
LTR Retroelements and Bird Adaptation to Arid Environments

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
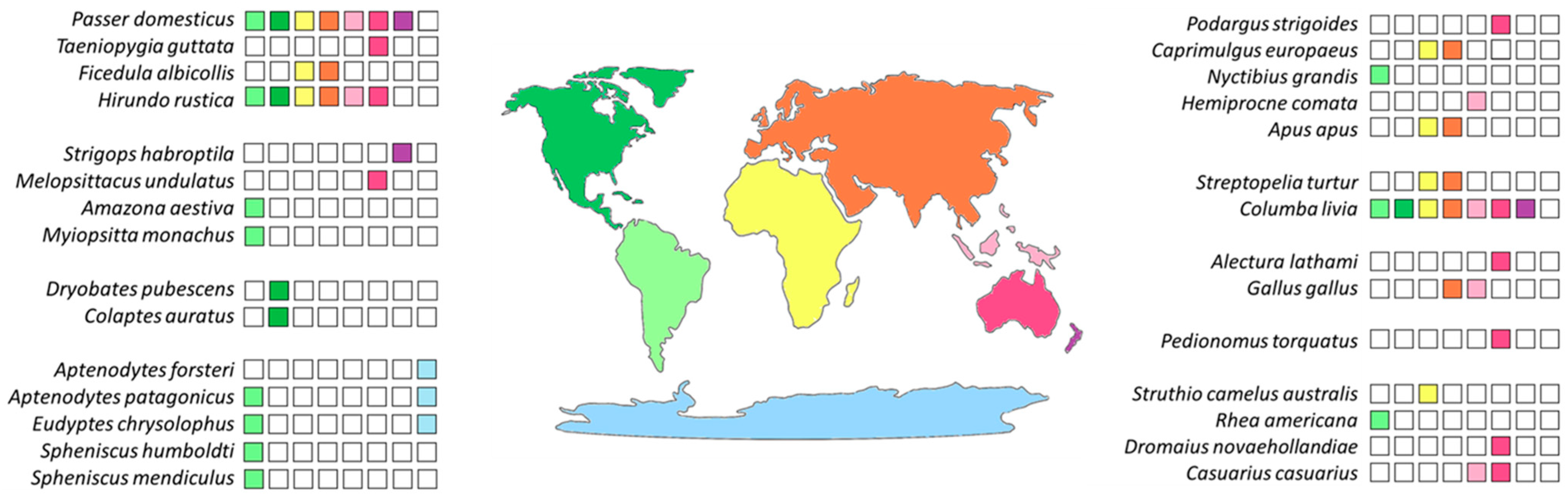
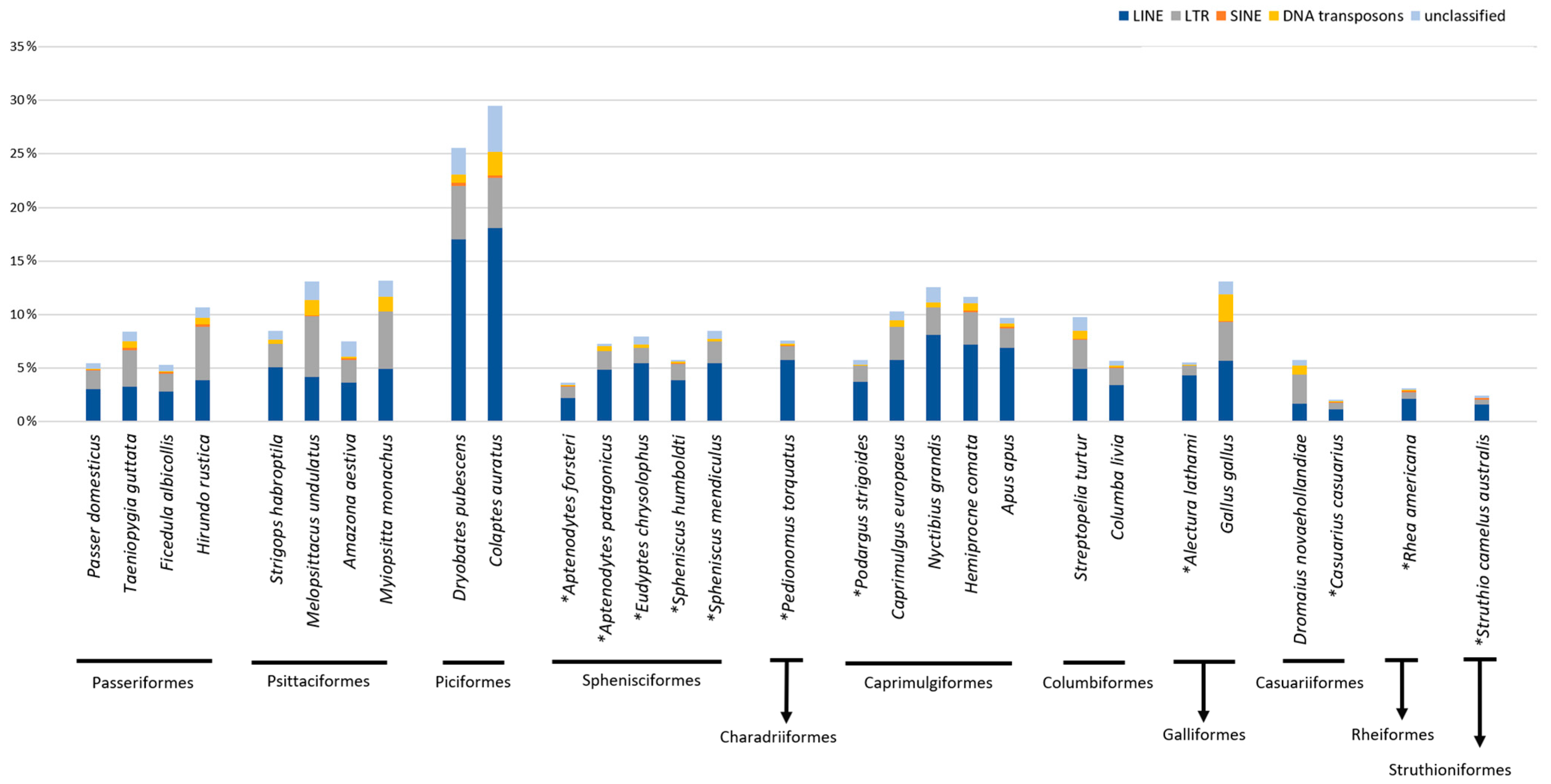
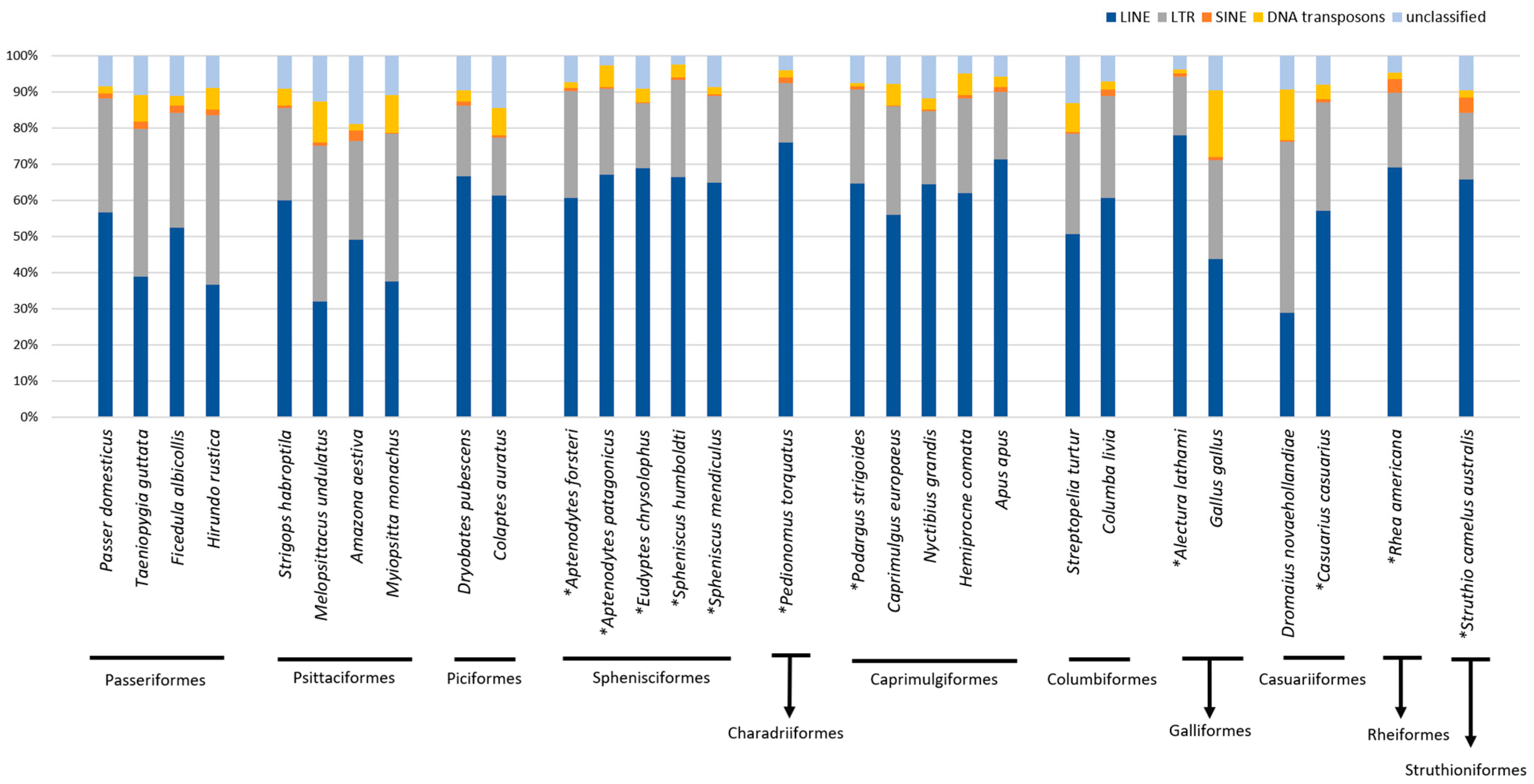
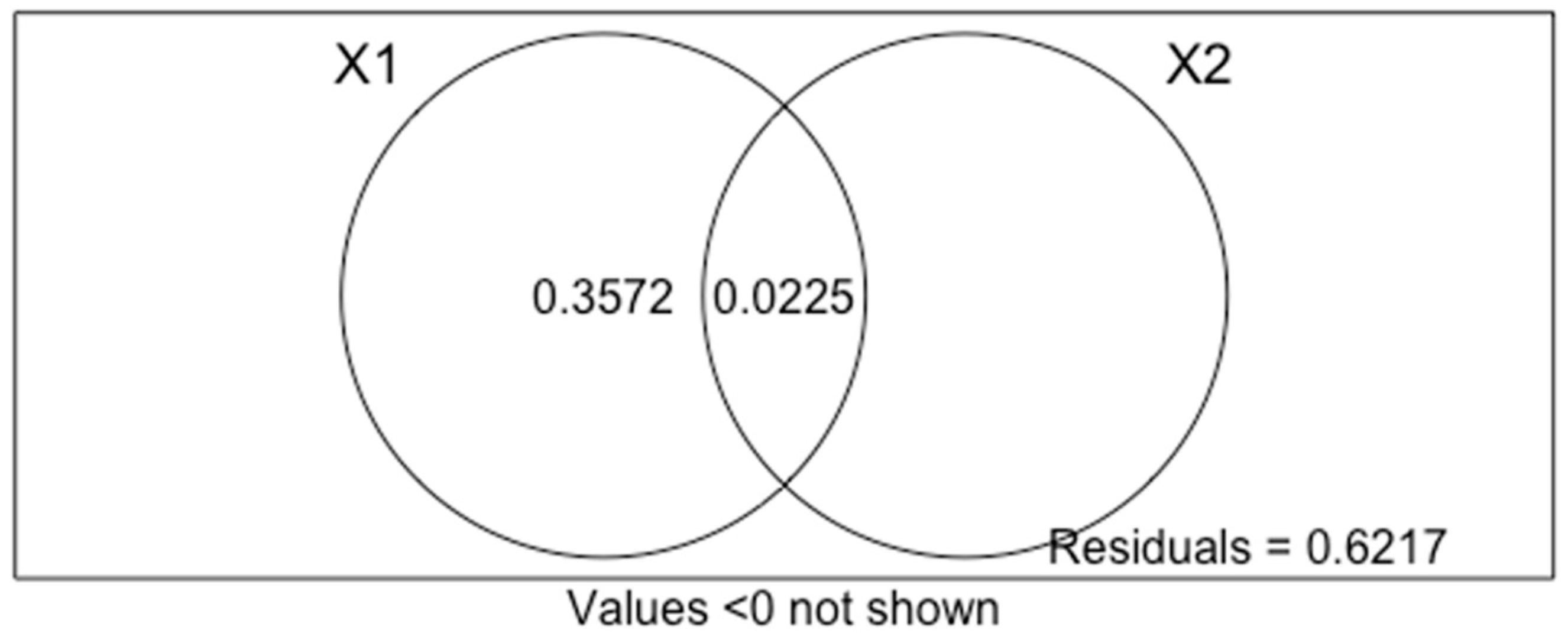
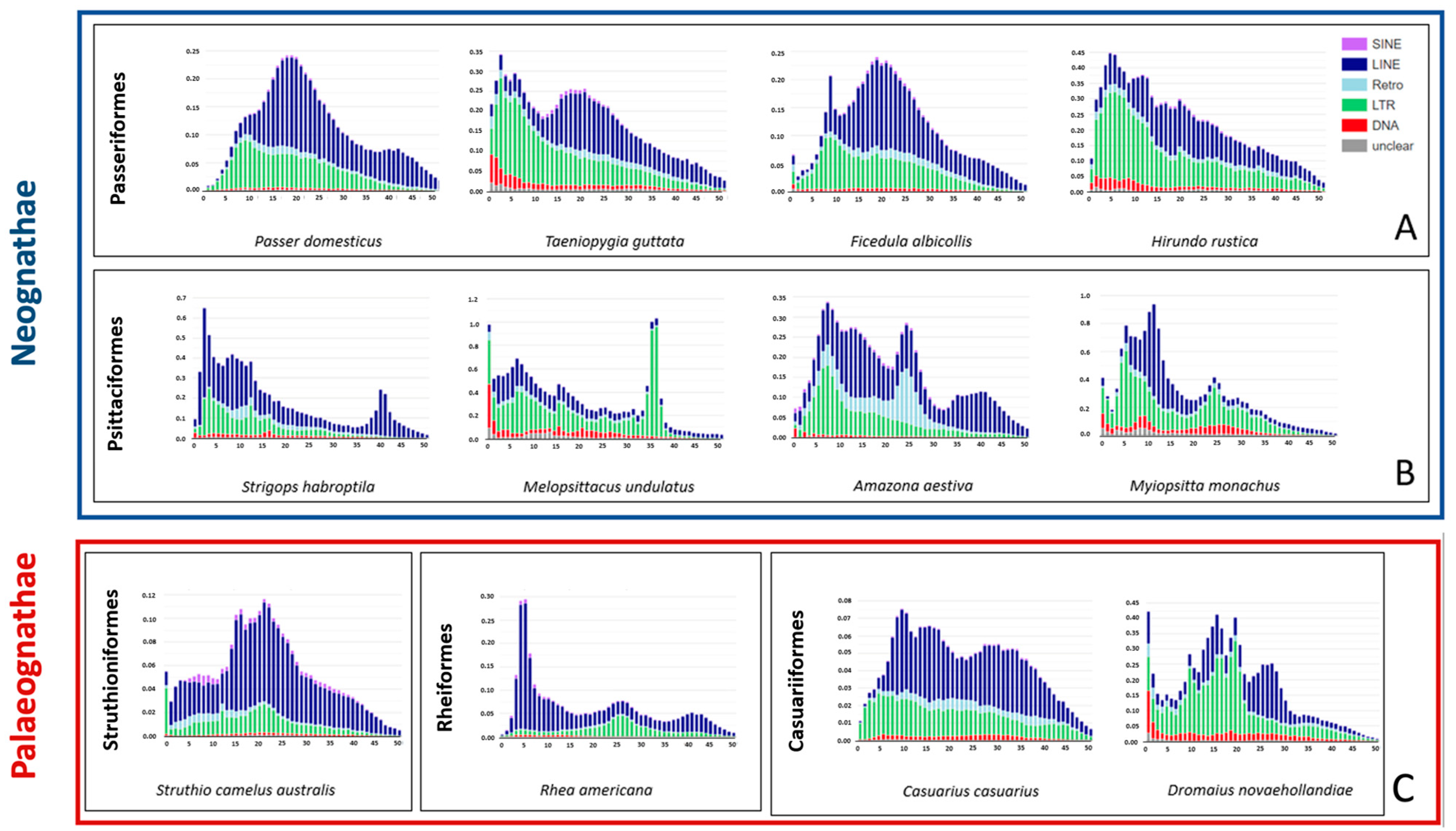
2. Results
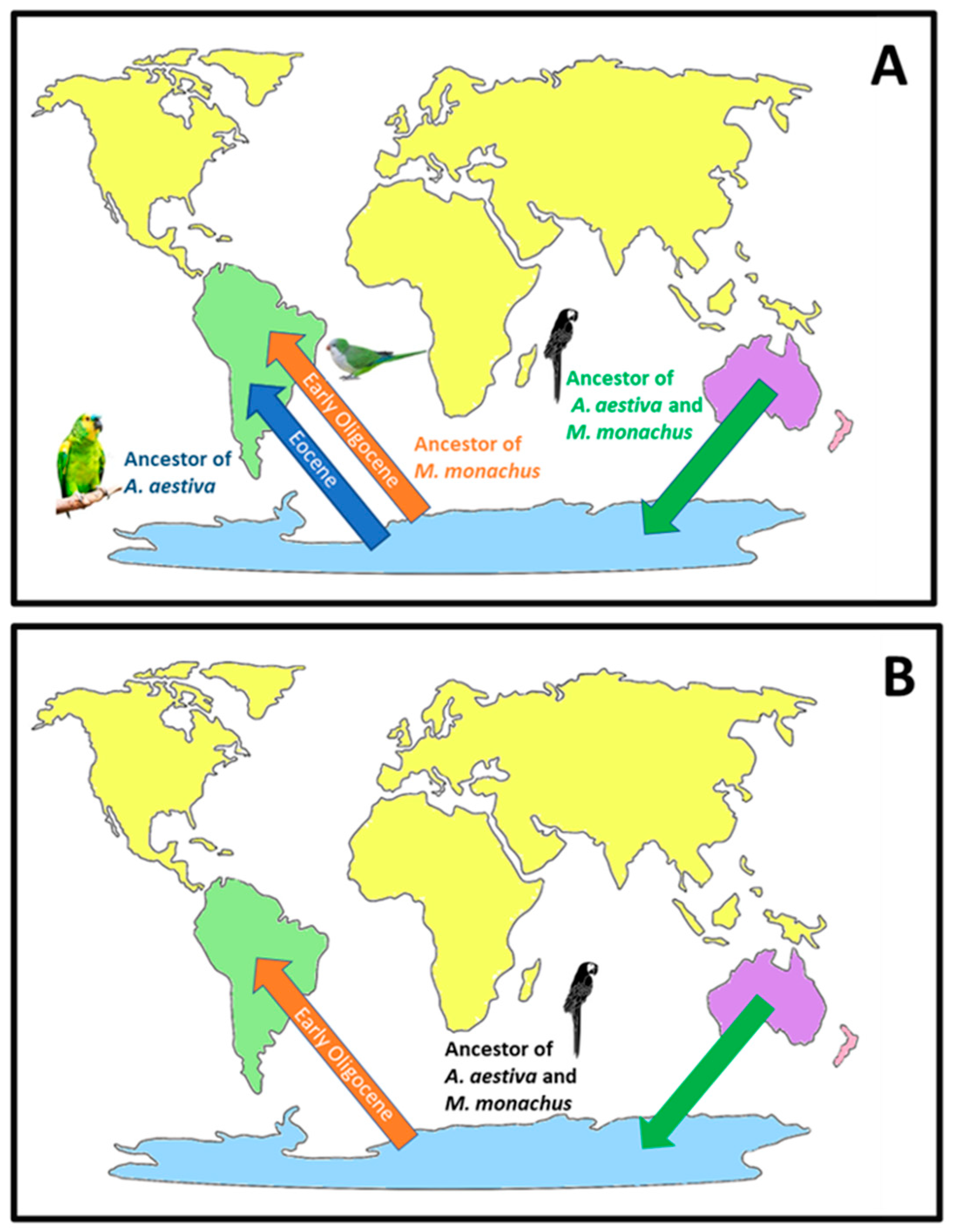
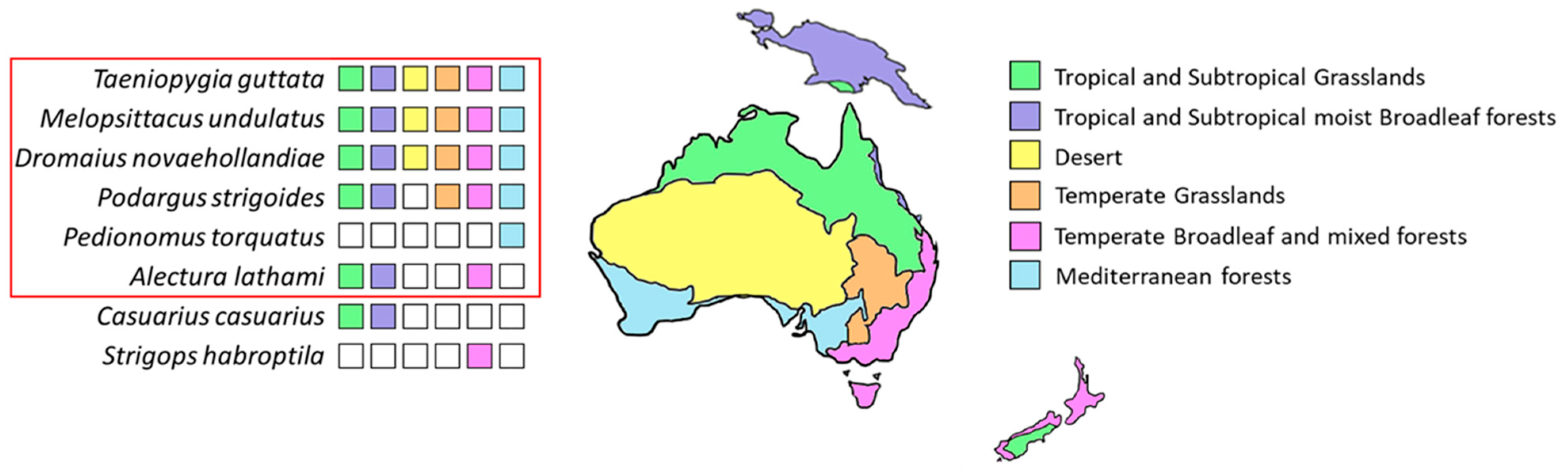
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Schrader, L.; Smith, J. The impact of transposable elements in adaptive evolution. Mol. Ecol. 2019, 28, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, A.M.; Ferrito, V.; Biscotti, M.A.; Canapa, A.; Capriglione, T. Transposable elements and stress in vertebrates: An overview. Int. J. Mol. Sci. 2021, 22, 1970. [Google Scholar] [CrossRef] [PubMed]
- Canapa, A.; Biscotti, M.A.; Barucca, M.; Carducci, F.; Carotti, E.; Olmo, E. Shedding light upon the complex net of genome size, genome composition and environment in chordates. Eur. Zool. J. 2020, 87, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Carducci, F.; Barucca, M.; Canapa, A.; Carotti, E.; Biscotti, M.A. Mobile Elements in Ray-Finned Fish Genomes. Life 2020, 10, 221. [Google Scholar] [CrossRef]
- Biscotti, M.A.; Carducci, F.; Olmo, E.; Canapa, A. Vertebrate genome size and the impact of transposable elements in genome evolution. In Evolution, Origin of Life, Concepts and Methods; Springer International Publishing: Cham, Switzerland, 2019; pp. 233–251. [Google Scholar]
