
Tea Polyphenols Protects Tracheal Epithelial Tight Junctions in Lung during Actinobacillus pleuropneumoniae Infection via Suppressing TLR-4/MAPK/PKC-MLCK Signaling
 , ,
, ,
Abstract
:1. Introduction
2. Results
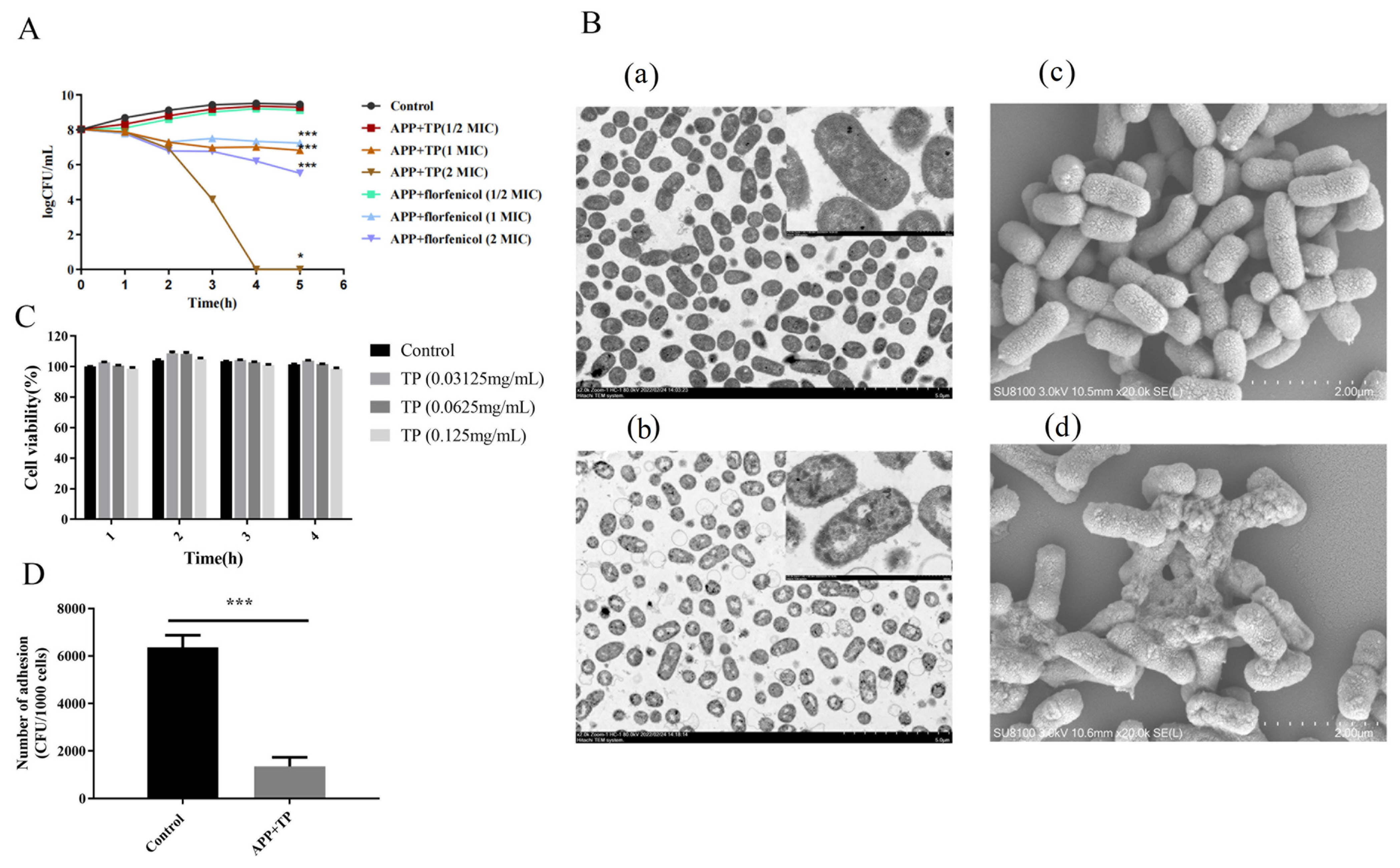
2.1. TP Inhibit the Growth of APP In Vitro
2.2. TP Affect the Cellular Integrity of APP
2.3. TP Effect on the Adhesion of APP Bacteria
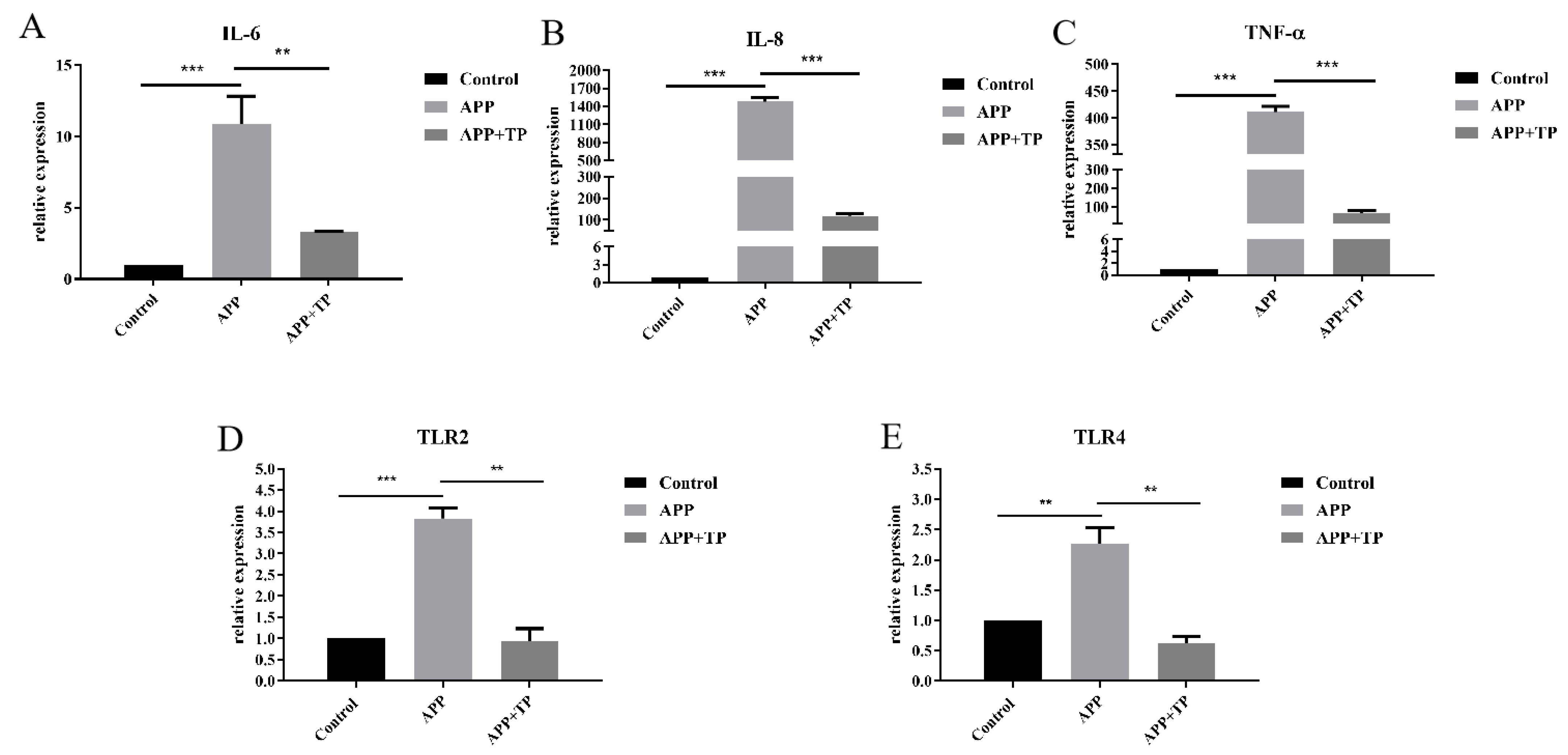
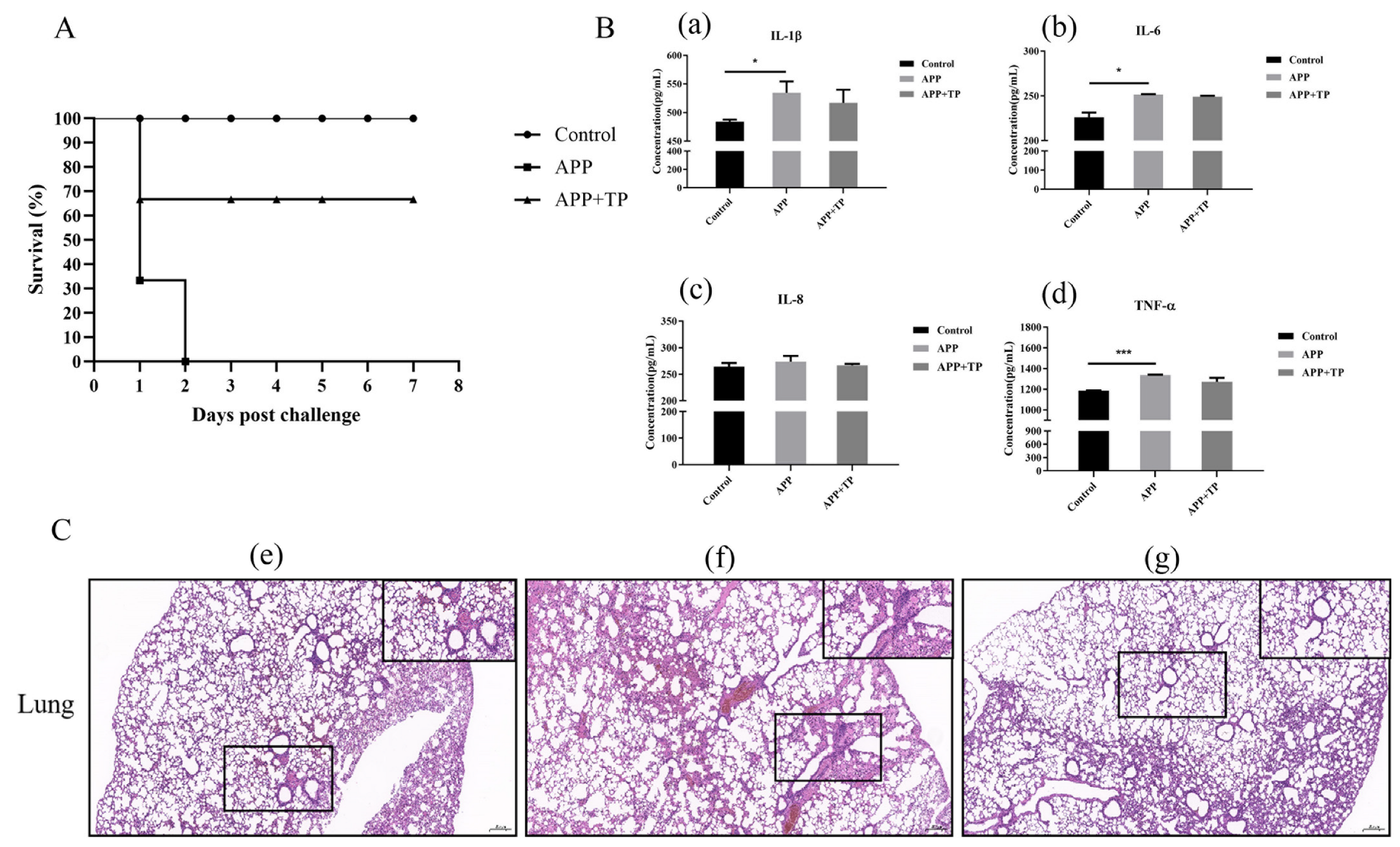
2.4. Effect of TP on Pro-Inflammatory Cytokine Secretion and the mRNA Level of TLR2 and TLR4 in NPTr Cells
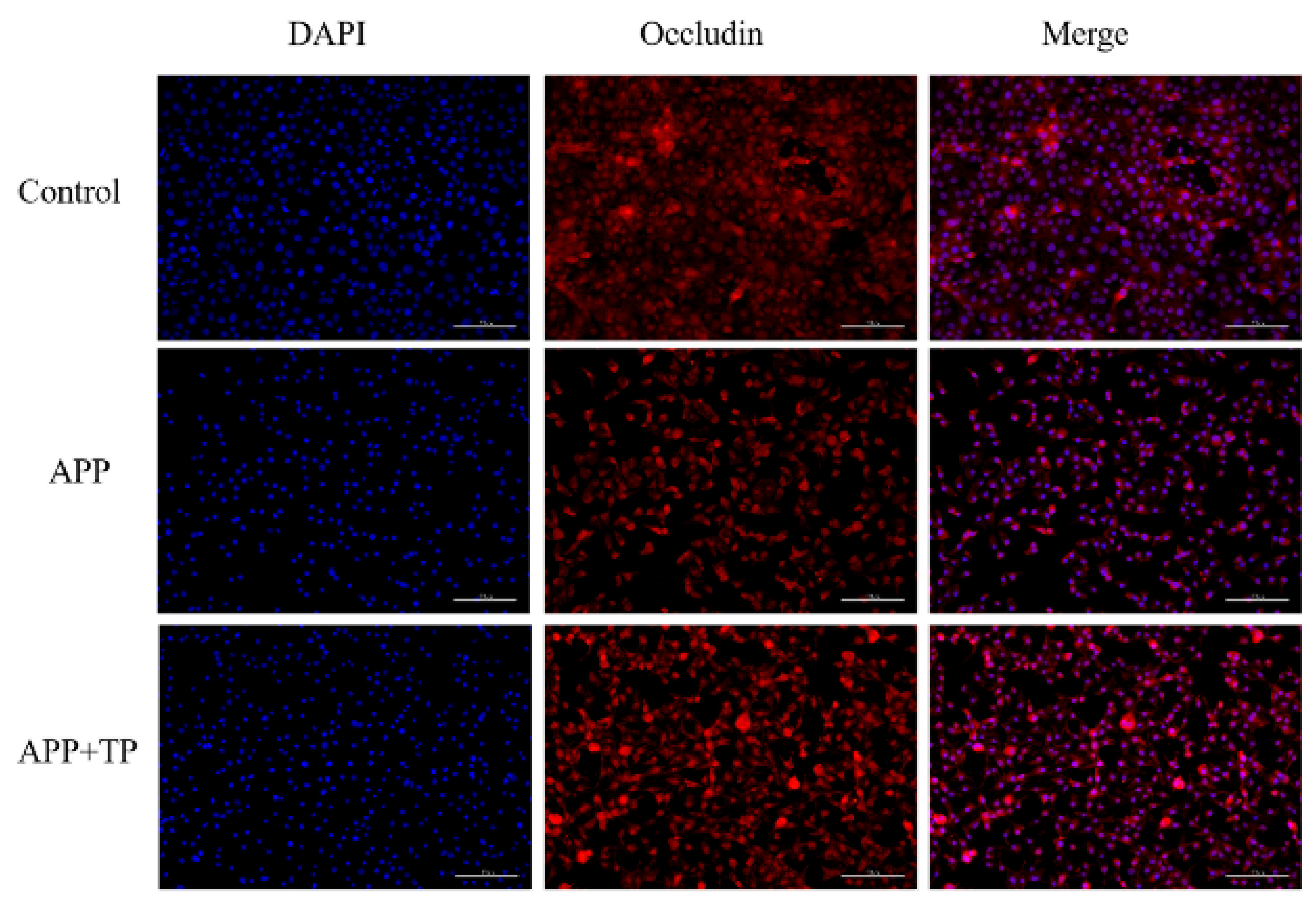
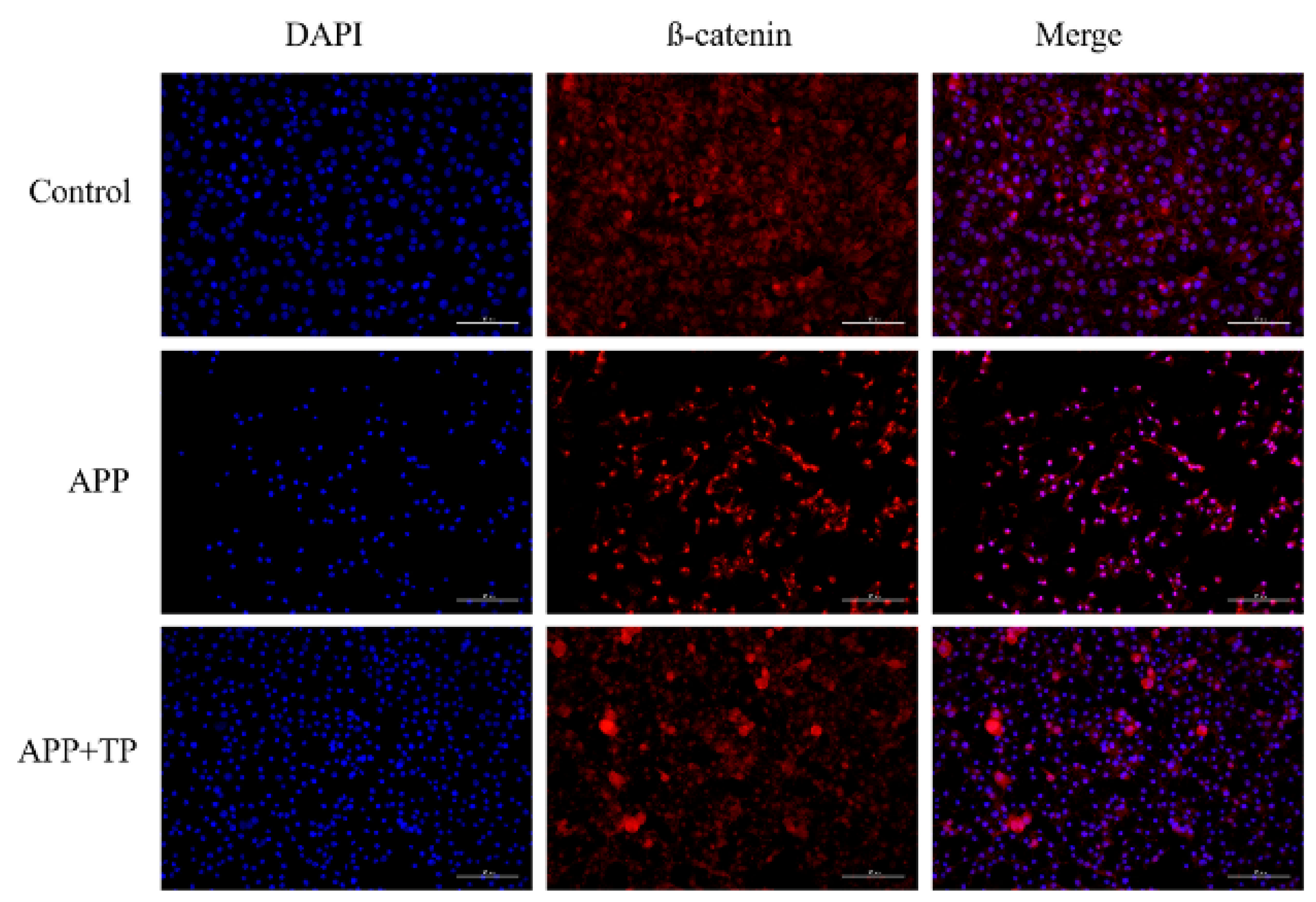
2.5. TP Decrease the Disruption of Cellular Junctions in NPTr Cells
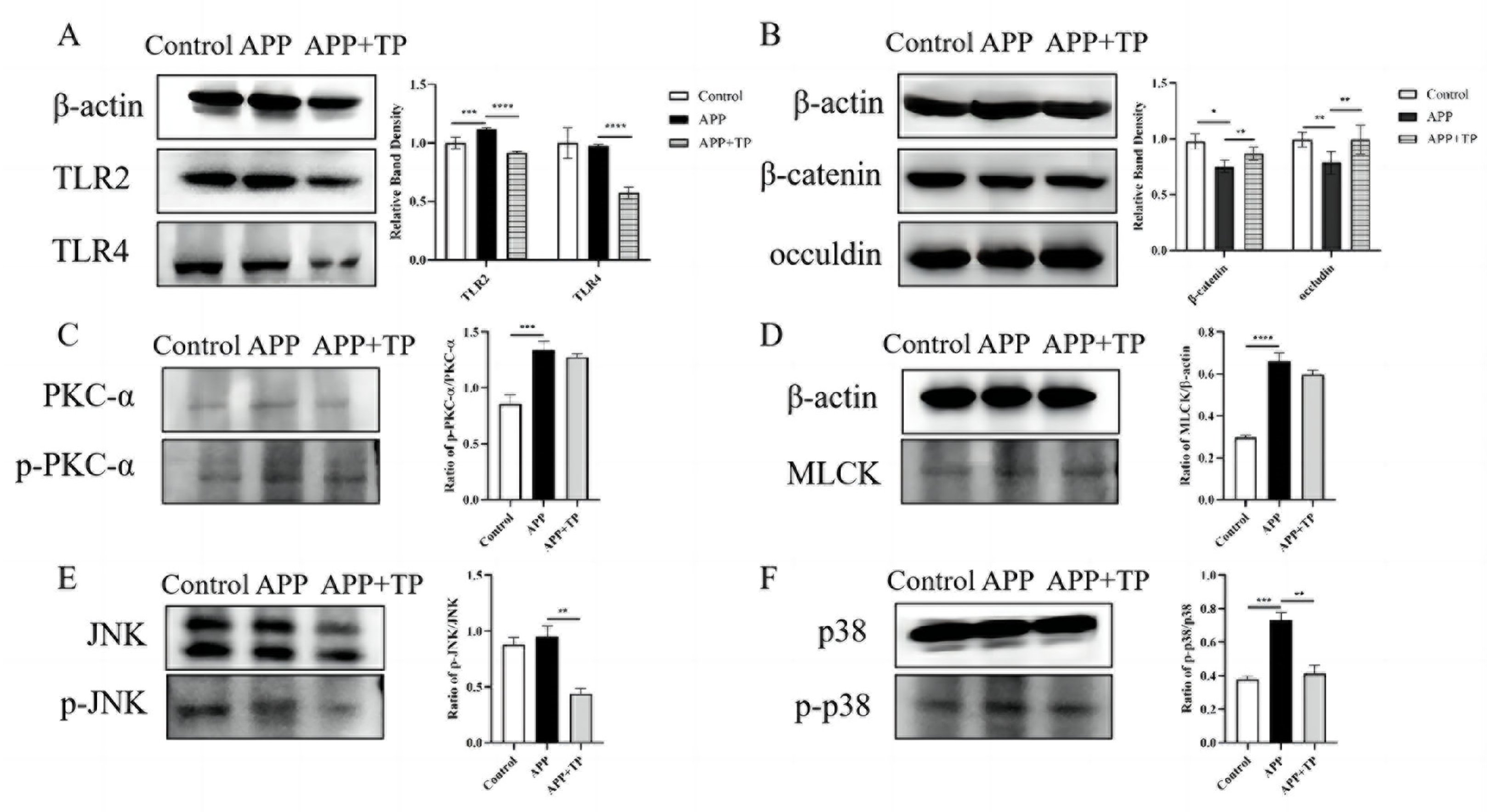
2.6. The Effect of TP on the Expression of Toll-like Receptor-Related Proteins in NPTr Cells
2.7. Effects of TP on PKC-α and MLCK Signaling Pathway Activated by APP
2.8. Effect of TP on the MAPK Signaling Pathway Activated by APP
2.9. Protective Effect of TP on Mice Infected by APP
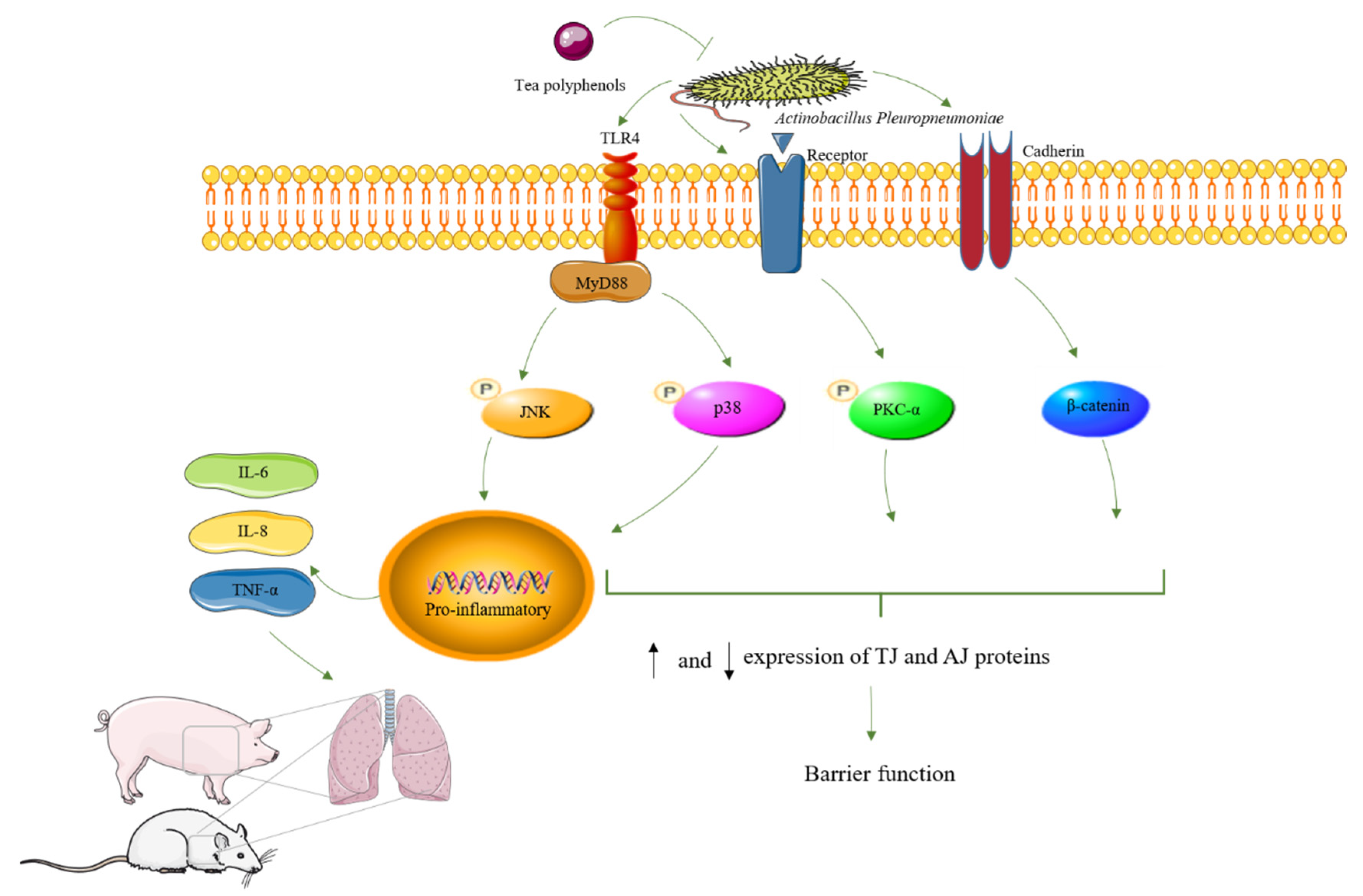
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Bacterial Culture, and TP
4.2. TP and APP Co-Cultivation Affects Bacteria Growth
4.3. Cytotoxicity Detection Assay
4.4. Adherence Assay
4.5. Analysis of Cytokine, TLR2 and TLR4 mRNA Expression Using qRT-PCR
4.6. Immunofluorescence Assay
4.7. Western Blotting
4.8. Animal Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caruana, M.; Vassallo, N. Tea Polyphenols in Parkinson’s Disease. Adv. Exp. Med. Biol. 2015, 863, 117–137. [Google Scholar] [PubMed]
- Guo, Y.J.; Sun, L.Q.; Yu, B.Y.; Qi, J. An integrated antioxidant activity fingerprint for commercial teas based on their capacities to scavenge reactive oxygen species. Food Chem. 2017, 237, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, X.; Lou, Y. Interactions of tea polyphenols with intestinal microbiota and their implication for cellular signal conditioning mechanism. J. Food Biochem. 2019, 43, e12953. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.-Q.; Liu, W.; Liu, W.-L.; Liang, R.-H.; Li, T.; Liu, C.-M.; Cao, Y.-L.; Niu, J.; Liu, Z. Characterization and Bioavailability of Tea Polyphenol Nanoliposome Prepared by Combining an Ethanol Injection Method with Dynamic High-Pressure Microfluidization. J. Agric. Food Chem. 2014, 62, 934–941. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, X.D.; Wu, C.D. The tea catechin epigallocatechin gallate suppresses cariogenic virulence factors of Streptococcus mutans. Antimicrob. Agents Chemother. 2011, 55, 1229–1236. [Google Scholar] [CrossRef] [Green Version]
