

Vitamin D Promotes Mucosal Barrier System of Fish Skin Infected with Aeromonas hydrophila through Multiple Modulation of Physical and Immune Protective Capacity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
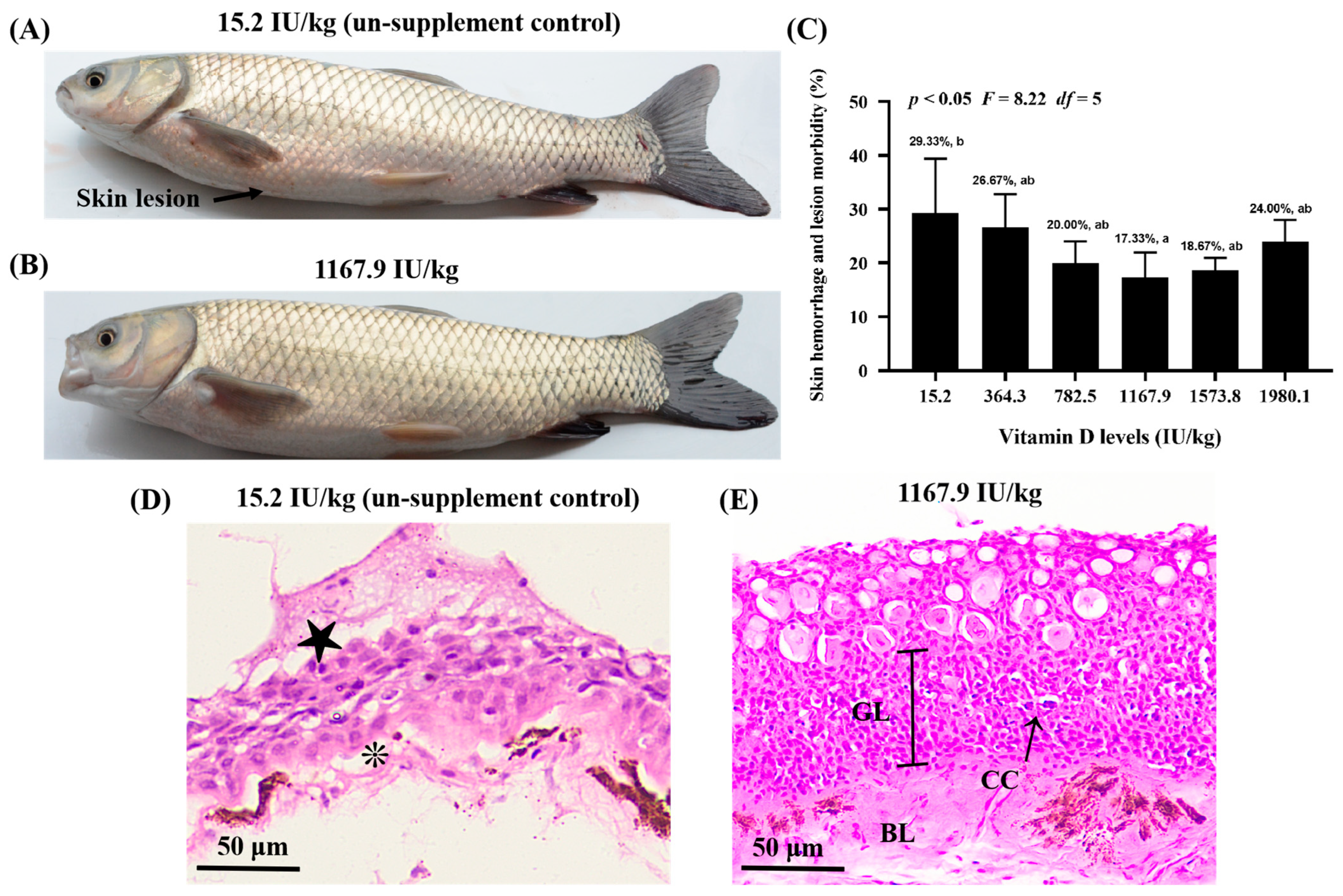
2.1. Disease-Resistant Phenotypes
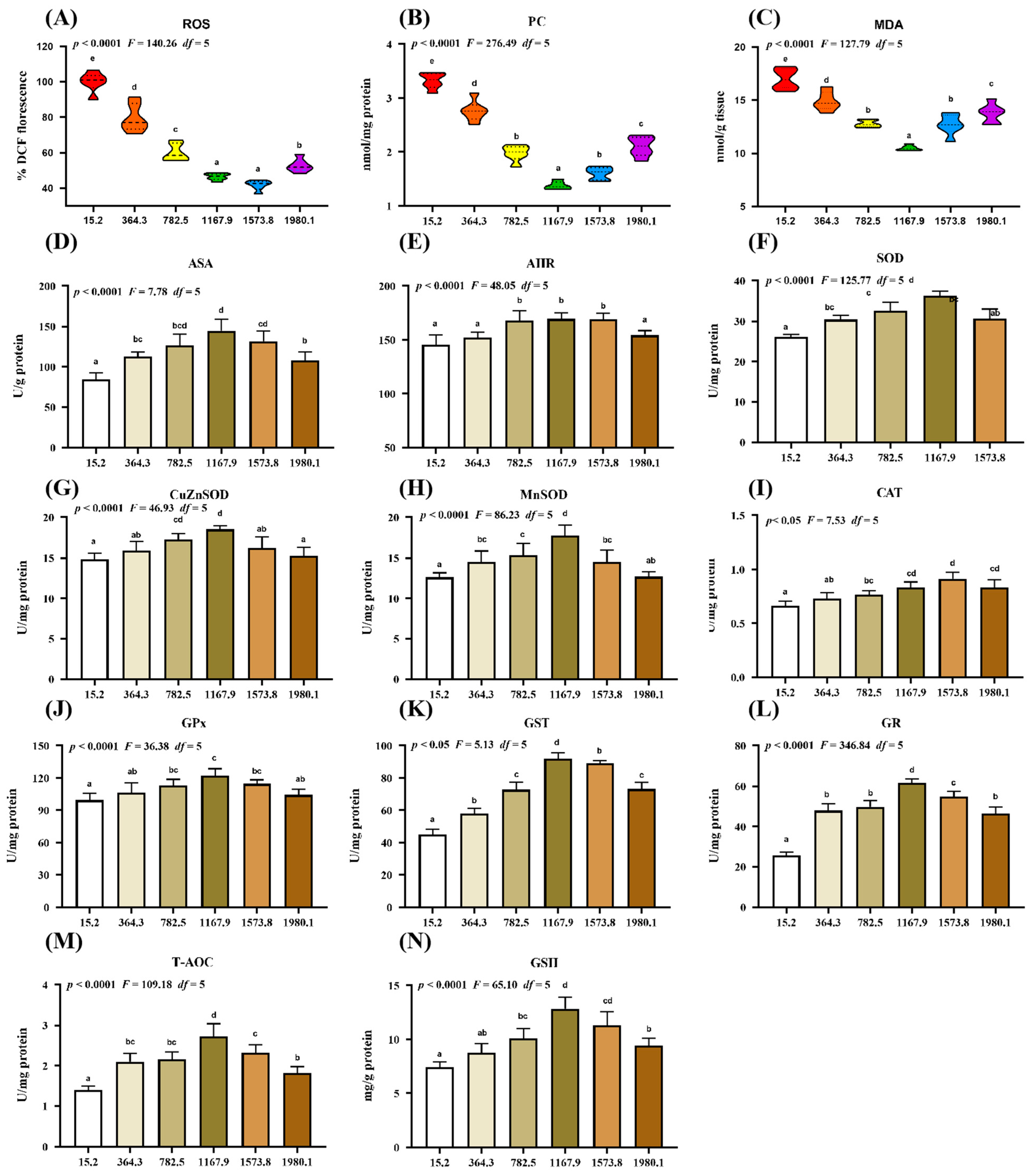
2.2. Oxidative Damage Biomarkers and Antioxidant Enzymes
2.3. Key Regulatory Genes Involved in the Physical Barrier Function of Skin
2.3.1. Key Regulatory Genes for Antioxidant-Related Parameters
2.3.2. Key Regulatory Gene for Apoptosis-Related Parameters
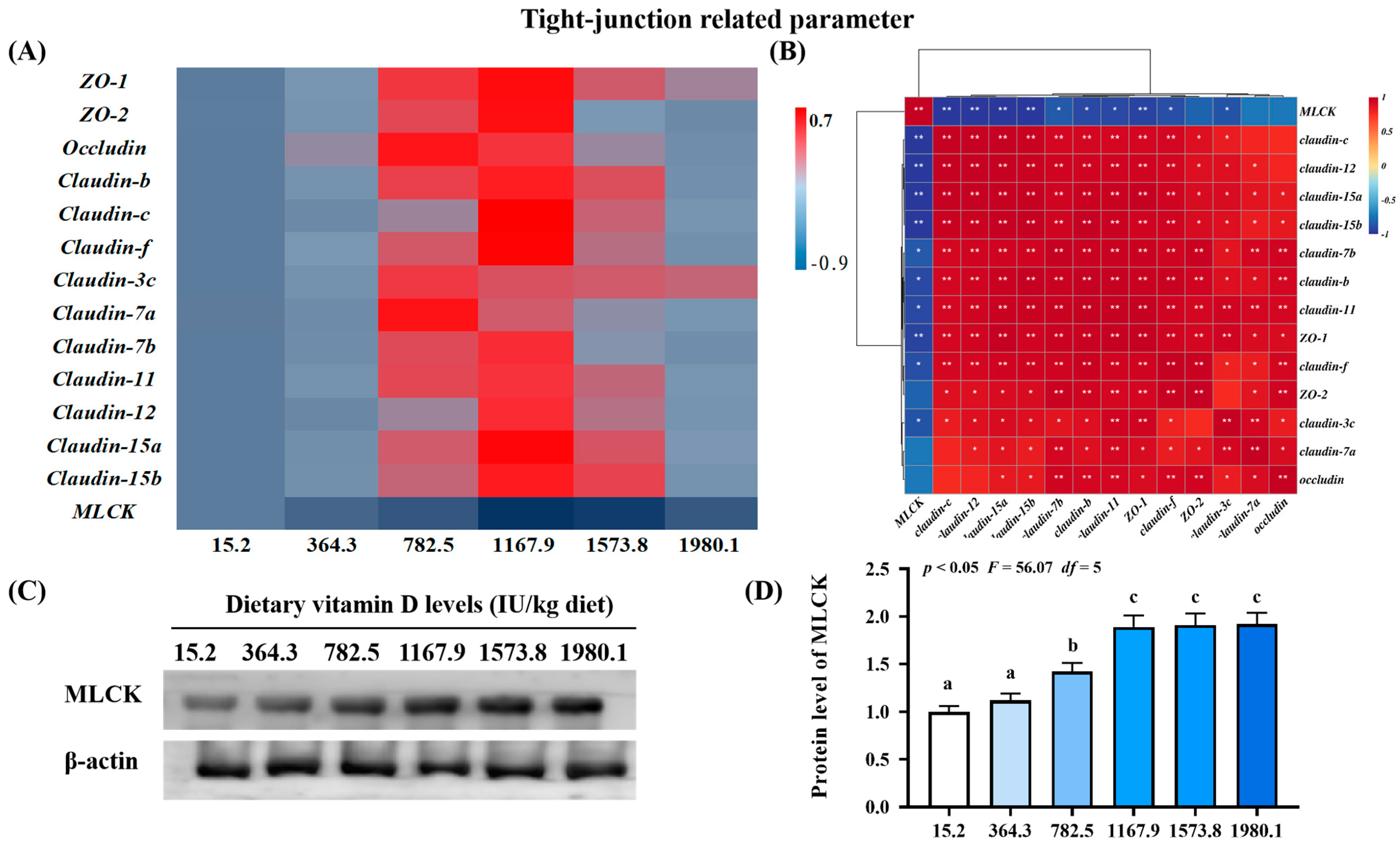
2.3.3. Key Regulatory Gene for Tight Junction-Related Parameters
2.4. Immune-Related Parameters and Key Regulatory Genes of Skin Immune Barrier Function
2.5. Expression of VDR
3. Discussion
3.1. Dietary Vitamin D Supplementation Enhanced Disease Resistance in Fish Skin
3.2. Dietary Vitamin D Supplementation Improved Physical Barrier Function in the Skin
3.2.1. Dietary Vitamin D Supplementation Enhanced Antioxidant Capacity in the Skin
3.2.2. Vitamin D Supplementation Decreased Cell Apoptosis, Mediated by the p38MAPK and JNK Pathways
3.2.3. Vitamin D Supplementation Enhanced Tight Junction Function via MLCK Inhibition
3.3. Vitamin D Supplementation Improved Immune Barrier Function in Fish Skin
3.4. Vitamin D Supplementation Improved VDR Expression in Fish Skin
4. Conclusions
5. Materials and Methods
5.1. Experimental Diets and Conditions
5.2. Challenge Trial and Sampling
5.3. Biochemical Analysis
5.4. Hematoxylin and Eosin Staining of Fish Skin
5.5. Total RNA Extraction and Real-Time qPCR
5.6. Western Blot Analysis
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Assefa, A.; Abunna, F. Maintenance of fish health in aquaculture: Review of epidemiological approaches for prevention and control of infectious disease of fish. Vet.-Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Qin, L.; Zhao, D.; Xiong, F.; Wang, G.; Zou, H.; Li, W.; Li, M.; Song, K.; Wu, S. Growth performance, immunity and intestinal microbiota of swamp eel (Monopterus albus) fed a diet supplemented with house fly larvae (Musca domestica). Aquac. Nutr. 2020, 26, 693–704. [Google Scholar] [CrossRef]
- Esteban, M.Á.; Cerezuela, R. Fish mucosal immunity: Skin. In Mucosal Health in Aquaculture; Academic Press: Cambridge, MA, USA, 2015; pp. 34–92. [Google Scholar] [CrossRef]
- Jiang, W.-D.; Zhang, L.; Feng, L.; Wu, P.; Liu, Y.; Kuang, S.-Y.; Li, S.-W.; Tang, L.; Mi, H.-F.; Zhang, L.; et al. New insight on the immune modulation and physical barrier protection caused by vitamin A in fish gills infected with Flavobacterium columnare. Front. Immunol. 2022, 13, 833455. [Google Scholar] [CrossRef] [PubMed]
