
Genome-Wide Identification and Analysis of the Plant Cysteine Oxidase (PCO) Gene Family in Brassica napus and Its Role in Abiotic Stress Response


Abstract
:1. Introduction
2. Results
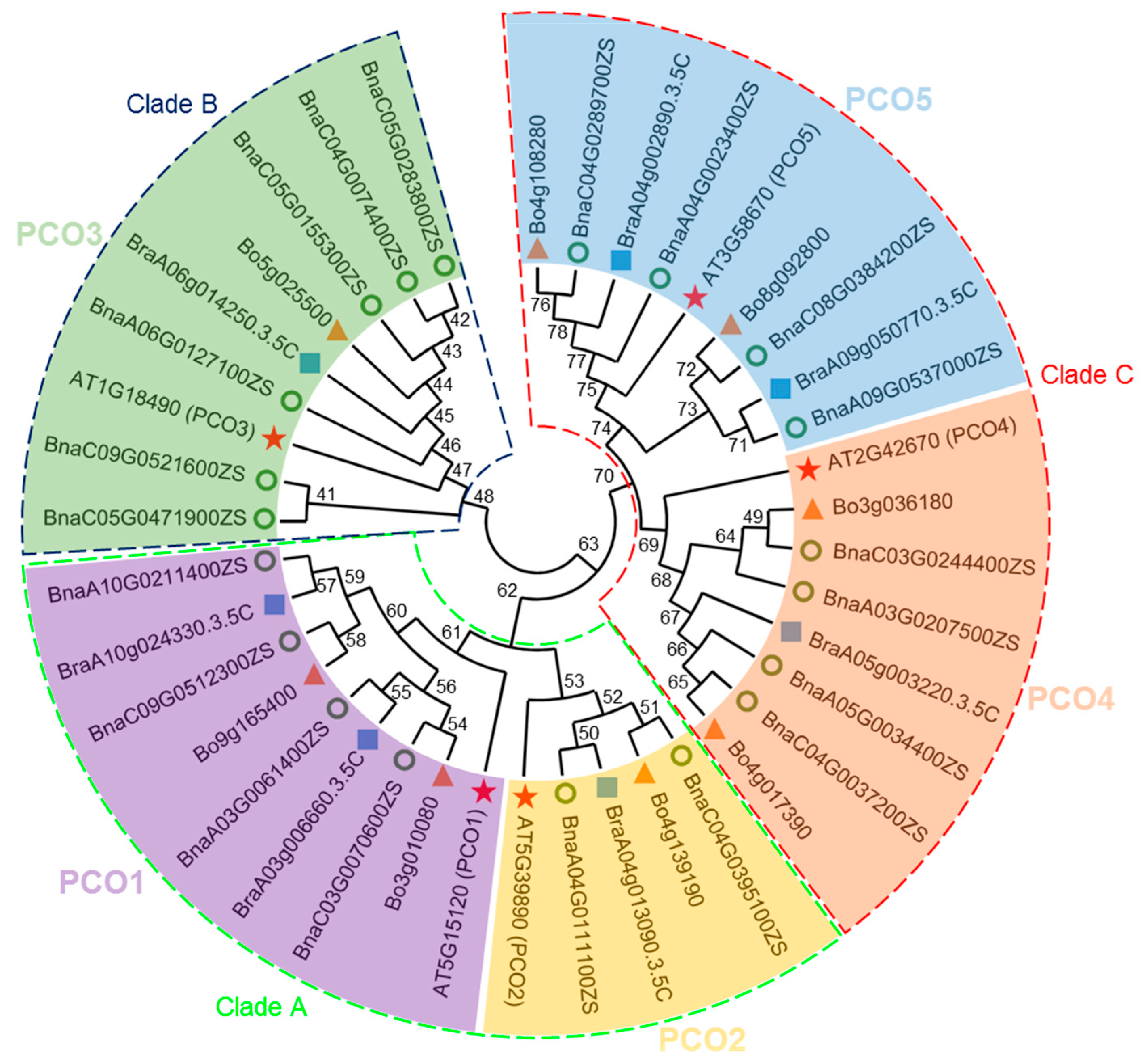
2.1. Identification and Classification of PCO Genes in B. napus, B. oleracea and B. rapa
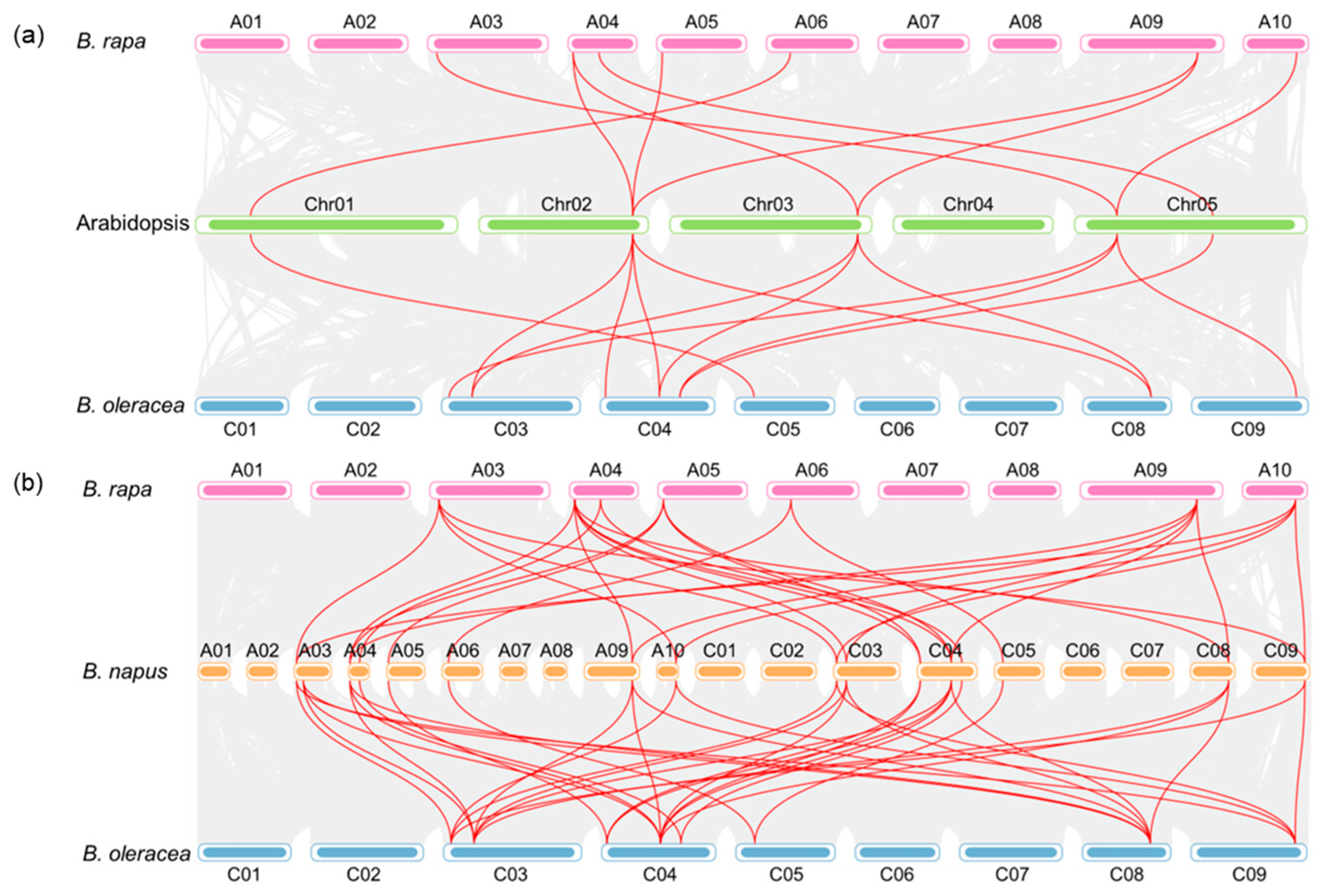
2.2. Chromosomal Distribution and Duplication of BnaPCOs
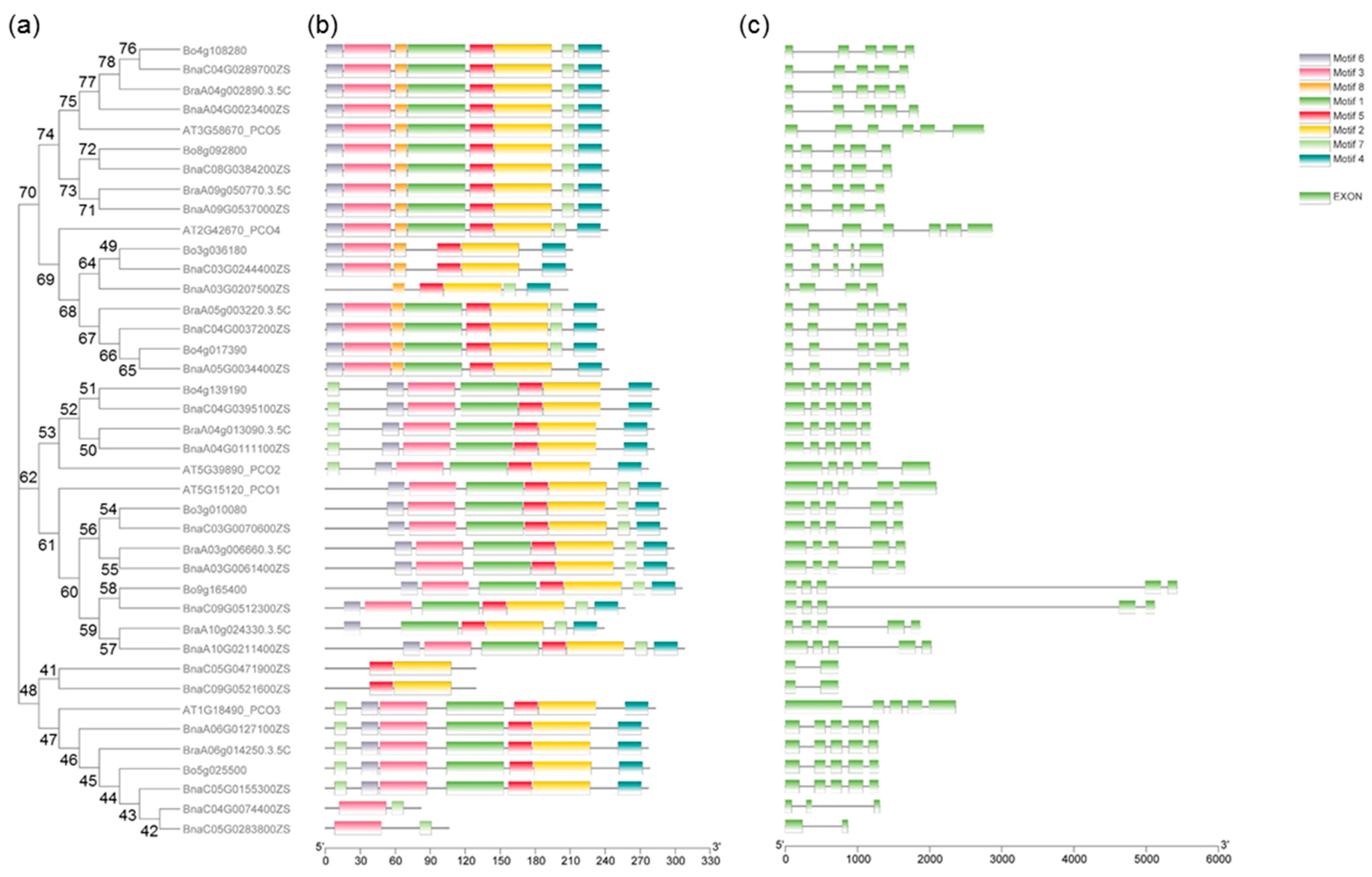
2.3. Gene Structures and Motif Analysis of PCOs in B. napus
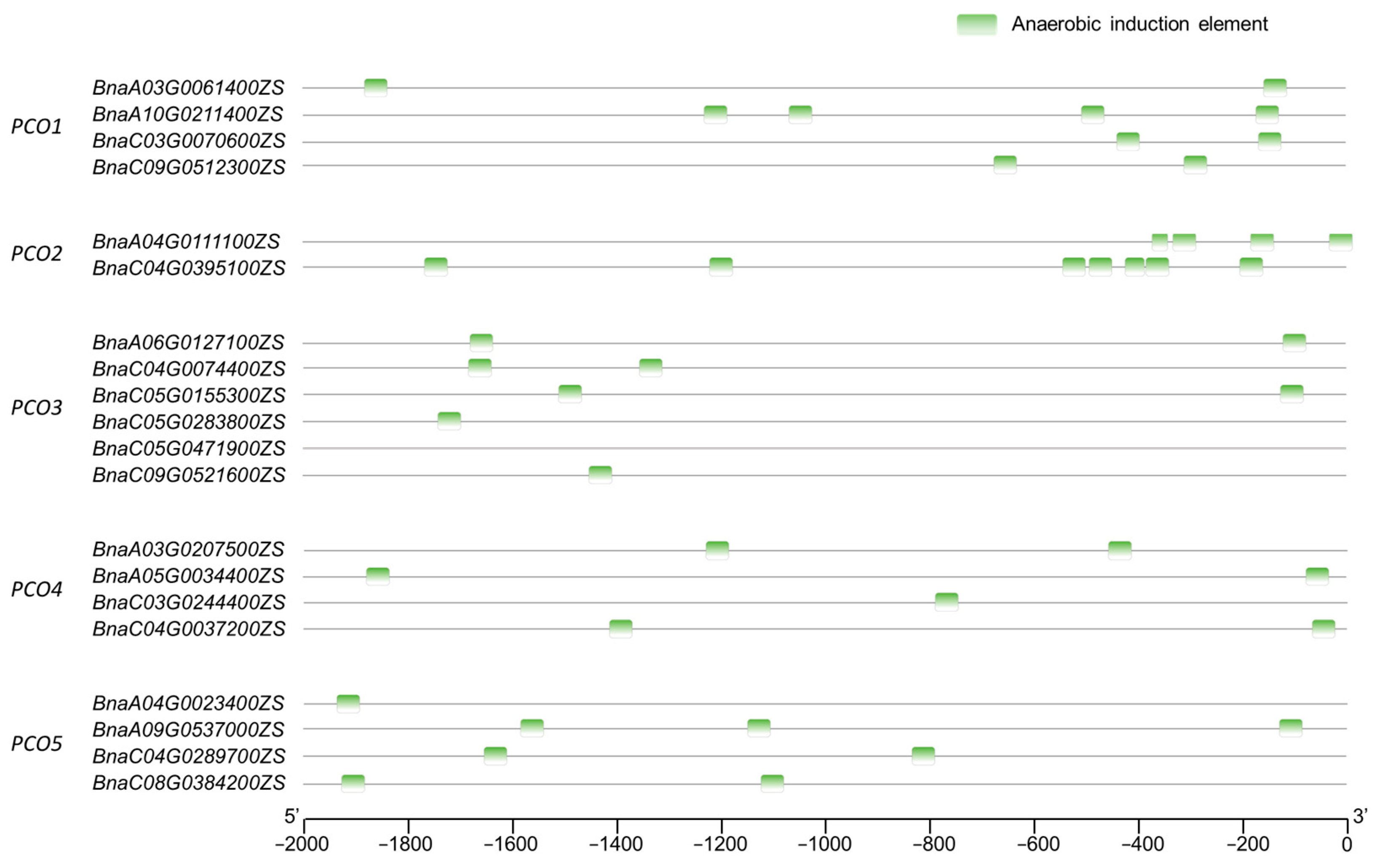
2.4. Cis-Element Analysis of BnaPCOs
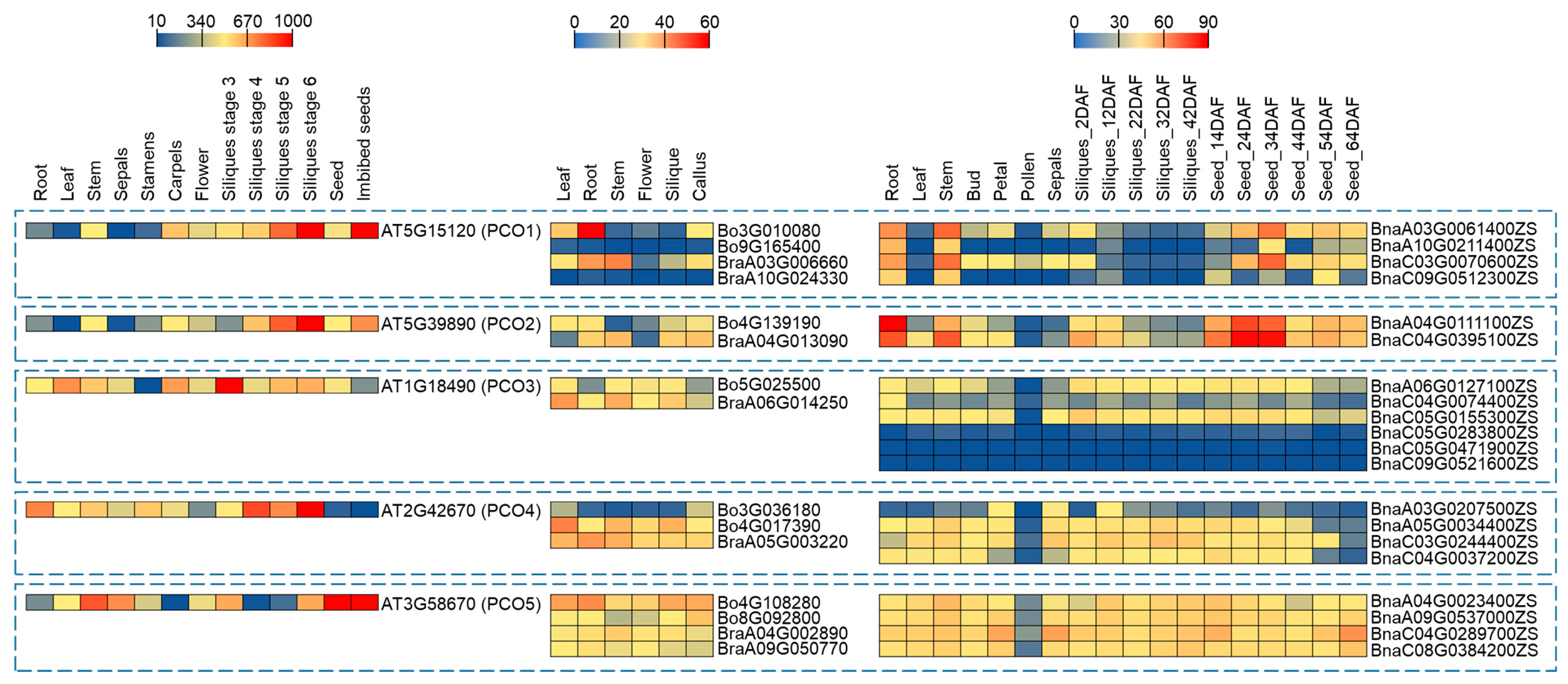
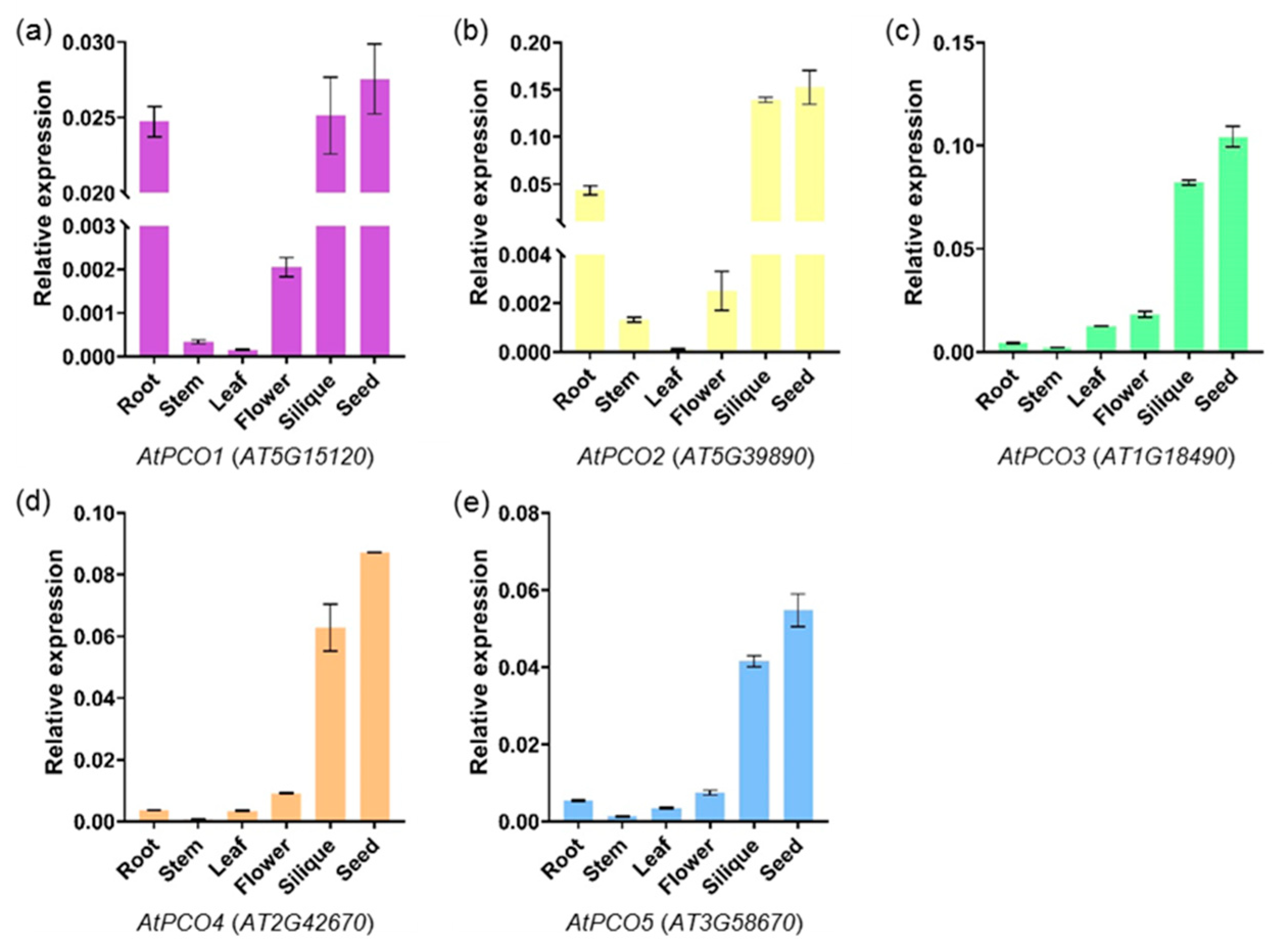
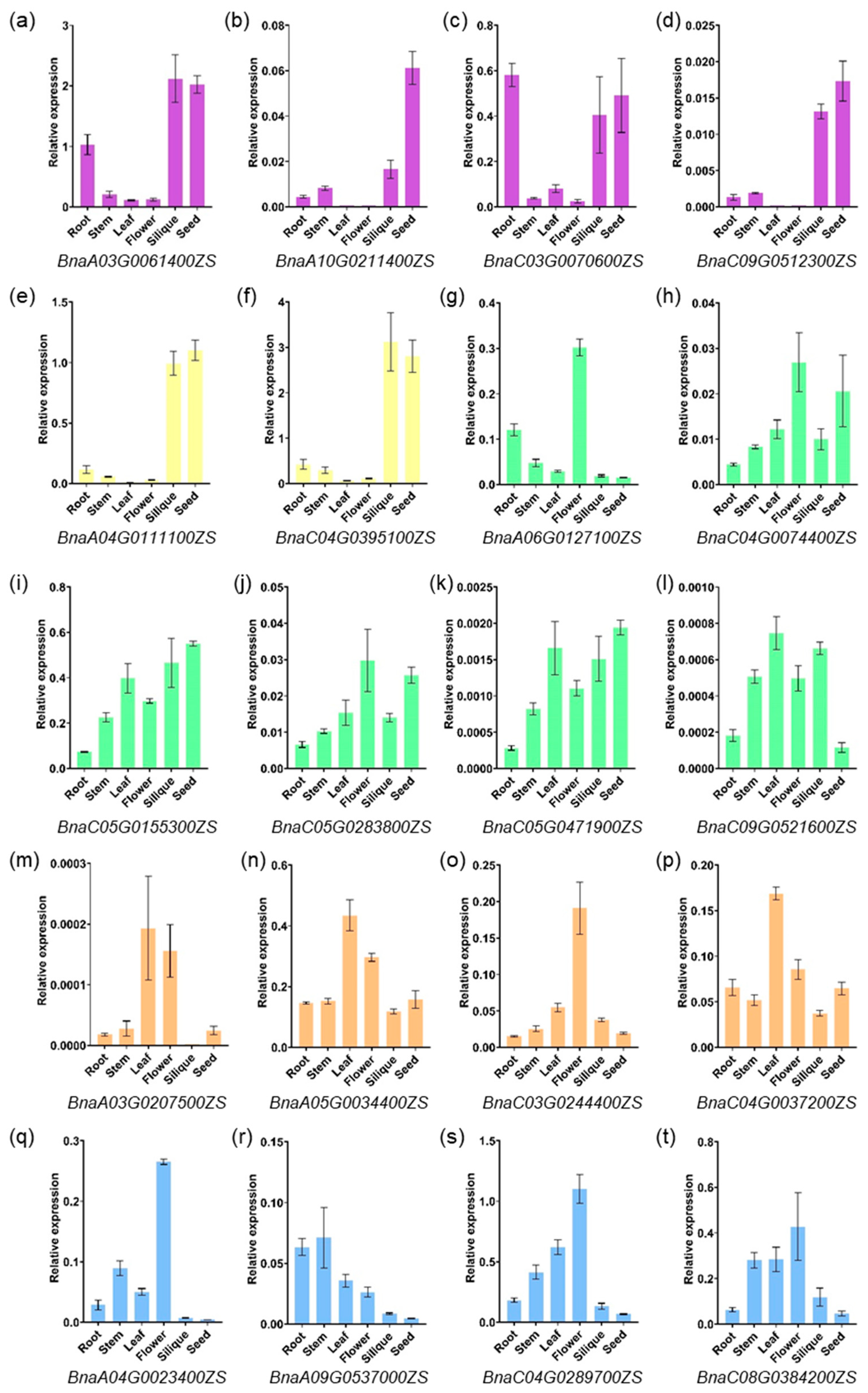
2.5. Expression Profiling of PCO Genes in Different Tissues
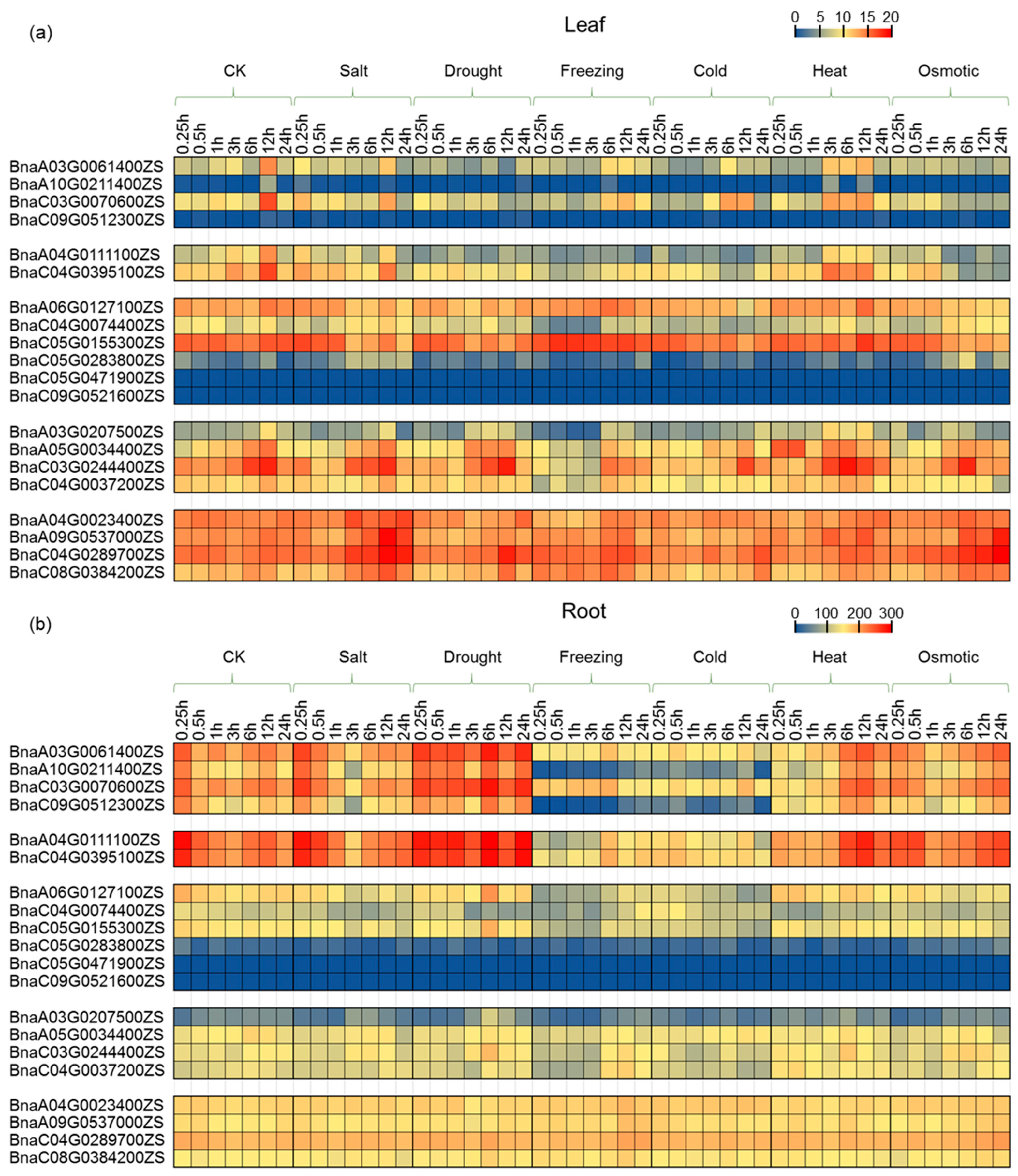
2.6. Expression Profiling of PCO Genes under Abiotic Stress Treatment
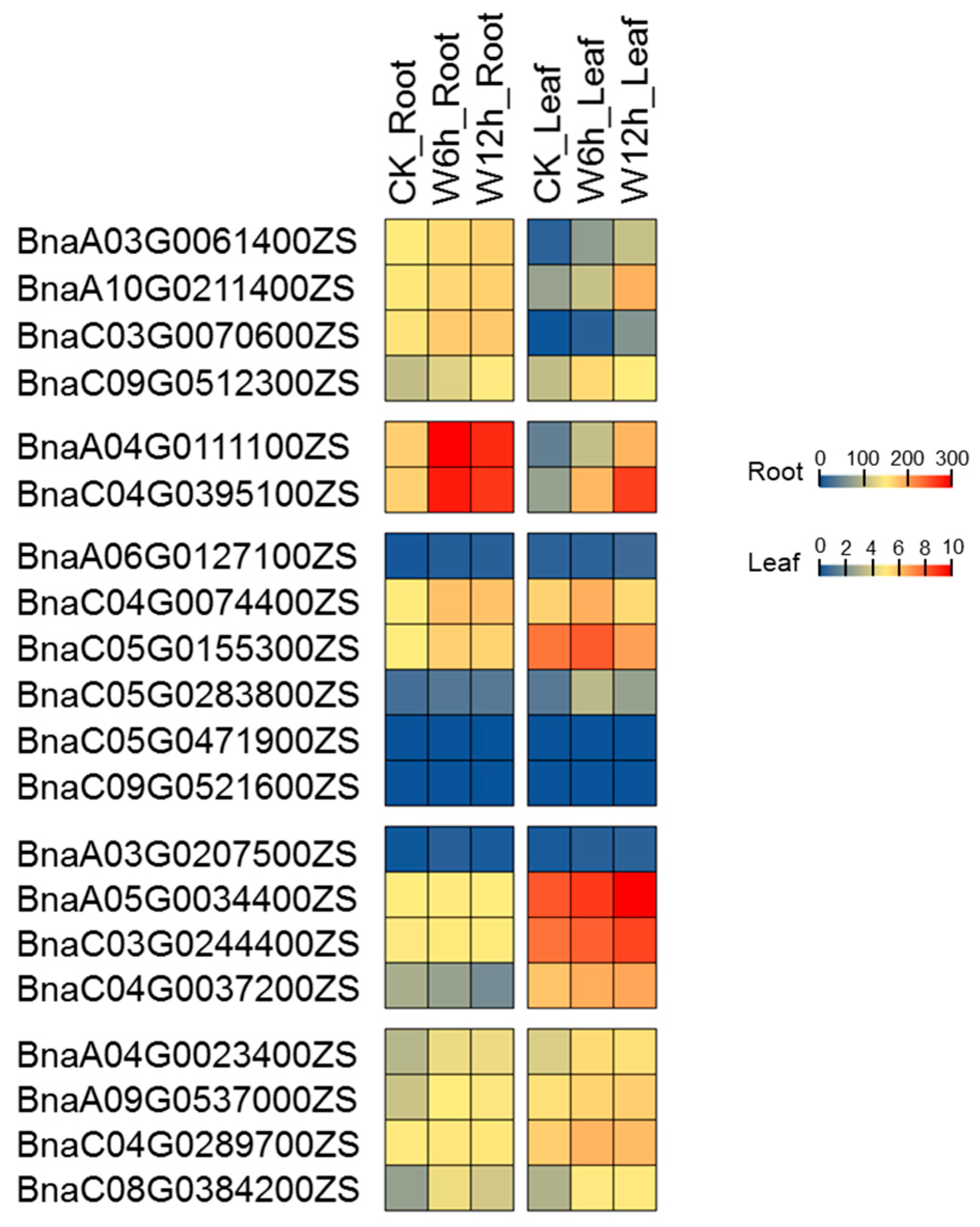
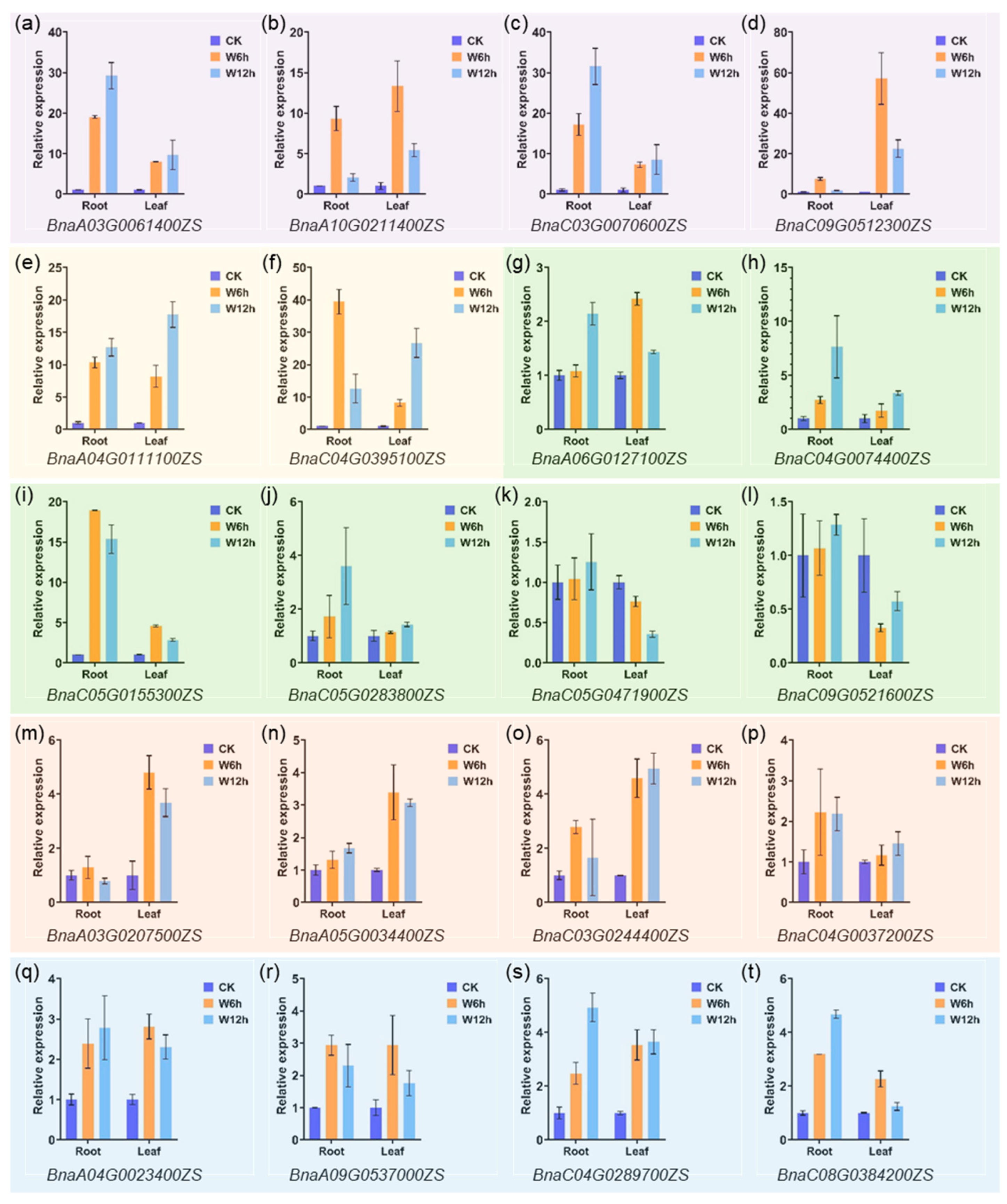
2.7. Expression Profiling of PCO Genes under Waterlogging Stress
3. Discussion
4. Materials and Methods
4.1. Identification of the PCO Gene Family
4.2. Phylogenetic Analysis, Chromosomal Locations and Syntenic Relationship
4.3. Analysis of Gene Structure, Motif Composition and Cis-Element
4.4. Plant Materials and Treatments
4.5. RNA-Seq and Heat Map Analysis of the PCO Transcriptome Data
4.6. Quantitative Real-Time RT-PCR (qRT-PCR) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chung, H.S.; Wang, S.B.; Venkatraman, V.; Murray, C.I.; Van, E.J.E. Cysteine oxidative posttranslational modifications: Emerging regulation in the cardiovascular system. Circ. Res. 2013, 112, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Romero, L.C.; Aroca, M.Á.; Laureano-Marín, A.M.; Moreno, I.; García, I.; Gotor, C. Cysteine and cysteine-related signaling pathways in Arabidopsis thaliana. Mol. Plant 2014, 7, 264–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, A.J.; Kim, S.B.; Ji, C.H.; Han, D.; Lee, S.J.; Lee, S.H.; Lee, M.J.; Lee, J.S.; Ciechanover, A.; Kim, B.Y.; et al. The N-terminal cysteine is a dual sensor of oxygen and oxidative stress. Proc. Natl. Acad. Sci. USA 2021, 118, e2107993118. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.J.; Tedds, H.M.; Labandera, A.M.; Bailey, M.; White, M.D.; Hartman, S.; Sprigg, C.; Mogg, S.L.; Osborne, R.; Dambire, C.; et al. Oxygen-dependent proteolysis regulates the stability of angiosperm polycomb repressive complex 2 subunit VERNALIZATION 2. Nat. Commun. 2018, 9, 5438. [Google Scholar] [CrossRef] [Green Version]
