
Cytogenetics Meets Genomics: Cytotaxonomy and Genomic Relationships among Color Variants of the Asian Arowana Scleropages formosus

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. SatDNA Content of S. formosus Genome
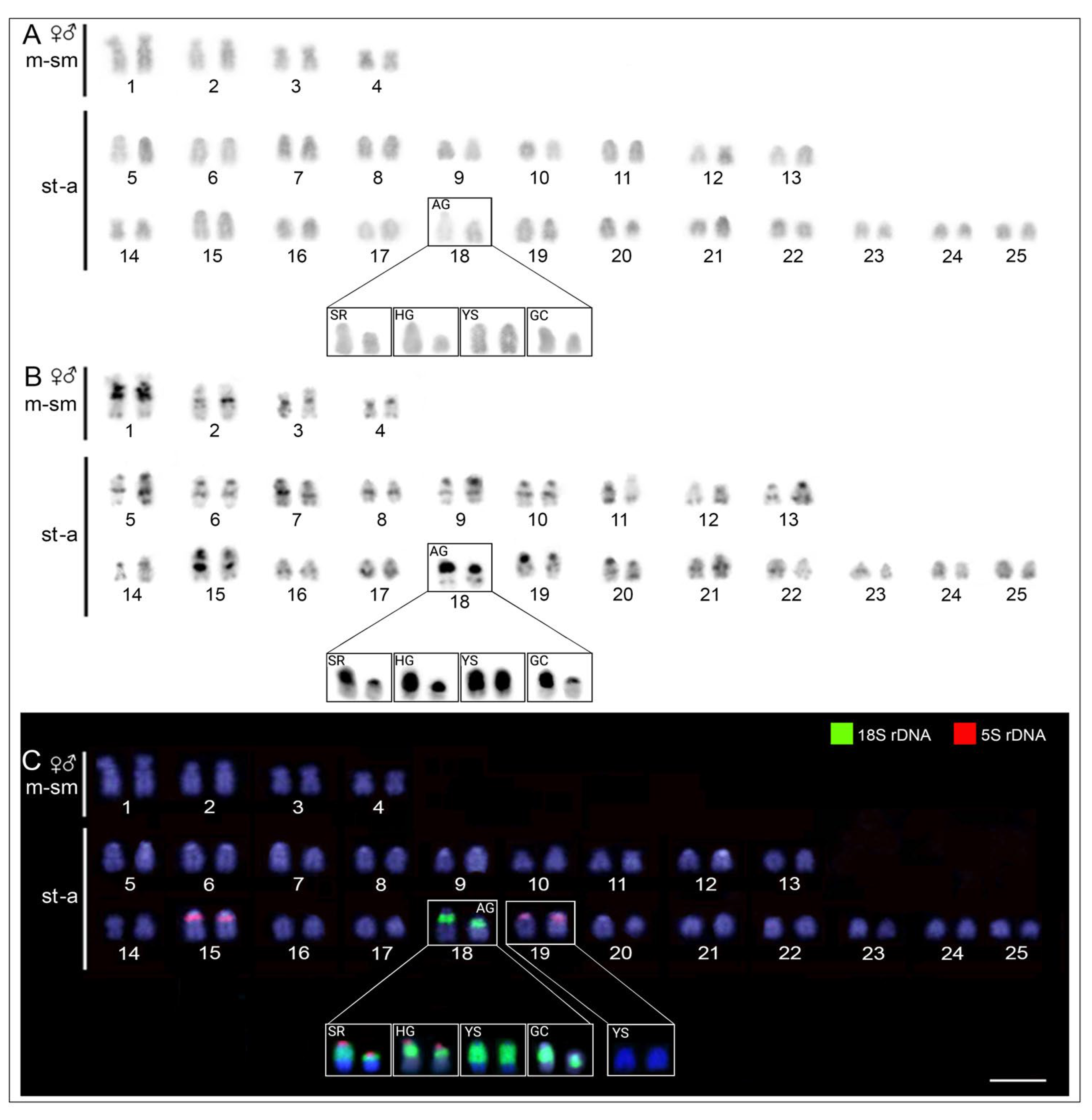
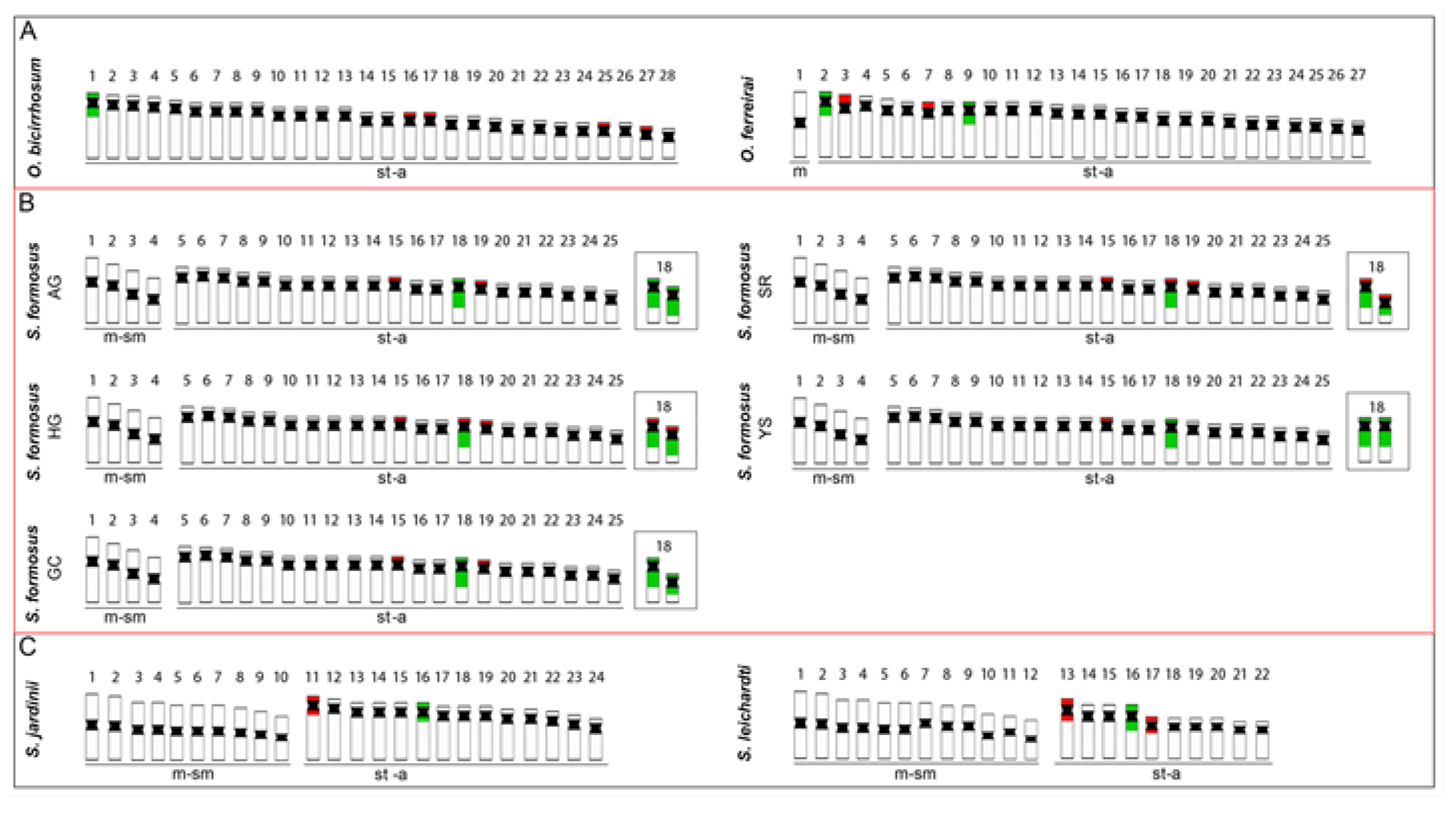
2.2. Karyotypes and C-Banding
2.3. Chromosomal Location of 18S and 5S rDNA
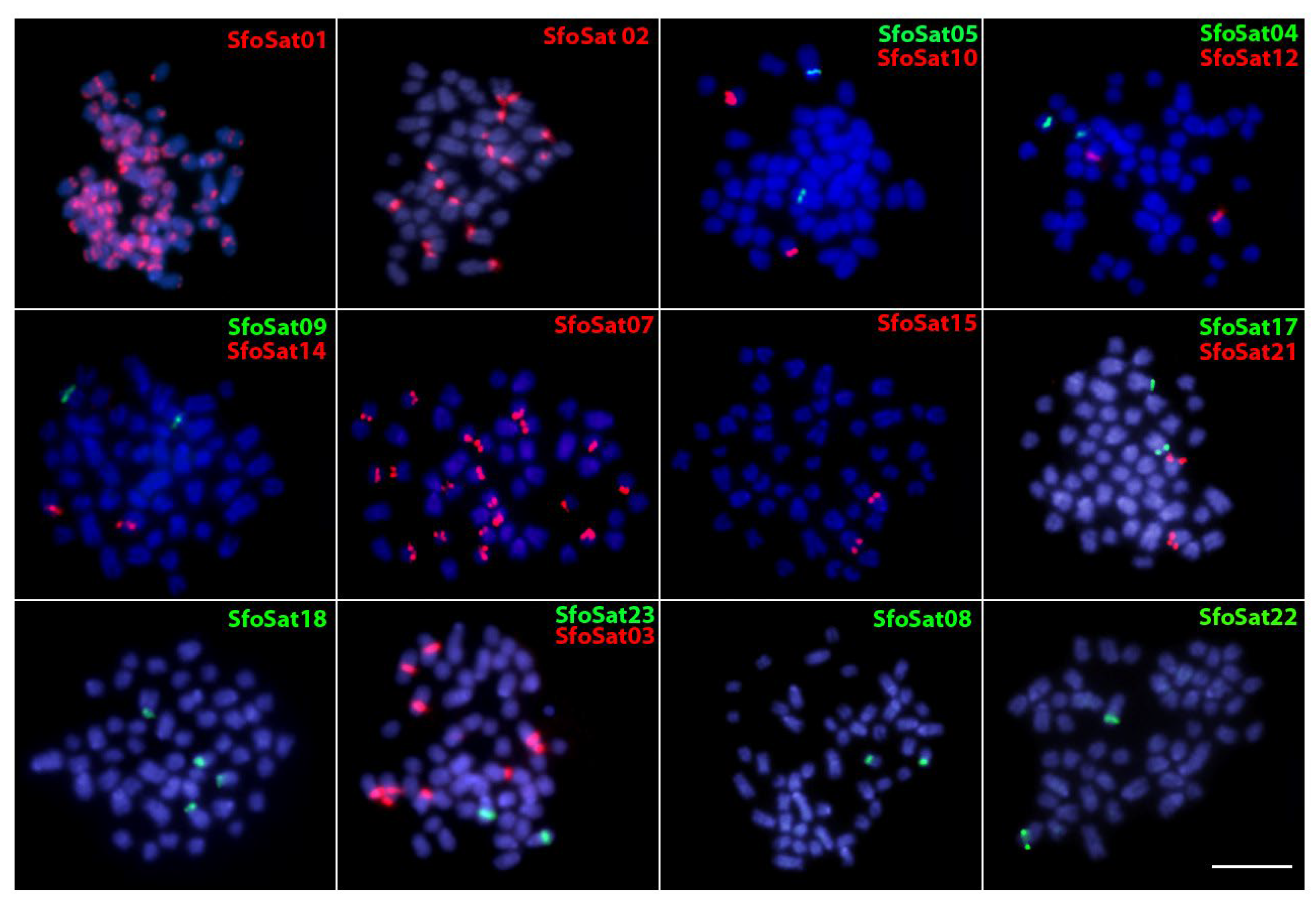
2.4. Chromosomal Location of SatDNAs of S. formosus
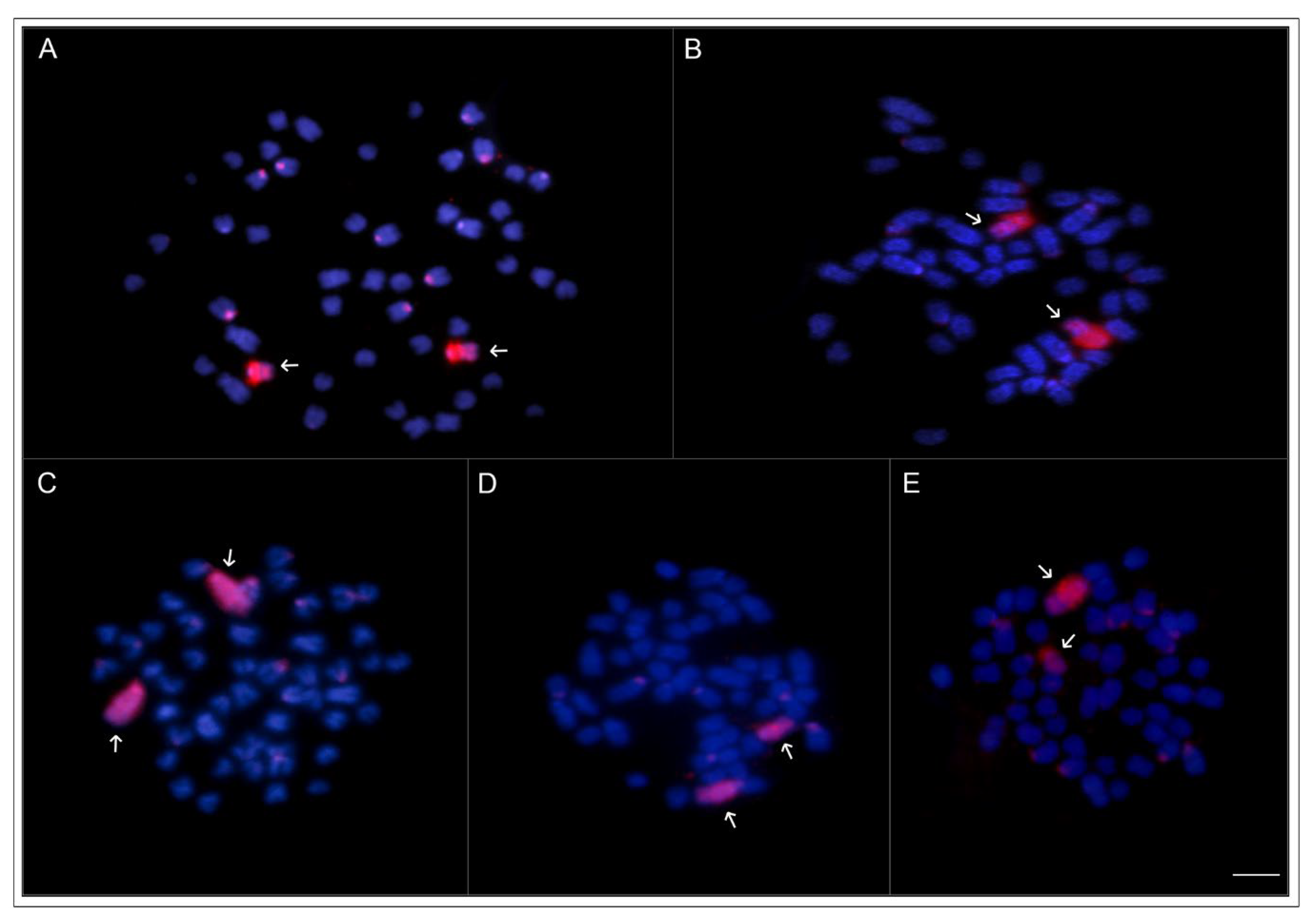
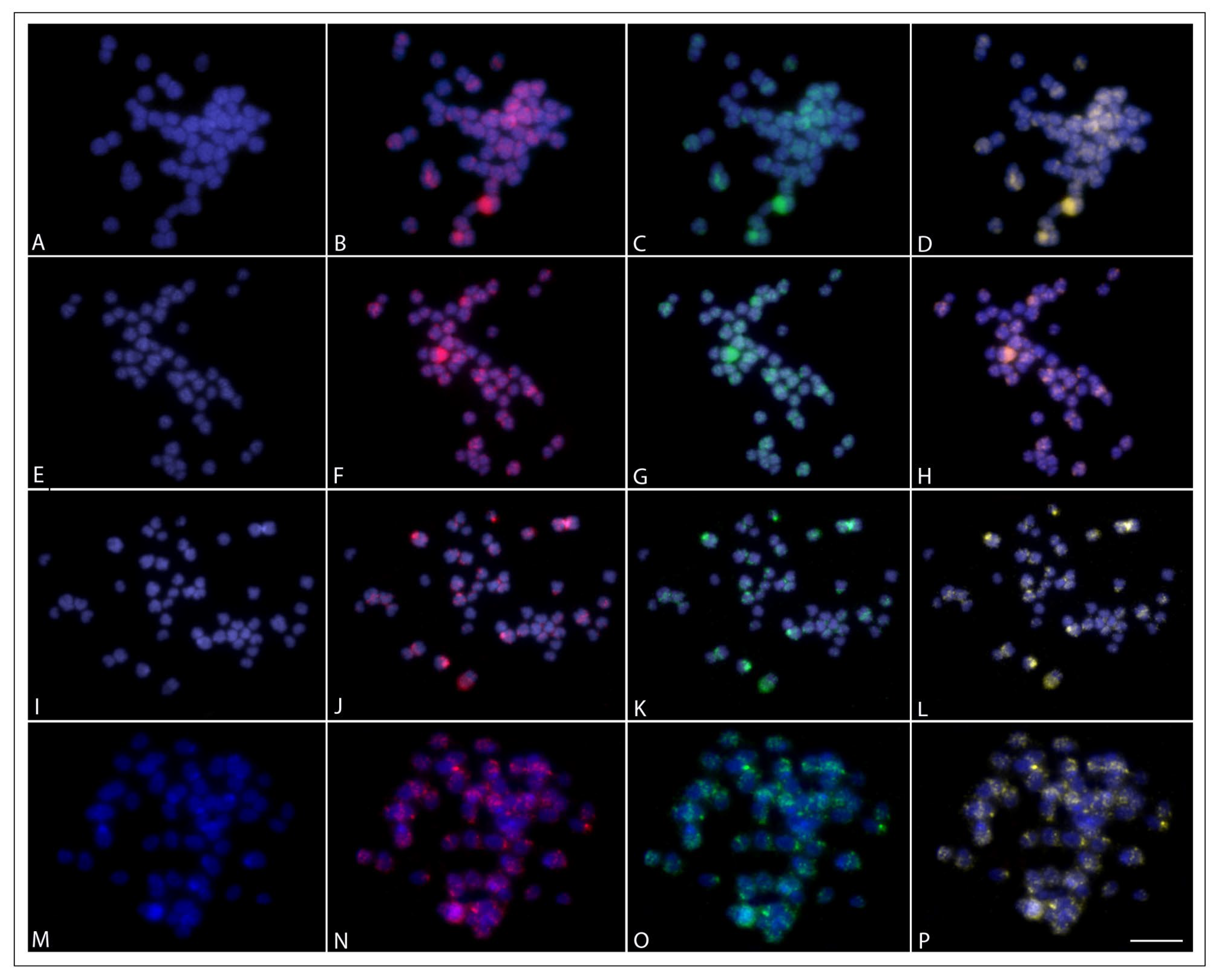
2.5. Whole Chromosome Paint Hybridization (WCP) and Comparative Genomic Hybridization (CGH)
3. Discussion
4. Materials and Methods
4.1. Individuals and Conventional Cytogenetics
4.2. DNA Extraction and Genome Sequencing
4.3. Bioinformatic Analyses
4.4. Primer Design and DNA Amplification via Polymerase Chain Reaction (PCR)
4.5. Fluorescence in Situ Hybridization (FISH)
4.6. Microdissection and Preparation of Chromosome Painting Probes
4.7. Comparative Genomic Hybridization (CGH)
4.8. Microscopy and Image Processing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilton, E.J.; Lavoué, S. A review of the systematic biology of fossil and living bony-tongue fishes, Osteoglossomorpha (Actinopterygii: Teleostei). Neotrop. Ichthyol. 2018, 16, 1–35. [Google Scholar] [CrossRef]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; Wiley: Hoboken, NJ, USA, 2016. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Eschemeyer’s Catalog of Fishes: Genera, Species, References. 2023. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 10 March 2023).
- Pusey, B.J.; Fisher, C.; Maclaine, J. On the nature of Scleropages leichardti Günther, 1864 (Pisces: Osteoglossidae). Zootaxa 2016, 4173, 75. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.R. Scleropages inscriptus, a new fish species from the Tananthayi or Tenasserim River basin, Malay Peninsula of Myanmar (Osteoglossidae: Osteoglossiformes). Aqua Int. J. Ichthyol. 2012, 18, 113–118. [Google Scholar]
- Medipally, S.R.; Yusoff, F.M.; Sharifhuddin, N.; Shariff, M. Sustainable aquaculture of Asian arowana—A review. J. Environ. Biol. 2016, 37, 829–838. [Google Scholar]
- Yue, G.H.; Li, Y.; Lim, L.C.; Orban, L. Monitoring the genetic diversity of three Asian arowana (Scleropages formosus) captive stocks using AFLP and microsatellites. Aquaculture 2004, 237, 89–102. [Google Scholar] [CrossRef]
- Larson, H.; Vidthayanon, C. Scleropages formosus. The IUCN Red List of Threatened Species. 2019. pp. 1–10. Available online: https://www.iucnredlist.org/ (accessed on 15 January 2023).
- Mohd-Shamsudin, M.I.; Fard, M.Z.; Mather, P.B.; Suleiman, Z.; Hassan, R.; Othman, R.Y.; Bhassu, S. Molecular characterization of relatedness among colour variants of Asian Arowana (Scleropages formosus). Gene 2011, 490, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.H.; Chang, A.; Yuzer, A.; Suwanto, A. Current Knowledge on the Biology and Aquaculture of the Endangered Asian Arowana. Rev. Fish. Sci. Aquac. 2019, 28, 193–210. [Google Scholar] [CrossRef]
