
UV-C Seed Surface Sterilization and Fe, Zn, Mg, Cr Biofortification of Wheat Sprouts as an Effective Strategy of Bioelement Supplementation

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
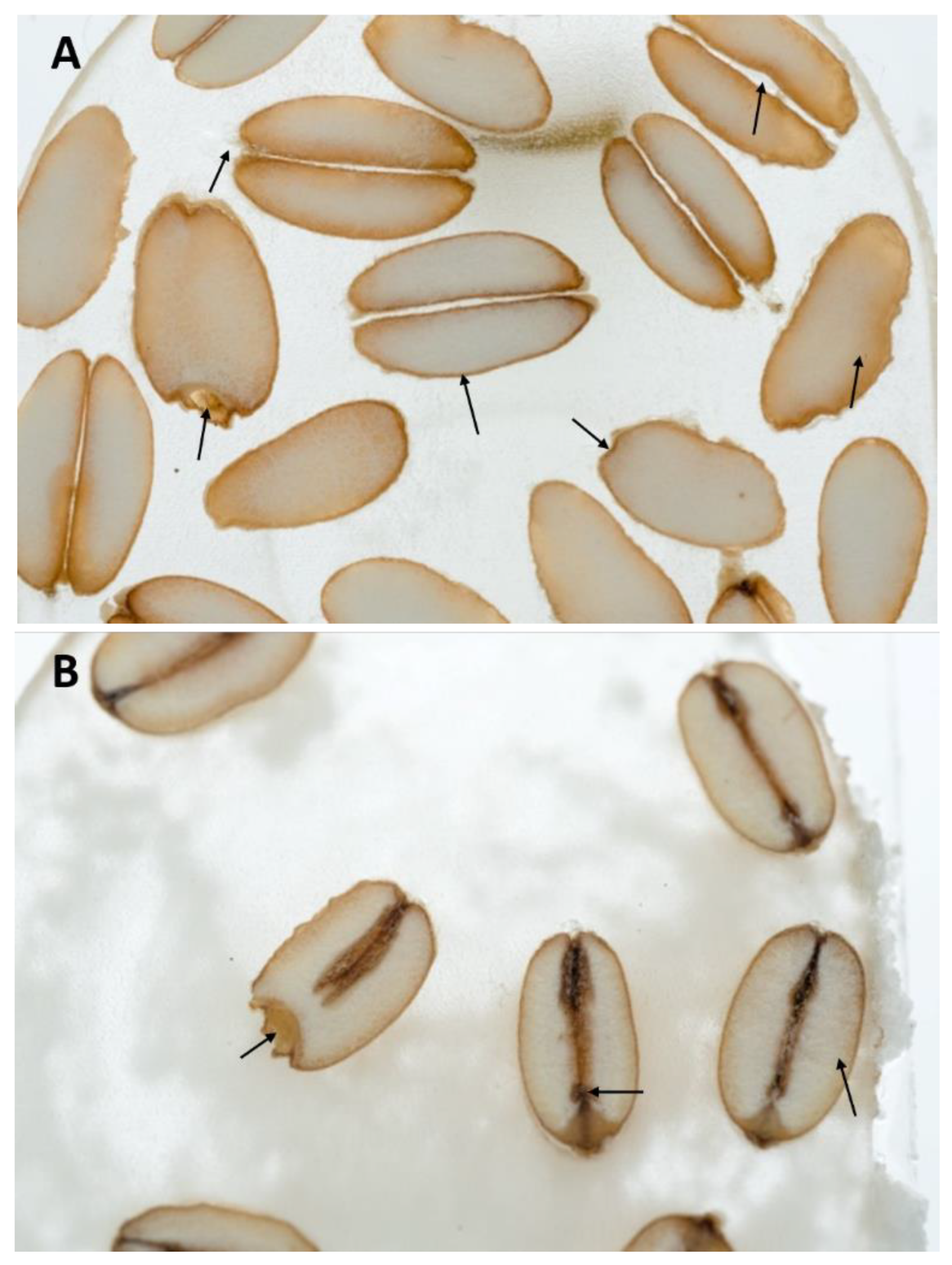
2.1. Sample Preparation and Macroscopic Analysis of Wheat Seeds
2.2. Seed Germination Energy
2.3. The Influence of Seed UV-C Sterilization on Biofortification Efficiency
2.4. Wheat Biofortification with Nutrients
3. Materials and Methods
3.1. Ion Solutions and AAS Standards
3.2. Plant Material
3.3. UV-C Exposure
3.4. Sample Preparation and Analysis of Wheat Seeds via Scanning Electron Microscopy (SEM)
3.5. Hydroponic Cultivation
3.6. Seed Germination Energy
3.7. Collection and Mineralization of Sprout Samples
3.8. The AAS Conditions
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sigel, A.; Sigel, H.; Sigel, R.K.O. Neurodegenerative Diseases and Metal Ions; John Wiley & Sons, Ltd.: Chichester, UK, 2006. [Google Scholar] [CrossRef]
- Lee, T.G.; Park, J.-W.; Shon, H.K.; Moon, D.W.; Choi, W.W.; Li, K.; Chung, J.H. Biochemical imaging of tissues by SIMS for biomedical applications. Appl. Surf. Sci. 2008, 255, 1241–1248. [Google Scholar] [CrossRef]
- Al-Fartusie, F.; Mohssan, S. Essential Trace Elements and Their Vital Roles in Human Body. Indian J. Adv. Chem. Sci. 2017, 5, 127–136. [Google Scholar]
- McDowell, L.R. Minerals in Animal and Human Nutrition, 2nd ed.; Elsevier Science: Amsterdam, The Netherlands, 2003; p. 660. [Google Scholar]
- Hurrell, R.F. Bioavailability of iron. Eur. J. Clin. Nutr. 1997, 51, 4–8. [Google Scholar]
- Osredkar, J. Copper and Zinc, Biological Role and Significance of Copper/Zinc Imbalance. J. Clin. Toxicol. S 2011, 3, 2161. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.S. Zinc deficiency: Has been known of for 40 years but ignored by global health organisations. BMJ 2003, 326, 409. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mertz, W. Chromium in human nutrition: A review. J. Nutr. 1993, 123, 626–633. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.B.; Lukaski, H.C. Chromium. Adv. Nutr. 2018, 9, 505–506. [Google Scholar] [CrossRef] [Green Version]
- Kostov, K.; Halacheva, L. Role of Magnesium Deficiency in Promoting Atherosclerosis, Endothelial Dysfunction, and Arterial Stiffening as Risk Factors for Hypertension. Int. J. Mol. Sci. 2018, 19, 1724. [Google Scholar] [CrossRef] [Green Version]
