
Alterations in Antibiotic Susceptibility of Staphylococcus aureus and Klebsiella pneumoniae in Dual Species Biofilms

 , , and
, , and
Abstract
:1. Introduction
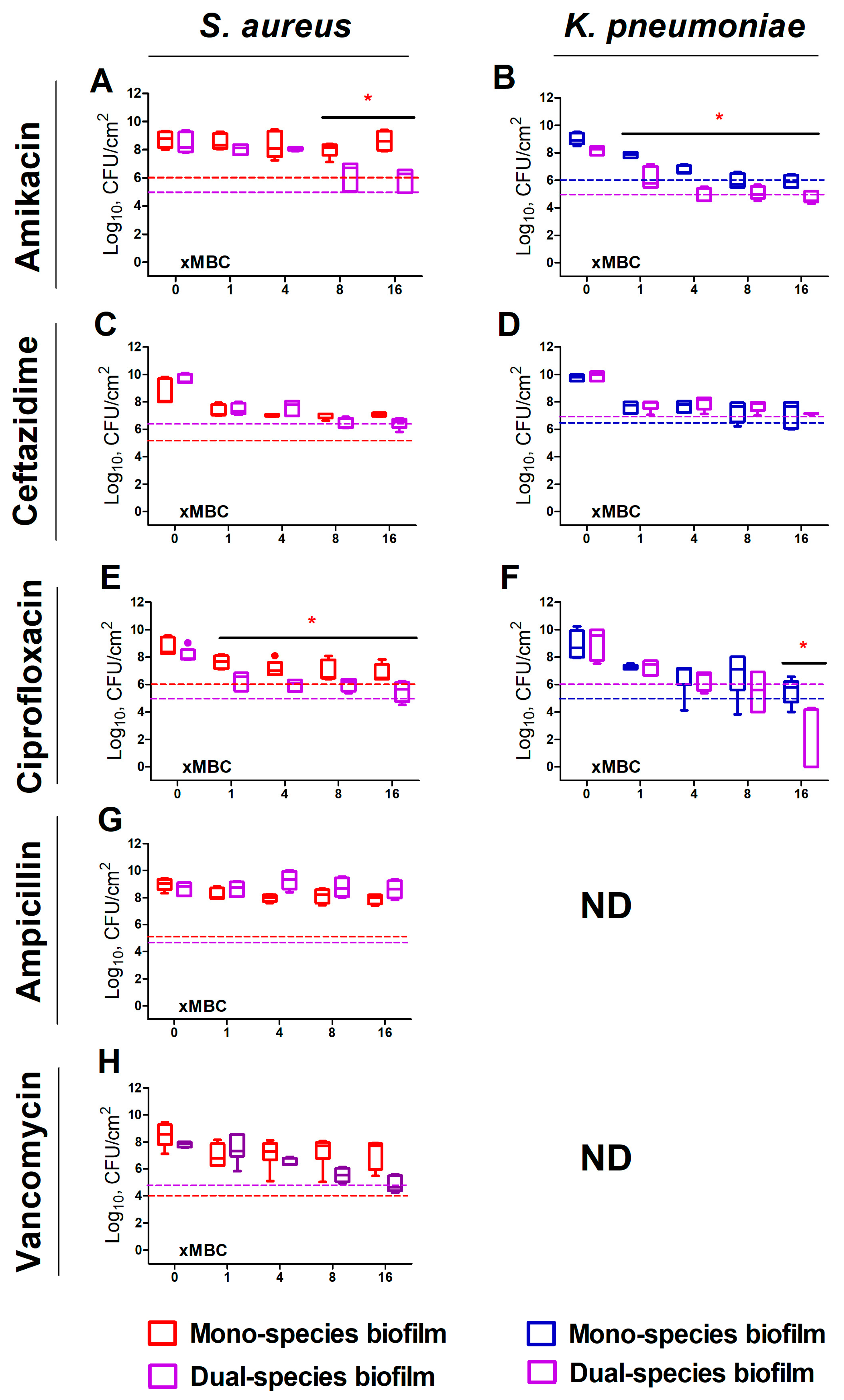
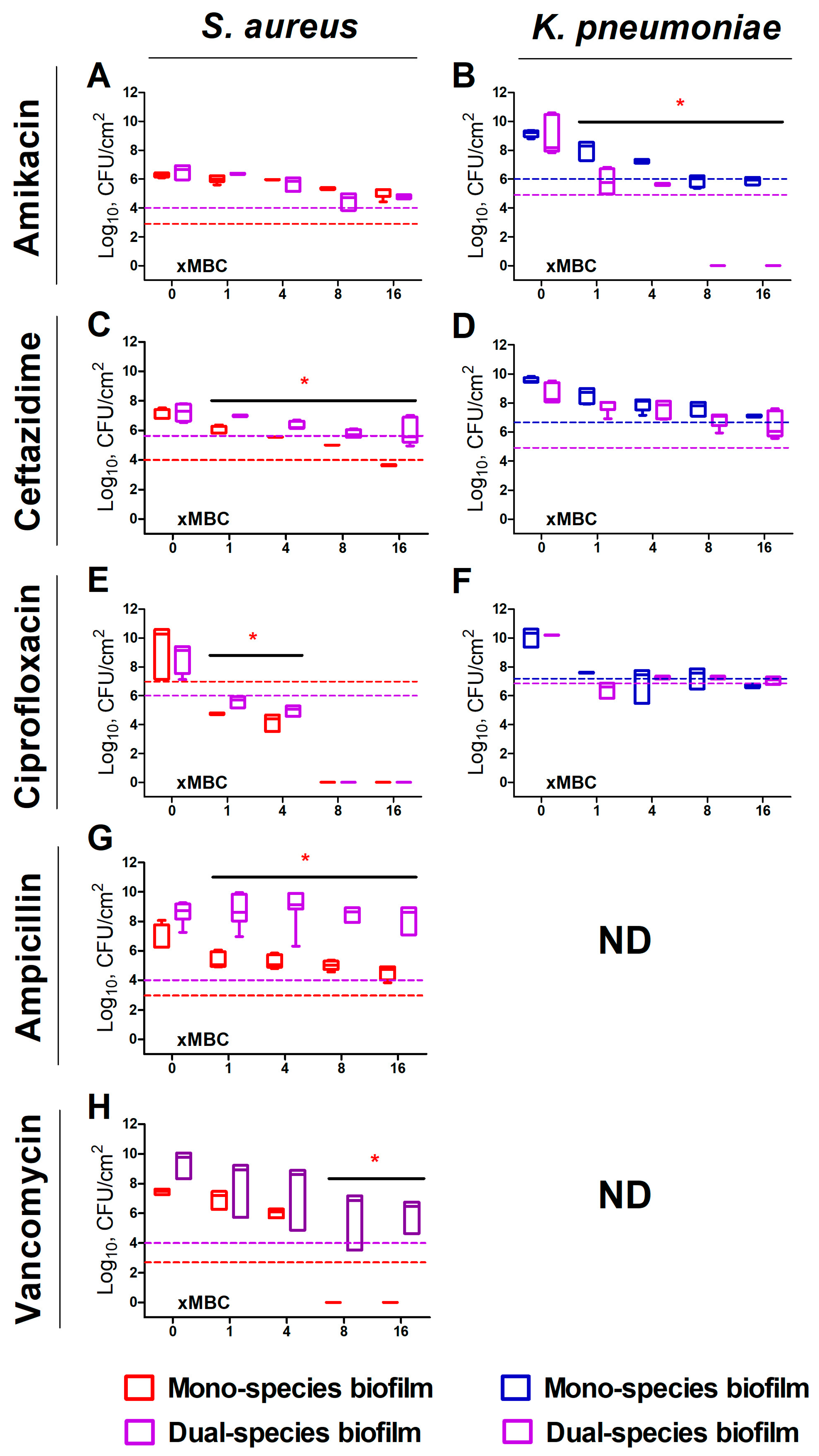
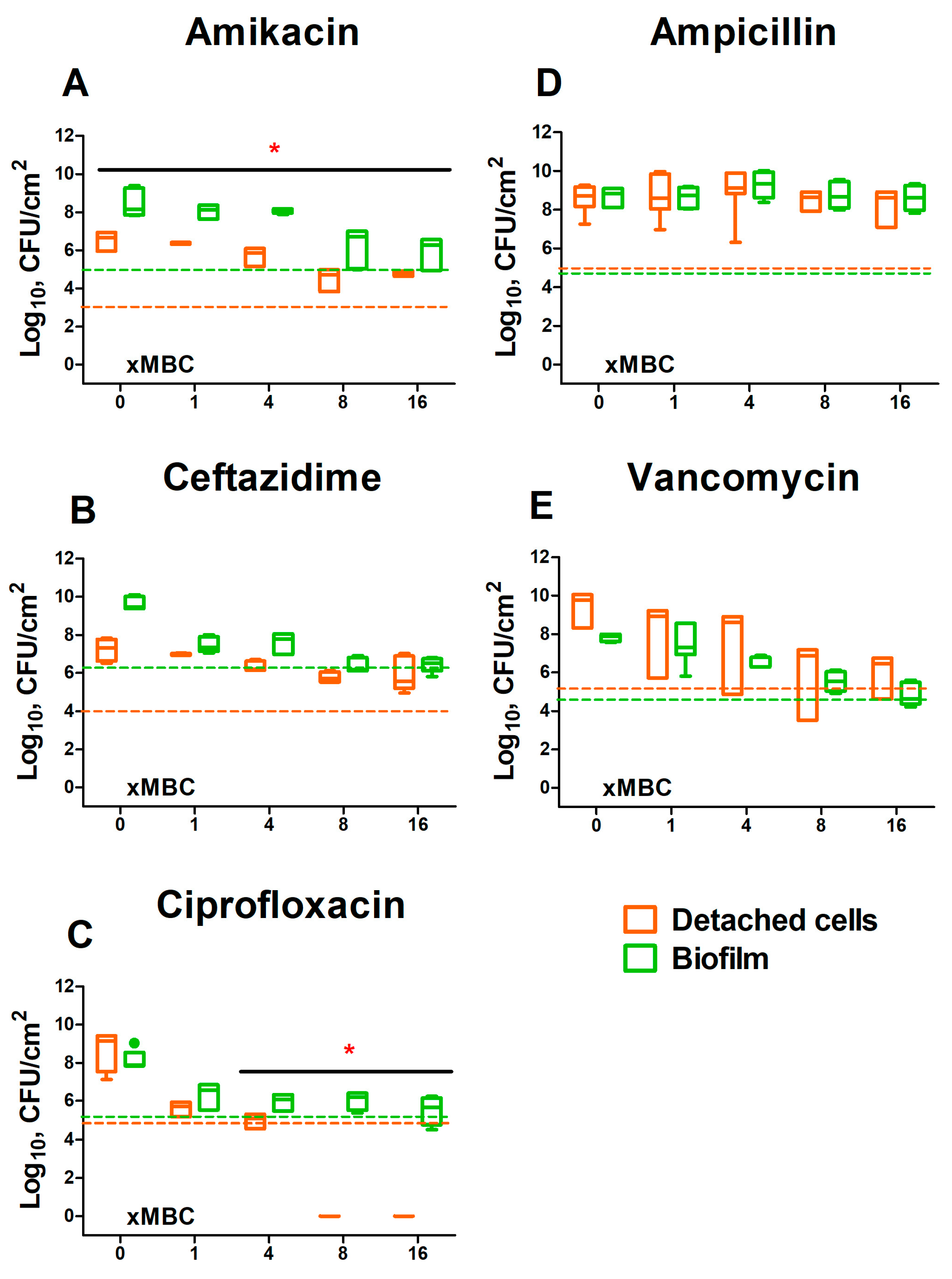
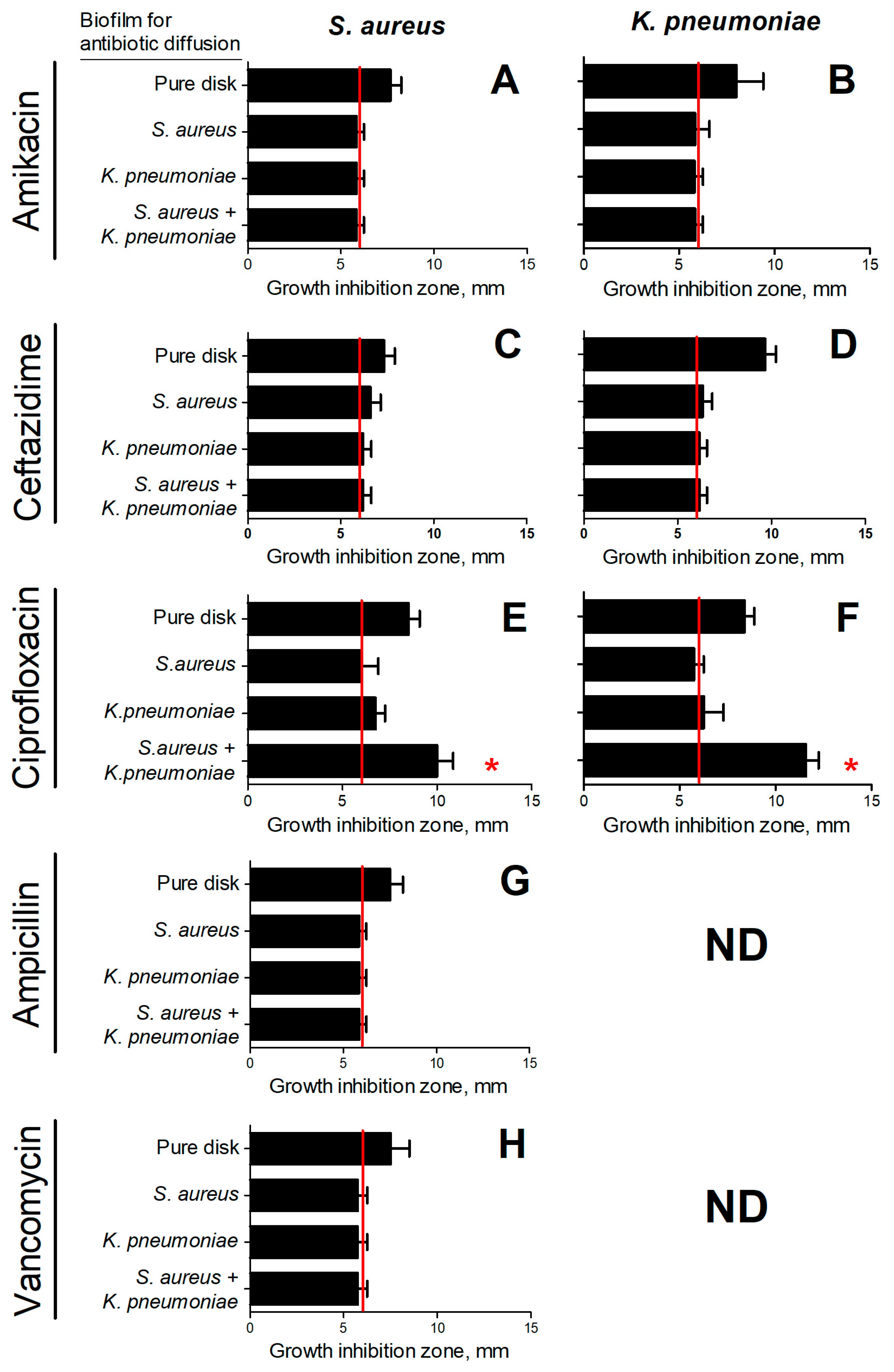
2. Results
2.1. S. aureus and K. pneumoniae Susceptibility to Antibiotics in Mixed Cultures
2.2. Evaluation of the Permeability of S. aureus and K. pneumoniae Mono- and Dual-Species Biofilms for Antimicrobials
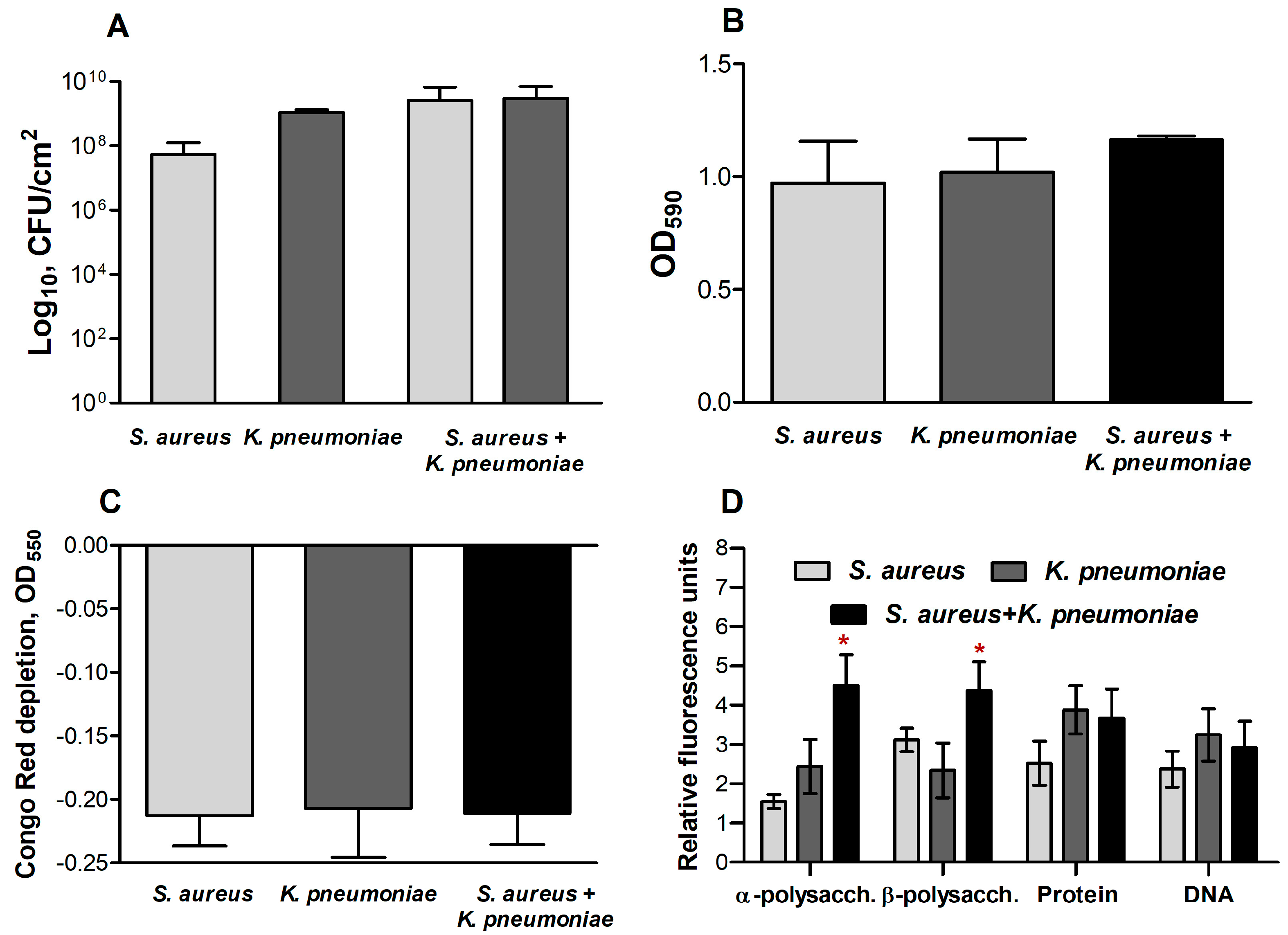
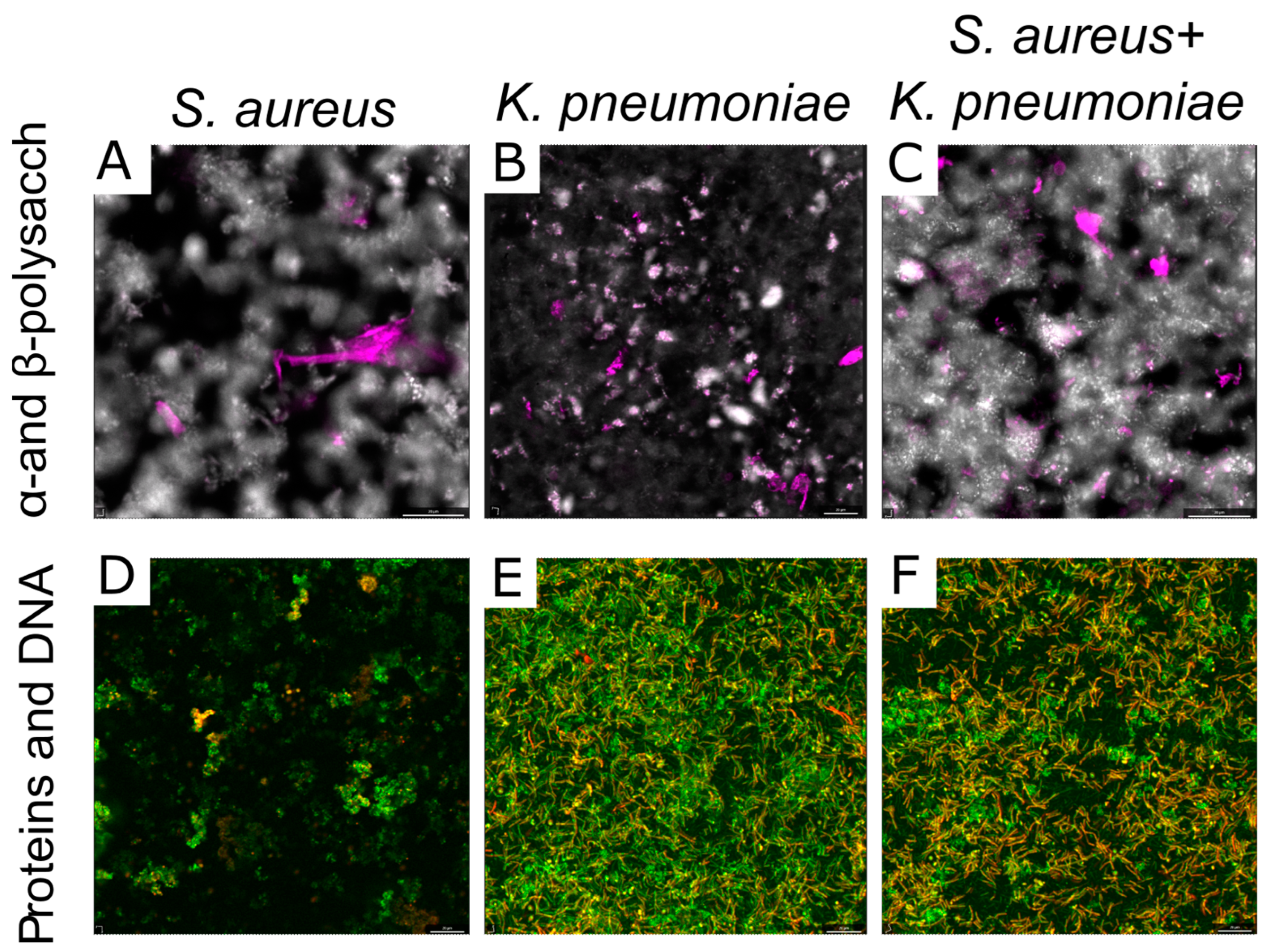
2.3. The Biochemical Composition of Extracellular Matrix in S. aureus and K. pneumoniae Mono- and Dual-Species Biofilms
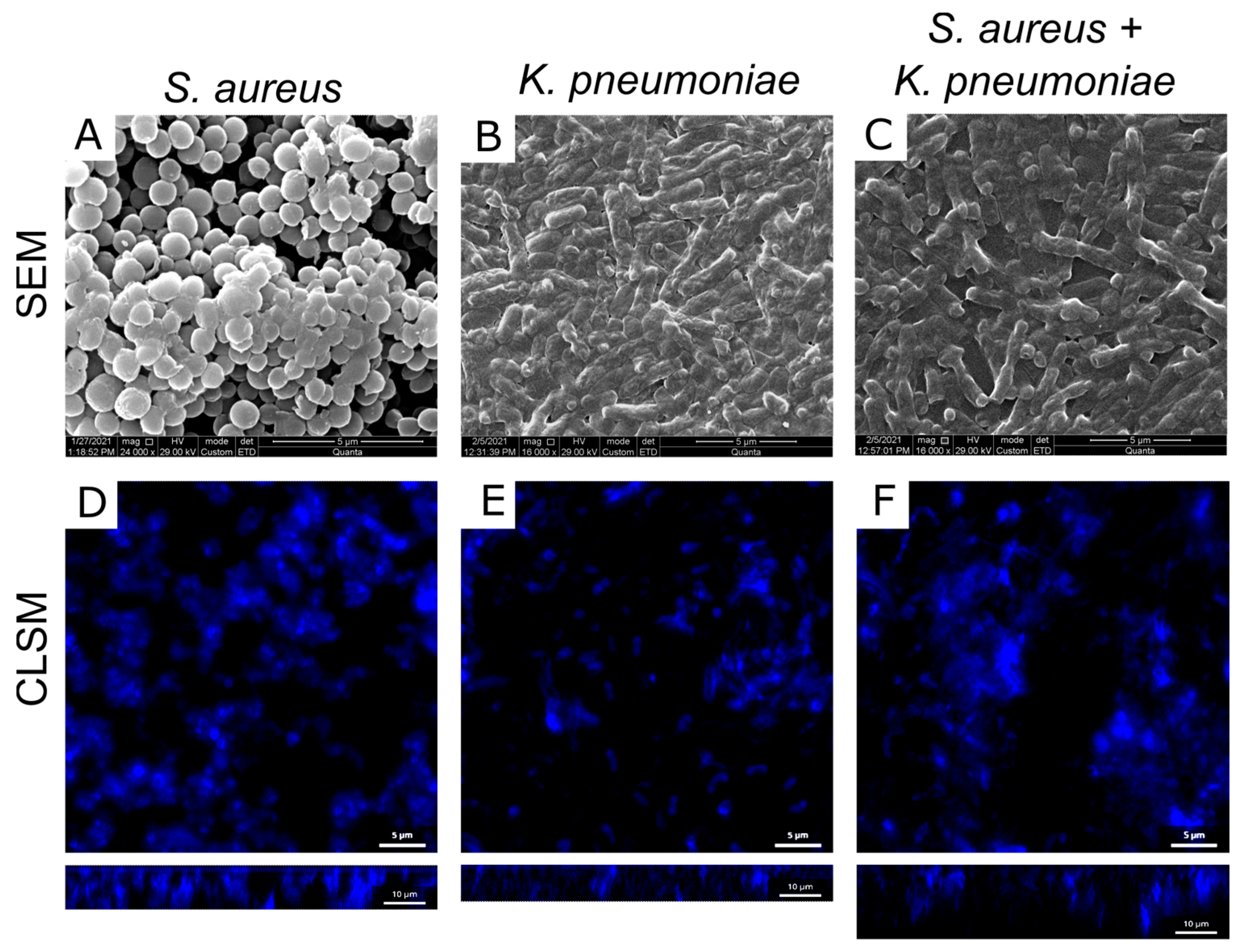
2.4. Microscopy
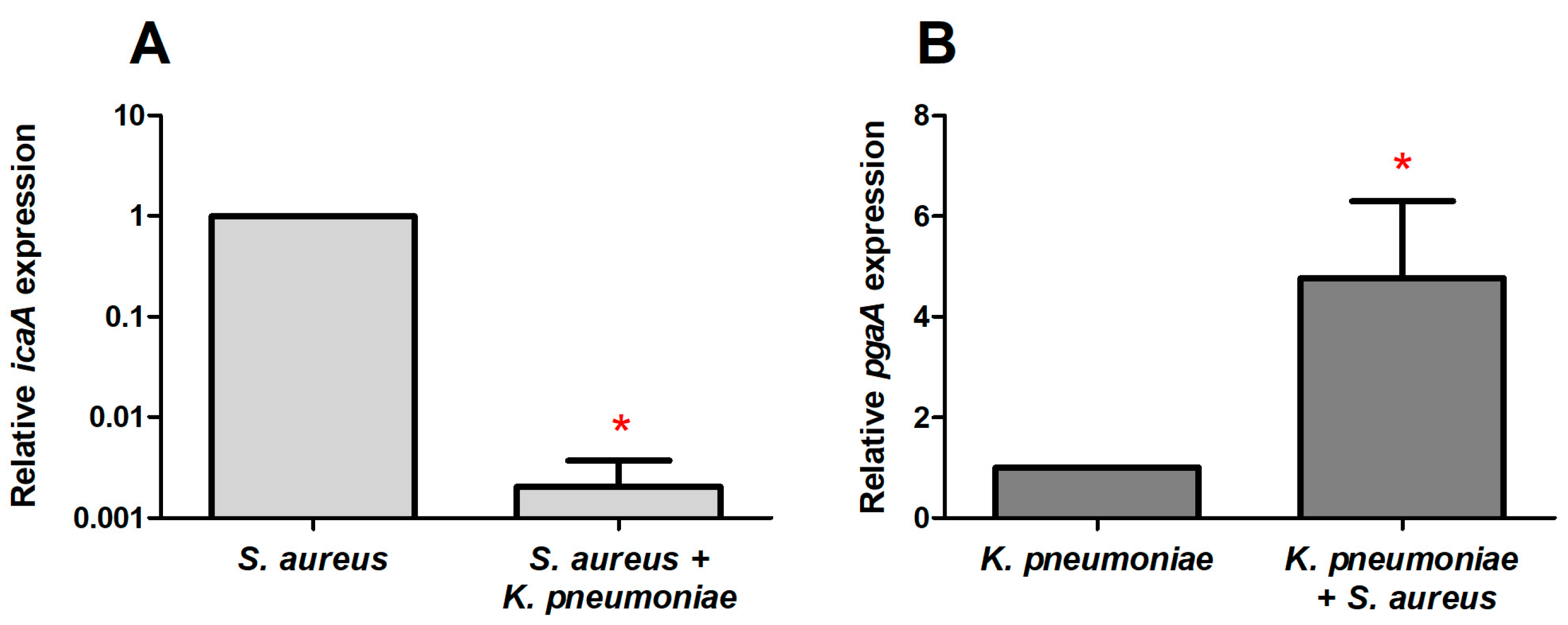
2.5. The Expression of Genes Involved in the Synthesis of the Extracellular Matrix in Mono- and Dual-Species Biofilms of S. aureus–K. pneumoniae
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Media
4.2. Determination of Minimum Inhibitory Concentration and Minimum Bactericidal Concentration
4.3. Biofilm Assays
4.4. CFUs Counting
4.5. The Quantification of Matrix Composition
4.6. Microscopy
4.7. Penetration of Antimicrobials Throw Biofilm Matrix
4.8. RNA Isolation and Real-Time One-Step qRT-PCR
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frieri, M.; Kumar, K.; Boutin, A. Antibiotic resistance. J. Infect. Public Health 2017, 10, 369–378. [Google Scholar] [CrossRef]
- Jiang, Y.; Geng, M.; Bai, L. Targeting Biofilms Therapy: Current Research Strategies and Development Hurdles. Microorganisms 2020, 8, 1222. [Google Scholar] [CrossRef]
- Schulze, A.; Mitterer, F.; Pombo, J.P.; Schild, S. Biofilms by bacterial human pathogens: Clinical relevance development, composition and regulation therapeutical strategies. Microb. Cell 2021, 8, 28–56. [Google Scholar] [CrossRef]