- Lavoué, S. Testing a time hypothesis in the biogeography of the arowana genus Scleropages (Osteoglossidae). J. Biogeogr. 2015, 42, 2427–2439. [Google Scholar] [CrossRef]
- Alshari, N.F.M.A.; Lavoué, S.; Sulaiman, M.A.M.; Khaironizam, M.Z.; Nor, S.A.M.; Aziz, F. Pleistocene paleodrainages explain the phylogeographic structure of Malaysian populations of Asian arowana better than their chromatic variation. Endanger Species Res. 2021, 46, 205–214. [Google Scholar] [CrossRef]
- Kumazawa, Y.; Nishida, M. Molecular phylogeny of osteoglossoids: A new model for Gondwanian origin and plate tectonic transportation of the Asian arowana. Mol. Biol. Evol. 2000, 17, 1869–1878. [Google Scholar] [CrossRef]
- Pouyaud, L.; Sudarto, T.G.; Teugels, G. The different colour varieties of the asian arowana Scleropages formosus (Osteoglossidae) are distinct species: Morphologic and genetic evidences. Cybium 2003, 27, 287–305. [Google Scholar]
- Kottelat, M.; Widjanarti, E. The fishes of Danau Sentarum National Park and the Kapuas Lakes Area, Kalimantan Barat, Indonesia. Raffles Bull. Zool. Suppl. 2005, 13, 139–173. [Google Scholar]
- Kottelat, M. The fishes of the inland waters of Southeast Asia: A catalogue and core bibliography of the fishes known to occur in freshwaters, mangroves and estuaries. Raffles Bull. Zool. 2013, 27, 1–663. [Google Scholar]
- Mu, X.D.; Wang, X.; Song, H.; Yang, Y.; Luo, D.; Gu, D.; Xu, M.; Liu, C.; Luo, J.; Hu, Y. Mitochondrial DNA as effective molecular markers for the genetic variation and phylogeny of the family Osteoglossidae. Gene 2012, 511, 320–325. [Google Scholar] [CrossRef]
- Mu, X.; Wang, X.; Song, H.; Yang, Y.; Luo, D.; Gu, D.; Xu, M.; Liu, C.; Luo, J.; Hu, Y. Genetic diversity and phylogeny of the family Osteoglossidae by the nuclear 18S ribosomal RNA and implications for its conservation. Biochem. Syst. Ecol. 2013, 51, 280–287. [Google Scholar] [CrossRef]
- Barby, F.; Ráb, P.; Lavoué, S.; Ezaz, T.; Bertollo, L.A.C.; Kilian, A.; Maruyama, S.R.; Oliveira, E.A.; Artoni, R.F.; Santos, M.H.; et al. From Chromosomes to Genome: Insights into the Evolutionary Relationships and Biogeography of Old World Knifefishes (Notopteridae; Osteoglossiformes). Genes 2018, 9, 306. [Google Scholar] [CrossRef]
- Barby, F.F.; Ráb, P.; Lavoué, S.; Ezaz, T.; Bertollo, L.A.C.; Kilian, A.; Maruyama, S.R.; Oliveira, E.A.; Artoni, R.F.; Santos, M.H.; et al. Emerging patterns of genome organization in Notopteridae species (Teleostei, Osteoglossiformes) as revealed by Zoo-FISH and Comparative Genomic Hybridization (CGH). Sci. Rep. 2019, 9, 1112. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Ráb, P.; Ezaz, T.; Bertollo, L.A.C.; Lavoué, S.; Oliveira, E.A.; Sember, A.; Molina, W.F.; Souza, F.H.S.; Majtánová, Z.; et al. Deciphering the Evolutionary History of Arowana Fishes (Teleostei, Osteoglossiformes, Osteoglossidae): Insight from Comparative Cytogenomics. Int. J. Mol. Sci. 2019, 20, 4296. [Google Scholar] [CrossRef]