- Carducci, F.; Biscotti, M.A.; Barucca, M.; Canapa, A. Transposable elements in vertebrates: Species evolution and environmental adaptation. Eur. Zool. J. 2019, 86, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Canapa, A.; Barucca, M.; Biscotti, M.A.; Forconi, M.; Olmo, E. Transposons, genome size, and evolutionary insights in animals. Cytogenet. Genome Res. 2015, 147, 217–239. [Google Scholar] [CrossRef]
- Chénais, B.; Caruso, A.; Hiard, S.; Casse, N. The impact of transposable elements on eukaryotic genomes: From genome size increase to genetic adaptation to stressful environments. Gene 2012, 509, 7–15. [Google Scholar] [CrossRef]
- Casacuberta, E.; González, J. The impact of transposable elements in environmental adaptation. Mol. Ecol. 2013, 6, 1503–1517. [Google Scholar] [CrossRef]
- Makarevitch, I.; Waters, A.J.; West, P.T.; Stitzer, M.; Hirsch, C.N.; Ross-Ibarra, J.; Springer, N.M. Transposable elements contribute to activation of maize genes in response to abiotic stress. PLoS Genet. 2015, 11, e1004915. [Google Scholar]
- Piacentini, L.; Fanti, L.; Specchia, V.; Bozzetti, M.P.; Berloco, M.; Palumbo, G.; Pimpinelli, S. Transposons, environmental changes, and heritable induced phenotypic variability. Chromosoma 2014, 123, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujino, K.; Hashida, S.N.; Ogawa, T.; Natsume, T.; Uchiyama, T.; Mikami, T.; Kishima, Y. Temperature controls nuclear import of Tam3 transposase in Antirrhinum. Plant J. 2011, 65, 146–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carotti, E.; Carducci, F.; Canapa, A.; Barucca, M.; Biscotti, M.A. Transposable element tissue-specific response to temperature stress in the stenothermal fish Puntius tetrazona. Animals 2023, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Carotti, E.; Carducci, F.; Greco, S.; Gerdol, M.; Di Marino, D.; Perta, N.; La Teana, A.; Canapa, A.; Barucca, M.; Biscotti, M.A. Transcriptional contribution of transposable elements in relation to salinity conditions in teleosts and silencing mechanisms involved. Int. J. Mol. Sci. 2022, 23, 5215. [Google Scholar] [CrossRef]
- Carotti, E.; Carducci, F.; Canapa, A.; Barucca, M.; Greco, S.; Gerdol, M.; Biscotti, M.A. Transposable elements and teleost migratory behaviour. Int. J. Mol. Sci. 2021, 22, 602. [Google Scholar] [CrossRef] [PubMed]
- Carducci, F.; Biscotti, M.A.; Forconi, M.; Barucca, M.; Canapa, A. An intriguing relationship between teleost Rex3 retroelement and environmental temperature. Biol. Lett. 2019, 15, 20190279. [Google Scholar] [CrossRef] [Green Version]
