
Drug-Inducible Gene Therapy Effectively Reduces Spontaneous Seizures in Kindled Rats but Creates Off-Target Side Effects in Inhibitory Neurons
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Design of a Drug-Inducible Gene Therapy
2.2. In Vitro Testing of AAV-SmonCeiT
2.3. In Vivo Testing of AAV-SmonCeiT
2.4. Evidence for “Leak” Expression in the Absence of Doxycycline
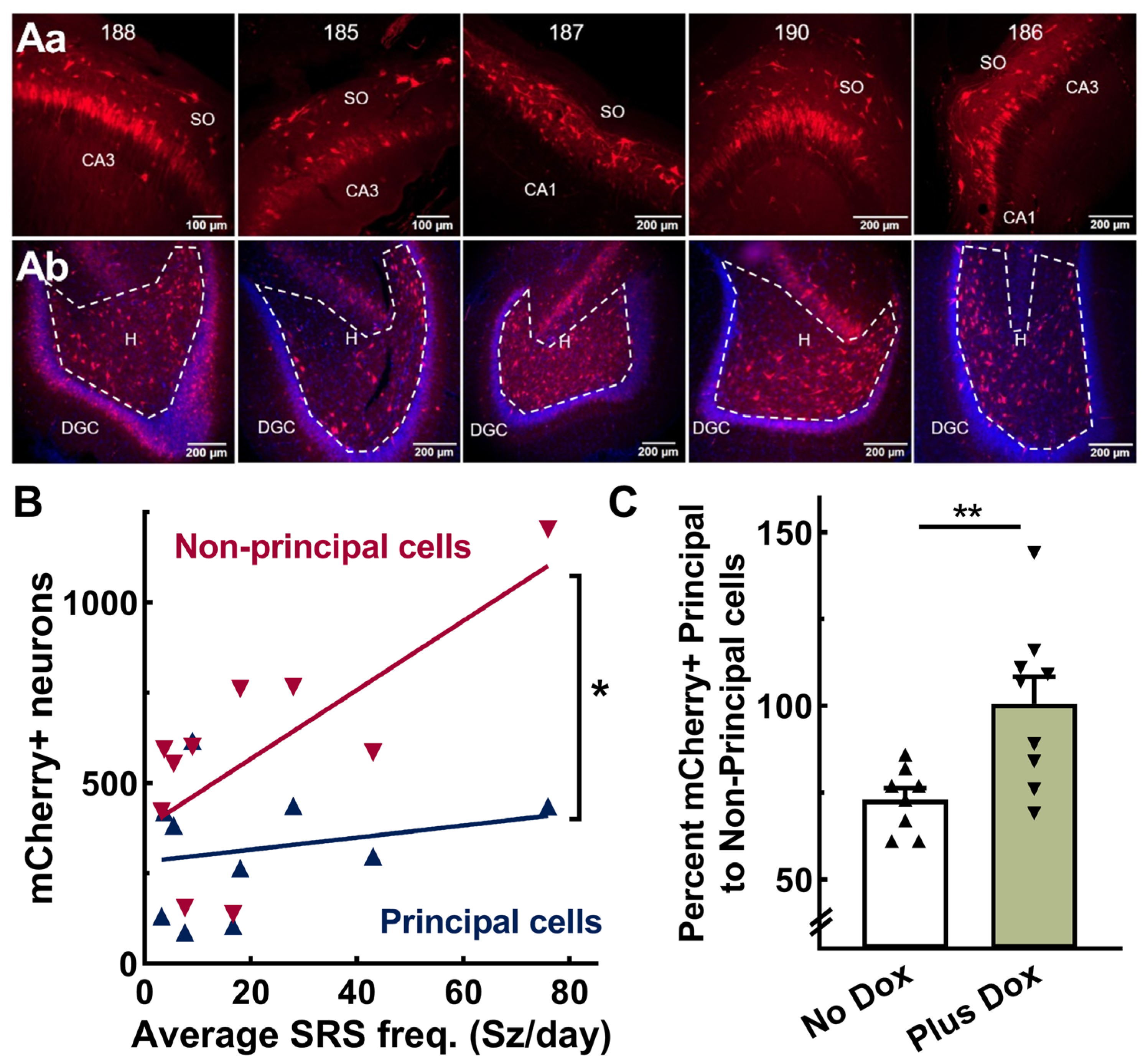
2.5. AAV-SmonCeiT Expression in the Hippocampus
3. Discussion
4. Materials and Methods
4.1. Molecular Cloning
4.2. Stereotactic Injection of AAV and Implantation of Electrodes
4.3. Kindling and Electroencephalogram (EEG) Recordings
4.4. Imaging Studies of AAV Distribution
4.5. DNA and RNA Isolation and Real Time Quantitative PCR (RT-qPCR) Protocols in Rats
4.6. Histology of Parvalbumin-Expressing Hippocampal Interneurons in Rats
4.7. Cre-Dependent AAV-SmonCeiT Expression in Mice
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asadi-Pooya, A.A.; Stewart, G.R.; Abrams, D.J.; Sharan, A. Prevalence and Incidence of Drug-Resistant Mesial Temporal Lobe Epilepsy in the United States. World Neurosurg. 2017, 99, 662–666. [Google Scholar] [CrossRef]
- Kwan, P.; Arzimanoglou, A.; Berg, A.T.; Brodie, M.J.; Allen Hauser, W.; Mathern, G.; Moshé, S.L.; Perucca, E.; Wiebe, S.; French, J. Definition of drug resistant epilepsy: Consensus proposal by the ad hoc Task Force of the ILAE Commission on Therapeutic Strategies. Epilepsia 2010, 51, 1069–1077. [Google Scholar] [CrossRef]
- Wiebe, S.; Blume, W.T.; Girvin, J.P.; Eliasziw, M. A randomized, controlled trial of surgery for temporal-lobe epilepsy. N. Engl. J. Med. 2001, 345, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Engel, J., Jr. (Ed.) Surgical treatment of the epilepsies. In Surgical Treatment of the Epilepsies, 2nd ed.; Raven Press: New York, NY, USA, 1992; pp. 23–34. [Google Scholar]
- Yardi, R.; Irwin, A.; Kayyali, H.; Gupta, A.; Nair, D.; Gonzalez-Martinez, J.; Bingaman, W.; Najm, I.M.; Jehi, L.E. Reducing versus stopping antiepileptic medications after temporal lobe surgery. Ann. Clin. Transl. Neurol. 2014, 1, 115–123. [Google Scholar] [CrossRef] [PubMed]
- de Tisi, J.; Bell, G.S.; Peacock, J.L.; McEvoy, A.W.; Harkness, W.F.; Sander, J.W.; Duncan, J.S. The long-term outcome of adult epilepsy surgery, patterns of seizure remission, and relapse: A cohort study. Lancet 2011, 378, 1388–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, J., Jr.; Wiebe, S.; French, J.; Sperling, M.; Williamson, P.; Spencer, D.; Gumnit, R.; Zahn, C.; Westbrook, E.; Enos, B. Practice parameter: Temporal lobe and localized neocortical resections for epilepsy: Report of the Quality Standards Subcommittee of the American Academy of Neurology, in association with the American Epilepsy Society and the American Association of Neurological Surgeons. Neurology 2003, 60, 538–547. [Google Scholar] [CrossRef]
