (Fold Change) | p |
|---|---|---|---|---|---|
| 48 hvs. 24 h—upregulated genes | |||||
| MGM_04103 | FRE1 | Ferric reductase transmembrane component | 1667.0 | 6.21 | 2.22 × 10-6 |
| MGM_06015 | ATX1 | Putative cytosolic copper metallochaperone | 10,752.6 | 2.20 | 3.13 × 10-2 |
| MGM_03936 | CSA2 | Extracellular heme-binding protein | 873.2 | 5.31 | 3.13 × 10-2 |
| 72 hvs. 24 h—upregulated genes | |||||
| MGM_04103 | FRE1 | Ferric reductase transmembrane component | 1667.0 | 8.00 | 4.11 × 10-12 |
| MGM_03936 | CSA2 | Extracellular heme-binding protein | 873.2 | 9.07 | 4.16 × 10-10 |
| MGM_06015 | ATX1 | Putative cytosolic copper metallochaperone | 10,752.6 | 2.78 | 6.7 × 10-5 |
| MGM_05642 | Uncharacterized | 187.3 | 3.58 | 1.11 × 10-4 | |
| MGM_02771 | GST1 | Putative glutathione S-transferase 1 | 115.0 | 2.39 | 8.73 × 10-3 |
| MGM_02719 | Uncharacterized | 108.2 | 2.96 | 9.69 × 10-3 | |
| MGM_00619 | CSA1 | Cell wall protein 1 | 11,289.7 | 3.79 | 9.69 × 10-3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chudzik-Rząd, B.; Zalewski, D.; Kasela, M.; Sawicki, R.; Szymańska, J.; Bogucka-Kocka, A.; Malm, A. The Landscape of Gene Expression during Hyperfilamentous Biofilm Development in Oral Candida albicans Isolated from a Lung Cancer Patient. Int. J. Mol. Sci. 2023, 24, 368. https://doi.org/10.3390/ijms24010368
Chudzik-Rząd B, Zalewski D, Kasela M, Sawicki R, Szymańska J, Bogucka-Kocka A, Malm A. The Landscape of Gene Expression during Hyperfilamentous Biofilm Development in Oral Candida albicans Isolated from a Lung Cancer Patient. International Journal of Molecular Sciences. 2023; 24(1):368. https://doi.org/10.3390/ijms24010368
Chicago/Turabian StyleChudzik-Rząd, Beata, Daniel Zalewski, Martyna Kasela, Rafał Sawicki, Jolanta Szymańska, Anna Bogucka-Kocka, and Anna Malm. 2023. "The Landscape of Gene Expression during Hyperfilamentous Biofilm Development in Oral Candida albicans Isolated from a Lung Cancer Patient" International Journal of Molecular Sciences 24, no. 1: 368. https://doi.org/10.3390/ijms24010368
APA StyleChudzik-Rząd, B., Zalewski, D., Kasela, M., Sawicki, R., Szymańska, J., Bogucka-Kocka, A., & Malm, A. (2023). The Landscape of Gene Expression during Hyperfilamentous Biofilm Development in Oral Candida albicans Isolated from a Lung Cancer Patient. International Journal of Molecular Sciences, 24(1), 368. https://doi.org/10.3390/ijms24010368