- Gregory, T.R.; Nicol, J.A.; Tamm, H.; Kullman, B.; Kullman, K.; Leitch, I.J.; Murray, B.G.; Kapraun, D.F.; Greilhuber, J.; Bennett, M.D. Eukaryotic genome size databases. Nucleic Acids Res. 2007, 35, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Wright, N.A.; Gregory, T.R.; Witt, C.C. Metabolic ’engines’ of flight drive genome size reduction in birds. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132780. [Google Scholar] [CrossRef] [Green Version]
- Kapusta, A.; Suh, A.; Feschotte, C. Dynamics of genome size evolution in birds and mammals. Proc. Natl. Acad. Sci. USA 2017, 114, 1460–1469. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.L.; Hughes, M.K. Small genomes for better flyers. Nature 1995, 377, 391. [Google Scholar] [CrossRef]
- Organ, C.L.; Shedlock, A.M. Palaeogenomics of pterosaurs and the evolution of small genome size in flying vertebrates. Biol. Lett. 2009, 5, 47–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.D.L.; Gregory, T.R. The genome sizes of megabats (Chiroptera: Pteropodidae) are remarkably constrained. Biol. Lett. 2009, 5, 347–351. [Google Scholar] [CrossRef]
- Zhang, G.; Li, C.; Li, Q.; Li, B.; Larkin, D.M.; Lee, C.; Storz, J.F.; Antunes, A.; Greenwold, M.J.; Meredith, R.W.; et al. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hruska, J.P.; Manthey, J.D. De novo assembly of a chromosome-scale reference genome for the northern flicker Colaptes auratus. G3 2021, 11, jkaa026. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.D.; Moyle, R.G.; Boissinot, S. Multiple and independent phases of transposable element amplification in the genomes of Piciformes (Woodpeckers and Allies). Genome Biol. Evol. 2018, 10, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotero-Caio, C.G.; Platt, R.N., II; Suh, A.; Ray, D.A. Evolution and diversity of transposable elements in vertebrate genomes. Genome Biol. Evol. 2017, 9, 161–177. [Google Scholar] [CrossRef] [Green Version]
- Cracraft, J. Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc. R. Soc. B Biol. Sci. 2001, 268, 459–469. [Google Scholar] [CrossRef]
- Feduccia, A. Explosive evolution in tertiary birds and mammals. Science 1995, 267, 637–638. [Google Scholar] [CrossRef]
- Moyle, R.G.; Oliveros, C.H.; Andersen, M.J.; Hosner, P.A.; Benz, B.W.; Manthey, J.D.; Travers, S.L.; Brown, R.M.; Faircloth, B.C. Tectonic collision and uplift of Wallacea triggered the global songbird radiation. Nat. Commun. 2016, 7, 12709. [Google Scholar] [CrossRef] [Green Version]
- Cracraft, J. Continental drift, paleoclimatology, and the evolution and biogeography of birds. J. Zool. 1973, 169, 455–545. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Common, M.S.; Norton, T.W. Biodiversity: Its Conservation in Australia. Ambio 1992, 21, 258–265. [Google Scholar]