- Weits, D.A.; Kunkowska, A.B.; Kamps, N.C.W.; Portz, K.M.S.; Packbier, N.K.; Nemec, V.Z.; Gaillochet, C.; Lohmann, J.U.; Pedersen, O.; Dongen, J.T.; et al. An apical hypoxic niche sets the pace of shoot meristem activity. Nature 2019, 569, 714–717. [Google Scholar] [CrossRef]
- White, M.D.; Klecker, M.; Hopkinson, R.J.; Weits, D.A.; Mueller, C.; Naumann, C.; O’Neill, R.; Wickens, J.; Yang, J.; Brooks-Bartlett, J.C.; et al. Plant cysteine oxidases are dioxygenases that directly enable arginyl transferase-catalysed arginylation of N-end rule targets. Nat. Commun. 2017, 8, 14690. [Google Scholar] [CrossRef] [Green Version]
- Weits, D.A.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Hubberten, H.M.; Riegler, H.; Hoefgen, R.; Perata, P.; Dongen, J.T.; Licausi, F. Plant cysteine oxidases control the oxygen-dependent branch of the N-end-rule pathway. Nat. Commun. 2014, 5, 3425. [Google Scholar] [CrossRef] [Green Version]
- Licausi, F.; Dongen, J.T.; Giuntoli, B.; Novi, G.; Santaniello, A.; Geigenberger, P.; Perata, P. HRE1 and HRE2, two hypoxia-inducible ethylene response factors, affect anaerobic responses in Arabidopsis thaliana. Plant J. 2010, 62, 302–315. [Google Scholar] [CrossRef]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Wilson, I.W.; Yang, J.; Buerstenbinder, K.; Llewellyn, D.; Dennis, E.S.; Sauter, M.; Dolferus, R. Arabidopsis RAP2.2: An ethylene response transcription factor that is important for hypoxia survival. Plant Physiol. 2010, 153, 757–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.D.; Kamps, J.J.A.G.; East, S.; Taylor, K.L.J.; Flashman, E. The plant cysteine oxidases from Arabidopsis thaliana are kinetically tailored to act as oxygen sensors. J. Biol. Chem. 2018, 293, 11786–11795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.D.; Dalle, C.L.; Lavilla, P.M.; Iacopino, S.; Edwards, M.; Dunne, K.; Pires, E.; Levy, C.; McDonough, M.A.; Licausi, F.; et al. Structures of Arabidopsis thaliana oxygen-sensing plant cysteine oxidases 4 and 5 enable targeted manipulation of their activity. Proc. Natl. Acad. Sci. USA 2020, 117, 23140–23147. [Google Scholar] [CrossRef]