- De Oliveira, E.A.; Bertollo, L.A.C.; Ráb, P.; Ezaz, T.; Yano, C.F.; Hatanaka, T.; Jedege, O.I.; Tanomtong, A.; Liehr, T.; Sember, A.; et al. Cytogenetics, genomics and biodiversity of the South American and African Arapaimidae fish family (Teleostei, Osteoglossiformes). PLoS ONE 2019, 14, e0214225. [Google Scholar] [CrossRef]
- Majtánová, Z.; Symonová, R.; Arias-Rodriguez, L.; Sallan, L.; Ráb, P. “Holostei versus Halecostomi” Problem: Insight from Cytogenetics of Ancient Nonteleost Actinopterygian Fish, Bowfin Amia calva. J. Exp. Zool. Part B Mol. Dev. Evol. 2017, 328, 620–628. [Google Scholar] [CrossRef]
- Souza, F.H.S.; Perez, M.F.; Bertollo, L.A.C.; Oliveira, E.A.; Lavoué, S.; Gestich, C.C.; Ráb, P.; Ezaz, T.; Liehr, T.; Viana, O.F.; et al. Interspecific Genetic Differences and Historical Demography in South American Arowanas (Osteoglossiformes, Osteoglossidae, Osteoglossum). Genes 2019, 10, 693. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Howell, W. Vertebrate Genome Evolution in the Light of Fish Cytogenomics and rDNAomics. Genes 2018, 9, 96. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Robledillo, L.A.; Koblízkova, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef]
- Peona, V.; Kutschera, V.E.; Blom, M.P.K.; Irestedt, M.; Suh, A. Satellite DNA evolution in Corvoidea inferred from short and long reads. Mol. Ecol. 2022, 32, 1–18. [Google Scholar] [CrossRef]
- Sena, R.S.; Heringer, P.; Valeri, M.P.; Pereira, V.S.; Kuhn, G.C.S.; Svartman, M. Identification and characterization of satellite DNAs in two-toed sloths of the genus Choloepus (Megalonychidae, Xenarthra). Sci. Rep. 2020, 10, 19202. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; Cabrero, J.; López-León, M.D.; Camacho, J.P.M. Satellite DNA content illuminates the ancestry of a supernumerary (B) chromosome. Chromosoma 2017, 126, 487–500. [Google Scholar] [CrossRef]
- Goes, C.A.G.; Santos, R.Z.; Aguiar, W.R.C.; Alves, D.C.V.; Silva, D.M.Z.A.; Foresti, F.; Oliveira, C.; Utsunomia, R.; Porto-Foresti, F. Revealing the Satellite DNA History in Psalidodon and Astyanax Characid Fish by Comparative Satellitomics. Front. Genet. 2022, 13. [Google Scholar] [CrossRef]
- Kretschmer, R.; Goes, C.A.G.; Bertollo, L.A.C.; Ezaz, T.; Porto-Foresti, F.; Toma, G.A.; Utsunomia, R.; Cioffi, M.B. Satellitome analysis illuminates the evolution of ZW sex chromosomes of Triportheidae fishes (Teleostei: Characiformes). Chromosoma 2022, 131, 29–45. [Google Scholar] [CrossRef]
- Silva, D.M.Z.A.; Utsunomia, R.; Ruiz-Ruano, F.J.; Daniel, S.N.; Porto-OFresti, F.; Hashimoto, D.T.; Oliveira, C.; Camacho, J.P.M.; Foresti, F. High-throughput analysis unveils a highly shared satellite DNA library among three species of fish genus Astyanax. Sci. Rep. 2017, 7, 12726. [Google Scholar] [CrossRef] [PubMed]
- Arai, R. Fish Karyotypes: A Check List; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Bian, C.; Hu, Y.; Ravi, V.; Kuznetsov, I.S.; Shen, X.; Mu, X.; Sun, Y.; You, X.; Li, J.; Li, X.; et al. The Asian arowana (Scleropages formosus) genome provides new insights into the evolution of an early lineage of teleosts. Sci. Rep. 2016, 6, 24501. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.Y.; Kwan, H.Y.; Thevasagayam, N.M.; Prakki, S.R.S.; Kuznetsova, I.S.; Ngoh, S.Y.; Lim, Z.; Feng, F.; Chang, A.; Orbán, L. The first transcriptome and genetic linkage map for Asian arowana. Mol. Ecol. Resour. 2014, 14, 622–635. [Google Scholar] [CrossRef] [PubMed]