- Engel, J., Jr.; McDermott, M.P.; Wiebe, S.; Langfitt, J.T.; Stern, J.M.; Dewar, S.; Sperling, M.R.; Gardiner, I.; Erba, G.; Fried, I.; et al. Early surgical therapy for drug-resistant temporal lobe epilepsy: A randomized trial. JAMA 2012, 307, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Seidenberg, M.; Hermann, B.; Wyler, A.R.; Davies, K.; Dohan, F.C., Jr.; Leveroni, C. Neuropsychological outcome following anterior temporal lobectomy in patients with and without the syndrome of mesial temporal lobe epilepsy. Neuropsychology 1998, 12, 303. [Google Scholar] [CrossRef] [PubMed]
- Hrazdil, C.; Roberts, J.I.; Wiebe, S.; Sauro, K.; Vautour, M.; Hanson, A.; Murphy, W.; Pillay, N.; Federico, P.; Jetté, N. Patient perceptions and barriers to epilepsy surgery: Evaluation in a large health region. Epilepsy Behav. 2013, 28, 52–65. [Google Scholar] [CrossRef]
- Bell, B.; Lin, J.J.; Seidenberg, M.; Hermann, B. The neurobiology of cognitive disorders in temporal lobe epilepsy. Nat. Rev. Neurol. 2011, 7, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Samulski, R.J. Engineering adeno-associated virus vectors for gene therapy. Nat. Rev. Genet. 2020, 21, 255–272. [Google Scholar] [CrossRef] [PubMed]
- Shaimardanova, A.A.; Chulpanova, D.S.; Mullagulova, A.I.; Afawi, Z.; Gamirova, R.G.; Solovyeva, V.V.; Rizvanov, A.A. Gene and Cell Therapy for Epilepsy: A Mini Review. Front. Mol. Neurosci. 2022, 15, 868531. [Google Scholar] [CrossRef] [PubMed]
- Snowball, A.; Chabrol, E.; Wykes, R.C.; Shekh-Ahmad, T.; Cornford, J.H.; Lieb, A.; Hughes, M.P.; Massaro, G.; Rahim, A.A.; Hashemi, K.S.; et al. Epilepsy Gene Therapy Using an Engineered Potassium Channel. J. Neurosci. 2019, 39, 3159–3169. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; O’neill, N.; Maffei, B.; Zourray, C.; Almacellas-Barbanoj, A.; Carpenter, J.C.; Jones, S.P.; Leite, M.; Turner, T.J.; Moreira, F.C.; et al. On-demand cell-autonomous gene therapy for brain circuit disorders. Science 2022, 378, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Colasante, G.; Qiu, Y.; Massimino, L.; Di Berardino, C.; Cornford, J.H.; Snowball, A.; Weston, M.; Jones, S.P.; Giannelli, S.G.; Lieb, A.; et al. In vivo CRISPRa decreases seizures and rescues cognitive deficits in a rodent model of epilepsy. Brain 2020, 143, 891–905. [Google Scholar] [CrossRef] [Green Version]
- Simonato, M.; Iyengar, S.; Brooks-Kayal, A.; Collins, S.; Depaulis, A.; Howells, D.W.; Jensen, F.; Liao, J.; Macleod, M.R.; Patel, M. Identification and characterization of outcome measures reported in animal models of epilepsy: Protocol for a systematic review of the literature—A TASK2 report of the AES/ILAE Translational Task Force of the ILAE. Epilepsia 2017, 58, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.A.; Moon, J.; Kim, T.J.; Jun, J.S.; Park, B.; Byun, J.I.; Sunwoo, J.S.; Park, K.I.; Lee, S.T.; Jung, K.H.; et al. Clustering of spontaneous recurrent seizures separated by long seizure-free periods: An extended video-EEG monitoring study of a pilocarpine mouse model. PLoS ONE 2018, 13, e0194552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, P.J.; Thomson, K.; Billingsley, P.; Pruess, T.; Rueda, C.; Saunders, G.W.; Smith, M.D.; Metcalf, C.S.; Wilcox, K.S. Spontaneous recurrent seizures in an intra-amygdala kainate microinjection model of temporal lobe epilepsy are differentially sensitive to antiseizure drugs. Exp. Neurol. 2022, 349, 113954. [Google Scholar] [CrossRef]