- Masson, N.; Keeley, T.P.; Giuntoli, B.; White, M.D.; Puerta, M.L.; Perata, P.; Hopkinson, R.J.; Flashman, E.; Licausi, F.; Ratcliffe, P.J. Conserved N-terminal cysteine dioxygenases transduce responses to hypoxia in animals and plants. Science 2019, 365, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Kearney, L.J.; Madden, S.; Wilson, J.; Myers, W.K.; Gunawardana, D.M.; Pires, E.; Holdship, P.; Tumber, A.; Rickaby, R.E.M.; Flashman, E. Plant cysteine oxidase oxygen-sensing function is conserved in early land plants and algae. ACS Bio Med. Chem. Au 2022, 2, 521–528. [Google Scholar] [CrossRef]
- Taylor-Kearney, L.J.; Flashman, E. Targeting plant cysteine oxidase activity for improved submergence tolerance. Plant J. 2022, 109, 779–788. [Google Scholar] [CrossRef]
- Dirr, A.; Gunawardana, D.M.; Flashman, E. Kinetic measurements to investigate the oxygen-sensing properties of plant cysteine oxidases. Methods Mol. Biol. 2023, 2648, 207–230. [Google Scholar] [PubMed]
- Chen, Z.; Guo, Q.; Wu, G.; Wen, J.; Liao, S.; Xu, C. Molecular basis for cysteine oxidation by plant cysteine oxidases from Arabidopsis thaliana. J. Struct. Biol. 2021, 213, 107663. [Google Scholar] [CrossRef]
- Weits, D.A.; Zhou, L.; Giuntoli, B.; Carbonare, L.D.; Iacopino, S.; Piccinini, L.; Lombardi, L.; Shukla, V.; Bui, L.T.; Novi, G.; et al. Acquisition of hypoxia inducibility by oxygen sensing N-terminal cysteine oxidase in spermatophytes. Plant Cell Environ. 2023, 46, 322–338. [Google Scholar] [CrossRef]
- Xu, K.; Xu, X.; Fukao, T.; Canlas, P.; Maghirang-Rodriguez, R.; Heuer, S.; Ismail, A.M.; Bailey-Serres, J.; Ronald, P.C.; Mackill, D.J. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature 2006, 442, 705–708. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wu, J.; Wang, X. Genome triplication drove the diversification of Brassica plants. Hortic. Res. 2014, 1, 14024. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Chen, B.; Zhao, J.; Zhang, F.; Xie, T.; Xu, K.; Gao, G.; Yan, G.; Li, H.; Li, L.; et al. Genomic selection and genetic architecture of agronomic traits during modern rapeseed breeding. Nat. Genet. 2022, 54, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Plant genetics. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Li, Z.; Liu, Y.; Li, W. Role of tillage measures in mitigating waterlogging damage in rapeseed. BMC Plant Biol. 2023, 23, 231. [Google Scholar] [CrossRef] [PubMed]