- Uruen, C.; Chopo-Escuin, G.; Tommassen, J.; Mainar-Jaime, R.C.; Arenas, J. Biofilms as promoters of bacterial antibiotic resistance and tolerance. Antibiotics 2021, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Karygianni, L.; Ren, Z.; Koo, H.; Thurnheer, T. Biofilm Matrixome: Extracellular Components in Structured Microbial Communities. Trends Microbiol. 2020, 28, 668–681. [Google Scholar] [CrossRef]
- Luo, A.; Wang, F.; Sun, D.; Liu, X.; Xin, B. Formation, Development, and Cross-Species Interactions in Biofilms. Front. Microbiol. 2021, 12, 757327. [Google Scholar] [CrossRef]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef]
- Abebe, G.M. The Role of Bacterial Biofilm in Antibiotic Resistance and Food Contamination. Int. J. Microbiol. 2020, 2020, 1705814. [Google Scholar] [CrossRef] [PubMed]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Batoni, G.; Maisetta, G.; Esin, S. Therapeutic Potential of Antimicrobial Peptides in Polymicrobial Biofilm-Associated Infections. Int. J. Mol. Sci. 2021, 22, 482. [Google Scholar] [CrossRef]
- Bernardy, E.E.; Petit, R.A.; Raghuram, V.; Alexander, A.M.; Read, T.D.; Goldberg, J.B. Genotypic and Phenotypic Diversity of Staphylococcus aureus Isolates from Cystic Fibrosis Patient Lung Infections and Their Interactions with Pseudomonas aeruginosa. mBio 2020, 11, 00735-20. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Cendra, M.D.M.; Blanco-Cabra, N.; Pedraz, L.; Torrents, E. Optimal environmental and culture conditions allow the in vitro coexistence of Pseudomonas aeruginosa and Staphylococcus aureus in stable biofilms. Sci. Rep. 2019, 9, 16284. [Google Scholar] [CrossRef]
- Booth, S.C.; Rice, S.A. Influence of interspecies interactions on the spatial organization of dual species bacterial communities. Biofilm 2020, 2, 100035. [Google Scholar] [CrossRef]
- Orazi, G.; Ruoff, K.L.; O’Toole, G.A. Pseudomonas aeruginosa Increases the Sensitivity of Biofilm-Grown Staphylococcus aureus to Membrane-Targeting Antiseptics and Antibiotics. mBio 2019, 10, 01501-19. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Patel, S.K.S.; Kharga, K.; Kumar, R.; Kumar, P.; Pandohee, J.; Kulshresha, S.; Harjai, K.; Chhibber, S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules 2022, 27, 7584. [Google Scholar] [CrossRef]