- Zeidler, Z.; Brandt-Fontaine, M.; Leintz, C.; Krook-Magnuson, C.; Netoff, T.; Krook-Magnuson, E. Targeting the Mouse Ventral Hippocampus in the Intrahippocampal Kainic Acid Model of Temporal Lobe Epilepsy. Eneuro 2018, 5, ENEURO.0158-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Mtchedlishvili, Z.; Bertram, E.H.; Erisir, A.; Kapur, J. Selective loss of dentate hilar interneurons contributes to reduced synaptic inhibition of granule cells in an electrical stimulation-based animal model of temporal lobe epilepsy. J. Comp. Neurol. 2007, 500, 876–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukau, B.; Lanzer, M.; Melchers, F.; Wallon, G. Hermann Bujard (1934–2020)—Pioneering researcher and visionary science politician. EMBO J. 2020, 39, e106981. [Google Scholar] [CrossRef] [PubMed]
- Dey, D.; Eckle, V.-S.; Vitko, I.; Sullivan, K.A.; Lasiecka, Z.M.; Winckler, B.; Stornetta, R.L.; Williamson, J.M.; Kapur, J.; Perez-Reyes, E. A potassium leak channel silences hyperactive neurons and ameliorates status epilepticus. Epilepsia 2014, 55, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Manfredsson, F.P.; Burger, C.; Rising, A.C.; Zuobi-Hasona, K.; Sullivan, L.F.; Lewin, A.S.; Huang, J.; Piercefield, E.; Muzyczka, N.; Mandel, R.J. Tight Long-term Dynamic Doxycycline Responsive Nigrostriatal GDNF Using a Single rAAV Vector. Mol. Ther. 2009, 17, 1857–1867. [Google Scholar] [CrossRef] [PubMed]
- Loew, R.; Heinz, N.; Hampf, M.; Bujard, H.; Gossen, M. Improved Tet-responsive promoters with minimized background expression. BMC Biotechnol. 2010, 10, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, W.; La Celle, M.; Rhoads, R.E. Functional Characterization of the Internal Ribosome Entry Site of eIF4G mRNA. J. Biol. Chem. 1998, 273, 5006–5012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker-Haliski, M.; White, S.H. Validated animal models for antiseizure drug (ASD) discovery: Advantages and potential pitfalls in ASD screening. Neuropharmacology 2020, 167, 107750. [Google Scholar] [CrossRef]
- Lonser, R.R.; Sarntinoranont, M.; Morrison, P.F.; Oldfield, E.H. Convection-enhanced delivery to the central nervous system. J. Neurosurg. 2015, 122, 697–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorter, J.A.; van Vliet, E.A.; Lopes da Silva, F.H. Which insights have we gained from the kindling and post-status epilepticus models? J. Neurosci. Methods 2016, 260, 96–108. [Google Scholar] [CrossRef]
- Vong, L.; Ye, C.; Yang, Z.; Choi, B.; Chua, S., Jr.; Lowell, B.B. Leptin Action on GABAergic Neurons Prevents Obesity and Reduces Inhibitory Tone to POMC Neurons. Neuron 2011, 71, 142–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.-H.; Yu, N.-K.; Baek, G.-C.; Bakes, J.; Seo, D.; Nam, H.J.; Baek, S.H.; Lim, C.-S.; Lee, Y.-S.; Kaang, B.