
Topographically Distinguished Microbiome Taxonomy and Stress-Response Genes of Royal Belum Rainforest and Raja Muda Musa Peat Swamp Revealed through Metagenomic Inquisition

 , , ,
, , ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Taxonomic and Functional Comparisons of Tropical Rain Forest and Peat Swamp Soil Bacterium
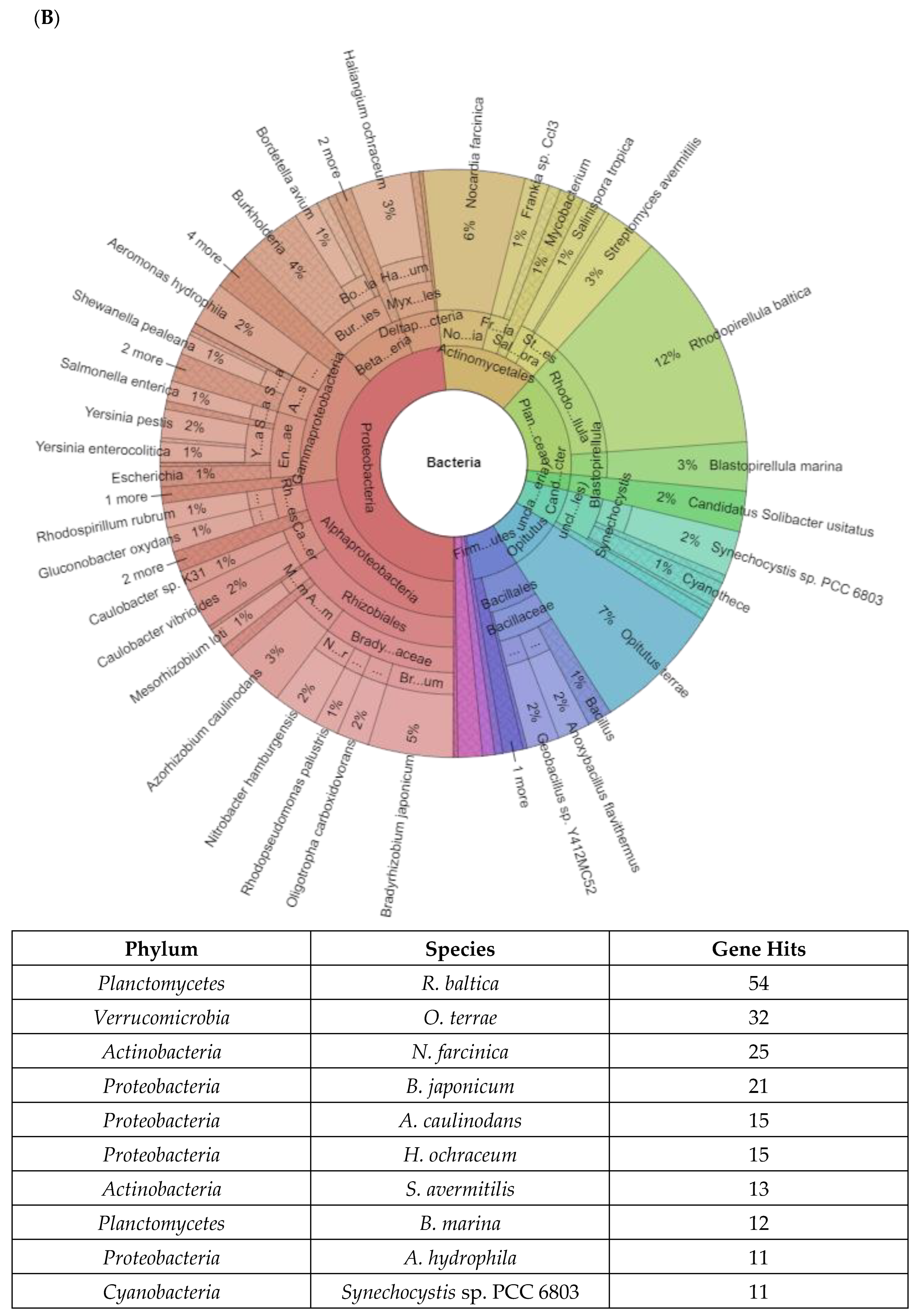
2.2. Taxonomy of Genes and Bacteria Diversity Involved in Stress Response
3. Discussion
4. Materials and Methods
4.1. Experimental Sites
4.2. Soil Bacterial Metagenome Extraction
4.3. Gene Annotation and Sequence Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Page, S.E.; Rieley, J.O.; Banks, C.J. Global and Regional Importance of the Tropical Peatland Carbon Pool: Tropical Peatland Carbon Pool. Glob. Chang. Biol. 2011, 17, 798–818. [Google Scholar] [CrossRef] [Green Version]
- Dargie, G.C.; Lewis, S.L.; Lawson, I.T.; Mitchard, E.T.A.; Page, S.E.; Bocko, Y.E.; Ifo, S.A. Age, Extent and Carbon Storage of the Central Congo Basin Peatland Complex. Nature 2017, 542, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Renou-Wilson, F. The Soils of Ireland; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Abdulkarim, B.; Yacob, M.R.; Abdullah, A.M.; Radam, A. Farmers’ Perceptions and Attitudes toward Forest Watershed Conservation of the North Selangor Peat Swamp Forest. J. Sustain. For. 2017, 10, 309–323. [Google Scholar] [CrossRef]
