
Dual-Species Biofilms: Biomass, Viable Cell Ratio/Cross-Species Interactions, Conjugative Transfer
 , and
, and
Abstract
:1. Introduction
2. Results
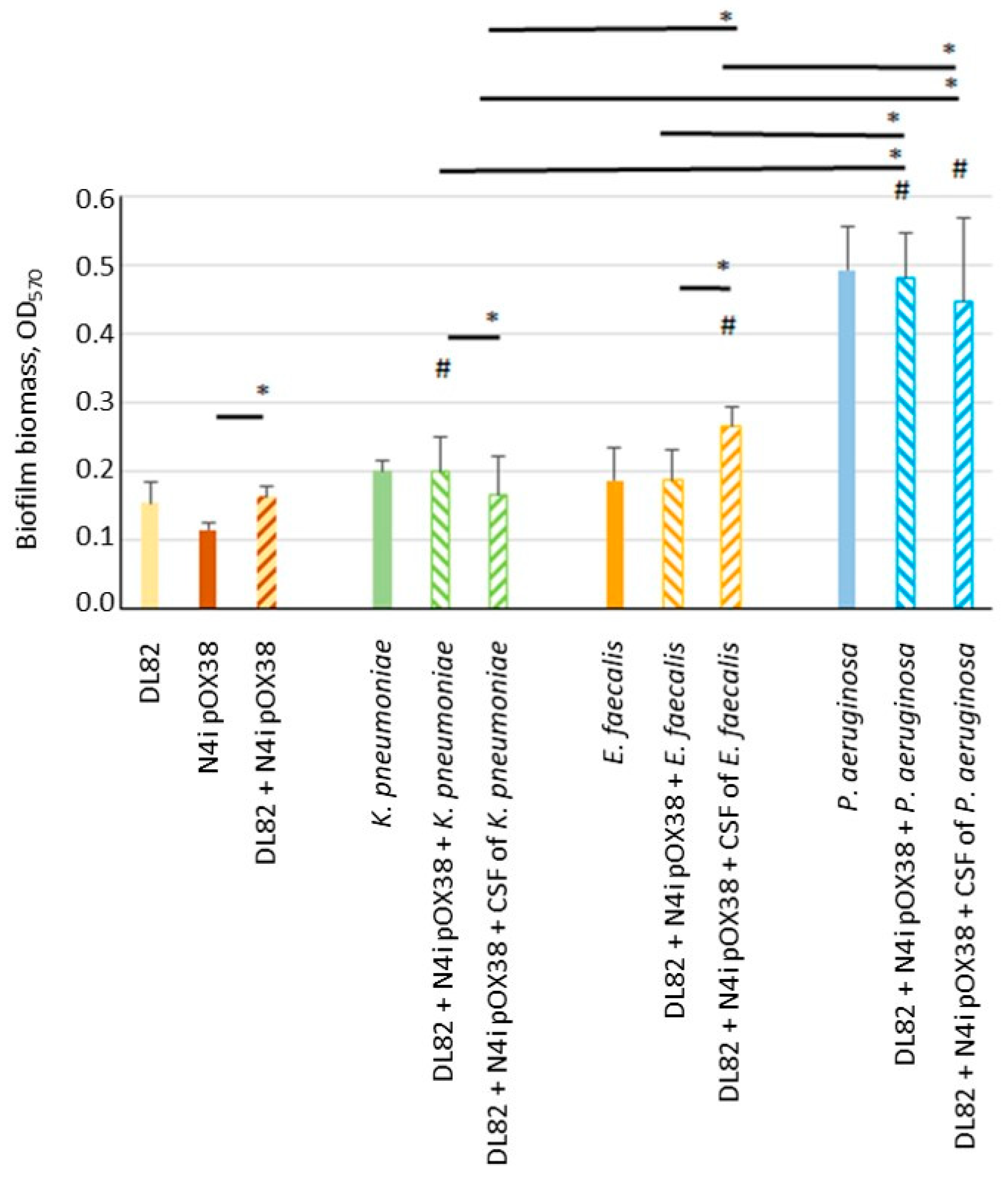
2.1. Biofilm Biomass
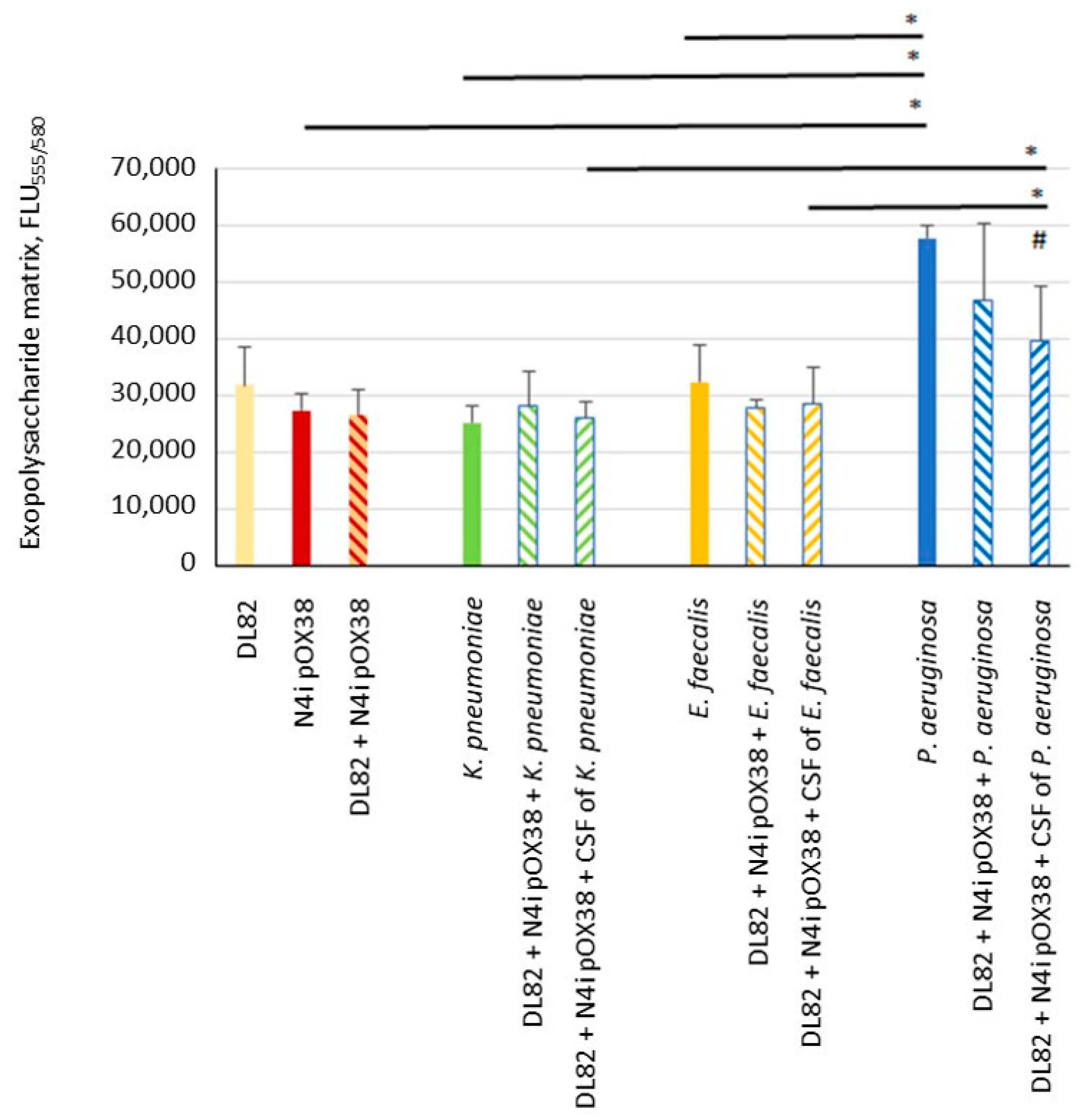
2.2. Biofilm Exopolysaccharide Matrix
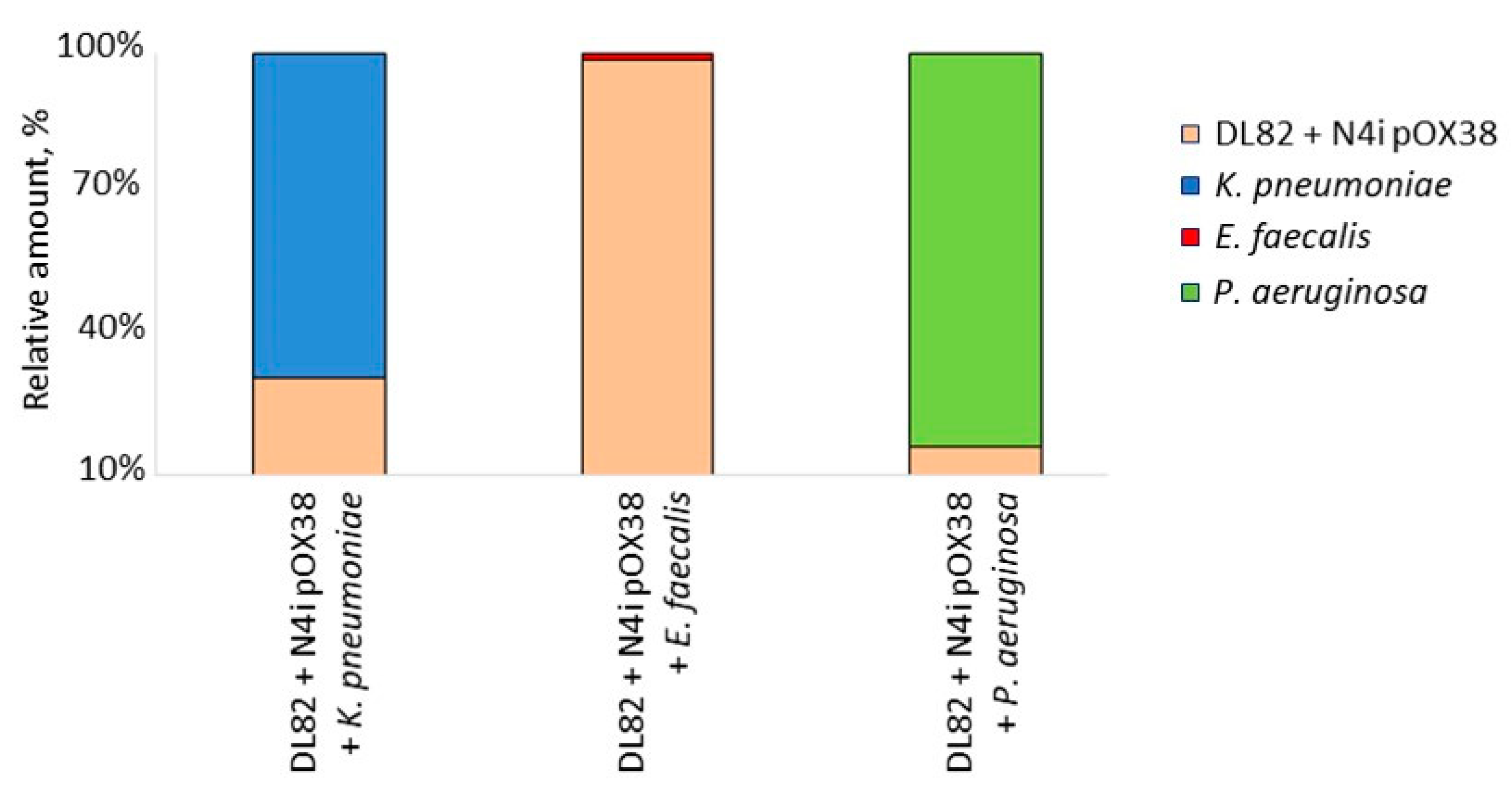
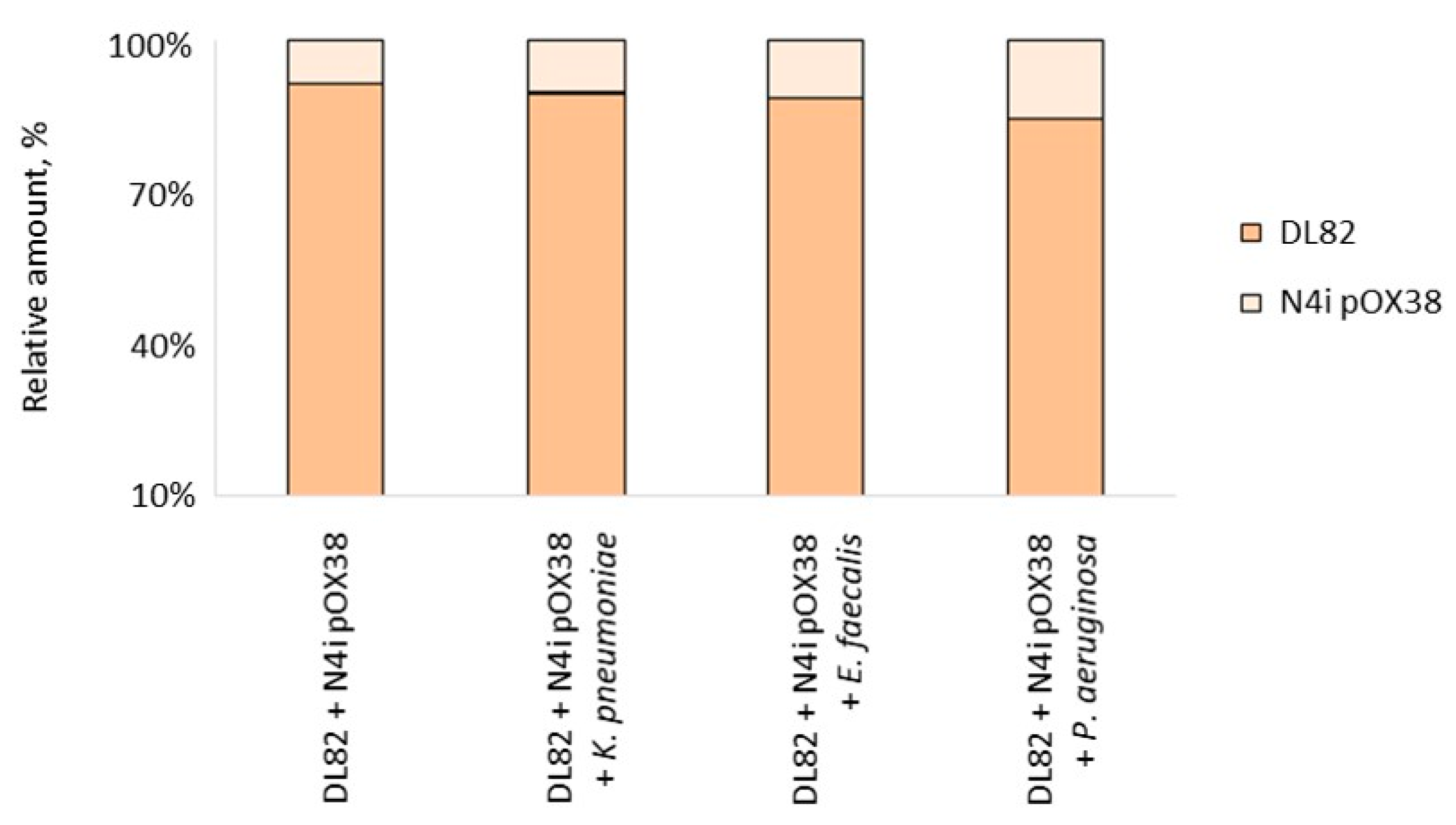
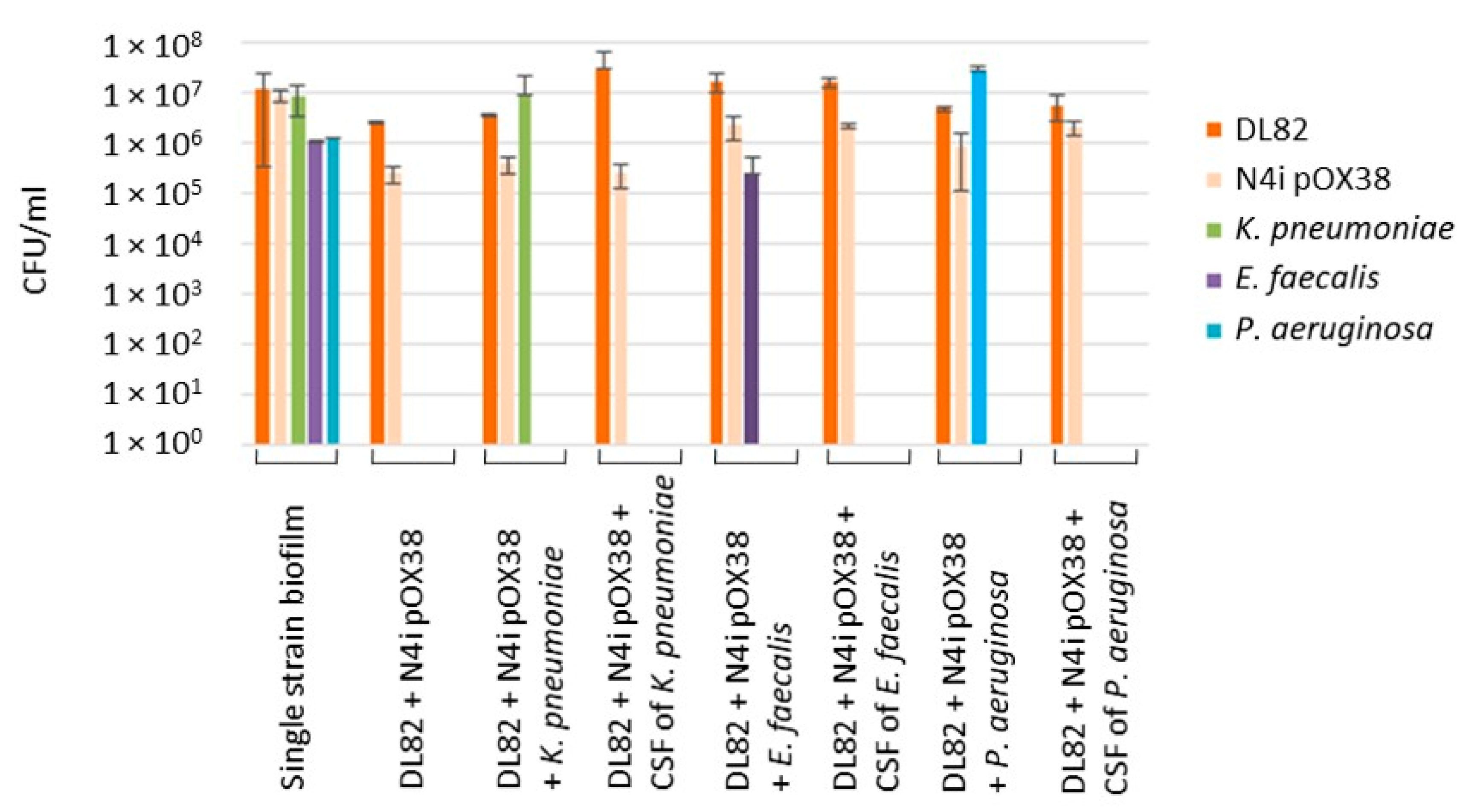
2.3. The Viability of Recipient E. coli DL82, Donor E. coli N4i pOX38 and Opportunistic Pathogenic Bacteria within Dual-Species Biofilms
2.4. The Frequency of Conjugation within Dual-Species Biofilm
2.5. AI-2 Activity in CFSs of Conjugation Mixtures
3. Discussion
4. Materials and Methods
4.1. Strains and Media
4.2. Cell-Free Supernatant (CFS)
4.3. Conjugation Assay
4.4. Biofilm Biomass Analysis
4.5. Biofilm Exopolysaccharide Matrix Analysis
4.6. AI-2 Production Assay
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Besemer, K. Biodiversity, community structure and function of biofilms in stream ecosystems. Res. Microbiol. 2015, 10, 774–781. [Google Scholar] [CrossRef]
- Römling, U.; Kjelleberg, S.; Normark, S.; Nyman, L.; Uhlin, B.E.; Åkerlund, B. Microbial biofilm formation: A need to act. J. Intern. Med. 2014, 276, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Mizan, M.F.; Jahid, I.K.; Ha, S.D. Microbial biofilms in seafood: A food-hygiene challenge. Food Microbiol. 2015, 49, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Larsen, L.H.; Lorenzen, J.; Hall-Stoodley, L.; Kikhney, J.; Moter, A.; Thomsen, T.R. Microbiological diagnosis of device-related biofilm infections. APMIS 2017, 125, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.M.; Pracser, N.; Thalguter, S.; Fischel, K.; Rammer, N.; Pospíšilová, L.; Alispahic, M.; Wagner, M.; Rychli, K. Identification of biofilm hotspots in a meat processing environment: Detection of spoilage bacteria in multi-species biofilms. Int. J. Food Microbiol. 2020, 328, 108668. [Google Scholar] [CrossRef]