- Urushido, T. Karyotype of three species of fishes in the order Osteoglossiformes. Chromosom. Inf. Serv. 1975, 18, 20–22. [Google Scholar]
- Ellegren, H. Evolutionary stasis: The stable chromosomes of birds. Trends Ecol. Evol. 2010, 25, 283–291. [Google Scholar] [CrossRef]
- Mandáková, T.; Heenan, P.B.; Lysak, M.A. Island species radiation and karyotypic stasis in Pachycladon allopolyploids. BMC Evol. Biol. 2010, 10, 367. [Google Scholar] [CrossRef]
- Molina, W.F. Chromosomal changes and stasis in marine fish groups. In Fish Cytogenetics, 1st ed.; Pisano, E., Ozouf-Costaz, C., Foresti, F., Kapoor, B., Eds.; Science Publishers: Cambridge, UK, 2007; pp. 69–110. [Google Scholar]
- Sadílek, D.; Nguyen, P.; Koç, H.; Kovařík, F.; Yağmur, E.A.; Yağmur, F. Molecular cytogenetics of Androctonus scorpions: An oasis of calm in the turbulent karyotype evolution of the diverse family Buthidae. Biol. J. Linn. Soc. 2015, 115, 69–76. [Google Scholar] [CrossRef]
- Charlesworth, B.; Lande, R.; Slatkin, M. A Neo-Darwinian Commentary on Macroevolution. Evolution 1982, 36, 474–498. [Google Scholar]
- Hansen, T.F. Stabilizing selection and the Comparative Analysis of Adaptation. Evolution 1997, 51, 1341–1351. [Google Scholar] [CrossRef]
- Motta-Neto, C.C.; Cioffi, M.B.; Costa, G.W.W.F.; Amorim, K.D.J.; Bertollo, L.A.C.; Artoni, R.F.; Molina, W.F. Overview on Karyotype Stasis in Atlantic Grunts (Eupercaria, Haemulidae) and the Evolutionary Extensions for Other Marine Fish Groups. Front. Mar. Sci. 2019, 6, 628. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2018, 127, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Franco, W.; Ferreira, R.; Bertollo, L.A.C. Chromosomes as tools for discovering biodiversity—The case of erythrinidae fish family. In Recent Trends in Cytogenet Studies; Methodology and Applcations; Tirunilai, P., Ed.; InTech: Rijeka, Croatia, 2012; pp. 125–146. [Google Scholar]
- Deon, G.A.; Glugoski, L.; Vicari, M.R.; Nogaroto, V.; Sassi, F.M.C.; Cioffi, M.B.; Liehr, T.; Bertollo, L.A.C.; Moreira-Filho, O. Highly Rearranged Karyotypes and Multiple Sex Chromosome Systems in Armored Catfishes from the Genus Harttia (Teleostei, Siluriformes). Genes 2020, 11, 1366. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.; Garcia, C.; Matoso, D.A.; Jesus, I.S.; Cioffi, M.B.; Bertollo, L.A.C.; Zuanon, J.; Feldberg, E. The Bunocephalus coracoideus Species Complex (Siluriformes, Aspredinidae). Signs of a Speciation Process through Chromosomal, Genetic and Ecological Diversity. Front. Genet. 2017, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Pazza, R.; Kavalco, K.F.; Bertollo, L.A.C. Chromosome polymorphism in Astyanax fasciatus (Teleostei, Characidae). 1. Karyotype analysis, Ag-NORs and mapping of the 18S and 5S ribosomal genes in sympatric karyotypes and their possible hybrid forms. Cytogenet. Genome Res. 2006, 112, 313–319. [Google Scholar] [CrossRef]