- Kayum, M.A.; Park, J.I.; Nath, U.K.; Biswas, M.K.; Kim, H.T.; Nou, I.S. Genome-wide expression profiling of aquaporin genes confer responses to abiotic and biotic stresses in Brassica rapa. BMC Plant Biol. 2017, 17, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Wu, J.; Qi, W.; Coulter, J.A.; Fang, Y.; Li, X.; Liu, L.; Jin, J.; Niu, Z.; Yue, J.; et al. Screening and verification of reference genes for analysis of gene expression in winter rapeseed (Brassica rapa L.) under abiotic stress. PLoS ONE 2020, 15, e0236577. [Google Scholar] [CrossRef]
- Tan, X.; Long, W.; Zeng, L.; Ding, X.; Cheng, Y.; Zhang, X.; Zou, X. Melatonin-induced transcriptome variation of rapeseed seedlings under salt stress. Int. J. Mol. Sci. 2019, 20, 5355. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Walk, T.C.; Han, P.; Chen, L.; Shen, X.; Li, Y.; Gu, C.; Xie, L.; Hu, X.; Liao, X.; et al. Genome-wide identification and analysis of high-affinity nitrate transporter 2 (NRT2) family genes in rapeseed (Brassica napus L.) and their responses to various stresses. BMC Plant Biol. 2020, 20, 464. [Google Scholar] [CrossRef]
- Wang, Z.; Han, Y.; Luo, S.; Rong, X.; Song, H.; Jiang, N.; Li, C.; Yang, L. Calcium peroxide alleviates the waterlogging stress of rapeseed by improving root growth status in a rice-rape rotation field. Front. Plant Sci. 2022, 13, 1048227. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhao, J.; Favero, D.S.; Qiu, J.; Roalson, E.H.; Neff, M.M. Insights into the evolution and diversification of the AT-hook Motif Nuclear Localized gene family in land plants. BMC Plant Biol. 2014, 14, 266. [Google Scholar] [CrossRef] [Green Version]
- Maire, P.; Wuarin, J.; Schibler, U. The role of cis-acting promoter elements in tissue-specific albumin gene expression. Science 1989, 244, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.N.; Chiang-Hsieh, Y.F.; Chien, C.H.; Zheng, H.Q.; Lee, T.Y.; Wu, N.Y.; Tseng, K.C.; Hou, P.F.; Chang, W.C. Delineation of condition specific Cis- and Trans-acting elements in plant promoters under various Endo- and exogenous stimuli. BMC Genom. 