- McLean, R.J.; Kakirde, K.S. Enhancing metagenomics investigations of microbial interactions with biofilm technology. Int. J. Mol. Sci. 2013, 14, 22246–22257. [Google Scholar] [CrossRef]
- Schulze, A.; Mitterer, F.; Pombo, J.P.; Schild, S. Biofilms by bacterial human pathogens: Clinical relevance—Development, composition and regulation—Therapeutical strategies. Microb. Cell. 2021, 8, 28–56. [Google Scholar] [CrossRef]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Marra, M.; Zummo, S.; Biondo, C. Urinary tract infections: The current scenario and future prospects. Pathogens 2023, 12, 623. [Google Scholar] [CrossRef]
- Lila, A.S.A.; Rajab, A.A.H.; Abdallah, M.H.; Rizvi, S.M.D.; Moin, A.; Khafagy, E.S.; Tabrez, S.; Hegazy, W.A.H. Biofilm lifestyle in recurrent urinary tract infections. Life 2023, 13, 148. [Google Scholar] [CrossRef]
- Oliveira, A.; Sousa, J.C.; Silva, A.C.; Melo, L.D.R.; Sillankorva, S. Chestnut honey and bacteriophage application to control Pseudomonas aeruginosa and Escherichia coli biofilms: Evaluation in an ex vivo wound model. Front. Microbiol. 2018, 9, 1725. [Google Scholar] [CrossRef] [PubMed]
- Juarez, G.E.; Galván, E.M. Role of nutrient limitation in the competition between uropathogenic strains of Klebsiella pneumoniae and Escherichia coli in mixed biofilms. Biofouling 2018, 34, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Galván, E.M.; Mateyca, C.; Ielpi, L. Role of interspecies interactions in dual-species biofilms developed in vitro by uropathogens isolated from polymicrobial urinary catheter-associated bacteriuria. Biofouling 2016, 32, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O.; Travier, L.; Latour-Lambert, P.; Fontaine, T.; Magnus, J.; Denamur, E.; Ghigo, J.M. Screening of Escherichia coli species biodiversity reveals new biofilm-associated antiadhesion polysaccharides. MBio 2011, 2, e00043-11. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, H.J.; Ning, S.J.; Gao, Y.L. The response of type 2 quorum sensing in Klebsiella pneumoniae to a fluctuating culture environment. DNA Cell Biol. 2012, 31, 455–459. [Google Scholar] [CrossRef]
