
An HIV-1/HIV-2 Chimeric Envelope Glycoprotein Generates Binding and Neutralising Antibodies against HIV-1 and HIV-2 Isolates

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Expression and Antigenicity of HIV1/HIV2 Chimeric Envelope
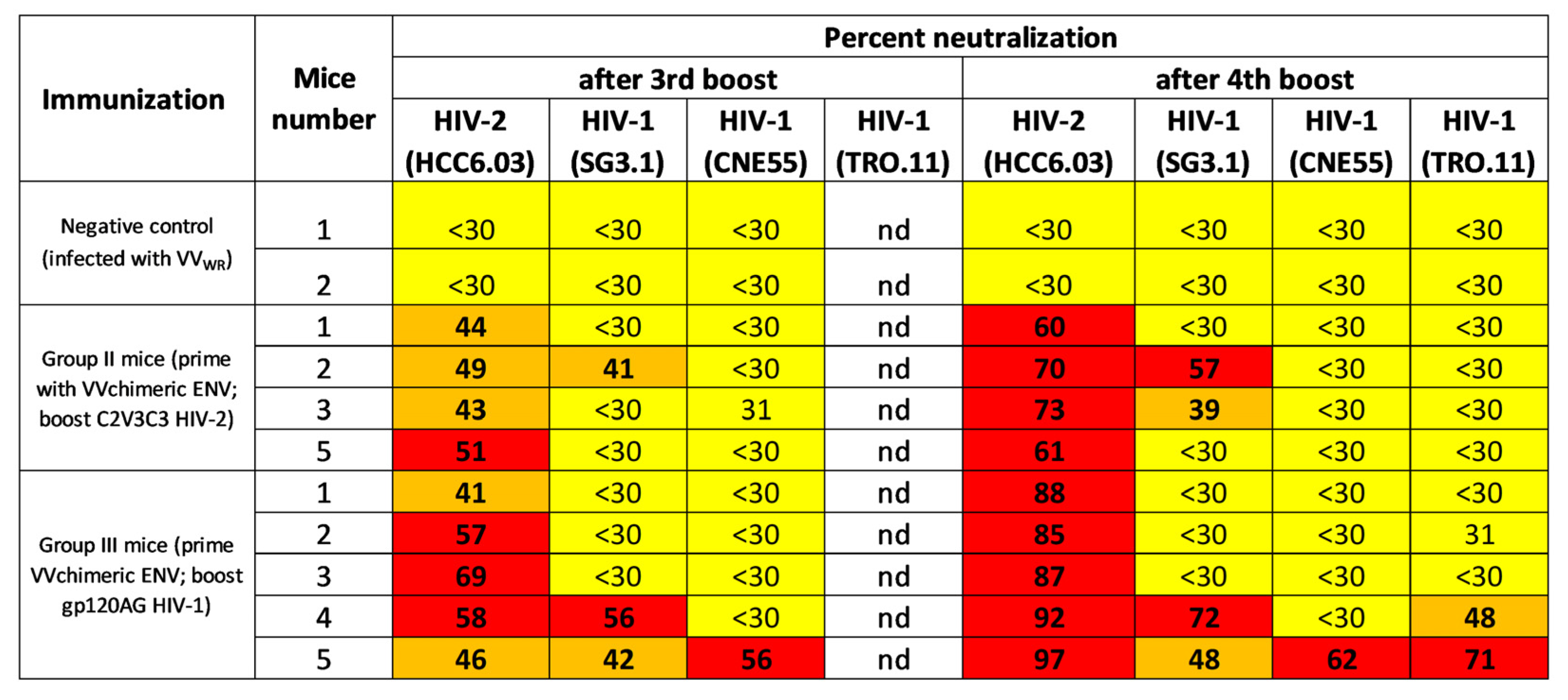
2.2. Chimeric Env Induces HIV-1 and HIV2 Binding and Neutralising Antibodies in Mice
2.3. Epitope Mapping Analyses
3. Discussion
4. Materials and Methods
4.1. Cells, Plasmids, Viruses, and Antibodies
4.2. Production and Expression of Recombinant Vaccinia Viruses
4.3. Characterization of Chimeric Envelope Glycoproteins
4.4. Balb/c Mice Immunizations
4.5. Envelope-Specific Antibody Binding in Mice Sera
4.6. Neutralization Assays
4.7. Epitope Mapping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faria, N.R.; Esbjörnsson, J.; Lemey, P. Phylogeographic Insights into the Origins and Epidemic History of the Human Immunodeficiency Virus Type 2. In Encyclopedia of AIDS; Springer Science: New York, NY, USA, 2013. [Google Scholar]
- van Tienen, C.; van der Loeff, M.S. Epidemiology of HIV-2 Infection in West Africa. In Encyclopedia of AIDS; Hope, T.J., Stevenson, M., Richman, D., Eds.; Springer: New York, NY, USA, 2016; pp. 1–11. [Google Scholar]
- UNAIDS Global HIV & AIDS Statistics—Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 12 May 2023).