- Siti-Munirah, M.Y.; Suhaimi-Miloko, Z.; Ahmad, M.I.Z. Thismia belumensis (Thismiaceae), a remarkable new species from The Royal Belum State Park, Gerik, Perak, Peninsular Malaysia. PhytoKeys 2021, 172, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.W.; Soepadmo, E.; Yap, S.K. The Malaysian Heritage and Scientific Expedition to Belum: Temengor Forest Reserve; Perak, Malaysia. Malay. Nat. J. 1995, 48, 133–146. [Google Scholar]
- Kanniah, K.D.; Tan, K.P.; Cracknell, A.P.; Huete, A.R.; Idris, N.H.; Lau, A.M.S.; Abd Rahman, M.Z.; Rasib, A.W.; Ahmad, A. Assessment of Biophysical Properties of Royal Belum Tropical Forest, Malaysia: Biophysical Properties of Royal Belum Forest. Singap. J. Trop. Geogr. 2018, 39, 90–106. [Google Scholar] [CrossRef]
- Kedri, F.K.; Hamzah, Z.; Sukri, N.S.; Yaacob, S.H.; Majid, A.; Mokhtar, N.K.; Amir, N. Distribution and Ecology of Rafflesia in Royal Belum State Park, Perak, Malaysia. Int. J. Eng. Technol. 2018, 7, 292–296. [Google Scholar] [CrossRef]
- Misni, A.; Rasam, A.R.; Buyadi, S.N. Spatial Analysis of Habitat Conservation for Hornbills: A Case Study of Royal Belum-Temengor Forest Complex in Perak State Park, Malaysia. Malaysia. Pertanika J. Soc. Sci. Humanit. 2017, 25, 11–20. [Google Scholar]
- Schwabe, K.A.; Carson, R.T.; DeShazo, J.R.; Potts, M.D.; Reese, A.N.; Vincent, J.R. Creation of Malaysia’s Royal Belum State Park: A Case Study of Conservation in a Developing Country. J. Environ. Dev. 2015, 24, 54–81. [Google Scholar] [CrossRef] [Green Version]
- Malaysian Nature Society. MNS Position Statement. Kuala Lumpur, Malaysia. 2005. Available online: https://www.mns.my/ (accessed on 20 October 2022).