- Brito, P.H.; Rocha, E.P.; Xavier, K.B.; Gordo, I. Natural genome diversity of AI-2 quorum sensing in Escherichia coli: Conserved signal production but labile signal reception. Genome Biol. Evol. 2013, 1, 16–30. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Wang, Z.; Fu, Y.; Ai, Q.; Dong, Y.; Yu, J. Autoinducer-2 regulates Pseudomonas aeruginosa PAO1 biofilm formation and virulence production in a dose-dependent manner. BMC Microbiol. 2015, 15, 192. [Google Scholar] [CrossRef]
- Yang, Y.; Li, W.; Hou, B.; Zhang, C. Quorum sensing LuxS/autoinducer-2 inhibits Enterococcus faecalis biofilm formation ability. J. Appl. Oral. Sci. 2018, 26, e20170566. [Google Scholar] [CrossRef]
- Wang, Y.M.; Dong, W.L.; Odah, K.A.; Kong, L.C.; Ma, H.X. Transcriptome analysis reveals AI-2 relevant genes of multi-drug resistant Klebsiella pneumoniae in response to Eugenol at sub-MIC. Front. Microbiol. 2019, 10, 1159. [Google Scholar] [CrossRef]
- Chen, L.; Wilksch, J.J.; Liu, H.; Zhang, X.; Torres, V.V.L.; Bi, W.; Mandela, E.; Cao, J.; Li, J.; Lithgow, T.; et al. Investigation of LuxS-mediated quorum sensing in Klebsiella pneumoniae. J. Med. Microbiol. 2020, 69, 402–413. [Google Scholar] [CrossRef]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signaling in bacteria. FEMS Microbiol. Rev. 2013, 37, 156–181. [Google Scholar] [CrossRef] [PubMed]
- Laganenka, L.; Sourjik, V. Autoinducer 2-dependent Escherichia coli biofilm formation is enhanced in a dual-species coculture. Appl. Environ. Microbiol. 2018, 84, e02638-17. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, Y.; Wu, H.; Hóiby, N.; Molin, S.; Song, Z.J. Current understanding of multi-species biofilms. Int. J. Oral. Sci. 2011, 3, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.; Banin, E. Multi-species biofilms: Living with friendly neighbors. FEMS Microbiol Rev. 2012, 36, 990–1004. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, O.; Ghigo, J.M. Multi-species biofilms: How to avoid unfriendly neighbors. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef] [PubMed]