2018, 19, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, C.; Wang, X.; Yu, J.; Wu, J.; Li, W.; Huang, J.; Dong, C.; Hua, W.; Liu, S. Comprehensive analysis of RNA-seq data reveals the complexity of the transcriptome in Brassica rapa. BMC Genom. 2013, 14, 689. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S.; et al. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Yu, L.; Wei, L.; Yu, P.; Wang, J.; Zhao, H.; Zhang, Y.; Zhang, S.; Yang, Z.; Chen, G.; et al. BnTIR: An online transcriptome platform for exploring RNA-seq libraries for oil crop Brassica napus. Plant Biotechnol. J. 2021, 19, 1895–1897. [Google Scholar] [CrossRef]
- Mendiondo, G.M.; Gibbs, D.J.; Szurman-Zubrzycka, M.; Korn, A.; Marquez, J.; Szarejko, I.; Maluszynski, M.; King, J.; Axcell, B.; Smart, K.; et al. Enhanced waterlogging tolerance in barley by manipulation of expression of the N-end rule pathway E3 ligase PROTEOLYSIS6. Plant Biotechnol. J. 2016, 14, 40–50. [Google Scholar] [CrossRef]
- Klok, E.J.; Wilson, I.W.; Wilson, D.; Chapman, S.C.; Ewing, R.M.; Somerville, S.C.; Peacock, W.J.; Dolferus, R.; Dennis, E.S. Expression profile analysis of the low-oxygen response in Arabidopsis root cultures. Plant Cell 2002, 14, 2481–2494. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.; Chen, Z.J. Genomic and expression plasticity of polyploidy. Curr. Opin. Plant Biol. 2010, 13, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, R.; Wu, X.; Wang, J. Homoeolog expression bias and expression level dominance (ELD) in four tissues of natural allotetraploid Brassica napus. BMC Genom. 2020, 21, 330. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Wei, W.; Tao, J.J.; Lu, X.; Bian, X.H.; Hu, Y.; Cheng, T.; Yin, C.C.; Zhang, W.K.; Chen, S.Y.; et al. Nuclear factor Y subunit GmNFYA competes with GmHDA13 for interaction with GmFVE to positively regulate salt tolerance in soybean. Plant Biotechnol. J. 2021, 19, 2362–2379. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Dogra, V.; Joshi, R.; Kumar, S. Stress-responsive cis-regulatory elements underline podophyllotoxin biosynthesis and better performance of Sinopodophyllum hexandrum under water deficit conditions. Front. Plant Sci. 2021, 12, 751846. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.S.; Hussain, M.A.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-wide analysis, identification, and expression pattern in response to multiple hormones and abiotic stress conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef]