- Elmassry, M.M.; Chung, E.; Cao, J.J.; Hamood, A.N.; Shen, C.L. Osteoprotective effect of green tea polyphenols and annatto-extracted tocotrienol in obese mice is associated with enhanced microbiome vitamin K2 biosynthetic pathways. J. Nutr. Biochem. 2020, 86, 108492. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X. Interactions of tea polyphenols with intestinal microbiota and their implication for anti-obesity. J. Sci. Food Agric. 2020, 100, 897–903. [Google Scholar] [CrossRef]
- Cao, D.; Zhao, M.; Wan, C.; Zhang, Q.; Tang, T.; Liu, J.; Shao, Q.; Yang, B.; He, J.; Jiang, C. Role of tea polyphenols in delaying hyperglycemia-induced senescence in human glomerular mesangial cells via miR-126/Akt-p53-p21 pathways. Int. Urol. Nephrol. 2019, 51, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Shimojima, M.; Saijo, M.; Fukasawa, M. Several catechins and flavonols from green tea inhibit severe fever with thrombocytopenia syndrome virus infection in vitro. J. Infect. Chemother. 2021, 27, 32–39. [Google Scholar] [CrossRef]
- Reygaert, W.C. The antimicrobial possibilities of green tea. Front. Microbiol. 2014, 5, 434. [Google Scholar] [CrossRef]
- Lagha, A.B.; Grenier, D. Tea polyphenols inhibit the activation of NF-kappaB and the secretion of cytokines and matrix metalloproteinases by macrophages stimulated with Fusobacterium nucleatum. Sci. Rep. 2016, 6, 34520. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Deng, Y.; Wang, H.; Liu, W.; Zhuang, X.; Chu, W. Tea polyphenols as an antivirulence compound Disrupt Quorum-Sensing Regulated Pathogenicity of Pseudomonas aeruginosa. Sci. Rep. 2015, 5, 16158. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Guo, J.; Liu, H.; Zhang, J.; Chen, X.; Qiu, Y.; Fu, S. Tea polyphenols suppress growth and virulence-related factors of Haemophilus parasuis. J. Vet. Med. Sci. 2018, 80, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zheng, B.; Wang, J.; Li, G.; Cao, S.; Wen, Y.; Huang, X.; Zuo, Z.; Zhong, Z.; Gu, Y. Quinolone Resistance of Actinobacillus pleuropneumoniae Revealed through Genome and Transcriptome Analyses. Int. J. Mol. Sci. 2021, 22, 10036. [Google Scholar] [CrossRef]
- Opriessnig, T.; Gimenez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef]