- Kim, U.; Kim, J.H.; Oh, S.W. Review of multi-species biofilm formation from foodborne pathogens: Multi-species biofilms and removal methodology. Crit. Rev. Food Sci. Nutr. 2022, 62, 5783–5793. [Google Scholar] [CrossRef]
- Trizna, E.Y.; Yarullina, M.N.; Baidamshina, D.R.; Mironova, A.V.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Bidirectional alterations in antibiotics susceptibility in Staphylococcus aureus-Pseudomonas aeruginosa dual-species biofilm. Sci. Rep. 2020, 10, 14849. [Google Scholar] [CrossRef]
- Biswas, L.; Götz, F. Molecular Mechanisms of Staphylococcus and Pseudomonas Interactions in Cystic Fibrosis. Front. Cell. Infect. Microbiol. 2022, 11, 1383. [Google Scholar] [CrossRef] [PubMed]
- Orazi, G.; O’Toole, G.A. Pseudomonas aeruginosa Alters Staphylococcus aureus Sensitivity to Vancomycin in a Biofilm Model of Cystic Fibrosis Infection. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Guerra, M.E.S.; Destro, G.; Vieira, B.; Lima, A.S.; Ferraz, L.F.C.; Hakansson, A.P.; Darrieux, M.; Converso, T.R. Biofilms and Their Role in Disease Pathogenesis. Front. Cell. Infect. Microbiol. 2022, 12, 877995. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.Y. The emerging problems of Klebsiella pneumoniae infections: Carbapenem resistance and biofilm formation. FEMS Microbiol. Lett. 2016, 363, 219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, Q.; Liao, K.; Ni, Y.; Yu, Y.; Hu, B.; Sun, Z.; Huang, W.; Wang, Y.; Wu, A.; et al. Update of incidence and antimicrobial susceptibility trends of Escherichia coli and Klebsiella pneumoniae isolates from Chinese intra-abdominal infection patients. BMC Infect. Dis. 2017, 17, 776. [Google Scholar] [CrossRef] [PubMed]
- Choby, J.E.; Howard-Anderson, J.; Weiss, D.S. Hypervirulent Klebsiella pneumoniae clinical and molecular perspectives. J. Intern. Med. 2020, 287, 283–300. [Google Scholar] [CrossRef]