- Antonova, E.S.; Hammer, B.K. Quorum-sensing autoinducer molecules produced by members of a multispecies biofilm promote horizontal gene transfer to Vibrio cholerae. FEMS Microbiol. Lett. 2011, 1, 68–76. [Google Scholar] [CrossRef]
- Hola, V.; Ruzicka, F. The Formation of Poly-Microbial Biofilms on Urinary Catheters. In Urinary Tract Infections, 2nd ed.; Tenke, P., Ed.; Rendelőintézet: Budapest, Hungary, 2011; pp. 153–172. [Google Scholar] [CrossRef]
- Reisner, A.; Höller, B.M.; Molin, S.; Zechner, E.L. Synergistic effects in mixed Escherichia coli biofilms: Conjugative plasmid transfer drives biofilm expansion. J. Bacteriol. 2006, 188, 3582–3588. [Google Scholar] [CrossRef]
- Koraimann, G. Spread and persistence of virulence and antibiotic resistance genes: A ride on the F plasmid conjugation module. EcoSal Plus 2018, 8, 1–23. [Google Scholar] [CrossRef]
- Stephens, C.; Arismendi, T.; Wright, M.; Hartman, A.; Gonzalez, A.; Gill, M.; Pandori, M.; Hess, D. F Plasmids are the major carriers of antibiotic resistance genes in human-associated commensal Escherichia coli. mSphere 2020, 5, e00709-20. [Google Scholar] [CrossRef]
- Kuznetsova, M.V.; Maslennikova, I.L.; Pospelova, J.S.; Žgur Bertok, D.; Starčič Erjavec, M. Differences in recipient ability of uropathogenic Escherichia coli strains in relation with their pathogenic potential. Infect. Genet. Evol. 2022, 97, 105160. [Google Scholar] [CrossRef]
- Smith, R.S.; Iglewski, B.H. P. aeruginosa quorum-sensing systems and virulence. Curr. Opin. Microbiol. 2003, 6, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Laganenka, L.; Lee, J.W.; Malfertheiner, L.; Dieterich, C.L.; Fuchs, L.; Piel, J.; von Mering, C.; Sourjik, V.; Hardt, W.D. Chemotaxis and autoinducer-2 signalling mediate colonization and contribute to co-existence of Escherichia coli strains in the murine gut. Nat. Microbiol. 2023, 2, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Tchebotar, I.V.; Mayanskiy, A.N.; Mayanskiy, N.A. Matrix of microbial biofilms. Clin. Microbiol. Antimicrobial. Chemother. 2016, 18, 9–19. (In Russian) [Google Scholar]