- Lazarus, B.A.; Shah, M.M.; Hamdan, A.; Hassan, A.N.; Mohammad, M.S.; Hassim, H.A.; Noor, M.H.; Azizan, T.R.; Ahmad, H. Topographical Differences Impacting Wildlife Dynamics at Natural Saltlicks in the Royal Belum Rainforest. Asian J. Conserv. Biol. 2019, 8, 97–101. [Google Scholar]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A Global Atlas of the Dominant Bacteria Found in Soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N. Embracing the Unknown: Disentangling the Complexities of the Soil Microbiome. Nat. Rev. Microbiol. 2017, 15, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; van der Putten, W.H. Belowground Biodiversity and Ecosystem Functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Bodelier, P.L.E. Toward Understanding, Managing, and Protecting Microbial Ecosystems. Front. Microbiol. 2011, 2, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, K.S.; Leff, J.W.; Barberán, A.; Bates, S.T.; Betley, J.; Crowther, T.W.; Kelly, E.F.; Oldfield, E.E.; Shaw, E.A.; Steenbock, C.; et al. Biogeographic Patterns in Below-Ground Diversity in New York City’s Central Park Are Similar to Those Observed Globally. Proc. Biol. Sci. 2014, 281, 20141988. [Google Scholar] [CrossRef] [PubMed]
- Diamond, S.; Andeer, P.F.; Li, Z.; Crits-Christoph, A.; Burstein, D.; Anantharaman, K.; Lane, K.R.; Thomas, B.C.; Pan, C.; Northen, T.R.; et al. Mediterranean Grassland Soil C-N Compound Turnover Is Dependent on Rainfall and Depth, and Is Mediated by Genomically Divergent Microorganisms. Nat. Microbiol. 2019, 4, 1356–1367. [Google Scholar] [CrossRef] [Green Version]
- Guan, N.; Liu, L. Microbial Response to Acid Stress: Mechanisms and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Land, M.; Hauser, L.; Jun, S.-R.; Nookaew, I.; Leuze, M.R.; Ahn, T.-H.; Karpinets, T.; Lund, O.; Kora, G.; Wassenaar, T.; et al. Insights from 20 Years of Bacterial Genome Sequencing. Funct. Integr. Genom. 2015, 15, 141–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Girard, R.A.; Martin, T.; Edwards, J.; Thrash, J.C. Status of the Archaeal and Bacterial Census: An Update. MBio 2016, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Lok, C. Mining the Microbial Dark Matter. Nature 2015, 522, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and Function of the Global Topsoil Microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Hansen, M.A.; Hansen, L.H.; Jacquiod, S.; Sørensen, S.J. Bioinformatic Approaches Reveal Metagenomic Characterization of Soil Microbial Community. PLoS ONE 2014, 9, e93445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neal, A.L.; Bacq-Labreuil, A.; Zhang, X.; Clark, I.M.; Coleman, K.; Mooney, S.J.; Ritz, K.; Crawford, J.W. Soil as an extended composite phenotype of the microbial metagenome. Sci. Rep. 2020, 10, 10649. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of Soil Organic Matter via Biochemical and Physical Pathways of Litter Mass Loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Lal, R. Soil Organic Matter Content and Crop Yield. J. Soil Water Conserv. 2020, 75, 27A–32A. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.; Ni, K.; Wu, Z.; Zhang, J.; Yi, X.; Yang, X.; Ling, N.; You, Z.; Guo, S.; Ruan, J. Effect of Organic Substitution Rates on Soil Quality and Fungal Community Composition in a Tea Plantation with Long-Term Fertilization. Biol. Fertil. Soils 2020, 56, 633–646. [Google Scholar] [CrossRef]