- Arato, V.; Raso, M.M.; Gasperini, G.; Berlanda Scorza, F.; Micoli, F. Prophylaxis and Treatment against. Int. J. Mol. Sci. 2021, 22, 42. [Google Scholar] [CrossRef]
- Tanner, W.D.; Atkinson, R.M.; Goel, R.K.; Toleman, M.A.; Benson, L.S.; Porucznik, C.A.; VanDerslice, J.A. Horizontal transfer of the blaNDM-1 gene to Pseudomonas aeruginosa and Acinetobacter baumannii in biofilms. FEMS Microbiol. Lett. 2017, 364, 048. [Google Scholar] [CrossRef]
- Thuy, D.B.; Campbell, J.; Thuy, C.T.; Hoang, N.V.M.; Voong Vinh, P.; Nguyen, T.N.T.; Nguyen Ngoc Minh, C.; Pham, D.T.; Rabaa, M.A.; Lan, N.P.H.; et al. Colonization with Staphylococcus aureus and Klebsiella pneumoniae causes infections in a Vietnamese intensive care unit. Microb. Genom. 2021, 7, 000514. [Google Scholar] [CrossRef]
- Li, Y.; Wu, M.X. Reversal of Polymicrobial Biofilm Tolerance to Ciprofloxacin by Blue Light plus Carvacrol. Microorganisms 2021, 9, 2074. [Google Scholar] [CrossRef]
- Fisher, J.F.; Mobashery, S. β-Lactams against the Fortress of the Gram-Positive. Chem. Rev. 2021, 121, 3412–3463. [Google Scholar] [CrossRef]
- Thomsen, I.P.; Liu, G.Y. Targeting fundamental pathways to disrupt Staphylococcus aureus survival: Clinical implications of recent discoveries. JCI Insight 2018, 3, 98216. [Google Scholar] [CrossRef]
- Esposito, S.; Blasi, F.; Curtis, N.; Kaplan, S.; Lazzarotto, T.; Meschiari, M.; Mussini, C.; Peghin, M.; Rodrigo, C.; Vena, A.; et al. New Antibiotics for. Antibiotics 2023, 12, 742. [Google Scholar] [CrossRef]
- Lee, K.W.; Periasamy, S.; Mukherjee, M.; Xie, C.; Kjelleberg, S.; Rice, S.A. Biofilm development and enhanced stress resistance of a model, mixed-species community biofilm. ISME J. 2014, 8, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Lee, K.M.; Kim, D.; Yoon, S.S. Molecular Determinants of the Thickened Matrix in a Dual-Species Pseudomonas aeruginosa and Enterococcus faecalis Biofilm. Appl. Environ. Microbiol. 2017, 83, 01182-17. [Google Scholar] [CrossRef]