- Shenkin, A. Micronutrients in health and disease. Postgrad. Med. J. 2006, 82, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Biesalski, H.K.; Tinz, J. Micronutrients in the life cycle: Requirements and sufficient supply. NFS J. 2018, 11, 1–11. [Google Scholar] [CrossRef]
- Yin, X.; Yuan, L.; Lin, Z. Phytoremediation and Biofortification. Two Sides of One Coin. In Phytoremediation and Biofortification; Yin, X., Yuan, L., Eds.; Springer Briefs in Green Chemistry for Sustainability; Springer: Doetinchem, The Netherlands, 2012. [Google Scholar] [CrossRef]
- Huey, S.L.; Krisher, J.T.; Bhargava, A.; Friesen, V.M.; Konieczynski, E.M.; Mbuya, M.N.N.; Mehta, N.H.; Monterrosa, E.; Nyangaresi, A.M.; Mehta, S. Review of the Impact Pathways of Biofortified Foods and Food Products. Nutrients 2022, 14, 1200. [Google Scholar] [CrossRef] [PubMed]
- Beach, R.H.; Sulser, T.B.; Crimmins, A.; Cenacchi, N.; Cole, J.; Fukagawa, N.K.; Mason-D’Croz, D.; Myers, S.; Sarofim, M.C.; Smith, M.; et al. Combining the effects of increased atmospheric carbon dioxide on protein, iron, and zinc availability and projected climate change on global diets: A modelling study. Lancet Planet Health 2019, 3, e307–e317. [Google Scholar] [CrossRef] [Green Version]
- Combs, G.F., Jr. Selenium in global food systems. Brit. J. Nutr. 2001, 85, 517–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, G.; Nantel, G.; Shetty, P. The scourge of ‘hidden hunger’: Global dimensions of micronutrient deficiencies. Food Nutr. Agri. 2003, 32, 8–16. [Google Scholar]
- WHO; FAO. Better nutrition, better lives. In Proceedings of the Second International Conference on Nutrition (ICN2), Rome, Italy, 19–21 November 2014. [Google Scholar]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral Biofortification of Vegetables as a Tool to Improve Human Diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- Siwela, M.; Pillay, K.; Govender, L.; Lottering, S.; Mudau, F.N.; Modi, A.T.; Mabhaudhi, T. Biofortified crops for combating hidden hunger in South Africa: Availability, acceptability, micronutrient retention and bioavailability. Foods 2020, 9, 815. [Google Scholar] [CrossRef]
- Duborská, E.; Šebesta, M.; Matulová, M.; Zvěřina, O.; Urík, M. Current Strategies for Selenium and Iodine Biofortification in Crop Plants. Nutrients 2022, 14, 4717. [Google Scholar] [CrossRef]
- Blicharska, E.; Flieger, J.; Oszust, K.; Frąc, M.; Świeboda, R.; Kocjan, R. High-resolution continuum source atomic absorption spectrometry with microwave-assisted extraction for the determination of metals in vegetable sprouts. Anal. Lett. 2015, 48, 2272–2287. [Google Scholar] [CrossRef]