- Collaborators, G.H. Global, regional, and national sex-specific burden and control of the HIV epidemic, 1990–2019, for 204 countries and territories: The Global Burden of Diseases Study 2019. Lancet HIV 2021, 8, e633–e651. [Google Scholar]
- De Lay, P.R.; Benzaken, A.; Karim, Q.A.; Aliyu, S.; Amole, C.; Ayala, G.; Chalkidou, K.; Chang, J.; Clayton, M.; Couto, A.; et al. Ending AIDS as a public health threat by 2030: Time to reset targets for 2025. PLoS Med. 2021, 18, e1003649. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Crotty, S. HIV vaccinology: 2021 update. Semin. Immunol. 2021, 51, 101470. [Google Scholar] [CrossRef] [PubMed]
- Landais, E.; Huang, X.; Havenar-Daughton, C.; Murrell, B.; Price, M.A.; Wickramasinghe, L.; Ramos, A.; Bian, C.B.; Simek, M.; Allen, S.; et al. Broadly Neutralizing Antibody Responses in a Large Longitudinal Sub-Saharan HIV Primary Infection Cohort. PLoS Pathog. 2016, 12, e1005369. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Hu, Y.; Zhao, C.; Gao, H.; Greene, K.M.; Ren, L.; Ma, L.; Ruan, Y.; Sarzotti-Kelsoe, M.; Montefiori, D.C.; et al. Profiling the neutralizing antibody response in chronically HIV-1 CRF07_BC-infected intravenous drug users naive to antiretroviral therapy. Sci. Rep. 2017, 7, 46308. [Google Scholar] [CrossRef] [PubMed]
- Rusert, P.; Kouyos, R.D.; Kadelka, C.; Ebner, H.; Schanz, M.; Huber, M.; Braun, D.L.; Hoze, N.; Scherrer, A.; Magnus, C.; et al. Determinants of HIV-1 broadly neutralizing antibody induction. Nat. Med. 2016, 22, 1260–1267. [Google Scholar] [CrossRef]
- Hraber, P.; Korber, B.T.; Lapedes, A.S.; Bailer, R.T.; Seaman, M.S.; Gao, H.; Greene, K.M.; McCutchan, F.; Williamson, C.; Kim, J.H.; et al. Impact of clade, geography, and age of the epidemic on HIV-1 neutralization by antibodies. J. Virol. 2014, 88, 12623–12643. [Google Scholar] [CrossRef]
- Tomaras, G.D.; Binley, J.M.; Gray, E.S.; Crooks, E.T.; Osawa, K.; Moore, P.L.; Tumba, N.; Tong, T.; Shen, X.; Yates, N.L.; et al. Polyclonal B cell responses to conserved neutralization epitopes in a subset of HIV-1-infected individuals. J. Virol. 2011, 85, 11502–11519. [Google Scholar] [CrossRef]
- Simek, M.D.; Rida, W.; Priddy, F.H.; Pung, P.; Carrow, E.; Laufer, D.S.; Lehrman, J.K.; Boaz, M.; Tarragona-Fiol, T.; Miiro, G.; et al. Human immunodeficiency virus type 1 elite neutralizers: Individuals with broad and potent neutralizing activity identified by using a high-throughput neutralization assay together with an analytical selection algorithm. J. Virol. 2009, 83, 7337–7348. [Google Scholar] [CrossRef]
- Saunders, K.O.; Nicely, N.I.; Wiehe, K.; Bonsignori, M.; Meyerhoff, R.R.; Parks, R.; Walkowicz, W.E.; Aussedat, B.; Wu, N.R.; Cai, F.; et al. Vaccine Elicitation of High Mannose-Dependent Neutralizing Antibodies against the V3-Glycan Broadly Neutralizing Epitope in Nonhuman Primates. Cell Rep. 2017, 18, 2175–2188. [Google Scholar] [CrossRef]