- Bian, X.H.; Li, W.; Niu, C.F.; Wei, W.; Hu, Y.; Han, J.Q.; Lu, X.; Tao, J.J.; Jin, M.; Qin, H.; et al. A class B heat shock factor selected for during soybean domestication contributes to salt tolerance by promoting flavonoid biosynthesis. New Phytol. 2020, 225, 268–283. [Google Scholar] [CrossRef] [Green Version]
- Bailey-Serres, J.; Fukao, T.; Gibbs, D.J.; Holdsworth, M.J.; Lee, S.C.; Licausi, F.; Perata, P.; Voesenek, L.A.; Dongen, J.T. Making sense of low oxygen sensing. Trends Plant Sci. 2012, 17, 129–138. [Google Scholar] [CrossRef]
- Ambros, S.; Kotewitsch, M.; Wittig, P.R.; Bammer, B.; Mustroph, A. Transcriptional response of two Brassica napus cultivars to short-term hypoxia in the root zone. Front. Plant Sci. 2022, 13, 897673. [Google Scholar] [CrossRef]
- Hong, B.; Zhou, B.; Peng, Z.; Yao, M.; Wu, J.; Wu, X.; Guan, C.; Guan, M. Tissue-specific transcriptome and metabolome analysis reveals the response mechanism of Brassica napus to waterlogging stress. Int. J. Mol. Sci. 2023, 24, 6015. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, S.; Wei, L.; Huang, Y.; Liu, D.; Jia, Y.; Luo, C.; Lin, Y.; Liang, C.; Hu, Y.; et al. BnIR: A multi-omics database with various tools for Brassica napus research and breeding. Mol. Plant 2023, 16, 775–789. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.T.; Lu, X.; Song, Q.X.; Chen, H.W.; Wei, W.; Tao, J.J.; Bian, X.H.; Shen, M.; Ma, B.; Zhang, W.K.; et al. Selection for a zinc-finger protein contributes to seed oil increase during soybean domestication. Plant Physiol. 2017, 173, 2208–2224. [Google Scholar] [CrossRef] [Green Version]
- Xi, X.; Wei, K.; Gao, B.; Liu, J.; Liang, J.; Cheng, F.; Wang, X.; Wu, J. BrFLC5: A weak regulator of flowering time in Brassica rapa. Theor. Appl. Genet. 2018, 131, 2107–2116. [Google Scholar] [CrossRef]
- Ridge, S.; Brown, P.H.; Hecht, V.; Driessen, R.G.; Weller, J.L. The role of BoFLC2 in cauliflower (Brassica oleracea var. botrytis L.) reproductive development. J. Exp. Bot. 2015, 66, 125–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Chen, X.; Zhang, S.; Zhu, J.; Tang, B.; Wang, A.; Dong, L.; Zhang, Z.; Yu, C.; Sun, Y.; et al. The genome sequence archive family: Toward explosive data growth and diverse data types. Genom. Proteom. Bioinform. 2021, 19, 578–583. [Google Scholar] [CrossRef]