- Dunwell, J.M. Transgenic cereals: Current status and future prospects. J. Cereal Sci. 2014, 59, 419–434. [Google Scholar] [CrossRef]
- Kumar, S.; Palve, A.; Joshi, C.; Srivastava, R.K. Crop biofortification for iron (Fe), zinc (Zn) and vitamin A with transgenic approaches. Heliyon 2019, 5, e01914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Der Straeten, D.; Bhullar, N.K.; De Steur, H.; Gruissem, W.; MacKenzie, D.; Pfeiffer, W.; Bouis, H. Multiplying the efficiency and impact of biofortification through metabolic engineering. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Malik, K.A.; Maqbool, A. Transgenic crops for biofortification. Front. Sustain. Food Syst. 2020, 4, 182. [Google Scholar] [CrossRef]
- Koç, E.; Karayiğit, B. Assessment of Biofortification Approaches Used to Improve Micronutrient-Dense Plants That Are a Sustainable Solution to Combat Hidden Hunger. J. Soil Sci. Plant Nutr. 2022, 22, 475–500. [Google Scholar] [CrossRef]
- Lorenz, K.; D’Appolonia, B. Cereal sprouts: Composition, nutritive value, food applications. CRC Cr. Rev. Food Sci. 1980, 13, 353–385. [Google Scholar] [CrossRef]
- Park, S.A.; Grusak, M.A.; Oh, M.M. Concentrations of minerals and phenolic compounds in three edible sprout species treated with iron-chelates during imbibition. Hortic. Environ. Biotechnol. 2014, 55, 471–478. [Google Scholar] [CrossRef]
- Wei, Y.; Shohag, M.; Ying, F.; Yang, X.; Wu, C.; Wang, Y.; Shohag, J.I. Effect of ferrous sulfate fortification in germinated brown rice on seed iron concentration and bioavailability. Food Chem. 2013, 138, 1952–1958. [Google Scholar] [CrossRef]
- Zhu, H. Accumulation and distribution of selenium in different parts and macromolecule of Se-enriched Tartary Buckwheat (Fagopyrum tataricum Gaertn.) during germination. Int. Food Res. J. 2014, 21, 991–997. [Google Scholar]
- Liu, K.; Chen, F.; Zhao, Y.; Gu, Z.; Yang, H. Selenium accumulation in protein fractions during germination of Se-enriched brown rice and molecular weights distribution of Se-containing proteins. Food Chem. 2011, 127, 1526–1531. [Google Scholar] [CrossRef]
- Lazo-Vélez, M.A.; Avilés-González, J.; Serna-Saldivar, S.O.; Temblador-Pérez, M.C. Optimization of wheat sprouting for production of selenium enriched kernels using response surface methodology and desirability function. LWT-Food Sci. Technol. 2016, 65, 1080–1086. [Google Scholar] [CrossRef]
- Krzepiłko, A.; Zych-Wężyk, I.; Święciło, A.; Molas, J.; Skwaryło-Bednarz, B. Effect of iodine biofortification of lettuce seedlings on their mineral composition and biological quality. J. Elem. 2016, 21, 1071–1080. [Google Scholar] [CrossRef]