- Saunders, K.O.; Verkoczy, L.K.; Jiang, C.; Zhang, J.; Parks, R.; Chen, H.; Housman, M.; Bouton-Verville, H.; Shen, X.; Trama, A.M.; et al. Vaccine Induction of Heterologous Tier 2 HIV-1 Neutralizing Antibodies in Animal Models. Cell Rep. 2017, 21, 3681–3690. [Google Scholar] [CrossRef] [PubMed]
- Barouch, D.H.; Tomaka, F.L.; Wegmann, F.; Stieh, D.J.; Alter, G.; Robb, M.L.; Michael, N.L.; Peter, L.; Nkolola, J.P.; Borducchi, E.N.; et al. Evaluation of a mosaic HIV-1 vaccine in a multicentre, randomised, double-blind, placebo-controlled, phase 1/2a clinical trial (APPROACH) and in rhesus monkeys (NHP 13-19). Lancet 2018, 392, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Acharya, P.; Kong, R.; Cheng, C.; Chuang, G.Y.; Liu, K.; Louder, M.K.; O’Dell, S.; Rawi, R.; Sastry, M.; et al. Epitope-based vaccine design yields fusion peptide-directed antibodies that neutralize diverse strains of HIV-1. Nat. Med. 2018, 24, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Gorman, J.; Geng, H.; Liu, Q.; Lin, Y.; Tsybovsky, Y.; Go, E.P.; Dey, B.; Andine, T.; Kwon, A.; et al. Interdomain Stabilization Impairs CD4 Binding and Improves Immunogenicity of the HIV-1 Envelope Trimer. Cell Host Microbe 2018, 23, 832–844.e6. [Google Scholar] [CrossRef]
- Asbach, B.; Kibler, K.V.; Kostler, J.; Perdiguero, B.; Yates, N.L.; Stanfield-Oakley, S.; Tomaras, G.D.; Kao, S.F.; Foulds, K.E.; Roederer, M.; et al. Priming with a Potent HIV-1 DNA Vaccine Frames the Quality of Immune Responses prior to a Poxvirus and Protein Boost. J. Virol. 2019, 93, e01529-18. [Google Scholar] [CrossRef]
- Dubrovskaya, V.; Tran, K.; Ozorowski, G.; Guenaga, J.; Wilson, R.; Bale, S.; Cottrell, C.A.; Turner, H.L.; Seabright, G.; O’Dell, S.; et al. Vaccination with Glycan-Modified HIV NFL Envelope Trimer-Liposomes Elicits Broadly Neutralizing Antibodies to Multiple Sites of Vulnerability. Immunity 2019, 51, 915–929.e7. [Google Scholar] [CrossRef]
- Escolano, A.; Gristick, H.B.; Abernathy, M.E.; Merkenschlager, J.; Gautam, R.; Oliveira, T.Y.; Pai, J.; West, A.P., Jr.; Barnes, C.O.; Cohen, A.A.; et al. Immunization expands B cells specific to HIV-1 V3 glycan in mice and macaques. Nature 2019, 570, 468–473. [Google Scholar] [CrossRef]
- Francica, J.R.; Laga, R.; Lynn, G.M.; Muzikova, G.; Androvic, L.; Aussedat, B.; Walkowicz, W.E.; Padhan, K.; Ramirez-Valdez, R.A.; Parks, R.; et al. Star nanoparticles delivering HIV-1 peptide minimal immunogens elicit near-native envelope antibody responses in nonhuman primates. PLoS Biol. 2019, 17, e3000328. [Google Scholar] [CrossRef]
- Calado, R.; Duarte, J.; Borrego, P.; Marcelino, J.M.; Bartolo, I.; Martin, F.; Figueiredo, I.; Almeida, S.; Graca, L.; Vitor, J.; et al. A Prime-Boost Immunization Strategy with Vaccinia Virus Expressing Novel gp120 Envelope Glycoprotein from a CRF02_AG Isolate Elicits Cross-Clade Tier 2 HIV-1 Neutralizing Antibodies. Vaccines 2020, 8, 171. [Google Scholar] [CrossRef]