- Degrandi, T.M.; Gunski, R.J.; Garnero, A.V.; Oliveira, E.H.C.; Kretschmer, R.; Souza, M.S.; Barcellos, S.A.; Hass, I. The distribution of 45S rDNA sites in bird chromosomes suggests multiple evolutionary histories. Genet. Mol. Biol. 2020, 43, e20180331. [Google Scholar] [CrossRef]
- Yano, C.F.; Merlo, M.A.; Portela-Bens, S.; Cioffi, M.B.; Bertollo, L.A.C.; Santos-Júnior, C.D.; Rebordinos, L. Evolutionary Dynamics of Multigene Families in Triportheus (Characiformes, Triportheidae): A Transposon Mediated Mechanism? Front. Mar. Sci. 2020, 7, 6. [Google Scholar] [CrossRef]
- Gunski, R.J.; Kretschmer, R.; Souza, M.S.; Furo, I.O.; Barcellos, S.A.; Costa, A.L.; Cioffi, M.B.; Oliveira, E.H.C.; Garnero, A.D.V. Evolution of Bird Sex Chromosomes Narrated by Repetitive Sequences: Unusual W Chromosome Enlargement in Gallinula melanops (Aves: Gruiformes: Rallidae). Cytogenet. Genome Res. 2019, 158, 152–159. [Google Scholar] [CrossRef]
- Salvadori, S.; Deiana, A.M.; Deidda, F.; Lobina, C.; Mulas, A.; Coluccia, E. XX/XY sex chromosome system and chromosome markers in the snake eel Ophisurus serpens (Anguilliformes: Ophichtidae). Mar. Biol. Res. 2018, 14, 158–164. [Google Scholar] [CrossRef]
- Dover, G. Molecular drive: A cohesive mode of species evolution. Nature 1982, 299, 111–117. [Google Scholar] [CrossRef]
- Goffová, I.; Fajkus, J. The rDNA Loci—Intersections of Replication, Transcription, and Repair Pathways. Int. J. Mol. Sci. 2021, 22, 1302. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Chromosome spreading of associated transposable elements and ribosomal DNA in the fish Erythrinus erythrinus. Implications for genome change and karyoevolution in fish. BMC Evol. Biol. 2010, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Symonová, R.; Majtánová, Z.; Sember, A.; Staaks, G.B.O.; Bohlen, J.; Freyhof, J.; Rábová, M.; Ráb, P. Genome differentiation in a species pair of coregonine fishes: An extremely rapid speciation driven by stress-activated retrotransposons mediating extensive ribosomal DNA multiplications. BMC Evol. Biol. 2013, 13, 42. [Google Scholar] [CrossRef] [PubMed]
- Collares-Pereira, M.J.; Ráb, P. NOR polymorphism in the Iberian species Chondrostoma lusitanicum (Pisces: Cyprinidae)—Re-examination by FISH. Genetica 1999, 105, 301–303. [Google Scholar] [CrossRef] [PubMed]
- Ghigliotti, L.; Near, T.J.; Ferrando, S.; Vacchi, M.; Pisano, E. Cytogenetic diversity in the Antarctic plunderfishes (Notothenioidei: Artedidraconidae). Antarct. Sci. 2010, 22, 805–814. [Google Scholar] [CrossRef]
- Sassi, F.M.C.; Oliveira, E.A.; Bertollo, L.A.C.; Nirchio, M.; Hatanaka, T.; Marinho, M.M.F.; Moreira-Filho, O.; Aroutiounian, R.; Liehr, T.; Al-Rikabi, A.B.H.; et al. Chromosomal Evolution and Evolutionary Relationships of Lebiasina Species (Characiformes, Lebiasinidae). Int. J. Mol. Sci. 2019, 20, 2944. [Google Scholar] [CrossRef]