- Graff, I.; Høie, S.; Totland, G.; Lie, Ø. Three different levels of dietary vitamin D3 fed to first-feeding fry of Atlantic salmon (Salmo salar L.): Effect on growth, mortality, calcium content and bone formation. Aquac. Nutr. 2002, 8, 103–111. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, D.; Yongyut, P.; Li, G.; Esteban, M.Á.; Jintasataporn, O.; Deng, J.; Zhang, W.; Ai, Q.; Mai, K.; et al. Vitamin D3 deficiency induced intestinal inflammatory response of turbot through nuclear factor-κB/inflammasome pathway, accompanied by the mutually exclusive apoptosis and autophagy. Front. Immunol. 2022, 13, 986593. [Google Scholar] [CrossRef]
- He, S.; Ding, M.; Ray, G.W.; Yang, Q.; Tan, B.; Dong, X.; Chi, S.; Liu, H.; Zhang, S. Effect of dietary vitamin D levels on growth, serum biochemical parameters, lipid metabolism enzyme activities, fatty acid synthase and hepatic lipase mRNA expression for orange-spotted grouper (Epinephelus coioides) in growth mid-stage. Aquac. Nutr. 2021, 27, 655–665. [Google Scholar] [CrossRef]
- Li, X.; Chen, F.; Huang, D.; Guo, Y.; Wu, Y.; Wu, C.; Zhang, W.; Mai, K. Interactions of dietary carbohydrate and vitamin D3 on growth performance, insulin signaling pathway and glucose metabolism in juvenile abalone Haliotis discus hannai. Aquaculture 2021, 542, 736908. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.-N.; Jiang, W.-D.; Wu, P.; Liu, Y.; Kuang, S.-Y.; Tang, L.; Li, S.-W.; Jin, X.-W.; Ren, H.-M.; et al. Vitamin D serves as a modulator of immune organs in grass carp (Ctenopharyngodon idella) infected with Aeromonas hydrophila. Aquaculture 2023, 565, 739144. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.-N.; Jiang, W.-D.; Wu, P.; Liu, Y.; Kuang, S.-Y.; Tang, L.; Li, S.-W.; Jin, X.-W.; Ren, H.-M.; et al. An emerging role of vitamin D3 in amino acid absorption in different intestinal segments of on-growing grass carp (Ctenopharyngodon idella). Anim. Nutr. 2022, 10, 305–318. [Google Scholar] [CrossRef]
- Yuan, Z.; Wu, P.; Feng, L.; Jiang, W.; Liu, Y.; Kuang, S.; Tang, L.; Zhou, X. Dietary choline inhibited the gill apoptosis in association with the p38MAPK and JAK/STAT3 signalling pathways of juvenile grass carp (Ctenopharyngodon idella). Aquaculture 2020, 529, 735699. [Google Scholar] [CrossRef]
- Dash, S.; Das, S.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar] [CrossRef]
- Liu, Y.; Meng, F.; Wang, S.; Xia, S.; Wang, R. Vitamin D3 mitigates lipopolysaccharide-induced oxidative stress, tight junction damage and intestinal inflammatory response in yellow catfish, Pelteobagrus fulvidraco. Comp. Biochem. Physiol. Part C 2021, 243, 108982. [Google Scholar] [CrossRef]
- Liu, S.; Wang, X.; Bu, X.; Lin, Z.; Li, E.; Shi, Q.; Zhang, M.; Qin, J.G.; Chen, L. Impact of Dietary Vitamin D3 Supplementation on Growth, Molting, Antioxidant Capability, and Immunity of Juvenile Chinese Mitten Crabs (Eriocheir sinensis) by Metabolites and Vitamin D Receptor. J. Agric. Food Chem. 2021, 69, 12794–12806. [Google Scholar] [CrossRef] [PubMed]