- Sørensen, S.J.; Bailey, M.; Hansen, L.H.; Kroer, N.; Wuertz, S. Studying plasmid horizontal transfer in situ: A critical review. Nat. Rev. Microbiol. 2005, 3, 700–710. [Google Scholar] [CrossRef]
- Król, J.E.; Wojtowicz, A.J.; Rogers, L.M.; Heuer, H.; Smalla, K.; Krone, S.M.; Top, E.M. Invasion of E. coli biofilms by antibiotic resistance plasmids. Plasmid 2013, 70, 110–119. [Google Scholar] [CrossRef]
- Cook, L.C.; Dunny, G.M. The influence of biofilms in the biology of plasmids. Microbiol Spectr. 2014, 2, 0012. [Google Scholar] [CrossRef]
- Ferrières, L.; Hancock, V.; Klemm, P. Specific selection for virulent urinary tract infectious Escherichia coli strains during catheter-associated biofilm formation. FEMS Immunol. Med. Microbiol. 2007, 51, 212–219. [Google Scholar] [CrossRef]
- Keogh, D.; Tay, W.H.; Ho, Y.Y.; Dale, J.L.; Chen, S.; Umashankar, S.; Williams, R.B.H.; Chen, S.L.; Dunny, G.M.; Kline, K.A. Enterococcal Metabolite Cues Facilitate Interspecies Niche Modulation and Polymicrobial Infection. Cell Host Microbe 2016, 20, 493–503. [Google Scholar] [CrossRef]
- Lopes, S.P.; Machado, I.; Pereira, M.O. Role of planktonic and sessile extracellular metabolic byproducts on Pseudomonas aeruginosa and Escherichia coli intra and interspecies relationships. J. Ind. Microbiol. Biotechnol. 2011, 38, 133–140. [Google Scholar] [CrossRef]
- Machado, I.; Lopes, S.P.; Sousa, A.M.; Pereira, M.O. Adaptive response of single and binary Pseudomonas aeruginosa and Escherichia coli biofilms to benzalkonium chloride. J. Basic Microbiol. 2012, 52, 43–52. [Google Scholar] [CrossRef]