- Gibbons, J.G.; Branco, A.T.; Godinho, S.A.; Yu, S.; Lemos, B. Concerted copy number variation balances ribosomal DNA dosage in human and mouse genomes. Proc. Natl. Acad. Sci. USA 2015, 112, 2485–2490. [Google Scholar] [CrossRef]
- Lavrinienko, A.; Jernfors, T.; Koskimäki, J.J.; Pirttilä, A.M.; Watts, P.C. Does Intraspecific Variation in rDNA Copy Number Affect Analysis of Microbial Communities? Trends Microbiol. 2021, 29, 19–27. [Google Scholar] [CrossRef]
- Utsunomia, R.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Goes, C.A.G.; Melo, S.; Ramos, L.P.; Oliveira, C.; Porto-Foresti, F.; Foresti, F.; Hashimoto, D.T. Satellitome landscape analysis of Megaleporinus macrocephalus (Teleostei, Anostomidae) reveals intense accumulation of satellite sequences on the heteromorphic sex chromosome. Sci. Rep. 2019, 9, 5856. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Singchat, W.; Jehangir, M.; Suntronpong, A.; Pnathum, T.; Malaivijitnond, S.; Srikulnath, K. Dark Matter of Primate Genomes: Satellite DNA Repeats and Their Evolutionary Dynamics. Cells 2020, 9, 2714. [Google Scholar] [CrossRef]
- Dos Santos, R.Z.; Calegari, R.M.; Silva, D.M.Z.A.; Ruiz-Ruano, F.J.; Melo, S.; Oliveira, C.; Foresti, F.; Uliano-Silva, M.; Porto-Foresti, F.; Utsunomia, R. A Long-Term Conserved Satellite DNA That Remains Unexpanded in Several Genomes of Characiformes Fish Is Actively Transcribed. Genome Biol. Evol. 2021, 13, evab002. [Google Scholar] [CrossRef]
- Tajima, F. Simple methods for testing the molecular evolutionary clock hypothesis. Genetics 1993, 135, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Robles, F.; Herrán, R.; Ludwig, A.; Rejón, C.R.; Rejón, M.R.; Garrido-Ramos, M.A. Evolution of ancient satellite DNAs in sturgeon genomes. Gene 2004, 338, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, M.E.; Santini, F.; Brock, C.; Harmon, L.J. Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. Proc. Natl. Acad. Sci. USA 2009, 106, 13410–13414. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.M.; Tan, M.H.; Croft, L.J.; Hammer, M.P.; Gan, H.M. Whole Genome Sequencing of the Asian Arowana (Scleropages formosus) Provides Insights into the Evolution of Ray-Finned Fishes. Genome Biol. Evol. 2015, 7, 2885–2895. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B.; Sniegowski, P.; Stephan, W. The evolutionary dynamics of repetitive DNA in eukaryotes. Nature 1994, 371, 215–220. [Google Scholar] [CrossRef]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The Centromere Paradox: Stable Inheritance with Rapidly Evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef]
- O’Neill, R.J.; Eldridge, M.D.B.; Metcalfe, C.J. Centromere Dynamics and Chromosome Evolution in Marsupials. J. Hered. 2004, 95, 375–381. [Google Scholar] [CrossRef]
- Melters, D.P.; Bradnam, K.R.; Young, H.A.; Telis, N.; May, M.R.; Rybu, J.G.; Sebra, R.; Peluso, P.; Eid, J.; Rank, D.; et al. Comparative analysis of tandem repeats from hundreds of species reveals unique insights into centromere evolution. Genome Biol. 2013, 14, R10. [Google Scholar] [CrossRef]
- Plohl, M.; Meštrović, N.; Mravinac, B. Centromere identity from the DNA point of view. Chromosoma 2014, 123, 313–325. [Google Scholar] [CrossRef]