- Haynes, B.F.; Wiehe, K.; Borrow, P.; Saunders, K.O.; Korber, B.; Wagh, K.; McMichael, A.J.; Kelsoe, G.; Hahn, B.H.; Alt, F.; et al. Strategies for HIV-1 vaccines that induce broadly neutralizing antibodies. Nat. Rev. Immunol. 2023, 23, 142–158. [Google Scholar] [CrossRef]
- Burton, D.R.; Mascola, J.R. Antibody responses to envelope glycoproteins in HIV-1 infection. Nat. Immunol. 2015, 16, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, K.E.; Wagh, K.; Korber, B.; Barouch, D.H. Vaccines and Broadly Neutralizing Antibodies for HIV-1 Prevention. Annu. Rev. Immunol. 2020, 38, 673–703. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.E.; Bekker, L.G.; Laher, F.; Malahleha, M.; Allen, M.; Moodie, Z.; Grunenberg, N.; Huang, Y.; Grove, D.; Prigmore, B.; et al. Vaccine Efficacy of ALVAC-HIV and Bivalent Subtype C gp120-MF59 in Adults. N. Engl. J. Med. 2021, 384, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Pena, A.T.; Korzun, J.; et al. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef] [PubMed]
- Julien, J.P.; Lee, J.H.; Ozorowski, G.; Hua, Y.; Torrents de la Pena, A.; de Taeye, S.W.; Nieusma, T.; Cupo, A.; Yasmeen, A.; Golabek, M.; et al. Design and structure of two HIV-1 clade C SOSIP.664 trimers that increase the arsenal of native-like Env immunogens. Proc. Natl. Acad. Sci. USA 2015, 112, 11947–11952. [Google Scholar] [CrossRef]
- Sanders, R.W.; van Gils, M.J.; Derking, R.; Sok, D.; Ketas, T.J.; Burger, J.A.; Ozorowski, G.; Cupo, A.; Simonich, C.; Goo, L.; et al. HIV-1 VACCINES. HIV-1 neutralizing antibodies induced by native-like envelope trimers. Science 2015, 349, aac4223. [Google Scholar] [CrossRef]
- Pugach, P.; Ozorowski, G.; Cupo, A.; Ringe, R.; Yasmeen, A.; de Val, N.; Derking, R.; Kim, H.J.; Korzun, J.; Golabek, M.; et al. A native-like SOSIP.664 trimer based on an HIV-1 subtype B env gene. J. Virol. 2015, 89, 3380–3395. [Google Scholar] [CrossRef]
- Zhang, P.; Narayanan, E.; Liu, Q.; Tsybovsky, Y.; Boswell, K.; Ding, S.; Hu, Z.; Follmann, D.; Lin, Y.; Miao, H.; et al. A multiclade env-gag VLP mRNA vaccine elicits tier-2 HIV-1-neutralizing antibodies and reduces the risk of heterologous SHIV infection in macaques. Nat. Med. 2021, 27, 2234–2245. [Google Scholar] [CrossRef]
- Escolano, A.; Gristick, H.B.; Gautam, R.; DeLaitsch, A.T.; Abernathy, M.E.; Yang, Z.; Wang, H.; Hoffmann, M.A.G.; Nishimura, Y.; Wang, Z.; et al. Sequential immunization of macaques elicits heterologous neutralizing antibodies targeting the V3-glycan patch of HIV-1 Env. Sci. Transl. Med. 2021, 13, eabk1533. [Google Scholar] [CrossRef]
- Julg, B.; Barouch, D.H. Neutralizing antibodies for HIV-1 prevention. Curr. Opin. HIV AIDS 2019, 14, 318–324. [Google Scholar] [CrossRef]