- Moormeier, D.E.; Endres, J.L.; Mann, E.E.; Sadykov, M.R.; Horswill, A.R.; Rice, K.C.; Fey, P.D.; Bayles, K.W. Use of microfluidic technology to analyze gene expression during Staphylococcus aureus biofilm formation reveals distinct physiological niches. Appl. Environ. Microbiol. 2013, 79, 3413–3424. [Google Scholar] [CrossRef]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential gene expression profiling of Staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococci in the human microbiome: The role of host and interbacterial interactions. Curr. Opin. Microbiol. 2020, 53, 71–77. [Google Scholar] [CrossRef]
- Vuotto, C.; Longo, F.; Balice, M.P.; Donelli, G.; Varaldo, P.E. Antibiotic Resistance Related to Biofilm Formation in Klebsiella pneumoniae. Pathogens 2014, 3, 743–758. [Google Scholar] [CrossRef]
- Radlinski, L.; Rowe, S.E.; Kartchner, L.B.; Maile, R.; Cairns, B.A.; Vitko, N.P.; Gode, C.J.; Lachiewicz, A.M.; Wolfgang, M.C.; Conlon, B.P. Pseudomonas aeruginosa exoproducts determine antibiotic efficacy against Staphylococcus aureus. PLoS Biol. 2017, 15. [Google Scholar] [CrossRef]
- Kvich, L.; Crone, S.; Christensen, M.H.; Lima, R.; Alhede, M.; Staerk, D.; Bjarnsholt, T. Investigation of the Mechanism and Chemistry Underlying Staphylococcus aureus. J. Bacteriol. 2022, 204, e0017422. [Google Scholar] [CrossRef] [PubMed]
- Abdi-Ali, A.; Mohammadi-Mehr, M.; Alaei, Y.A. Bactericidal activity of various antibiotics against biofilm-producing Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2006, 27, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, H.; Charron-Mazenod, L.; Lewenza, S. Extracellular DNA Chelates Cations and Induces Antibiotic Resistance in Pseudomonas aeruginosa Biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef]
- Anderl, J.N.; Franklin, M.J.; Stewart, P.S. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob. Agents Chemother. 2000, 44, 1818–1824. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Antimicrob. Chemother. 2010, 65, 1955–1958. [Google Scholar] [CrossRef]