- Chalopin, D.; Naville, M.; Plard, F.; Galiana, D.; Volff, J.-N. Comparative analysis of transposable elements highlights mobilome diversity and evolution in vertebrates. Genome Biol. Evol. 2015, 7, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, C.H.; Field, D.J.; Ksepka, D.T.; Barker, F.K.; Aleixo, A.; Andersen, M.J.; Alström, P.; Benz, B.W.; Braun, E.L.; Braun, M.J.; et al. Earth history and the passerine superradiation. Proc. Natl. Acad. Sci. USA 2019, 116, 7916–7925. [Google Scholar] [CrossRef] [Green Version]
- Owen, C.L.; Marshall, D.C.; Hill, K.B.R.; Simon, C. How the aridification of Australia structured the biogeography and influenced the diversification of a large lineage of Australian cicadas. Syst. Biol. 2017, 66, 569–589. [Google Scholar] [CrossRef]
- Brennan, I.G.; Oliver, P.M. Mass turnover and recovery dynamics of a diverse Australian continental radiation. Evol. Int. J. Org. Evol. 2017, 71, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, H.; Frankl-Vilches, C.; Bakker, A.; Mayr, G.; Nikolaus, G.; Boerno, S.T.; Klages, S.; Timmermann, B.; Gahr, M. An Unbiased Molecular Approach Using 3’-UTRs Resolves the Avian Family-Level Tree of Life. Mol. Biol. Evol. 2021, 38, 108–127. [Google Scholar] [CrossRef]
- Claramunt, S.; Cracraft, J. A new time tree reveals Earth history’s imprint on the evolution of modern birds. Sci. Adv. 2015, 1, e1501005. [Google Scholar] [CrossRef] [Green Version]