- Ki, Y.-W.; Park, J.H.; Lee, J.E.; Shin, I.C.; Koh, H.C. JNK and p38 MAPK regulate oxidative stress and the inflammatory response in chlorpyrifos-induced apoptosis. Toxicol. Lett. 2013, 218, 235–245. [Google Scholar] [CrossRef]
- Niessen, C.M. Tight junctions/adherens junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2525–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Black, E.D.; Witkowski, E.D.; Lencer, W.I.; Guerriero, V.; Schneeberger, E.E.; Turner, J.R. Myosin light chain phosphorylation regulates barrier function by remodeling tight junction structure. J. Cell Sci. 2006, 119, 2095–2106. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.-D.; Zheng, X.; Feng, L.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X. The antioxidant status, apoptosis, intercellular integrity and immune function of grass carp (Ctenopharyngodon idella) head kidney and spleen fed different levels of pyridoxine. Aquac. Nutr. 2020, 26, 613–630. [Google Scholar] [CrossRef]
- Bishop, E.L.; Ismailova, A.; Dimeloe, S.K.; Hewison, M.; White, J.H. Vitamin D and immune regulation: Antibacterial, antiviral, anti-Inflammatory. JBMR Plus 2021, 5, e10405. [Google Scholar] [CrossRef] [PubMed]
- Roussel, P.; Delmotte, D. The diversity of epithelial secreted mucins. Curr. Org. Chem. 2004, 8, 413–437. [Google Scholar] [CrossRef]
- Raj, V.S.; Fournier, G.; Rakus, K.; Ronsmans, M.; Ouyang, P.; Michel, B.; Delforges, C.; Costes, B.; Farnir, F.; Leroy, B.; et al. Skin mucus of Cyprinus carpio inhibits cyprinid herpesvirus 3 binding to epidermal cells. Vet. Res. 2011, 42, 92. [Google Scholar] [CrossRef] [Green Version]
- Christakos, S.; Dhawan, P.; Verstuyf, A.; Verlinden, L.; Carmeliet, G. Vitamin D: Metabolism, Molecular Mechanism of Action, and Pleiotropic Effects. Physiol. Rev. 2016, 96, 365–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Zhang, Y.-G.; Xia, Y.; Sun, J. Imbalance of autophagy and apoptosis in intestinal epithelium lacking the vitamin D receptor. FASEB J. 2019, 33, 11845–11856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Lu, R.; Zhang, Y.-G.; Sun, J. Vitamin D Receptor Deletion Leads to the Destruction of Tight and Adherens Junctions in Lungs. Tissue Barriers 2018, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, V.; White, J.H. Species-specific regulation of innate immunity by vitamin D signaling. J. Steroid Biochem. Mol. Biol. 2016, 164, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q.; et al. The draft genome of the grass carp (Ctenopharyngodon idellus) provides insights into its evolution and vegetarian adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of World Fisheries and Aquaculture 2020: Sustainability in Action; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2020; pp. 9–65. [Google Scholar]
- MAF. China Fishery Statistical Yearbook; Chinese Ministry of Agriculture, China Agriculture Press: Beijing, China, 2021; p. 25. (In Chinese) [Google Scholar]