- Cohen, Y.Z.; Caskey, M. Broadly neutralizing antibodies for treatment and prevention of HIV-1 infection. Curr. Opin. HIV AIDS 2018, 13, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Spencer, D.A.; Shapiro, M.B.; Haigwood, N.L.; Hessell, A.J. Advancing HIV Broadly Neutralizing Antibodies: From Discovery to the Clinic. Front. Public Health 2021, 9, 690017. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, S.; Luthra, K. An Overview of Human Anti-HIV-1 Neutralizing Antibodies against Diverse Epitopes of HIV-1. ACS Omega 2023, 8, 7252–7261. [Google Scholar] [CrossRef] [PubMed]
- Kwong, P.D.; Mascola, J.R. HIV-1 Vaccines Based on Antibody Identification, B Cell Ontogeny, and Epitope Structure. Immunity 2018, 48, 855–871. [Google Scholar] [CrossRef]
- Cizmeci, D.; Lofano, G.; Rossignol, E.; Dugast, A.S.; Kim, D.; Cavet, G.; Nguyen, N.; Tan, Y.C.; Seaman, M.S.; Alter, G.; et al. Distinct clonal evolution of B-cells in HIV controllers with neutralizing antibody breadth. Elife 2021, 10, e62648. [Google Scholar] [CrossRef]
- Visseaux, B.; Damond, F.; Matheron, S.; Descamps, D.; Charpentier, C. Hiv-2 molecular epidemiology. Infect. Genet. Evol. 2016, 46, 233–240. [Google Scholar] [CrossRef]
- Azevedo-Pereira, J.M.; Santos-Costa, Q. HIV Interaction With Human Host: HIV-2 As a Model of a Less Virulent Infection. AIDS Rev. 2016, 18, 44–53. [Google Scholar]
- Ceccarelli, G.; Giovanetti, M.; Sagnelli, C.; Ciccozzi, A.; d’Ettorre, G.; Angeletti, S.; Borsetti, A.; Ciccozzi, M. Human Immunodeficiency Virus Type 2: The Neglected Threat. Pathogens 2021, 10, 1377. [Google Scholar] [CrossRef]
- Matheron, S.; Pueyo, S.; Damond, F.; Simon, F.; Lepretre, A.; Campa, P.; Salamon, R.; Chene, G.; Brun-Vezinet, F. Factors associated with clinical progression in HIV-2 infected-patients: The French ANRS cohort. AIDS 2003, 17, 2593–2601. [Google Scholar] [CrossRef]
- Esbjornsson, J.; Mansson, F.; Kvist, A.; da Silva, Z.J.; Andersson, S.; Fenyo, E.M.; Isberg, P.E.; Biague, A.J.; Lindman, J.; Palm, A.A.; et al. Long-term follow-up of HIV-2-related AIDS and mortality in Guinea-Bissau: A prospective open cohort study. Lancet HIV 2018, 6, e25–e31. [Google Scholar] [CrossRef]
- Corey, L.; Gilbert, P.B.; Tomaras, G.D.; Haynes, B.F.; Pantaleo, G.; Fauci, A.S. Immune correlates of vaccine protection against HIV-1 acquisition. Sci. Transl. Med. 2015, 7, 310rv7. [Google Scholar] [CrossRef]
- Sadanand, S.; Suscovich, T.J.; Alter, G. Broadly Neutralizing Antibodies Against HIV: New Insights to Inform Vaccine Design. Annu. Rev. Med. 2016, 67, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, D.C.; Roederer, M.; Morris, L.; Seaman, M.S. Neutralization tiers of HIV-1. Curr. Opin. HIV AIDS 2018, 13, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, J.M.; Borrego, P.; Rocha, C.; Barroso, H.; Quintas, A.; Novo, C.; Taveira, N. Potent and broadly reactive HIV-2 neutralizing antibodies elicited by a vaccinia virus vector prime-C2V3C3 polypeptide boost immunization strategy. J. Virol. 2010, 84, 12429–12436. [Google Scholar] [CrossRef] [PubMed]