- Hoflack, G.; Maes, D.; Mateusen, B.; Verdonck, M.; de Kruif, A. Efficacy of tilmicosin phosphate (Pulmotil premix) in feed for the treatment of a clinical outbreak of Actinobacillus pleuropneumoniae infection in growing-finishing pigs. J. Vet. Med. B Infect. Dis. Vet. Public. Health 2001, 48, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Chiers, K.; De Waele, T.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F. Virulence factors of Actinobacillus pleuropneumoniae involved in colonization, persistence and induction of lesions in its porcine host. Vet. Res. 2010, 41, 65. [Google Scholar] [CrossRef] [Green Version]
- Bosse, J.T.; Janson, H.; Sheehan, B.J.; Beddek, A.J.; Rycroft, A.N.; Kroll, J.S.; Langford, P.R. Actinobacillus pleuropneumoniae: Pathobiology and pathogenesis of infection. Microbes Infect. 2002, 4, 225–235. [Google Scholar] [CrossRef]
- Stringer, O.W.; Bosse, J.T.; Lacouture, S.; Gottschalk, M.; Fodor, L.; Angen, O.; Velazquez, E.; Penny, P.; Lei, L.; Langford, P.R.; et al. Proposal of Actinobacillus pleuropneumoniae serovar 19, and reformulation of previous multiplex PCRs for capsule-specific typing of all known serovars. Vet. Microbiol. 2021, 255, 109021. [Google Scholar] [CrossRef]
- Bosse, J.T.; Li, Y.; Sarkozi, R.; Fodor, L.; Lacouture, S.; Gottschalk, M.; Casas Amoribieta, M.; Angen, O.; Nedbalcova, K.; Holden, M.T.G.; et al. Proposal of serovars 17 and 18 of Actinobacillus pleuropneumoniae based on serological and genotypic analysis. Vet. Microbiol. 2018, 217, 1–6. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, Y.; Li, L.; Zhou, R.; Xiao, S.; Wan, Y.; Zhang, S.; Wang, K.; Li, W.; Li, L.; et al. Genome biology of Actinobacillus pleuropneumoniae JL03, an isolate of serotype 3 prevalent in China. PLoS ONE 2008, 3, e1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosse, J.T.; Li, Y.; Walker, S.; Atherton, T.; Fernandez Crespo, R.; Williamson, S.M.; Rogers, J.; Chaudhuri, R.R.; Weinert, L.A.; Oshota, O.; et al. Identification of dfrA14 in two distinct plasmids conferring trimethoprim resistance in Actinobacillus pleuropneumoniae. J. Antimicrob. Chemother. 2015, 70, 2217–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilaro, A.; Novell, E.; Enrique-Tarancon, V.; Balielles, J.; Vilalta, C.; Martinez, S.; Fraile Sauce, L.J. Antimicrobial Susceptibility Pattern of Porcine Respiratory Bacteria in Spain. Antibiotics 2020, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Vanni, M.; Merenda, M.; Barigazzi, G.; Garbarino, C.; Luppi, A.; Tognetti, R.; Intorre, L. Antimicrobial resistance of Actinobacillus pleuropneumoniae isolated from swine. Vet. Microbiol. 2012, 156, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Dayao, D.; Gibson, J.S.; Blackall, P.J.; Turni, C. Antimicrobial resistance genes in Actinobacillus pleuropneumoniae, Haemophilus parasuis and Pasteurella multocida isolated from Australian pigs. Aust. Vet. J. 2016, 94, 227–231. [Google Scholar] [CrossRef]