- Kong, W.-G.; Li, S.-S.; Chen, X.-X.; Huang, Y.-Q.; Tang, Y.; Wu, Z.-X. A study of the damage of the intestinal mucosa barrier structure and function of Ctenopharyngodon idella with Aeromonas hydrophila. Fish Physiol. Biochem. 2017, 43, 1223–1235. [Google Scholar] [CrossRef]
- Song, X.; Zhao, J.; Bo, Y.; Liu, Z.; Wu, K.; Gong, C. Aeromonas hydrophila induces intestinal inflammation in grass carp (Ctenopharyngodon idella): An experimental model. Aquaculture 2014, 434, 171–178. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, L.; Jiang, W.; Wu, P.; Liu, Y.; Kuang, S.; Tang, L.; Zhou, X. Lysine deficiency impaired growth performance and immune response and aggravated inflammatory response of the skin, spleen and head kidney in grown-up grass carp (Ctenopharyngodon idella). Anim. Nutr. 2021, 7, 556–568. [Google Scholar] [CrossRef]
- Lu, Z.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Li, S.-W.; Liu, X.-A.; et al. Mannan oligosaccharides application: Multipath restriction from Aeromonas hydrophila infection in the skin barrier of grass Carp (Ctenopharyngodon idella). Front. Immunol. 2021, 12, 742107. [Google Scholar] [CrossRef]
- Ko, J.-Y.; Kim, E.-A.; Lee, J.-H.; Kang, M.-C.; Lee, J.-S.; Kim, J.-S.; Jung, W.-K.; Jeon, Y.-J. Protective effect of aquacultured flounder fish-derived peptide against oxidative stress in zebrafish. Fish Shellfish Immunol. 2014, 36, 320–323. [Google Scholar] [CrossRef]
- Ahmad, R.; Tripathi, A.; Tripathi, P.; Singh, S.; Singh, R.; Singh, R.K. Malondialdehyde and protein carbonyl as biomarkers for oxidative stress and disease progression in patients with chronic myeloid leukemia. Vivo 2008, 22, 525–528. [Google Scholar]
- Chen, J.; Zhou, X.Q.; Feng, L.; Liu, Y.; Jiang, J. Effects of glutamine on hydrogen peroxide-induced oxidative damage in intestinal epithelial cells of Jian carp (Cyprinus carpio var. Jian). Aquaculture 2009, 288, 285–289. [Google Scholar] [CrossRef]
- Wen, H.; Feng, L.; Jiang, W.; Liu, Y.; Jiang, J.; Li, S.; Tang, L.; Zhang, Y.; Kuang, S.; Zhou, X. Dietary tryptophan modulates intestinal immune response, barrier function, antioxidant status and gene expression of TOR and Nrf2 in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2014, 40, 275–287. [Google Scholar] [CrossRef]
- Giuliani, M.E.; Regoli, F. Identification of the Nrf2–Keap1 pathway in the European eel Anguilla anguilla: Role for a transcriptional regulation of antioxidant genes in aquatic organisms. Aquat. Toxicol. 2014, 150, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-P.; Jiang, W.-D.; Liu, Y.; Jiang, J.; Wu, P.; Zhao, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; et al. Exogenous phospholipids supplementation improves growth and modulates immune response and physical barrier referring to NF-κB, TOR, MLCK and Nrf2 signaling factors in the intestine of juvenile grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2015, 47, 46–62. [Google Scholar] [CrossRef]