- Mikell, I.; Sather, D.N.; Kalams, S.A.; Altfeld, M.; Alter, G.; Stamatatos, L. Characteristics of the earliest cross-neutralizing antibody response to HIV-1. PLoS Pathog. 2011, 7, e1001251. [Google Scholar] [CrossRef]
- Marcelino, J.M.; Borrego, P.; Nilsson, C.; Familia, C.; Barroso, H.; Maltez, F.; Doroana, M.; Antunes, F.; Quintas, A.; Taveira, N. Resistance to antibody neutralization in HIV-2 infection occurs in late stage disease and is associated with X4 tropism. AIDS 2012, 26, 2275–2284. [Google Scholar] [CrossRef]
- Rocha, C.; Duarte, J.; Borrego, P.; Calado, R.; Marcelino, J.M.; Tendeiro, R.; Valadas, E.; Sousa, A.E.; Taveira, N. Potency of HIV-2-specific antibodies increase in direct association with loss of memory B cells. AIDS 2017, 31, 2431–2433. [Google Scholar] [CrossRef]
- Rocha, C.; Calado, R.; Borrego, P.; Marcelino, J.M.; Bartolo, I.; Rosado, L.; Cavaco-Silva, P.; Gomes, P.; Familia, C.; Quintas, A.; et al. Evolution of the human immunodeficiency virus type 2 envelope in the first years of infection is associated with the dynamics of the neutralizing antibody response. Retrovirology 2013, 10, 110. [Google Scholar] [CrossRef]
- Scheid, J.F.; Mouquet, H.; Ueberheide, B.; Diskin, R.; Klein, F.; Oliveira, T.Y.; Pietzsch, J.; Fenyo, D.; Abadir, A.; Velinzon, K.; et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011, 333, 1633–1637. [Google Scholar] [CrossRef]
- Pantophlet, R.; Ollmann Saphire, E.; Poignard, P.; Parren, P.W.; Wilson, I.A.; Burton, D.R. Fine mapping of the interaction of neutralizing and nonneutralizing monoclonal antibodies with the CD4 binding site of human immunodeficiency virus type 1 gp120. J. Virol. 2003, 77, 642–658. [Google Scholar] [CrossRef]
- McInerney, T.L.; McLain, L.; Armstrong, S.J.; Dimmock, N.J. A human IgG1 (b12) specific for the CD4 binding site of HIV-1 neutralizes by inhibiting the virus fusion entry process, but b12 Fab neutralizes by inhibiting a postfusion event. Virology 1997, 233, 313–326. [Google Scholar] [CrossRef]
- McKeating, J.A.; Thali, M.; Furman, C.; Karwowska, S.; Gorny, M.K.; Cordell, J.; Zolla-Pazner, S.; Sodroski, J.; Weiss, R.A. Amino acid residues of the human immunodeficiency virus type I gp120 critical for the binding of rat and human neutralizing antibodies that block the gp120-sCD4 interaction. Virology 1992, 190, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Davenport, Y.W.; West, A.P., Jr.; Bjorkman, P.J. Structure of an HIV-2 gp120 in Complex with CD4. J. Virol. 2016, 90, 2112–2118. [Google Scholar] [CrossRef] [PubMed]
- Yusim, K.; Korber, B.T.; Brander, C.; Barouch, D.; de Boer, R.; Haynes, B.F.; Koup, R.; Moore, J.P.; Walker, B.D.; Watkins, D. HIV Molecular Immunology 2015; Los Alamos National Lab. (LANL): Los Alamos, NM, USA, 2016.