- Ito, H. How to activate heat-responsible retrotransposon ONSEN in Brassicaceae species. Methods Mol. Biol. 2021, 2250, 189–194. [Google Scholar]
- Jiao, Y.; Deng, X.W. A genome-wide transcriptional activity survey of rice transposable element-related genes. Genome Biol. 2007, 8, R28. [Google Scholar] [CrossRef] [Green Version]
- Lopes, F.R.; Jjingo, D.; da Silva, C.R.; Andrade, A.C.; Marraccini, P.; Teixeira, J.B.; Carazzolle, M.F.; Pereira, G.A.; Pereira, L.F.; Vanzela, A.L.; et al. Transcriptional activity, chromosomal distribution and expression effects of transposable elements in Coffea genomes. PLoS ONE 2013, 8, e78931. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A.; Al-Shomrani, B.M.; Simenc, M.; Alharbi, S.N.; Alqahtani, F.H.; Al-Fageeh, M.B.; Manee, M.M. Comparative analysis of transposable elements provides insights into genome evolution in the genus Camelus. BMC Genom. 2021, 22, 842. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, T.; Segawa, T.; Mori, H.; Campos, P.F.; Hongoh, Y.; Endo, H.; Akiyoshi, A.; Kohno, N.; Nishida, S.; Wu, J. Phylogenomics and morphology of extinct paleognaths reveal the origin and evolution of the ratites. Curr. Biol. 2017, 27, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Fänger, H. Avitaxonomicon Website. 2023. Available online: https://www.bird-phylogeny.de (accessed on 13 January 2022).