- Martínez-Sena, T.; Soluyanova, P.; Guzmán, C.; Valdivielso, J.M.; Castell, J.V.; Jover, R. The Vitamin D Receptor Regulates Glycerolipid and Phospholipid Metabolism in Human Hepatocytes. Biomolecules 2020, 10, 493. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.; Shaha, C. Role of glutathione S-transferases in oxidative stress–induced male germ cell apoptosis. Free Radic. Biol. Med. 2000, 29, 1015–1027. [Google Scholar] [CrossRef]
- Kobayashi, N.; DeLano, F.A.; Schmid-Schönbein, G.W. Oxidative Stress Promotes Endothelial Cell Apoptosis and Loss of Microvessels in the Spontaneously Hypertensive Rats. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2114–2121. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S. Molecular signaling in death receptor and mitochondrial pathways of apoptosis (Review). Int. J. Oncol. 2003, 22, 15–20. [Google Scholar] [CrossRef]
- Cai, G.; Si, M.; Li, X.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis of rat Sertoli cells through Fas-Fas ligand and mitochondrial pathway. Environ. Toxicol. 2019, 34, 424–433. [Google Scholar] [CrossRef]
- Tanel, A.; Averill-Bates, D.A. Activation of the death receptor pathway of apoptosis by the aldehyde acrolein. Free Radic. Biol. Med. 2007, 42, 798–810. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Liu, W.-H.; Kao, P.-H.; Wang, J.-J.; Chang, L.-S. Involvement of p38 MAPK- and JNK-modulated expression of Bcl-2 and Bax in Naja nigricollis CMS-9-induced apoptosis of human leukemia K562 cells. Toxicon 2010, 55, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.; Ma, C.; Guo, X.; Huang, Y.; Tang, R.; Karrow, N.A.; Wang, C. Vitamin D3 modulates yellow catfish (Pelteobagrus fulvidraco) immune function in vivo and in vitro and this involves the vitamin D3/VDR-type I interferon axis. Dev. Comp. Immunol. 2020, 107, 103644. [Google Scholar] [CrossRef]
- González-Mariscal, L.; Betanzos, A.; Nava, P.; Jaramillo, B. Tight junction proteins. Prog. Biophys. Mol. Biol. 2003, 81, 1–44. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-J.; Jiang, W.-D.; Feng, L.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; et al. Dietary vitamin C deficiency depresses the growth, head kidney and spleen immunity and structural integrity by regulating NF-κB, TOR, Nrf2, apoptosis and MLCK signaling in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2016, 52, 111–138. [Google Scholar] [CrossRef]
- Koenderman, L.; Buurman, W.; Daha, M.R. The innate immune response. Immunol. Lett. 2014, 162, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Kato, G.; Takano, T.; Sakai, T.; Matsuyama, T.; Nakayasu, C. Vibrio anguillarum bacterin uptake via the gills of Japanese flounder and subsequent immune responses. Fish Shellfish Immunol. 2013, 35, 1591–1597. [Google Scholar] [CrossRef] [PubMed]
- Tatiya-Aphiradee, N.; Chatuphonprasert, W.; Jarukamjorn, K. Ethanolic Garcinia mangostana extract and α-mangostin improve dextran sulfate sodium-induced ulcerative colitis via the suppression of inflammatory and oxidative responses in ICR mice. J. Ethnopharmacol. 2021, 265, 113384. [Google Scholar] [CrossRef]