- CNCB-NGDC Members and Partners. Database resources of the National Genomics Data Center, China National Center for Bioinformation in 2022. Nucleic Acids Res. 2022, 50, 27–38. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Nucleotide Length (bp) | Amino Acid | Molecular Weight (KD) | PI | Genome Location | Number of Introns | Number of Exons | |
|---|---|---|---|---|---|---|---|---|
| PCO1 | BnaA03G0061400ZS | 897 | 299 | 33.22 | 7.5 | ChrA03: 2,917,715–2,919,518 | 4 | 5 |
| BnaA10G0211400ZS | 924 | 308 | 34.18 | 8.01 | ChrA10: 22,378,464–22,380,486 | 4 | 5 | |
| BnaC03G0070600ZS | 879 | 293 | 32.75 | 7.77 | ChrC03: 3,619,461–3,621,089 | 4 | 5 | |
| BnaC09G0512300ZS | 771 | 257 | 28.66 | 5.91 | ChrC09: 61,478,635–61,483,755 | 4 | 5 | |
| Bo3G010080 | 876 | 292 | 32.62 | 7.52 | 3,923,348–3,924,992 | 4 | 5 | |
| Bo9G165400 | 918 | 306 | 33.96 | 8.78 | 60,734,007–60,728,419 | 4 | 5 | |
| BraA03G006660 | 897 | 299 | 33.3 | 7.5 | 2,874,949–2,876,803 | 4 | 5 | |
| BraA10G024330 | 717 | 239 | 26.63 | 5.14 | 16,636,583–16,634,714 | 4 | 5 | |
| PCO2 | BnaA04G0111100ZS | 846 | 282 | 31.35 | 8.21 | ChrA04: 12,952,829–12,954,006 | 4 | 5 |
| BnaC04G0395100ZS | 858 | 286 | 31.75 | 8.21 | ChrC04: 52,079,647–52,080,830 | 4 | 5 | |
| Bo4G139190 | 858 | 286 | 31.75 | 8.21 | 46,652,407–46,653,590 | 4 | 5 | |
| BraA04G013090 | 846 | 282 | 31.34 | 8.02 | 9,741,294–9,742,752 | 4 | 5 | |
| PCO3 | BnaA06G0127100ZS | 831 | 277 | 30.61 | 5 | ChrA06: 7,448,257–7,449,546 | 4 | 5 |
| BnaC04G0074400ZS | 246 | 82 | 8.65 | 4.39 | ChrC04: 6,512,883–6,514,191 | 2 | 3 | |
| BnaC05G0155300ZS | 831 | 277 | 30.62 | 5.01 | ChrC05: 9,956,060–9,957,351 | 4 | 5 | |
| BnaC05G0283800ZS | 318 | 106 | 11.58 | 4.17 | ChrC05: 24,420,297–24,421,163 | 1 | 2 | |
| BnaC05G0471900ZS | 387 | 129 | 14.3 | 8.6 | ChrC05: 52,043,598–52,044,332 | 1 | 2 | |
| BnaC09G0521600ZS | 387 | 129 | 14.23 | 8.37 | ChrC09: 62,173,896–62,174,630 | 1 | 2 | |
| Bo5G025500 | 834 | 278 | 30.67 | 5.01 | 9,490,230–9,491,521 | 4 | 5 | |
| BraA06G014250 | 831 | 277 | 30.55 | 4.89 | 7,467,637–7,469,108 | 4 | 5 | |
| PCO4 | BnaA03G0207500ZS | 624 | 208 | 23.51 | 6.5 | ChrA03: 10,842,878–10,844,151 | 3 | 4 |