- Klepaka, T. Construcion of axial-symmetric polymetric extrudates of complex forms. Polimery 2008, 53, 390–395. (In Polish) [Google Scholar] [CrossRef]
- Arscott, S.; Goldman, I. Biomass effects and selenium accumulation in sprouts of three vegetable species grown in selenium-enriched conditions. Hortscience 2012, 47, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Zielińska-Dawidziak, M.; Siger, A. Effect of elevated accumulation of iron in ferritin on the antioxidants content in soybean sprouts. Eur. Food Res. Technol. 2012, 234, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Zou, T.; Xu, N.; Hu, G.; Pang, J.; Xu, H. Biofortification of soybean sprouts with zinc and bioaccessibility of zinc in the sprouts. J. Sci. Food Agric. 2014, 94, 3053–3060. [Google Scholar] [CrossRef]
- Aloo, S.O.; Ofosu, F.K.; Kilonzi, S.M.; Shabbir, U.; Oh, D.H. Edible Plant Sprouts: Health Benefits, Trends, and Opportunities for Novel Exploration. Nutrients 2021, 13, 2882. [Google Scholar] [CrossRef]
- He, W.; Yang, H.; Gu, G. Effect of Bacteria-mineral water produced from bio-reacted fowl dung on seed germination of wheat (Triticum aestivum) and rice (Oryza sativa L.). Environ. Progr. 2008, 27, 125–132. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, S.; Mishra, S.; Chauha, D.K.; Dubey, N.K. Micronutrients and their diverse role in agricultural crops: Advances and future prospective. Acta Physiol. Plant. 2015, 37, 139. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauha, D.K.; Dubey, N.K.; Rai, A.K. Silicon-mediated alleviation of Cr (VI) toxicity in wheat seedlings as evidenced by chlorophyll florescence, laser induced breakdown spectroscopy and anatomical changes. Ecotoxicol. Environ. Saf. 2015, 113, 133–144. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Rupiasih, N.N.; Vidyasagar, P.B. Effect of UV-C radiation and hypergravity on germination, growth and content of chlorophyll of wheat seedlings. AIP Conf. Proc. 2016, 1719, 030035-1–030035-6. [Google Scholar] [CrossRef]
- Partap, M.; Solanki, V.A. Impact of black point incited by Alternaria alternata on wheat trade, seed quality and seed germination. Indian Phytopathol. 2016, 69, 74–76. [Google Scholar]
- Rajput, M.A.; Pathan, M.A.; Lodhi, A.M.; Shah, G.S.; Khanzada, K.A. Studies on seed-borne fungi of wheat in Sindh Province and their effect on seed germination. Pak. J. Bot. 2005, 37, 181–185. [Google Scholar]