- Tavares, E.S.; Baker, A.J.; Pereira, S.L.; Miyaki, C.Y. Phylogenetic relationships and historical biogeography of neotropical parrots (Psittaciformes: Psittacidae: Arini) inferred from mitochondrial and nuclear DNA sequences. Syst. Biol. 2006, 55, 454–470. [Google Scholar] [CrossRef] [Green Version]
- Furo, I.O.; Kretschmer, R.; O’Brien, P.C.; Pereira, J.C.; Garnero, A.D.V.; Gunski, R.J.; O’Connor, R.E.; Griffin, D.K.; Gomes, A.J.B.; Ferguson-Smith, M.A.; et al. Chromosomal evolution in the phylogenetic context: A remarkable karyotype reorganization in neotropical parrot Myiopsitta monachus (psittacidae). Front. Genet. 2020, 11, 721. [Google Scholar] [CrossRef]
- Huang, Z.; De O Furo, I.; Liu, J.; Peona, V.; Gomes, A.J.B.; Cen, W.; Huang, H.; Zhang, Y.; Chen, D.; Xue, T.; et al. Recurrent chromosome reshuffling and the evolution of neo-sex chromosomes in parrots. Nat. Commun. 2022, 13, 944. [Google Scholar] [CrossRef]
- Miskey, C.; Izsvák, Z.; Kawakami, K.; Ivics, Z. DNA transposons in vertebrate functional genomics. Cell. Mol. Life Sci. 2005, 62, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Lohe, A.R.; Moriyama, E.N.; Lidholm, D.A.; Hartl, D.L. Horizontal trasmission, vertical inactivation, and stochastuc loss of mariner-like transable elements. Mol. Biol. Evol. 1995, 12, 62–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, A.L.; Jones, N.C.; Pevzner, P.A. De novo identification of repeat families in large genomes. Bioinformatics 2005, 21, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- The UniProt Consortium UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5–5. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 10 May 2020).
- Sundaram, V.; Wysocka, J. Transposable elements as a potent source of diverse cis-regulatory sequences in mammalian genomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschetti, R.; Palazzo, A.; Lorusso, P.; Viggiano, L.; Massimiliano Marsano, R. “What You Need, Baby, I Got It”: Transposable elements as suppliers of cis-operating sequences in Drosophila. Biology 2020, 9, 25. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superorder | Order | Species |
|---|---|---|
| Neognathae | Caprimulgiformes | Apus apus |
| Caprimulgus europaeus | ||
| Hemiprocne comata | ||
| Nyctibius grandis | ||
| Podargus strigoides | ||
| Charadriiformes | Pedionomus torquatus | |
| Columbiformes | Columba livia | |
| Streptopelia turtur | ||
| Galliformes | Alectura lathami | |
| Gallus gallus | ||
| Passeriformes | Ficedula albicollis | |
| Hirundo rustica | ||
| Passer domesticus | ||
| Taeniopygia guttata | ||
| Piciformes | Colaptes auratus | |
| Dryobates pubescens | ||
| Psittaciformes | Amazona aestiva | |
| Melopsittacus undulatus | ||
| Myiopsitta monachus | ||
| Strigops habroptila | ||
| Sphenisciformes | Aptenodytes forsteri | |
| Aptenodytes patagonicus | ||
| Eudyptes chrysolophus | ||
| Spheniscus humboldti | ||
| Spheniscus mendiculus | ||
| Palaeognathae | Casuariiformes | Casuarius casuarius |
| Dromaius novaehollandiae | ||
| Rheiformes | Rhea americana | |
| Struthioniformes | Struthio camelus australis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carotti, E.; Tittarelli, E.; Canapa, A.; Biscotti, M.A.; Carducci, F.; Barucca, M. LTR Retroelements and Bird Adaptation to Arid Environments. Int. J. Mol. Sci. 2023, 24, 6332. https://doi.org/10.3390/ijms24076332
Carotti E, Tittarelli E, Canapa A, Biscotti MA, Carducci F, Barucca M. LTR Retroelements and Bird Adaptation to Arid Environments. International Journal of Molecular Sciences. 2023; 24(7):6332. https://doi.org/10.3390/ijms24076332
Chicago/Turabian StyleCarotti, Elisa, Edith Tittarelli, Adriana Canapa, Maria Assunta Biscotti, Federica Carducci, and Marco Barucca. 2023. "LTR Retroelements and Bird Adaptation to Arid Environments" International Journal of Molecular Sciences 24, no. 7: 6332. https://doi.org/10.3390/ijms24076332
APA StyleCarotti, E., Tittarelli, E., Canapa, A., Biscotti, M. A., Carducci, F., & Barucca, M. (2023). LTR Retroelements and Bird Adaptation to Arid Environments. International Journal of Molecular Sciences, 24(7), 6332. https://doi.org/10.3390/ijms24076332