- Talbert, P.B.; Henikoff, S. The genetics and epigenetics of satellite centromeres. Genome Res. 2022, 32, 608–615. [Google Scholar] [CrossRef]
- Völker, M.; Ráb, P. Direct chromosome preparation from regenerating fish fin tissue. In Fish Cytogenetic Techniques; Ozouf-Costaz, C., Pisan, E., Foresti, F., Toledo, L.F.A., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 37–41. [Google Scholar]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef]
- Smit, A.F.A.; Hubley, R.; Green, P. RepeatMasker. Available online: http://repeatmasker.org (accessed on 15 January 2023).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Martins, C.; Ferreira, I.A.; Oliveira, C.; Foresti, F.; Galetti, P.M. A tandemly repetitive centromeric DNA sequence of the fish Hoplias malabaricus (Characiformes: Erythrinidae) is derived from 5S rDNA. Genetica 2006, 127, 133. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Martins, C.; Centofante, L.; Jacobina, U.; Bertollo, L.A.C. Chromosomal Variability among Allopatric Populations of Erythrinidae Fish Hoplias malabaricus: Mapping of Three Classes of Repetitive DNAs. Cytogenet. Genome Res. 2009, 125, 132–141. [Google Scholar] [CrossRef]
- Pendás, A.M.; Móran, P.; Freije, J.P.; Garcia-Vásquez, E. Chromosomal location and nucleotide sequence of two tandem repeats of the Atlantic salmon 5S rDNA. Cytogenet. Cell Genet. 1994, 67, 31–36. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Cioffi, M.B. Fish-FISH: Molecular cytogenetics in fish species. In Fluorescence In Situ Hybridization (FISH); Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 429–443. [Google Scholar]
- Yang, F.; Trifonov, V.; Ng, B.; Kosyakova, N.; Carter, N.P. Generation of paint probes by flow-sorted and microdissected chromosomes. In Fluorescence In Situ Hybridization (FISH)—Application Guide; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 35–52. [Google Scholar]
- Zwick, M.S.; Hanson, R.E.; Islam-Faridi, M.N.; Stelly, D.M.; Wing, R.A.; Price, H.J.; McKnight, T.D. A rapid procedure for the isolation of C 0 t-1 DNA from plants. Genome 1997, 40, 138–142. [Google Scholar] [CrossRef]
- Symonová, R.; Sember, A.; Majtánová, Z.; Ráb, P. Characterization of fish genomes by GISH and CGH. In Fish Cytogenetic Techniques (Chondrichthyans and Teleosts); Ozouf-Costaz, C., Pisano, E., Foresti, F., Almeida Toledo, L.F., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 118–131. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SatDNA Family | SF | RUL | Abundance | Divergence | A + T (%) |
|---|---|---|---|---|---|
| * SfoSat01-180 | 180 | 0.025654546 | 7.06 | 36.7 | |
| * SfoSat02-39 | 39 | 0.016109651 | 11.84 | 56.4 | |
| * SfoSat03-198 | 198 | 0.010209052 | 2.27 | 52.5 | |
| * SfoSat04-109 | 109 | 0.001647104 | 1.88 | 59.6 | |
| * SfoSat05-107 | 107 | 0.001390976 | 7.70 | 55.1 | |
| SfoSat06-4000 | 4000 | 0.001339709 | 7.11 | 51.4 | |
| * SfoSat07-156 | 156 | 0.001159315 | 2.80 | 57.1 | |
| * SfoSat08-111 | 111 | 0.001183768 | 6.09 | 60.4 | |