- Noble, R.E. Effects of UV-irradiation on seed germination. Sci. Total Environ. 2002, 299, 173–176. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Kot, A.; Szczerba, H.; Nowak, M.; Muszyńska, M. A review of conventional PCR assays for the detection of selected phytopathogens of wheat. J Mol. Microbiol. Biotechnol. 2017, 27, 175–189. [Google Scholar] [CrossRef]
- Singh, V.K.; Devi, A.; Pathania, S.; Kumar, V.; Tripathi, D.K.; Sharma, S.; Chauhan, D.K.; Singh, V.K.; Zorba, V. Spectroscopic investigation of wheat grains (Triticum aestivum) infected by wheat seed gall nematodes (Anguina tritici). Biocat. Agri. Biotech. 2017, 9, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Badridze, G.; Kacharava, N.; Chkhubianishvili, E.; Rapava, L.; Kikvidze, M.; Chanishvili, S.; Shakarishvili, N.; Mazanishvili, L.; Chigladze, L. Effect of UV radiation and artificial acid rain on productivity of wheat. Russ. J. Ecol. 2016, 47, 158–166. [Google Scholar] [CrossRef]
- Thomas, D.T.T.; Puthur, J.T. UV radiation priming: A means of amplifying the inherent potential for abiotic stress tolerance in crop plants. Environ. Exp. Bot. 2017, 138, 57–66. [Google Scholar] [CrossRef]
- Brown, J.; Lu, T.; Stevens, C.; Khan, V.; Lu, J.; Wilson, C.; Collins, D.; Wilson, M.; Igwegbe, E.; Chalutz, E.; et al. The effect of low dose ultraviolet light-C seed treatment on induced resistance in cabbage to black rot (Xanthomonas campestris pv. campestris). Crop Prot. 2001, 20, 873–883. [Google Scholar] [CrossRef]
- Kacharava, N.; Chanishvili, S.; Badridze, G.; Chkhubianishvili, E.; Janukashvili, N. Effect of seed irradiationon the content of antioxidants in leaves of kidney bean, cabbage and beet cultivars. Aust. J. Crop. Sci. 2009, 3, 137–145. [Google Scholar]
- Ouhibi, C.; Attia, H.; Rebah, F.; Msilini, N.; Chebbi, M.; Aarrouf, J.; Urban, L.; Lachaal, M. Salt stress mitigation by seed priming with UV-C in lettuce plants, growth, antioxidant activity and phenolic compounds. Plant Phys. Biochem. 2014, 83, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Shetta, N.D.; Areaf, I.M. Impact of ultraviolet-c radiation on seed germination and chlorophyll concentration of some woody trees grown in Saudi Arabia. J. Agric. Food Env. Sci. 2009, 8, 1–21. [Google Scholar]
- Brazaitytė, A.; Viršilė, A.; Jankauskienė, J.; Sakalauskienė, S.; Samuolienė, G.; Sirtautas, R.; Novičkovas, A.; Dabašinskas, L.; Miliauskienė, J.; Vaštakaitė, V.; et al. Effect of supplemental UV-A irradiation in solid-state lighting on the growth and phytochemical content of microgreens. Int. Agrophysics 2015, 29, 13–22. [Google Scholar] [CrossRef]