-K. Optimization of AAV expression cassettes to improve packaging capacity and transgene expression in neurons. Mol. Brain 2014, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Kügler, S.; Kilic, E.; Bähr, M. Human synapsin 1 gene promoter confers highly neuron-specific long-term transgene expression from an adenoviral vector in the adult rat brain depending on the transduced area. Gene Ther. 2003, 10, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jirsa, V.K.; Stacey, W.C.; Quilichini, P.P.; Ivanov, A.I.; Bernard, C. On the nature of seizure dynamics. Brain 2014, 137 Pt 8, 2210–2230. [Google Scholar] [CrossRef] [Green Version]
- Straub, J.; Gawda, A.; Ravichandran, P.; McGrew, B.; Nylund, E.; Kang, J.; Burke, C.; Vitko, I.; Scott, M.; Williamson, J.; et al. Characterization of kindled VGAT-Cre mice as a new animal model of temporal lobe epilepsy. Epilepsia 2020, 61, 2277–2288. [Google Scholar] [CrossRef] [PubMed]
- Pitkanen, A.; Buckmaster, P.S.; Galanopoulou, A.S.; Moshe, S.L. Models of Seizures and Epilepsy, 2nd ed.; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Patel, A.J.; Honoré, E.; Maingret, F.; Lesage, F.; Fink, M.; Duprat, F.; Lazdunski, M. A mammalian two pore domain mechano-gated S-like K+ channel. EMBO J. 1998, 17, 4283–4290. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, J.L.; Jappelli, R.; Scheeff, E.D.; Manning, G.; Obata, K.; Brenner, S.; Callaway, E.M. Short promoters in viral vectors drive selective expression in mammalian inhibitory neurons, but do not restrict activity to specific inhibitory cell-types. Front. Neural Circuits 2009, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Roney, I.J.; Rudner, A.D.; Couture, J.-F.; Kærn, M. Improvement of the reverse tetracycline transactivator by single amino acid substitutions that reduce leaky target gene expression to undetectable levels. Sci. Rep. 2016, 6, 27697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Deyn, P.P.; Marescau, B.; Macdonald, R.L. Epilepsy and the GABA-hypothesis a brief review and some examples. Acta Neurol. Belg. 1990, 90, 65–81. [Google Scholar]
- Keaveney, M.K.; Rahsepar, B.; Tseng, H.A.; Fernandez, F.R.; Mount, R.A.; Ta, T.; White, J.A.; Berg, J.; Han, X. CaMKIIalpha-Positive Interneurons Identified via a microRNA-Based Viral Gene Targeting Strategy. J. Neurosci. 2020, 40, 9576–9588. [Google Scholar] [CrossRef] [PubMed]
- Lusby, E.; Fife, K.; Berns, K. Nucleotide sequence of the inverted terminal repetition in adeno-associated virus DNA. J. Virol. 1980, 34, 402–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szulc, J.; Wiznerowicz, M.; Sauvain, M.O.; Trono, D.; Aebischer, P. A versatile tool for conditional gene expression and knockdown. Nat. Methods 2006, 3, 109–116. [Google Scholar] [CrossRef]
- Cai, D.; Cohen, K.B.; Luo, T.; Lichtman, J.W.; Sanes, J.R. Improved tools for the Brainbow toolbox. Nat. Methods 2013, 10, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.-W.; Shaner, N.C.; Cheng, Z.; Tsien, R.Y.; Campbell, R.E. Exploration of New Chromophore Structures Leads to the Identification of Improved Blue Fluorescent Proteins. Biochemistry 2007, 46, 5904–5910. [Google Scholar] [CrossRef] [PubMed]