- Dayao, D.; Gibson, J.S.; Blackall, P.J.; Turni, C. Antimicrobial resistance in bacteria associated with porcine respiratory disease in Australia. Vet. Microbiol. 2014, 171, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.C.; Chan, J.P.; Yeh, K.S.; Chang, C.C.; Hsuan, S.L.; Hsieh, Y.M.; Chang, Y.C.; Lai, T.C.; Lin, W.H.; Chen, T.H. Molecular characterization of enrofloxacin resistant Actinobacillus pleuropneumoniae isolates. Vet. Microbiol. 2010, 142, 309–312. [Google Scholar] [CrossRef]
- Kucerova, Z.; Hradecka, H.; Nechvatalova, K.; Nedbalcova, K. Antimicrobial susceptibility of Actinobacillus pleuropneumoniae isolates from clinical outbreaks of porcine respiratory diseases. Vet. Microbiol. 2011, 150, 203–206. [Google Scholar] [CrossRef]
- Pridmore, A.; Burch, D.; Lees, P. Determination of minimum inhibitory and minimum bactericidal concentrations of tiamulin against field isolates of Actinobacillus pleuropneumoniae. Vet. Microbiol. 2011, 151, 409–412. [Google Scholar] [CrossRef]
- Bercier, P.; Gottschalk, M.; Grenier, D. Effects of Actinobacillus pleuropneumoniae on barrier function and inflammatory response of pig tracheal epithelial cells. Pathog. Dis. 2019, 77, fty079. [Google Scholar] [CrossRef]
- Gon, Y.; Hashimoto, S. Role of airway epithelial barrier dysfunction in pathogenesis of asthma. Allergol. Int. 2018, 67, 12–17. [Google Scholar] [CrossRef]
- Matter, K.; Balda, M.S. Signalling to and from tight junctions. Nat. Rev. Mol. Cell Biol. 2003, 4, 225–236. [Google Scholar] [CrossRef]
- Pohunek, P. Development, structure and function of the upper airways. Paediatr. Respir. Rev. 2004, 5, 2–8. [Google Scholar] [CrossRef]
- Ganz, T. Epithelia: Not just physical barriers. Proc. Natl. Acad. Sci. USA 2002, 99, 3357–3358. [Google Scholar] [CrossRef]
- Jiao, J.; Wang, C.; Zhang, L. Epithelial physical barrier defects in chronic rhinosinusitis. Expert. Rev. Clin. Immunol. 2019, 15, 679–688. [Google Scholar] [CrossRef]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef]
- Shen, L.; Weber, C.R.; Raleigh, D.R.; Yu, D.; Turner, J.R. Tight junction pore and leak pathways: A dynamic duo. Annu. Rev. Physiol. 2011, 73, 283–309. [Google Scholar] [CrossRef] [Green Version]
- Weber, C.R. Dynamic properties of the tight junction barrier. Ann. N. Y. Acad. Sci. 2012, 1257, 77–84. [Google Scholar] [CrossRef]