- Shukla, U.C.; Kakkar, P. Effect of dual stress of ultraviolet-B radiation and cadmium on nutrient uptake of wheat seedlings. Commun. Soil Sci. Plant. 2002, 33, 1737–1749. [Google Scholar] [CrossRef]
- Ilyas, M.; Khan, M.J.; Murad, Z.; Satti, S.Z.; Ullah, A. Biofortification of Iron in Wheat Varieties Using Different Methods of Application. Gesunde Pflanz. 2022. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Sadana, U.S.; Manchanda, J.S.; Dhadli, H. Biofortification of wheat grains with zinc and iron in Typic Ustochrept soils of Punjab. Indian J. Fertil. 2009, 5, 13–20. [Google Scholar]
- Dhaliwal, S.S.; Sharma, V.; Shukla, A.K.; Verma, V.; Kaur, M.; Shivay, Y.S.; Nisar, S.; Gaber, A.; Brestic, M.; Barek, V.; et al. Biofortification—A Frontier Novel Approach to Enrich Micronutrients in Field Crops to Encounter the Nutritional Security. Molecules 2022, 27, 1340. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Lu, L.-L.; Yang, X.-E.; Feng, Y.; Wei, Y.-Y.; Hao, H.-L.; Stoffella, P.J.; He, Z.-L. Uptake, translocation, and remobilization of zinc absorbed at different growth stages by rice genotypes of different Zn densities. J. Agric. Food Chem. 2010, 58, 6767–6773. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil. 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Phattarakul, N.; Rerkasem, B.; Li, L.J.; Wu, L.H.; Zou, C.Q.; Ram, H.; Sohu, V.S.; Kang, B.S.; Surek, H.; Kalayci, M.; et al. Biofortification of rice grain with zinc through zinc fertilization in different countries. Plant Soil. 2012, 361, 131–141. [Google Scholar] [CrossRef]
- Cakmak, I.; Kalayci, M.; Kaya, Y.; Torun, A.A.; Aydin, N.; Wang, Y.; Arisoy, Z.; Erdem, H.; Yazici, A.; Gokmen, O.; et al. Biofortification and localization of zinc in wheat grain. J. Agric. Food Chem. 2010, 58, 9092–9102. [Google Scholar] [CrossRef] [PubMed]
- Velu, G.; Ortiz-Monasterio, I.; Cakmak, I.; Hao, Y.; Singh, R.P. Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 2014, 59, 365–372. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Peterson, K.M.; Souza, E.J. Agronomic performance of low phytic acid wheat. Crop. Sci. 2006, 46, 2623–2629. [Google Scholar] [CrossRef]
- Liu, Z.H.; Wang, H.Y.; Zhang, G.P.; Chen, P.D.; Liu, D.J. Genotypic and spike positional difference in grain phytase activity, phytate, inorganic phosphorus, iron and zinc contents in wheat (Triticum aestivum L.). J. Cereal Sci. 2006, 44, 212–219. [Google Scholar] [CrossRef]