- Bellich, B.; Lagatolla, C.; Tossi, A.; Benincasa, M.; Cescutti, P.; Rizzo, R. Influence of Bacterial Biofilm Polysaccharide Structure on Interactions with Antimicrobial Peptides: A Study on. Int. J. Mol. Sci. 2018, 19, 1685. [Google Scholar] [CrossRef]
- Gordon, V.; Bakhtiari, L.; Kovach, K. From molecules to multispecies ecosystems: The roles of structure in bacterial biofilms. Phys. Biol. 2019, 16, 041001. [Google Scholar] [CrossRef]
- Balducci, E.; Papi, F.; Capialbi, D.E.; Del Bino, L. Polysaccharides’ Structures and Functions in Biofilm Architecture of Antimicrobial-Resistant (AMR) Pathogens. Int. J. Mol. Sci. 2023, 24, 4030. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.P.; França, Â.; Pereira, M.O.; Cerca, N. RNA-based qPCR as a tool to quantify and to characterize dual-species biofilms. Sci. Rep. 2019, 9, 13639. [Google Scholar] [CrossRef]
- Jadooa, M.M.; Al-Mousawi, H.T.M.; Al-bderee, N.M.H. Evaluation of the effects of ZnO NPs biosynthesis on Gene Expression for Hemolysin Hla Gene of S. aureus Bacteria Isolated from Diabetic Foot Ulcer. HIV Nurs. 2023, 23, 358–363. [Google Scholar]
- Speziale, P.; Pietrocola, G. The Multivalent Role of Fibronectin-Binding Proteins A and B (FnBPA and FnBPB) of. Front. Microbiol. 2020, 11, 2054. [Google Scholar] [CrossRef]
- Fluckiger, U.; Ulrich, M.; Steinhuber, A.; Döring, G.; Mack, D.; Landmann, R.; Goerke, C.; Wolz, C. Biofilm formation, icaADBC transcription, and polysaccharide intercellular adhesin synthesis by staphylococci in a device-related infection model. Infect. Immun. 2005, 73, 1811–1819. [Google Scholar] [CrossRef]
- Chew, S.C.; Kundukad, B.; Seviour, T.; van der Maarel, J.R.C.; Yang, L.; Rice, S.A.; Doyle, P.; Kjelleberg, S. Dynamic Remodeling of Microbial Biofilms by Functionally Distinct Exopolysaccharides. Mbio 2014, 5, e01536-14. [Google Scholar] [CrossRef] [PubMed]
- Baidamshina, D.R.; Trizna, E.Y.; Holyavka, M.G.; Bogachev, M.I.; Artyukhov, V.G.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Kayumov, A.R. Targeting microbial biofilms using Ficin, a nonspecific plant protease. Sci. Rep. 2017, 7, 46068. [Google Scholar] [CrossRef]