- Cerqueira, L.; Oliveira, J.A.; Nicolau, A.; Azevedo, N.F.; Vieira, M.J. Biofilm formation with mixed cultures of Pseudomonas aeruginosa/Escherichia coli on silicone using artificial urine to mimic urinary catheters. Biofouling 2013, 29, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Castonguay, M.H.; van der Schaaf, S.; Koester, W.; Krooneman, J.; van der Meer, W.; Harmsen, H.; Landini, P. Biofilm formation by Escherichia coli is stimulated by synergistic interactions and co-adhesion mechanisms with adherence-proficient bacteria. Res. Microbiol. 2006, 5, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, M.V.; Maslennikova, I.L.; Karpunina, T.I.; Nesterova, L.Y.; Demakov, V.A. Interactions of Pseudomonas aeruginosa in predominant biofilm or planktonic forms of existence in mixed culture with Escherichia coli in vitro. Can. J. Microbiol. 2013, 59, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Gritsenko, V.A.; Mrugova, T.M.; Kurlayev, P.P.; Belozertseva, Y.P.; Borisov, S.D. Antagonistic relationship Pseudomonas aeruginosa with gram-negative bacteria. Bull. OSC 2016, 4, 1–5. (In Russian) [Google Scholar]
- González Barrios, A.F.; Zuo, R.; Hashimoto, Y.; Yang, L.; Bentley, W.E.; Wood, T.K. Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum-sensing regulator (MqsR, B3022). J. Bacteriol. 2006, 1, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Goh, S.G.; You, L.; Yuan, Q.; Mohapatra, S.; Gin, K.Y.; Chen, B. Low concentration quaternary ammonium compounds promoted antibiotic resistance gene transfer via plasmid conjugation. Sci. Total Environ. 2023, 887, 163781. [Google Scholar] [CrossRef]
- Cho, J.; Jenneson, S.; Lane, M.; Macfadyen, A.; Van Rietschoten, S. The effects of altering autoinducer-2 concentration on transfer efficiencies of the F and RPI plasmids to the Quorum sensing recipient Escherichia coli strain AB 1157. JEMI 2003, 3, 8–14. [Google Scholar]
- Rijavec, M.; Rijavec, M.; Starčič Erjavec, M.; Ambrožič Avguštin, J.; Reissbrodt, R.; Fruth, A.; Križan-Hergouth, V.; Žgur-Bertok, D. High prevalence of multidrug resistance and random distribution of mobile genetic elements among uropathogenic Escherichia coli (UPEC) of the four major phylogenetic groups. Curr. Microbiol. 2006, 53, 158–162. [Google Scholar] [CrossRef]