- Zolla-Pazner, S.; Zhong, P.; Revesz, K.; Volsky, B.; Williams, C.; Nyambi, P.; Gorny, M.K. The cross-clade neutralizing activity of a human monoclonal antibody is determined by the GPGR V3 motif of HIV type 1. AIDS Res. Hum. Retrovir. 2004, 20, 1254–1258. [Google Scholar] [CrossRef]
- Gorny, M.K.; Conley, A.J.; Karwowska, S.; Buchbinder, A.; Xu, J.Y.; Emini, E.A.; Koenig, S.; Zolla-Pazner, S. Neutralization of diverse human immunodeficiency virus type 1 variants by an anti-V3 human monoclonal antibody. J. Virol. 1992, 66, 7538–7542. [Google Scholar] [CrossRef] [PubMed]
- Decker, J.M.; Bibollet-Ruche, F.; Wei, X.; Wang, S.; Levy, D.N.; Wang, W.; Delaporte, E.; Peeters, M.; Derdeyn, C.A.; Allen, S.; et al. Antigenic conservation and immunogenicity of the HIV coreceptor binding site. J. Exp. Med. 2005, 201, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Escolano, A.; Dosenovic, P.; Nussenzweig, M.C. Progress toward active or passive HIV-1 vaccination. J. Exp. Med. 2017, 214, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Borrego, P.; Marcelino, J.M.; Rocha, C.; Doroana, M.; Antunes, F.; Maltez, F.; Gomes, P.; Novo, C.; Barroso, H.; Taveira, N. The role of the humoral immune response in the molecular evolution of the envelope C2, V3 and C3 regions in chronically HIV-2 infected patients. Retrovirology 2008, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Lee, J.H.; Doores, K.J.; Murin, C.D.; Julien, J.P.; McBride, R.; Liu, Y.; Marozsan, A.; Cupo, A.; Klasse, P.J.; et al. Supersite of immune vulnerability on the glycosylated face of HIV-1 envelope glycoprotein gp120. Nat. Struct. Mol. Biol. 2013, 20, 796–803. [Google Scholar] [CrossRef]
- Daniels, C.N.; Saunders, K.O. Antibody responses to the HIV-1 envelope high mannose patch. Adv. Immunol. 2019, 143, 11–73. [Google Scholar]
- Sok, D.; Doores, K.J.; Briney, B.; Le, K.M.; Saye-Francisco, K.L.; Ramos, A.; Kulp, D.W.; Julien, J.P.; Menis, S.; Wickramasinghe, L.; et al. Promiscuous glycan site recognition by antibodies to the high-mannose patch of gp120 broadens neutralization of HIV. Sci. Transl. Med. 2014, 6, 236ra63. [Google Scholar] [CrossRef] [PubMed]
- Taveira, N.C.; Bex, F.; Burny, A.; Robertson, D.; Ferreira, M.O.; Moniz-Pereira, J. Molecular characterization of the env gene from a non-syncytium-inducing HIV-2 isolate (HIV-2ALI). AIDS Res. Hum. Retrovir. 1994, 10, 223–224. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Moss, B. New vaccinia virus recombination plasmids incorporating a synthetic late promoter for high level expression of foreign proteins. Nucleic Acids Res. 1990, 18, 4285–4286. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Rose, N.F.; Roberts, A.; Buonocore, L.; Rose, J.K. Glycoprotein exchange vectors based on vesicular stomatitis virus allow effective boosting and generation of neutralizing antibodies to a primary isolate of human immunodeficiency virus type 1. J. Virol. 2000, 74, 10903–10910. [Google Scholar] [CrossRef]