- Fu, C.-L.; Ye, Y.-L.; Lee, Y.-L.; Chiang, B.-L. Effects of overexpression of IL-10, IL-12, TGF-beta and IL-4 on allergen induced change in bronchial responsiveness. Respir. Res. 2006, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Shao, R.; Liu, J.; Lan, Y.; Liao, X.; Zhang, J.; Xu, W.; Mai, K.; Ai, Q.; Wan, M. Vitamin D impacts on the intestinal health, immune status and metabolism in turbot (Scophthalmus maximus L.). Br. J. Nutr. 2022, 128, 2083–2096. [Google Scholar] [CrossRef]
- Liu, J.; Shao, R.; Lan, Y.; Liao, X.; Zhang, J.; Mai, K.; Ai, Q.; Wan, M. Vitamin D3 protects turbot (Scophthalmus maximus L.) from bacterial infection. Fish Shellfish Immunol. 2021, 118, 25–33. [Google Scholar] [CrossRef]
- Wang, T.; Husain, M. The expanding repertoire of the IL-12 cytokine family in teleost fish: Identification of three paralogues each of the p35 and p40 genes in salmonids, and comparative analysis of their expression and modulation in Atlantic salmon Salmo salar. Dev. Comp. Immunol. 2014, 46, 194–207. [Google Scholar] [CrossRef]
- Su, Y.-N.; Wu, P.; Feng, L.; Jiang, W.-D.; Jiang, J.; Zhang, Y.-A.; Figueiredo-Silva, C.; Zhou, X.-Q.; Liu, Y. The improved growth performance and enhanced immune function by DL-methionyl-DL-methionine are associated with NF-kappa B and TOR signalling in intestine of juvenile grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2018, 74, 101–118. [Google Scholar] [CrossRef]
- Bollrath, J.; Greten, F.R. IKK/NF-κB and STAT3 pathways: Central signalling hubs in inflammation-mediated tumour promotion and metastasis. EMBO Rep. 2009, 10, 1314–1319. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; Tang, Q.; Guo, X.; Karow, N.A.; Wang, C. High dose of dietary vitamin D3 modulated the yellow catfish (Pelteobagrus fulvidraco) splenic innate immune response after Edwardsiella ictaluri infection. Fish Shellfish Immunol. 2020, 100, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Mazor, K.M.; Stipanuk, M.H. GCN2- and eIF2α-phosphorylation-independent, but ATF4-dependent, induction of CARE-containing genes in methionine-deficient cells. Amino Acids 2016, 48, 2831–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, M.J.; Jafri, M.; Wehmeier, K.R.; Onstead-Haas, L.M.; Mooradian, A.D. Inhibition of endoplasmic reticulum stress and oxidative stress by vitamin D in endothelial cells. Free Radic. Biol. Med. 2016, 99, 1–10. [Google Scholar] [CrossRef]