- Pearce, S.C.; Al-Jawadi, A.; Kishida, K.; Yu, S.; Hu, M.; Fritzky, L.F.; Edelblum, K.L.; Gao, N.; Ferraris, R.P. Marked differences in tight junction composition and macromolecular permeability among different intestinal cell types. BMC Biol. 2018, 16, 19. [Google Scholar] [CrossRef] [Green Version]
- Yuksel, H.; Turkeli, A. Airway epithelial barrier dysfunction in the pathogenesis and prognosis of respiratory tract diseases in childhood and adulthood. Tissue Barriers 2017, 5, e1367458. [Google Scholar] [CrossRef]
- Schleimer, R.P. Immunopathogenesis of Chronic Rhinosinusitis and Nasal Polyposis. Annu. Rev. Pathol. 2017, 12, 331–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruser, L.; Bogdan, S. Adherens Junctions on the Move-Membrane Trafficking of E-Cadherin. Cold Spring Harb. Perspect. Biol. 2017, 9, a029140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coopman, P.; Djiane, A. Adherens Junction and E-Cadherin complex regulation by epithelial polarity. Cell Mol. Life Sci. 2016, 73, 3535–3553. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, A.; Rafaels, N.M.; McGirt, L.Y.; Ivanov, A.I.; Georas, S.N.; Cheadle, C.; Berger, A.E.; Zhang, K.; Vidyasagar, S.; Yoshida, T.; et al. Tight junction defects in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2011, 127, 773–786.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, H.; Marano, C.W.; Peralta Soler, A.; Mullin, J.M. Modification of tight junction function by protein kinase C isoforms. Adv. Drug Deliv. Rev. 2000, 41, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y.; Zhu, H.; Yao, X.M.; Qian, J.P.; Yang, J.; Pan, X.D.; Chen, X.D. Metformin regulates tight junction of intestinal epithelial cells via MLCK-MLC signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5239–5246. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, L.; Tapia, R.; Chamorro, D. Crosstalk of tight junction components with signaling pathways. Biochim. Biophys. Acta 2008, 1778, 729–756. [Google Scholar] [CrossRef] [Green Version]
- Cong, X.; Kong, W. Endothelial tight junctions and their regulatory signaling pathways in vascular homeostasis and disease. Cell. Signal. 2020, 66, 109485. [Google Scholar] [CrossRef]
- Yang, C.S.; Chen, G.; Wu, Q. Recent scientific studies of a traditional chinese medicine, tea, on prevention of chronic diseases. J. Tradit. Complement. Med. 2014, 4, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhang, L.; Wan, X.; Zhan, J.; Ho, C.-T. Focusing on the recent progress of tea polyphenol chemistry and perspectives. Food Sci. Hum. Wellness 2022, 11, 437–444. [Google Scholar] [CrossRef]