- Kayumov, A.R.; Nureeva, A.A.; Trizna, E.Y.; Gazizova, G.R.; Bogachev, M.I.; Shtyrlin, N.V.; Pugachev, M.V.; Sapozhnikov, S.V.; Shtyrlin, Y.G. New Derivatives of Pyridoxine Exhibit High Antibacterial Activity against Biofilm-Embedded Staphylococcus Cells. Biomed. Res. Int. 2015, 10, 890968. [Google Scholar] [CrossRef]
- Kayumov, A.R.; Khakimullina, E.N.; Sharafutdinov, I.S.; Trizna, E.Y.; Latypova, L.Z.; Lien, H.T.; Margulis, A.B.; Bogachev, M.I.; Kurbangalieva, A.R. Inhibition of biofilm formation in Bacillus subtilis by new halogenated furanones. J. Antibiot. 2015, 68, 297–301. [Google Scholar] [CrossRef]
- Leclercq, R.; Canton, R.; Brown, D.F.J.; Giske, C.G.; Heisig, P.; MacGowan, A.P.; Mouton, J.W.; Nordmann, P.; Rodloff, A.C.; Rossolini, G.M.; et al. EUCAST expert rules in antimicrobial susceptibility testing. Clin. Microbiol. Infect. 2013, 19, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005, 1, 1B.1. [Google Scholar] [CrossRef]
- Lafi, R.; Montasser, I.; Hafiane, A. Adsorption of congo red dye from aqueous solutions by prepared activated carbon with oxygen-containing functional groups and its regeneration. Adsorpt. Sci. Technol. 2019, 37, 160–181. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2001, 44, 121–129. [Google Scholar] [CrossRef]
- Gomes, A.I.; Stuchi, L.P.; Siqueira, N.M.G.; Henrique, J.B.; Vicentini, R.; Ribeiro, M.L.; Darrieux, M.; Ferraz, L.F.C. Selection and validation of reference genes for gene expression studies in Klebsiella pneumoniae using Reverse Transcription Quantitative real-time PCR. Sci. Rep. 2018, 8, 9001. [Google Scholar] [CrossRef]
- Liu, M.; Wu, X.; Li, J.; Liu, L.; Zhang, R.-g.; Shao, D.; Du, X. The specific anti-biofilm effect of gallic acid on Staphylococcus aureus by regulating the expression of the ica operon. Food Control 2017, 73, 613–618. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. aureus | K. pneumoniae | |||
|---|---|---|---|---|
| MIC | MBC | MIC | MBC | |
| Amikacin | 8 | 32 | 4 | 4 |
| Ceftazidime | 8 | 32 | 32 | 256 |
| Ciprofloxacin | 0.25 | 16 | 0.5 | 0.5 |
| Ampicillin | 0.25 | 2 | ND | ND |