- Kollitz, E.M.; Hawkins, M.B.; Whitfield, G.K.; Kullman, S.W. Functional diversification of vitamin D receptor paralogs in teleost fish after a whole genome duplication event. Endocrinology 2014, 155, 4641–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollitz, E.M.; Zhang, G.; Hawkins, M.B.; Whitfield, G.K.; Reif, D.M.; Kullman, S.W. Molecular cloning, functional characterization, and evolutionary analysis of vitamin D receptors isolated from basal vertebrates. PLoS ONE 2015, 10, e0122853. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-H.; Su, C.-H.; Tseng, D.-Y.; Ding, F.-C.; Hwang, P.-P. Action of vitamin D and the receptor, VDRa, in calcium handling in zebrafish (Danio rerio). PLoS ONE 2012, 7, e45650. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.-J. Vitamin D receptor signaling regulates craniofacial cartilage development in zebrafish. J. Dev. Biol. 2019, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Shang, G.; Wang, W.; Chen, X.; Lou, Q.; Zhai, G.; Li, D.; Du, Z.; Ye, Y.; Jin, X.; et al. Fatty acid oxidation in zebrafish adipose tissue is promoted by 1α,25(OH)2D3. Cell Rep. 2017, 19, 1444–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Wu, F.; Wen, H.; Leng, X.J.; Liu, W. Effects of dietary vitamin D3 on growth and body composition of juvenile grass carp (Ctenopharyngodon idella). Freshw. Fish 2009, 39, 5. (In Chinese) [Google Scholar]
- Miao, L.H.; Ge, X.P.; Xie, J.; Liu, B.; Wang, K.B.; Zhu, J.; Ren, M.C.; Zhou, Q.L.; Pan, L.K.; Chen, R.L. Dietary vitamin D3 requirement of Wuchang bream (Megalobrama amblycephala). Aquaculture 2015, 436, 104–109. [Google Scholar] [CrossRef]
- Wang, L.S.; Xu, H.; Wang, Y.; Wang, C.A.; Li, J.N.; Zhao, Z.G.; Luo, L.; Du, X.; Xu, Q.Y. Effects of the supplementation of vitamin D 3 on the growth and vitamin D metabolites in juvenile Siberian sturgeon (Acipenser baerii). Fish Physiol. Biochem. 2017, 43, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Shiau, S.Y.; Hwang, J.Y. Vitamin D requirements of Juvenile Hybird tilapia (Oreochromis niloticus × O.aureus). Nippon Suisan Gakk. 1993, 59, 553–558. [Google Scholar] [CrossRef]
- Darias, M.J.; Mazurais, D.; Koumoundouros, G.; Glynatsi, N.; Christodoulopoulou, S.; Huelvan, C.; Desbruyeres, E.; Gall, M.M.L.; Quazuguel, P.; Cahu, C.L.; et al. Dietary vitamin D3 affects digestive system ontogenesis and ossification in European sea bass (Dicentrachus labrax, Linnaeus, 1758). Aquaculture 2010, 298, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Li, D.B.; Shao, S.S.; Zhang, G.W.; Zhou, D.G.; Li, X.W. Effects of dietary vitamin D3 on growth and body composition of Asian swamp eel (Monopterus albus). Anim. Nutr. 2015, 27, 1145–1151. (In Chinese) [Google Scholar]
- Liu, X.-W.; Feng, L.; Jiang, W.-D.; Wu, P.; Yang, D.-M.; Tang, L.; Kuang, S.-Y.; Shi, H.-Q.; Zhou, X.-Q.; Liu, Y. Novel insights into the intestinal immune regulatory effects of (2-Carboxyethyl) dimethylsulfonium Bromide (Br-DMPT) in on-growing grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2020, 98, 534–550. [Google Scholar] [CrossRef]
- Sun, H.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Yang, Q.-H.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q.; Feng, L. Betaine supplementations enhance the intestinal immunity of on-growing grass carp (Ctenopharyngodon idella): Partly related to TOR and NF-κB signaling pathways. Aquaculture 2020, 518, 734846. [Google Scholar] [CrossRef]