| BnaA05G0034400ZS | 729 | 243 | 27.11 | 6.03 | ChrA05: 1,925,460–1,927,173 | 4 | 5 | |
| BnaC03G0244400ZS | 636 | 212 | 23.54 | 8.04 | ChrC03: 14,957,859–14,959,210 | 4 | 5 | |
| BnaC04G0037200ZS | 717 | 239 | 26.81 | 6.42 | ChrC04: 3,448,037–3,452,967 | 4 | 5 | |
| Bo3G036180 | 636 | 212 | 23.53 | 8.04 | 15,839,677–15,841,028 | 4 | 5 | |
| Bo4G017390 | 717 | 239 | 26.75 | 6.23 | 3,496,211–3,497,884 | 4 | 5 | |
| BraA05G003220 | 717 | 239 | 26.72 | 6.42 | 1,719,860–1,722,377 | 4 | 5 | |
| PCO5 | BnaA04G0023400ZS | 729 | 243 | 27.23 | 6.84 | ChrA04: 1,525,382–1,527,222 | 4 | 5 |
| BnaA09G0537000ZS | 729 | 243 | 27.13 | 6.59 | ChrA09: 56,193,470–56,194,841 | 4 | 5 | |
| BnaC04G0289700ZS | 729 | 243 | 27.22 | 6.78 | ChrC04: 39,729,996–39,732,404 | 4 | 5 | |
| BnaC08G0384200ZS | 729 | 243 | 27.2 | 6.5 | ChrC08: 44,790,558–44,792,029 | 4 | 5 | |
| Bo4G108280 | 729 | 243 | 27.22 | 6.78 | 34,730,028–34,731,729 | 4 | 5 | |
| Bo8G092800 | 729 | 243 | 27.2 | 6.5 | 41,444,747–41,446,184 | 4 | 5 | |
| BraA04G002890 | 729 | 243 | 27.22 | 6.99 | 1,612,409–1,614,775 | 4 | 5 | |
| BraA09G050770 | 729 | 243 | 27.12 | 6.59 | 36,671,235–36,668,985 | 4 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, X.; Cao, Y.; Zhi, X.; Ma, N. Genome-Wide Identification and Analysis of the Plant Cysteine Oxidase (PCO) Gene Family in Brassica napus and Its Role in Abiotic Stress Response. Int. J. Mol. Sci. 2023, 24, 11242. https://doi.org/10.3390/ijms241411242
Bian X, Cao Y, Zhi X, Ma N. Genome-Wide Identification and Analysis of the Plant Cysteine Oxidase (PCO) Gene Family in Brassica napus and Its Role in Abiotic Stress Response. International Journal of Molecular Sciences. 2023; 24(14):11242. https://doi.org/10.3390/ijms241411242
Chicago/Turabian StyleBian, Xiaohua, Yifan Cao, Ximin Zhi, and Ni Ma. 2023. "Genome-Wide Identification and Analysis of the Plant Cysteine Oxidase (PCO) Gene Family in Brassica napus and Its Role in Abiotic Stress Response" International Journal of Molecular Sciences 24, no. 14: 11242. https://doi.org/10.3390/ijms241411242
APA StyleBian, X., Cao, Y., Zhi, X., & Ma, N. (2023). Genome-Wide Identification and Analysis of the Plant Cysteine Oxidase (PCO) Gene Family in Brassica napus and Its Role in Abiotic Stress Response. International Journal of Molecular Sciences, 24(14), 11242. https://doi.org/10.3390/ijms241411242