- Piaszczyk, W.; Lasota, J.; Błońska, E. Effect of Organic Matter Released from Deadwood at Different Decomposition Stages on Physical Properties of Forest Soil. Forests 2019, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Kuzyakov, Y.; Bore, E.; Dippold, M. Soil Organic Matter Formation: Convergence and Divergence of Three Carbon Pathways: Stabilization, Recycling and Losses. In Proceedings of the 22nd EGU General Assembly, Online, 4–8 May 2020. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct Evidence for Microbial-Derived Soil Organic Matter Formation and Its Ecophysiological Controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Kleber, M. The Contentious Nature of Soil Organic Matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef]
- Paul, E.A. The Nature and Dynamics of Soil Organic Matter: Plant Inputs, Microbial Transformations, and Organic Matter Stabilization. Soil Biol. Biochem. 2016, 98, 109–126. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of Soil Organic Matter as an Ecosystem Property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, K.; Ahmad, M.F.; Yaacob, N.S.; Ikram, W.M.; Maniyam, M.N.; Abdullah, H.; Katayama, T.; Komatsu, K.; Kuwahara, V.S. Enhancement of Targeted Microalgae Species Growth Using Aquaculture Sludge Extracts. Heliyon 2020, 6, e04556. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, K.; Onodera, T.; Kohzu, A.; Syutsubo, K.; Imai, A. Characterization of Dissolved Organic Matter in Wastewater during Aerobic, Anaerobic, and Anoxic Treatment Processes by Molecular Size and Fluorescence Analyses. Water Res. 2020, 171, 115459. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, K.; Yaacob, N.S.; Abdullah, H.; Ahmad, M.F.; Maniyam, M.N.; Hashim, E.F.; Sjahrir, F.; Ikram, W.M.; Komatsu, K.; Kuwahara, V.S. Growth-Promoting Effects of Marine Microalgae Species Using Tropical Forest Soil Extracts. J. Appl. Biol. Biotechnol. 2022, 10, 1–3. [Google Scholar]
- Finn, D.; Yu, J.; Penton, C.R. Soil Quality Shapes the Composition of Microbial Community Stress Response and Core Cell Metabolism Functional Genes. Appl. Soil Ecol. 2020, 148, 103483. [Google Scholar] [CrossRef]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete Genome Sequence of the Model Actinomycete Streptomyces Coelicolor A3. Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Chain, P.S.G.; Denef, V.J.; Konstantinidis, K.T.; Vergez, L.M.; Agulló, L.; Reyes, V.L.; Hauser, L.; Córdova, M.; Gómez, L.; González, M.; et al. Burkholderia Xenovorans LB400 Harbors a Multi-Replicon, 9.73-Mbp Genome Shaped for Versatility. Proc. Natl. Acad. Sci. USA 2006, 103, 15280–15287. [Google Scholar] [CrossRef] [Green Version]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three Genomes from the Phylum Acidobacteria Provide Insight into the Lifestyles of These Microorganisms in Soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef] [Green Version]
- Rocca, J.D.; Simonin, M.; Blaszczak, J.R.; Ernakovich, J.G.; Gibbons, S.M.; Midani, F.S.; Washburne, A.D. The Microbiome Stress Project: Toward a Global Meta-Analysis of Environmental Stressors and Their Effects on Microbial Communities. Front. Microbiol. 2018, 9, 3272. [Google Scholar] [CrossRef]