- Jiang, W.-D.; Deng, Y.-P.; Liu, Y.; Qu, B.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Wu, P.; Zhang, Y.-A.; et al. Dietary leucine regulates the intestinal immune status, immune-related signalling molecules and tight junction transcript abundance in grass carp (Ctenopharyngodon idella). Aquaculture 2015, 444, 134–142. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Tang, L.; Hu, K.; Liu, Y.; Jiang, W.-D.; Jiang, J.; Wu, P.; Chen, G.-F.; Li, S.-H.; Kuang, S.-Y.; et al. Effect of dietary lysine on growth, intestinal enzymes activities and antioxidant status of sub-adult grass carp (Ctenopharyngodon idella). Fish Physiol. Biochem. 2014, 40, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Damiran, D.; Preston, N.; McKinnon, J.; Jonker, A.; Christensen, D.; McAllister, T.A.; Yu, P. Effects of barley-based diets with 3 different rumen-degradable protein balances on performance and carcass characteristics of feedlot steers. Prof. Anim. Sci. 2014, 30, 432–443. [Google Scholar] [CrossRef]
- Refat, B.; Christensen, D.A.; McKinnon, J.J.; Yang, W.; Beattie, A.D.; McAllister, T.A.; Eun, J.-S.; Abdel-Rahman, G.A.; Yu, P. Effect of fibrolytic enzymes on lactational performance, feeding behavior, and digestibility in high-producing dairy cows fed a barley silage-based diet. J. Dairy Sci. 2018, 101, 7971–7979. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhou, X.-Q.; Jiang, W.-D.; Wu, P.; Liu, Y.; Ren, H.-M.; Jin, X.-W.; Feng, L. Vitamin D Promotes Mucosal Barrier System of Fish Skin Infected with Aeromonas hydrophila through Multiple Modulation of Physical and Immune Protective Capacity. Int. J. Mol. Sci. 2023, 24, 11243. https://doi.org/10.3390/ijms241411243
Zhang Y, Zhou X-Q, Jiang W-D, Wu P, Liu Y, Ren H-M, Jin X-W, Feng L. Vitamin D Promotes Mucosal Barrier System of Fish Skin Infected with Aeromonas hydrophila through Multiple Modulation of Physical and Immune Protective Capacity. International Journal of Molecular Sciences. 2023; 24(14):11243. https://doi.org/10.3390/ijms241411243
Chicago/Turabian StyleZhang, Yao, Xiao-Qiu Zhou, Wei-Dan Jiang, Pei Wu, Yang Liu, Hong-Mei Ren, Xiao-Wan Jin, and Lin Feng. 2023. "Vitamin D Promotes Mucosal Barrier System of Fish Skin Infected with Aeromonas hydrophila through Multiple Modulation of Physical and Immune Protective Capacity" International Journal of Molecular Sciences 24, no. 14: 11243. https://doi.org/10.3390/ijms241411243
APA StyleZhang, Y., Zhou, X. -Q., Jiang, W. -D., Wu, P., Liu, Y., Ren, H. -M., Jin, X. -W., & Feng, L. (2023). Vitamin D Promotes Mucosal Barrier System of Fish Skin Infected with Aeromonas hydrophila through Multiple Modulation of Physical and Immune Protective Capacity. International Journal of Molecular Sciences, 24(14), 11243. https://doi.org/10.3390/ijms241411243