- Starčič Erjavec, M.; Petkovšek, Ž.; Kuznetsova, M.V.; Maslennikova, I.L.; Žgur-Bertok, D. Strain ŽP—The first bacterial conjugation-based “kill”-“anti-kill” antimicrobial system. Plasmid 2015, 82, 28–34. [Google Scholar] [CrossRef]
- Bassler, B.L.; Greenberg, E.P.; Stevens, A.M. Cross-species induction of luminescence in the quorum-sensing bacterium Vibrio harveyi. J. Bacteriol. 1997, 179, 4043–4045. [Google Scholar] [CrossRef]
- Guglielmetti, E.; Korhonen, J.M.; Heikkinen, J.; Morelli, L.; Von Wright, A. Transfer of plasmid-mediated resistance to tetracycline in pathogenic bacteria from fish and aquaculture environments. FEMS Microbiol. Lett. 2009, 293, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005. [Google Scholar] [CrossRef]
- Surette, M.G.; Bassler, B.L. Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1998, 95, 7046–7050. [Google Scholar] [CrossRef]
- Zorina, A.S.; Maksimova, Y.G.; Demakov, V.A. Biofilm formation by monocultures and mixed cultures of Alcaligenes faecalis 2 and Rhodococcus ruber GT 1. Microbiology 2019, 88, 164–171. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conjugation Mixture | № | Frequency of Conjugation | |
|---|---|---|---|
| R + D + Opportunistic Pathogenic Bacteria | R + D + CFSs of Opportunistic Pathogenic Bacteria | ||
| E. coli DL82 + E. coli N4i pOX38 | 1 | 4.72 × 10−4 ± 2.28 × 10−4 | |
| E. coli DL82 + E. coli N4i pOX38+ K. pneumoniae | 2 | 2.69 × 10−4 ± 1.01 × 10−4 | 4.93 × 10−5 ± 3.66 × 10−5 P1–2 = 0.0495 1 |
| E. coli DL82 + E. coli N4i pOX38 + E. faecalis | 3 | 1.78 × 10−5 ± 5.38 × 10−6 P1–3 = 0.0495 P2–3 = 0.0495 | 1.93 × 10−5 ± 4.17 × 10−7 P1–3 = 0.0495 |
| E. coli DL82 + E. coli N4i pOX38+ P. aeruginosa | 4 | 2.93 × 10−5 ± 3.07 × 10−5 P1–4 = 0.0495 P2–4 = 0.0495 | 0.00 × 100 P1–4 = 0.0495 P2–4 = 0.0495 P3–4 = 0.0495 |
| Cell-Free Supernatant (CSF) of Strains/Conjugation Mixtures | № | Induction of Luminescence, % |
|---|---|---|