| Vancomycin | 2 | 64 | ND | ND |
| Dye | SYBR Green | Sypro Orange | ConA-TMR | CFW |
|---|---|---|---|---|
| Excitation Wavelength | 497 nm | 470 nm | 552 nm | 254 nm |
| Emission Wavelength | 520 nm | 570 nm | 578 nm | 432 nm |
| Target | eDNA | Proteins | α-polysaccharides | β-polysaccharides |
| Primer | Nucleotide Sequence | Reference |
|---|---|---|
| Kp-pgaA-for | 5′ CACCTGCAGACGCTCTCCTATGTC 3′ | This work |
| Kp-pgaA-rev | 5′ AAGAGGAGATGACCCAGCCGATG 3′ | This work |
| proC-for | 5′ GATTGCCGATATCGTCTTCG 3′ | [60] |
| proC-rev | 5′ GAGACCACCAGCGACTCTTT 3′ | [60] |
| icaA-for | 5′ AAGCCAACGCACTCAATCAAGG 3′ | [61] |
| icaA-rev | 5′ GGATTACCTGTAACCGCACCAAG 3′ | [61] |
| 16s rRNA-for | 5′ GGGACCCGCACAAGCGGTGG 3′ | [61] |
| 16s rRNA-rev | 5′ GGGTTGCGCTCGTTGCGGGA 3′ | [61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mironova, A.V.; Karimova, A.V.; Bogachev, M.I.; Kayumov, A.R.; Trizna, E.Y. Alterations in Antibiotic Susceptibility of Staphylococcus aureus and Klebsiella pneumoniae in Dual Species Biofilms. Int. J. Mol. Sci. 2023, 24, 8475. https://doi.org/10.3390/ijms24108475
Mironova AV, Karimova AV, Bogachev MI, Kayumov AR, Trizna EY. Alterations in Antibiotic Susceptibility of Staphylococcus aureus and Klebsiella pneumoniae in Dual Species Biofilms. International Journal of Molecular Sciences. 2023; 24(10):8475. https://doi.org/10.3390/ijms24108475
Chicago/Turabian StyleMironova, Anna V., Agniya V. Karimova, Mikhail I. Bogachev, Airat R. Kayumov, and Elena Y. Trizna. 2023. "Alterations in Antibiotic Susceptibility of Staphylococcus aureus and Klebsiella pneumoniae in Dual Species Biofilms" International Journal of Molecular Sciences 24, no. 10: 8475. https://doi.org/10.3390/ijms24108475
APA StyleMironova, A. V., Karimova, A. V., Bogachev, M. I., Kayumov, A. R., & Trizna, E. Y. (2023). Alterations in Antibiotic Susceptibility of Staphylococcus aureus and Klebsiella pneumoniae in Dual Species Biofilms. International Journal of Molecular Sciences, 24(10), 8475. https://doi.org/10.3390/ijms24108475