- Bouis, H.E.; Welch, R.M. Biofortification—A sustainable agricultural strategy for reducing micronutrient malnutrition in the Global South. Crop. Sci. 2010, 50, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Courtois, D.; Kasternmayer, P.; Clough, J.; Vigo, M.; Sabatier, M.; Arnaud, M.J. Magnesium enrichment and distribution in plants. Isot. Environ. Health Stud. 2003, 39, 273–279. [Google Scholar] [CrossRef]
- ISTA. International rules for seed testing. In Proceedings of the Ordinary General Meeting 2016, Tallinn, Estonia, 29 February 2016; Volume 2017. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion Concentration in Media (100 µg g−1) | Seed Germination Energy (%) | ||
|---|---|---|---|
| Variety A (Non-UV) | Variety A (UV) | Variety B (UV) | |
| 0 (control) | 100.0 ± 0.0 | 94.3 ± 0.6 | 95.3 ± 1.5 |
| Fe3+ | 98.3 ± 0.6 | 91.0 ± 0.0 | 91.0 ± 0.0 |
| Zn2+ | 97.0 ± 0.0 | 89.7 ± 1.2 | 92.0 ± 0.0 |
| Mg2+ | 97.7 ± 1.5 | 89.3 ± 0.6 | 90.3 ± 0.6 |
| Cr3+ | 88.0 ± 1.7 | 80.7 ± 1.5 | 79.3 ± 1.5 |
| Ion Concentration in Media (µg g−1) | Morphological Features of Sprout | |||
|---|---|---|---|---|
| Variety A (Non-UV) | Variety A (UV) | Variety B (UV) | ||
| Fe | 0 (control) | −/+ | −/+ | −/+ |
| 50 | −/+ | −/+ | −/+ | |
| 100 | + | + | + | |
| 200 | - | - | - | |
| Zn | 0 (control) | −/+ | −/+ | −/+ |
| 50 | −/+ | + | −/+ | |
| 100 | + | ++ | + | |
| 200 | −/+ | −/+ | - | |
| Mg | 0 (control) | −/+ | −/+ | −/+ |
| 50 | −/+ | −/+ | −/+ | |
| 100 | + | + | + | |
| 200 | + | + | + | |
| Cr | 0 (control) | −/+ | −/+ | −/+ |
| 50 | −/+ | −/+ | + | |
| 100 | −/+ | −/+ | + | |
| 200 | -- | -- | -- | |
| Elements | Concentrations of Elements (µg g−1) | ||
|---|---|---|---|
| 50 | 100 | 200 | |
| Iron (Fe) |  |  |  |
| Zinc (Zn) |  |  |  |
| Magnesium (Mg) |  |  |  |
| Chromium (Cr) |  |  |  |
| Control 1 |  | ||
| Mg Concentration in Media | BSE Micrograph with Graphic Arrangement of Measurement Points Made by SEM | Element Distribution Maps |
|---|---|---|
| 0 µg g−1 (Control) |  |  |
| 100 µg g−1 |  |  |
| Ion Concentration in Media (µg g−1) | Ion Concentration in Sprouts (µg g−1) | ||||
|---|---|---|---|---|---|
| 4 Days | 7 Days | ||||
| Variety A | Variety B | Variety A | Variety B | ||
| Fe | 0 (control) | 34.96 ± 3.96 c | 35.65 ± 0.85 d | 44.77 ± 2.30 c | 37.55 ± 2.43 c |
| 50 | 51.61 ± 1.36 bc | 53.03 ±2.83 c | 58.36 ± 3.02 c | 133.40 ± 8.37 b | |
| 100 | 75.89 ± 1.15 ab | 89.13 ± 9.79 b | 142.41 ± 3.91 b | 158.40 ± 9.26 a | |
| 200 | 85.20 ± 20.5 a | 114.96 ± 3.44 a | 163.27 ± 9.55 a | 163.47 ± 7.25 a | |
| Zn | 0 (control) | 22.30 ± 2.09 c | 16.11 ± 0.92 c | 19.54 ± 0.64 d | 16.72 ± 0.17 d |