| * SfoSat09-108 | 108 | 0.000889043 | 11.46 | 64.8 | |
| * SfoSat10-239 | 239 | 0.00089077 | 3.73 | 58.6 | |
| SfoSat11-1682 | 1682 | 0.000778082 | 9.24 | 50.2 | |
| * SfoSat12-337 | 337 | 0.000767312 | 6.28 | 58.8 | |
| SfoSat13-2034 | 2034 | 0.000692241 | 9.07 | 56.8 | |
| * SfoSat14-95 | 95 | 0.000607639 | 1.61 | 61.1 | |
| * SfoSat15-261 | 261 | 0.000255594 | 3.65 | 50.6 | |
| SfoSat16-6 | 6 | 0.000252204 | 15.04 | 50.0 | |
| * SfoSat17-699 | 699 | 0.000223762 | 2.97 | 50.6 | |
| * SfoSat18-767 | 767 | 0.000166812 | 4.07 | 43.0 | |
| SfoSat19-993 | 993 | 0.00012931 | 1.61 | 40.7 | |
| SfoSat20-677 | 677 | 0.000127314 | 0.87 | 54.2 | |
| * SfoSat21-651 | 1 | 651 | 0.000115523 | 4.55 | 38.2 |
| * SfoSat22-293 | 293 | 0.000112537 | 3.07 | 53.9 | |
| * SfoSat23-291 | 1 | 291 | 0.000103291 | 5.61 | 50.5 |
| SfoSat24-120 | 120 | 0.000099403 | 5.08 | 60.0 | |
| SfoSat25-616 | 616 | 0.000089154 | 1.77 | 55.9 |
| Species (Variety) | Phenotype (Code) | Sampling Site | n |
|---|---|---|---|
| S. formosus (green) | Asian Green (AG) | Aquarium trade, Song Khram river | (02♀02♂) |
| S. formosus (green) | Yellow TailSilver (YS) | Aquarium trade, Song Khram river | (04♀02♂) |
| S. formosus (golden) | Gold Crossback (GC) | Aquarium trade, Origin unknown | (02♀02♂) |
| S. formosus (golden) | High Back Golden (HG) | Aquarium trade, Origin unknown | (02♀02♂) |
| S. formosus (red) | Super Red (SR) | Aquarium trade, Origin unknown | (03♀02♂) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toma, G.A.; dos Santos, N.; dos Santos, R.; Rab, P.; Kretschmer, R.; Ezaz, T.; Bertollo, L.A.C.; Liehr, T.; Porto-Foresti, F.; Hatanaka, T.; et al. Cytogenetics Meets Genomics: Cytotaxonomy and Genomic Relationships among Color Variants of the Asian Arowana Scleropages formosus. Int. J. Mol. Sci. 2023, 24, 9005. https://doi.org/10.3390/ijms24109005
Toma GA, dos Santos N, dos Santos R, Rab P, Kretschmer R, Ezaz T, Bertollo LAC, Liehr T, Porto-Foresti F, Hatanaka T, et al. Cytogenetics Meets Genomics: Cytotaxonomy and Genomic Relationships among Color Variants of the Asian Arowana Scleropages formosus. International Journal of Molecular Sciences. 2023; 24(10):9005. https://doi.org/10.3390/ijms24109005
Chicago/Turabian StyleToma, Gustavo A., Natália dos Santos, Rodrigo dos Santos, Petr Rab, Rafael Kretschmer, Tariq Ezaz, Luiz A. C. Bertollo, Thomas Liehr, Fábio Porto-Foresti, Terumi Hatanaka, and et al. 2023. "Cytogenetics Meets Genomics: Cytotaxonomy and Genomic Relationships among Color Variants of the Asian Arowana Scleropages formosus" International Journal of Molecular Sciences 24, no. 10: 9005. https://doi.org/10.3390/ijms24109005
APA StyleToma, G. A., dos Santos, N., dos Santos, R., Rab, P., Kretschmer, R., Ezaz, T., Bertollo, L. A. C., Liehr, T., Porto-Foresti, F., Hatanaka, T., Tanomtong, A., Utsunomia, R., & Cioffi, M. B. (2023). Cytogenetics Meets Genomics: Cytotaxonomy and Genomic Relationships among Color Variants of the Asian Arowana Scleropages formosus. International Journal of Molecular Sciences, 24(10), 9005. https://doi.org/10.3390/ijms24109005