- Bansal, S.; Choudhary, S.; Sharma, M.; Kumar, S.S.; Lohan, S.; Bhardwaj, V.; Syan, N.; Jyoti, S. Tea: A native source of antimicrobial agents. Food Res. Int. 2013, 53, 568–584. [Google Scholar] [CrossRef]
- Ma, T.; Peng, W.; Liu, Z.; Gao, T.; Liu, W.; Zhou, D.; Yang, K.; Guo, R.; Duan, Z.; Liang, W.; et al. Tea polyphenols inhibit the growth and virulence of ETEC K88. Microb. Pathog. 2021, 152, 104640. [Google Scholar] [CrossRef]
- Auger, E.; Deslandes, V.; Ramjeet, M.; Contreras, I.; Nash, J.H.; Harel, J.; Gottschalk, M.; Olivier, M.; Jacques, M. Host-pathogen interactions of Actinobacillus pleuropneumoniae with porcine lung and tracheal epithelial cells. Infect. Immun. 2009, 77, 1426–1441. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Ortega, M.; Olivier, M.; Sizaret, P.Y.; Simon, G.; Meurens, F. Newborn pig trachea cell line cultured in air-liquid interface conditions allows a partial in vitro representation of the porcine upper airway tissue. BMC Cell Biol. 2014, 15, 14. [Google Scholar] [CrossRef]
- Cao, Q.; Wei, W.; Wang, H.; Wang, Z.; Lv, Y.; Dai, M.; Tan, C.; Chen, H.; Wang, X. Cleavage of E-cadherin by porcine respiratory bacterial pathogens facilitates airway epithelial barrier disruption and bacterial paracellular transmigration. Virulence 2021, 12, 2296–2313. [Google Scholar] [CrossRef]
- Fournier-Larente, J.; Morin, M.P.; Grenier, D. Green tea catechins potentiate the effect of antibiotics and modulate adherence and gene expression in Porphyromonas gingivalis. Arch. Oral Biol. 2016, 65, 35–43. [Google Scholar] [CrossRef]
- Lagha, A.B.; Groeger, S.; Meyle, J.; Grenier, D. Green tea polyphenols enhance gingival keratinocyte integrity and protect against invasion by Porphyromonas gingivalis. Pathog. Dis. 2018, 76, fty030. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, Z.; Wang, P.; Yu, B.; Liu, Y.; Xue, Y. Green tea polyphenols alleviate early BBB damage during experimental focal cerebral ischemia through regulating tight junctions and PKCalpha signaling. BMC Complement. Altern. Med. 2013, 13, 187. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ye, L.; Wang, X.; Liu, J.; Wang, Y.; Zhou, Y.; Ho, W. (-)-Epigallocatechin gallate inhibits endotoxin-induced expression of inflammatory cytokines in human cerebral microvascular endothelial cells. J. Neuroinflamm. 2012, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- He, W.Q.; Wang, J.; Sheng, J.Y.; Zha, J.M.; Graham, W.V.; Turner, J.R. Contributions of Myosin Light Chain Kinase to Regulation of Epithelial Paracellular Permeability and Mucosal Homeostasis. Int. J. Mol. Sci. 2020, 21, 993. [Google Scholar] [CrossRef] [Green Version]