| 50 | 118.79 ±10.48 b | 114.29 ± 1.96 b | 208.64 ± 45.71 c | 141.14 ±13.10 c | |
| 100 | 169.99 ±24.30 b | 117.44 ± 12.76 b | 301.32 ± 11.64 b | 208.92 ± 24.52 b | |
| 200 | 267.31 ±34.61 a | 305.12 ± 64.95 a | 560.53 ± 16.75 a | 320.06 ± 34.15 a | |
| Mg | 0 (control) | 1486.32 ± 7.92 c | 1464.48 ± 70.78 c | 1527.41 ± 60.30 d | 1473.89 ± 31.30 c |
| 50 | 1527.18 ±55.21 bc | 1657.53 ± 72.16 b | 1715.02 ± 13.35 c | 1625.96 ±57.53 c | |
| 100 | 1599.22 ±33.41 ab | 1717.31 ± 42.40 ab | 1928.44 ± 34.53 b | 1851.03 ± 92.55 b | |
| 200 | 1693.68 ± 39.14 a | 1813.90 ± 31.11 a | 2117.65 ± 111.59 a | 2015.14 ± 32.52 a | |
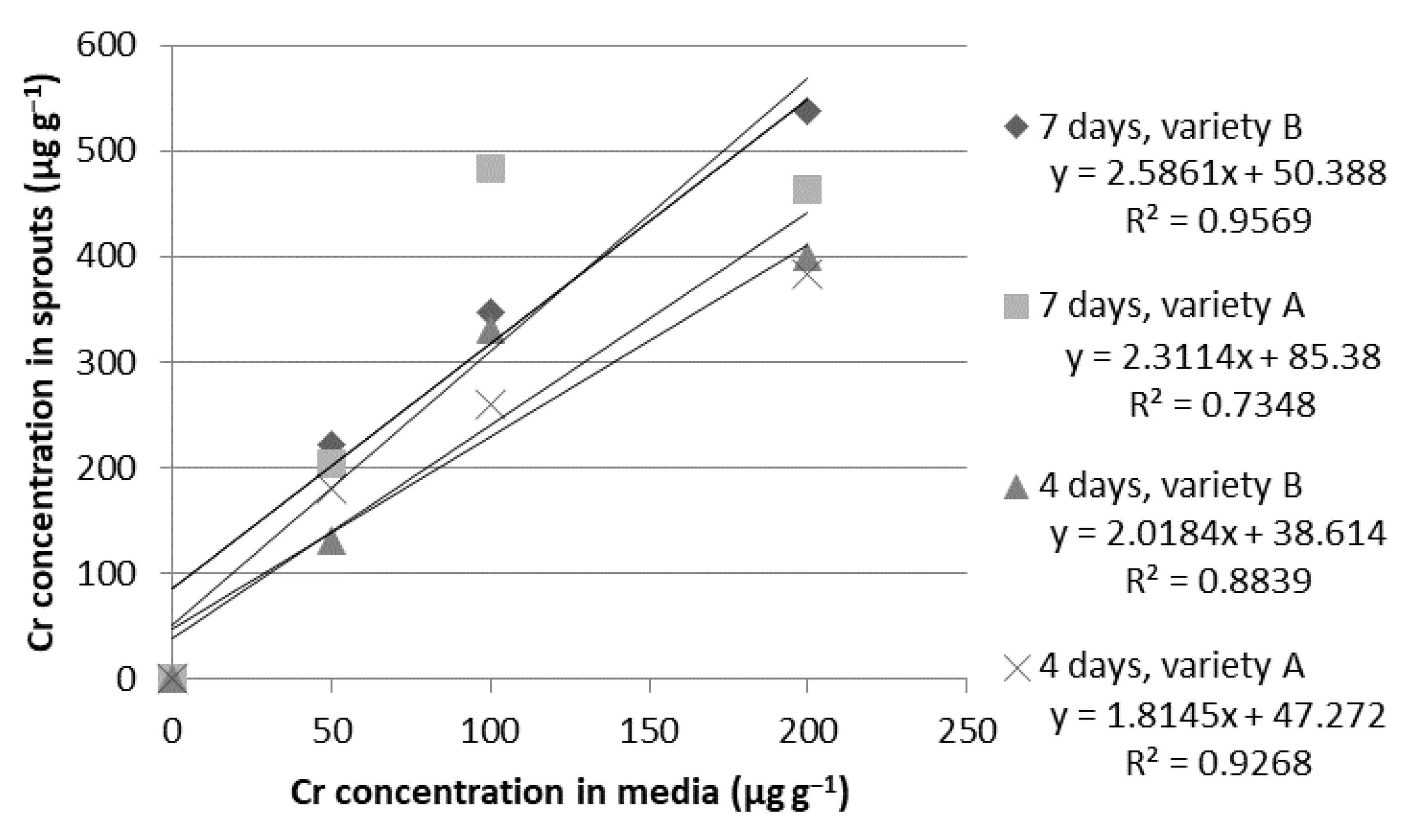
| Cr | 0 (control) | 0.00 ± 0.00 d | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 d |
| 50 | 179.79 ± 22.79 c | 130.42 ± 13.88 b | 203.60 ± 7.26 b | 221.64 ± 33.74 c | |
| 100 | 260.58 ± 19.13 b | 331.35 ± 10.82 a | 483.30 ± 28.46 a | 346.85 ± 63.34 b | |
| 200 | 383.80 ± 43.85 a | 399.12 ± 79.01 a | 463.60 ± 31.99 a | 538.19 ± 10.71 a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czarnek, K.; Tatarczak-Michalewska, M.; Dreher, P.; Rajput, V.D.; Wójcik, G.; Gierut-Kot, A.; Szopa, A.; Blicharska, E. UV-C Seed Surface Sterilization and Fe, Zn, Mg, Cr Biofortification of Wheat Sprouts as an Effective Strategy of Bioelement Supplementation. Int. J. Mol. Sci. 2023, 24, 10367. https://doi.org/10.3390/ijms241210367
Czarnek K, Tatarczak-Michalewska M, Dreher P, Rajput VD, Wójcik G, Gierut-Kot A, Szopa A, Blicharska E. UV-C Seed Surface Sterilization and Fe, Zn, Mg, Cr Biofortification of Wheat Sprouts as an Effective Strategy of Bioelement Supplementation. International Journal of Molecular Sciences. 2023; 24(12):10367. https://doi.org/10.3390/ijms241210367
Chicago/Turabian StyleCzarnek, Katarzyna, Małgorzata Tatarczak-Michalewska, Piotr Dreher, Vishnu D. Rajput, Grzegorz Wójcik, Anna Gierut-Kot, Agnieszka Szopa, and Eliza Blicharska. 2023. "UV-C Seed Surface Sterilization and Fe, Zn, Mg, Cr Biofortification of Wheat Sprouts as an Effective Strategy of Bioelement Supplementation" International Journal of Molecular Sciences 24, no. 12: 10367. https://doi.org/10.3390/ijms241210367
APA StyleCzarnek, K., Tatarczak-Michalewska, M., Dreher, P., Rajput, V. D., Wójcik, G., Gierut-Kot, A., Szopa, A., & Blicharska, E. (2023). UV-C Seed Surface Sterilization and Fe, Zn, Mg, Cr Biofortification of Wheat Sprouts as an Effective Strategy of Bioelement Supplementation. International Journal of Molecular Sciences, 24(12), 10367. https://doi.org/10.3390/ijms241210367