- Maniyam, N.; Abdullah, M.; Ahmad, H.; Hashim, M.F.; Sjahrir, E.F.; Komatsu, F.; Kuwahara, K.; Yaacob, V.S. Malaysian Virgin Soil Extracts as Natural Growth Enhancer for Targeted Green Microalgae Species. Appl. Sci. 2022, 12, 4060. [Google Scholar] [CrossRef]
- Yaacob, N.S.; Ahmad, M.F.; Sivam, A.; Hashim, E.F.; Maniyam, M.N.; Sjahrir, F.; Dzulkafli, N.F.; Wan Mohd Zamri, W.M.I.; Komatsu, K.; Kuwahara, V.S.; et al. The Effectiveness of Soil Extracts from Selangor Peat Swamp and Pristine Forest Soils on the Growth of Green Microalgae sp. Forests 2022, 13, 79. [Google Scholar] [CrossRef]
- Yaacob, N.S.; Ahmad, M.F.; Kawasaki, N.; Maniyam, M.N.; Abdullah, H.; Hashim, E.F.; Sjahrir, F.; Wan Mohd Zamri, W.M.I.; Komatsu, K.; Kuwahara, V.S. Kinetics Growth and Recovery of Valuable Nutrients from Selangor Peat Swamp and Pristine Forest Soils Using Different Extraction Methods as Potential Microalgae Growth Enhancers. Molecules 2021, 26, 653. [Google Scholar] [CrossRef] [PubMed]
- Mohd Zamri, W.; Sjahrir, W.M.; Yaacob, F.; Dzulkafli, N.S.; Ahmad, N.F.; Abdullah, M.F.; Maniyam, H.; Hashim, M.N.; Kawasaki, E.F.; Komatsu, N.; et al. Assessment of Aqueous Extraction Methods on Extractable Organic Matter and Hydrophobic/Hydrophilic Fractions of Virgin Forest Soils. Molecules 2021, 26, 2480. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest Soil Bacteria: Diversity, Involvement in Ecosystem Processes, and Response to Global Change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siles, J.A.; Margesin, R. Abundance and Diversity of Bacterial, Archaeal, and Fungal Communities along an Altitudinal Gradient in Alpine Forest Soils: What Are the Driving Factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Bao, Y.; Dolfing, J.; Guo, Z.; Chen, R.; Wu, M.; Li, Z.; Lin, X.; Feng, Y. Important Ecophysiological Roles of Non-Dominant Actinobacteria in Plant Residue Decomposition, Especially in Less Fertile Soils. Microbiome 2021, 9, 84. [Google Scholar] [CrossRef]
- Larsbrink, J.; McKee, L.S. Bacteroidetes Bacteria in the Soil: Glycan Acquisition, Enzyme Secretion, and Gliding Motility. Adv. Appl. Microbiol. 2020, 110, 63–98. [Google Scholar] [CrossRef]
- Sanseverino, I.; Pretto, P.; António, D.C.; Lahm, A.; Facca, C.; Loos, R.; Skejo, H.; Beghi, A.; Pandolfi, F.; Genoni, P.; et al. Metagenomics Analysis to Investigate the Microbial Communities and Their Functional Profile during Cyanobacterial Blooms in Lake Varese. Microb. Ecol. 2022, 83, 850–868. [Google Scholar] [CrossRef]
- Too, C.C.; Keller, A.; Sickel, W.; Lee, S.M.; Yule, C.M. Microbial Community Structure in a Malaysian Tropical Peat Swamp Forest: The Influence of Tree Species and Depth. Front. Microbiol. 2018, 9, 2859. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Samaddar, S.; Chatterjee, P.; Roy Choudhury, A.; Choi, J.; Choi, J.; Sa, T. Structural and Functional Shift in Soil Bacterial Community in Response to Long-Term Compost Amendment in Paddy Field. Appl. Sci. 2021, 11, 2183. [Google Scholar] [CrossRef]
- Penn, C.J.; Camberato, J.J. A Critical Review on Soil Chemical Processes That Control How Soil PH Affects Phosphorus Availability to Plants. Agriculture 2019, 9, 120. [Google Scholar] [CrossRef] [Green Version]