- McGrath, J.C.; Drummond, G.B.; McLachlan, E.M.; Kilkenny, C.; Wainwright, C.L. Guidelines for reporting experiments involving animals: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1573–1576. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Sun, H.; Rajasekaran, K.; Williamson, J.; Perez-Reyes, E.; Kapur, J. A novel therapeutic approach for treatment of catamenial epilepsy. Neurobiol. Dis. 2018, 111, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Lewczuk, E.; Joshi, S.; Williamson, J.; Penmetsa, M.; Shan, S.; Kapur, J. Electroencephalography and behavior patterns during experimental status epilepticus. Epilepsia 2017, 59, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Lothman, E.W.; Bertram, E.H.; Bekenstein, J.W.; Perlin, J.B. Self-sustaining limbic status epilepticus induced by ‘continuous’ hippocampal stimulation: Electrographic and behavioral characteristics. Epilepsy Res. 1989, 3, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Reeves, P.G.; Rossow, K.L.; Lindlauf, J. Development and Testing of the AIN-93 Purified Diets for Rodents: Results on Growth, Kidney Calcification and Bone Mineralization in Rats and Mice. J. Nutr. 1993, 123, 1923–1931. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Orientation | Sequence (5′-3′) |
|---|---|---|
| TREK-1 | F | TCCTCTTTGTGGCTCTCCCT |
| TREK-1 | R | ACACCTCGTTCTCGTAGCAG |
| mCherry | F | CTCCGACGGCCCCGTAATGC |
| mCherry | R | CGATGGTGTAGTCCTCGTTG |
| TA | F | CCGCCGTGGGCCACTTTACA |
| TA | R | ATCGTCAAGGGCGTCGGTCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, K.A.; Vitko, I.; Blair, K.; Gaykema, R.P.; Failor, M.J.; San Pietro, J.M.; Dey, D.; Williamson, J.M.; Stornetta, R.L.; Kapur, J.; et al. Drug-Inducible Gene Therapy Effectively Reduces Spontaneous Seizures in Kindled Rats but Creates Off-Target Side Effects in Inhibitory Neurons. Int. J. Mol. Sci. 2023, 24, 11347. https://doi.org/10.3390/ijms241411347
Sullivan KA, Vitko I, Blair K, Gaykema RP, Failor MJ, San Pietro JM, Dey D, Williamson JM, Stornetta RL, Kapur J, et al. Drug-Inducible Gene Therapy Effectively Reduces Spontaneous Seizures in Kindled Rats but Creates Off-Target Side Effects in Inhibitory Neurons. International Journal of Molecular Sciences. 2023; 24(14):11347. https://doi.org/10.3390/ijms241411347
Chicago/Turabian StyleSullivan, Kyle A., Iuliia Vitko, Kathryn Blair, Ronald P. Gaykema, Madison J. Failor, Jennifer M. San Pietro, Deblina Dey, John M. Williamson, Ruth L. Stornetta, Jaideep Kapur, and et al. 2023. "Drug-Inducible Gene Therapy Effectively Reduces Spontaneous Seizures in Kindled Rats but Creates Off-Target Side Effects in Inhibitory Neurons" International Journal of Molecular Sciences 24, no. 14: 11347. https://doi.org/10.3390/ijms241411347
APA StyleSullivan, K. A., Vitko, I., Blair, K., Gaykema, R. P., Failor, M. J., San Pietro, J. M., Dey, D., Williamson, J. M., Stornetta, R. L., Kapur, J., & Perez-Reyes, E. (2023). Drug-Inducible Gene Therapy Effectively Reduces Spontaneous Seizures in Kindled Rats but Creates Off-Target Side Effects in Inhibitory Neurons. International Journal of Molecular Sciences, 24(14), 11347. https://doi.org/10.3390/ijms241411347