- Walsh, S. Modulation of tight junction structure and function by cytokines. Adv. Drug Deliv. Rev. 2000, 41, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.J.; Liu, C.M.; Wong, M.Y.; Hou, L.T.; Chang, W.K. Interleukin 1beta-secreting cells in inflamed gingival tissue of adult periodontitis patients. Cytokine 1999, 11, 626–633. [Google Scholar] [CrossRef] [PubMed]
- Bloemen, V.; Schoenmaker, T.; de Vries, T.J.; Everts, V. IL-1beta favors osteoclastogenesis via supporting human periodontal ligament fibroblasts. J. Cell Biochem. 2011, 112, 1890–1897. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Kosmopoulos, M.; Christofides, A.; Drekolias, D.; Zavras, P.D.; Gargalionis, A.N.; Piperi, C. Critical Role of IL-8 Targeting in Gliomas. Curr. Med. Chem. 2018, 25, 1954–1967. [Google Scholar] [CrossRef]
- Sedger, L.M.; McDermott, M.F. TNF and TNF-receptors: From mediators of cell death and inflammation to therapeutic giants—Past, present and future. Cytokine Growth Factor. Rev. 2014, 25, 453–472. [Google Scholar] [CrossRef] [Green Version]
- Lagha, A.B.; Grenier, D. Tea polyphenols protect gingival keratinocytes against TNF-alpha-induced tight junction barrier dysfunction and attenuate the inflammatory response of monocytes/macrophages. Cytokine 2019, 115, 64–75. [Google Scholar] [CrossRef]
- Himes, S.R.; Sester, D.P.; Ravasi, T.; Cronau, S.L.; Sasmono, T.; Hume, D.A. The JNK are important for development and survival of macrophages. J. Immunol. 2006, 176, 2219–2228. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Hu, S.; Du, X.; Wen, Q.; Zhong, X.P.; Zhou, X.; Zhou, C.; Xiong, W.; Gao, Y.; Zhang, S.; et al. Vitamin B5 Reduces Bacterial Growth via Regulating Innate Immunity and Adaptive Immunity in Mice Infected with Mycobacterium tuberculosis. Front. Immunol. 2018, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Cario, E. Bacterial interactions with cells of the intestinal mucosa: Toll-like receptors and NOD2. Gut 2005, 54, 1182–1193. [Google Scholar] [CrossRef]
- Cario, E.; Gerken, G.; Podolsky, D.K. Toll-like receptor 2 controls mucosal inflammation by regulating epithelial barrier function. Gastroenterology 2007, 132, 1359–1374. [Google Scholar] [CrossRef]
- Cario, E. Barrier-protective function of intestinal epithelial Toll-like receptor 2. Mucosal Immunol. 2008, 1 (Suppl. S1), S62–S66. [Google Scholar] [CrossRef] [Green Version]