- Barrow, N.J. The Effects of PH on Phosphate Uptake from the Soil. Plant Soil 2017, 410, 401–410. [Google Scholar] [CrossRef]
- Aislabie, J.M.; Jordan, S.; Barker, G.M. Relation between Soil Classification and Bacterial Diversity in Soils of the Ross Sea Region, Antarctica. Geoderma 2008, 144, 9–20. [Google Scholar] [CrossRef]
- Ning, Z.; Zhang, M.; He, Z.; Cai, P.; Guo, C.; Wang, P. Spatial Pattern of Bacterial Community Diversity Formed in Different Groundwater Field Corresponding to Electron Donors and Acceptors Distributions at a Petroleum-Contaminated Site. Water 2018, 10, 842. [Google Scholar] [CrossRef] [Green Version]
- Pietri, J.A.; Brookes, P.C. Relationships between Soil PH and Microbial Properties in a UK Arable Soil. Soil Biol. Biochem. 2008, 40, 1856–1861. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. The Soil Microbiome-from Metagenomics to Metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef]
- Desjardins, P.; Conklin, D. NanoDrop Microvolume Quantitation of Nucleic Acids. J. Vis. Exp. 2010, 45, e2565. [Google Scholar] [CrossRef] [Green Version]
- Oberacker, P.; Stepper, P.; Bond, D.M.; Höhn, S.; Focken, J.; Meyer, V.; Schelle, L.; Sugrue, V.J.; Jeunen, G.-J.; Moser, T.; et al. Bio-On-Magnetic-Beads (BOMB): Open Platform for High-Throughput Nucleic Acid Extraction and Manipulation. PLoS Biol. 2019, 17, e3000107. [Google Scholar] [CrossRef] [Green Version]
- Kong, N.; Thao, K.; Huang, C.; Appel, M.; Lappin, S.; Knapp, L.; Kelly, L.; Weimer, B.C. Automated Library Construction Using KAPA Library Preparation Kits on the Agilent NGS Workstation Yields High-Quality Libraries for Whole-Genome Sequencing on the Illumina Platform; Agilent Technologies Application Note; Agilent Technologies: Santa Clara, CA, USA, 2014; Available online: http://www.agilent.com/cs/library/applications/5991-4296EN (accessed on 14 September 2022).
- Masotti, A.; Preckel, T. Analysis of Small RNAs with the Agilent 2100 Bioanalyzer. Nat. Methods 2006, 3, 658. [Google Scholar] [CrossRef]
- Sevim, V.; Lee, J.; Egan, R.; Clum, A.; Hundley, H.; Lee, J.; Everroad, R.C.; Detweiler, A.M.; Bebout, B.M.; Pett-Ridge, J.; et al. Shotgun Metagenome Data of a Defined Mock Community Using Oxford Nanopore, PacBio and Illumina Technologies. Sci. Data 2019, 6, 285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-Glance Quality Assessment of Illumina Second-Generation Sequencing Data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. MetaSPAdes: A New Versatile Metagenomic Assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Ruscheweyh, M.S.; Tappu, H.J. MEGAN Community Edition-Interactive Exploration and Analysis of Large-Scale Microbiome Sequencing Data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, F.; Paarmann, D.; Souza, D.; Olson, M.; Glass, R.E.; Rodriguez, R.; Wilke, J.; Wilkening, R. The Metagenomics RAST Server-a Public Resource for the Automatic Phylogenetic and Functional Analysis of Metagenomes. BMC Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boetzer, M.; Henkel, C.V.; Jansen, H.J.; Butler, D.; Pirovano, W. Scaffolding Pre-Assembled Contigs Using SSPACE. Bioinformatics 2011, 27, 578–579. [Google Scholar] [CrossRef] [Green Version]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Types | Sites | Phylum | Species |