| V. harveyi BB152 | 1 | 100.0 |
| E. coli K12 | 2 | 386.8 ± 42.9 P1–2 = 0.012 1 |
| E. coli DL82 | 3 | 3.2 ± 0.7 |
| E. coli N4i pOX38 | 4 | 1.3 ± 0.2 |
| E. coli DL82 + E. coli N4i pOX38 | 5 | 73.7 ± 35.5 P3–5 = 0.027 P4–5 = 0.016 |
| K. pneumoniae | 6 | 190.1 ± 57.9 P3–5 = 0.023 |
| E. coli DL82 + E. coli N4i pOX38+ K. pneumoniae | 7 | 77.4 ± 42.3 |
| E. coli DL82 + E. coli N4i pOX38 + CFS of K. pneumoniae | 8 | 96.2 ± 6.3 |
| E. faecalis | 9 | 0.1 ± 0.05 |
| E. coli DL82+ E. coli N4i pOX38 + E. faecalis | 10 | 0.7 ± 0.1 P5–10 = 0.027 |
| E. coli DL82+ E. coli N4i pOX38 + CFS of E. faecalis | 11 | 37.0 ± 4.7 P9–11 = 0.005 |
| P. aeruginosa | 12 | 1.5 ± 1.0 P2–12 = 0.010 |
| E. coli DL82 + E. coli N4i pOX38+ P. aeruginosa | 13 | 23.5 ± 14.6 P5–13 = 0.040 |
| E. coli DL82 + E. coli N4i pOX38+ CFS of P. aeruginosa | 14 | 29.4 ± 8.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuznetsova, M.V.; Pospelova, J.S.; Maslennikova, I.L.; Starčič Erjavec, M. Dual-Species Biofilms: Biomass, Viable Cell Ratio/Cross-Species Interactions, Conjugative Transfer. Int. J. Mol. Sci. 2023, 24, 14497. https://doi.org/10.3390/ijms241914497
Kuznetsova MV, Pospelova JS, Maslennikova IL, Starčič Erjavec M. Dual-Species Biofilms: Biomass, Viable Cell Ratio/Cross-Species Interactions, Conjugative Transfer. International Journal of Molecular Sciences. 2023; 24(19):14497. https://doi.org/10.3390/ijms241914497
Chicago/Turabian StyleKuznetsova, Marina V., Julia S. Pospelova, Irina L. Maslennikova, and Marjanca Starčič Erjavec. 2023. "Dual-Species Biofilms: Biomass, Viable Cell Ratio/Cross-Species Interactions, Conjugative Transfer" International Journal of Molecular Sciences 24, no. 19: 14497. https://doi.org/10.3390/ijms241914497
APA StyleKuznetsova, M. V., Pospelova, J. S., Maslennikova, I. L., & Starčič Erjavec, M. (2023). Dual-Species Biofilms: Biomass, Viable Cell Ratio/Cross-Species Interactions, Conjugative Transfer. International Journal of Molecular Sciences, 24(19), 14497. https://doi.org/10.3390/ijms241914497