- Nandi, A.; Lavine, C.L.; Wang, P.; Lipchina, I.; Goepfert, P.A.; Shaw, G.M.; Tomaras, G.D.; Montefiori, D.C.; Haynes, B.F.; Easterbrook, P.; et al. Epitopes for broad and potent neutralizing antibody responses during chronic infection with human immunodeficiency virus type 1. Virology 2010, 396, 339–348. [Google Scholar] [CrossRef]
- Mkhize, N.N.; Durgiah, R.; Ashley, V.; Archary, D.; Garrett, N.J.; Karim, Q.A.; Karim, S.S.; Moore, P.L.; Yates, N.; Passmore, J.A.; et al. Broadly neutralizing antibody specificities detected in the genital tract of HIV-1 infected women. AIDS 2016, 30, 1005–1014. [Google Scholar] [CrossRef]
- deCamp, A.; Hraber, P.; Bailer, R.T.; Seaman, M.S.; Ochsenbauer, C.; Kappes, J.; Gottardo, R.; Edlefsen, P.; Self, S.; Tang, H.; et al. Global panel of HIV-1 Env reference strains for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 2014, 88, 2489–2507. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HuMAbs | Epitopes | Chimeric Env gp120 | M Con. S D-11 gp120 | SF162 gp140 |
|---|---|---|---|---|
| PG9 | V1V2 | 0.58 | 21.37 | 0.23 |
| PG16 | V2 | 0.91 | 1.66 | 0.28 |
| 447-52D | V3 | 1.52 | 23.05 | 16.05 |
| 2G12 | N-linked glycans | 0.58 | 23.05 | 16.05 |
| B12 | CD4bs | 2.00 | 23.05 | 16.05 |
| VRCO1 | CD4bs | 0.36 | 23.05 | 14.63 |
| VRCO3 | CD4bs | 0.38 | 1.03 | 15.59 |
| HJ16 | CD4bs | 0.90 | 23.05 | 0.71 |
| 3BNC117 | CD4bs | 0.46 | 23.05 | 12.54 |
| 2F5 | MPER (gp41) | 0.91 | 0.42 | 16.05 |
| HIV + | Multiple | 2.63 | 34.43 | 16.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taveira, N.; Figueiredo, I.; Calado, R.; Martin, F.; Bártolo, I.; Marcelino, J.M.; Borrego, P.; Cardoso, F.; Barroso, H. An HIV-1/HIV-2 Chimeric Envelope Glycoprotein Generates Binding and Neutralising Antibodies against HIV-1 and HIV-2 Isolates. Int. J. Mol. Sci. 2023, 24, 9077. https://doi.org/10.3390/ijms24109077
Taveira N, Figueiredo I, Calado R, Martin F, Bártolo I, Marcelino JM, Borrego P, Cardoso F, Barroso H. An HIV-1/HIV-2 Chimeric Envelope Glycoprotein Generates Binding and Neutralising Antibodies against HIV-1 and HIV-2 Isolates. International Journal of Molecular Sciences. 2023; 24(10):9077. https://doi.org/10.3390/ijms24109077
Chicago/Turabian StyleTaveira, Nuno, Inês Figueiredo, Rita Calado, Francisco Martin, Inês Bártolo, José M. Marcelino, Pedro Borrego, Fernando Cardoso, and Helena Barroso. 2023. "An HIV-1/HIV-2 Chimeric Envelope Glycoprotein Generates Binding and Neutralising Antibodies against HIV-1 and HIV-2 Isolates" International Journal of Molecular Sciences 24, no. 10: 9077. https://doi.org/10.3390/ijms24109077
APA StyleTaveira, N., Figueiredo, I., Calado, R., Martin, F., Bártolo, I., Marcelino, J. M., Borrego, P., Cardoso, F., & Barroso, H. (2023). An HIV-1/HIV-2 Chimeric Envelope Glycoprotein Generates Binding and Neutralising Antibodies against HIV-1 and HIV-2 Isolates. International Journal of Molecular Sciences, 24(10), 9077. https://doi.org/10.3390/ijms24109077