|---|---|---|---|
| Rainforest | SK | 129 | 6264 |
| SP | 44 | 5560 | |
| SR | 81 | 3411 | |
| Peat Swamp Forest | RM1 | 128 | 5493 |
| RM2 | 100 | 3092 | |
| RM3 | 131 | 4305 |
| Phylum | Royal Belum Bacteria Abundances (%) | Raja Muda Musa Peat Swamp Bacteria Abundances (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| SK | SP | SR | Total | RM1 | RM2 | RM3 | Total | |
| Proteobacteria | 51.18 | 52.38 | 52.89 | 52.15 | 43.66 | 49.68 | 51.51 | 48.28 |
| Actinobacteria | 19.21 | 22.6 | 16.19 | 19.33 | 2.17 | 17.16 | 15.48 | 19.27 |
| Acidobacteria | 12.16 | 11.6 | 11.34 | 11.7 | 13.68 | 8.83 | 15.42 | 12.64 |
| Verrucomicrobia | 3.17 | 1.4 | 4.19 | 2.92 | 1.57 | 1.98 | 1.73 | 1.76 |
| Firmicutes | 2.97 | 2.6 | 2.86 | 2.81 | 3.6 | 6.98 | 3.79 | 4.79 |
| Bacteroidetes | 1.68 | 1.15 | 2.09 | 1.64 | 1.03 | 2.67 | 1.27 | 1.66 |
| Planctomycetes | 2.05 | 1.11 | 2.34 | 1.83 | 0.79 | 3.42 | 0.97 | 1.73 |
| Cyanobacteria | 1.75 | 1.59 | 1.78 | 1.71 | 1.63 | 1.91 | 1.72 | 1.75 |
| Chloroflexi | 1.47 | 2.29 | 1.3 | 1.69 | 1.65 | 2.02 | 1.58 | 1.75 |
| Nitrospirae | 0.67 | 0.2 | 1.14 | 0.67 | 0.23 | 0.26 | 0.27 | 0.25 |
| Deinococcus–Thermus | 0.46 | 0.04 | 0.42 | 0.31 | 0.56 | 0.56 | 0.54 | 0.55 |
| Others | 3.23 | 3.04 | 3.46 | 3.24 | 6.43 | 4.53 | 5.72 | 5.57 |
| Total | Protein-Coding Region | Functional Genes | Stress-Response Genes | Acidic Stress-Response Genes |
|---|---|---|---|---|
| Royal Belum | 1,391,841 | 421,409 | 11,751 | 177 |
| Raja Musa | 3,586,577 | 1,111,985 | 28,750 | 449 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, M.F.; Abdullah, H.; Hassan, M.N.; Jamaludin, M.I.; Sivam, A.; Komatsu, K.; Sapian, I.S.; Alias, H.; Mat Isa, M.N.; Kuwahara, V.S.; et al. Topographically Distinguished Microbiome Taxonomy and Stress-Response Genes of Royal Belum Rainforest and Raja Muda Musa Peat Swamp Revealed through Metagenomic Inquisition. Int. J. Mol. Sci. 2023, 24, 872. https://doi.org/10.3390/ijms24010872
Ahmad MF, Abdullah H, Hassan MN, Jamaludin MI, Sivam A, Komatsu K, Sapian IS, Alias H, Mat Isa MN, Kuwahara VS, et al. Topographically Distinguished Microbiome Taxonomy and Stress-Response Genes of Royal Belum Rainforest and Raja Muda Musa Peat Swamp Revealed through Metagenomic Inquisition. International Journal of Molecular Sciences. 2023; 24(1):872. https://doi.org/10.3390/ijms24010872
Chicago/Turabian StyleAhmad, Mohd Fadzli, Hasdianty Abdullah, Muhammad Naim Hassan, Muhammad Imran Jamaludin, Ashvini Sivam, Kazuhiro Komatsu, Irni Suhayu Sapian, Halimah Alias, Mohd Noor Mat Isa, Victor S. Kuwahara, and et al. 2023. "Topographically Distinguished Microbiome Taxonomy and Stress-Response Genes of Royal Belum Rainforest and Raja Muda Musa Peat Swamp Revealed through Metagenomic Inquisition" International Journal of Molecular Sciences 24, no. 1: 872. https://doi.org/10.3390/ijms24010872
APA StyleAhmad, M. F., Abdullah, H., Hassan, M. N., Jamaludin, M. I., Sivam, A., Komatsu, K., Sapian, I. S., Alias, H., Mat Isa, M. N., Kuwahara, V. S., & Yaacob, N. S. (2023). Topographically Distinguished Microbiome Taxonomy and Stress-Response Genes of Royal Belum Rainforest and Raja Muda Musa Peat Swamp Revealed through Metagenomic Inquisition. International Journal of Molecular Sciences, 24(1), 872. https://doi.org/10.3390/ijms24010872