- Gu, M.J.; Song, S.K.; Lee, I.K.; Ko, S.; Han, S.E.; Bae, S.; Ji, S.Y.; Park, B.C.; Song, K.D.; Lee, H.K.; et al. Barrier protection via Toll-like receptor 2 signaling in porcine intestinal epithelial cells damaged by deoxynivalnol. Vet. Res. 2016, 47, 25. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Chen, G.; Yang, W.; Xu, Y.; Xu, Y.; Huang, X.; Liu, J.; Feng, Y.; Xu, Y.; Liu, B. Hyaluronan ameliorates LPS-induced acute lung injury in mice via Toll-like receptor (TLR) 4-dependent signaling pathways. Int. Immunopharmacol. 2015, 28, 1050–1058. [Google Scholar] [CrossRef]
- Ding, J.; Liu, Q. Toll-like receptor 4: A promising therapeutic target for pneumonia caused by Gram-negative bacteria. J. Cell Mol. Med. 2019, 23, 5868–5875. [Google Scholar] [CrossRef] [Green Version]
- Sender, V.; Stamme, C. Lung cell-specific modulation of LPS-induced TLR4 receptor and adaptor localization. Commun. Integr. Biol. 2014, 7, e29053. [Google Scholar] [CrossRef]
- Dai, J.P.; Wang, Q.W.; Su, Y.; Gu, L.M.; Zhao, Y.; Chen, X.X.; Chen, C.; Li, W.Z.; Wang, G.F.; Li, K.S. Emodin Inhibition of Influenza A Virus Replication and Influenza Viral Pneumonia via the Nrf2, TLR4, p38/JNK and NF-kappaB Pathways. Molecules 2017, 22, 1754. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.C.; Huang, S.S.; Kuo, Y.H.; Ho, Y.L.; Yang, C.S.; Chang, Y.S.; Huang, G.J. Ugonin M, a Helminthostachys zeylanica Constituent, Prevents LPS-Induced Acute Lung Injury through TLR4-Mediated MAPK and NF-kappaB Signaling Pathways. Molecules 2017, 22, 573. [Google Scholar] [CrossRef] [Green Version]
- Morin, M.P.; Bedran, T.B.; Fournier-Larente, J.; Haas, B.; Azelmat, J.; Grenier, D. Green tea extract and its major constituent epigallocatechin-3-gallate inhibit growth and halitosis-related properties of Solobacterium moorei. BMC Complement. Altern. Med. 2015, 15, 48. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | Compound | MIC (mg/mL) | MBC (mg/mL) |
|---|---|---|---|
| TSB | TP | 0.625 | 1.25 |
| Gene | Nucleotide Sequence (5′-3′) | Tm (°C) |
|---|---|---|
| GAPDH | GGCTGCCCAGAACATCATCC | 60 |
| GACGCCTGCTTCACCACCTTCTTG | ||
| IL-6 | GGAACGCCTGGAAGAAGATG | 58 |
| ATCCACTCGTTCTGTGACTG | ||
| IL-8 | TTTCTGCAGCTCTCTGTGAGG | 58 |
| CTGCTGTTGTTGTTGCTTCTC | ||
| TNF-α | CGCATCGCCGTCTCCTACCA | 60 |
| GACGCCTGCTTCACCACCTTCTTG | ||
| TLR-2 | ACGGACTGTGGTGCATGAAG | 58 |
| GGACACGAAAGCGTCATAGC | ||
| TLR-4 | CATACAGAGCCGATGGTG | 58 |
| CCTGCTGAGAAGGCGATA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Liu, Z.; Gao, T.; Liu, W.; Yang, K.; Guo, R.; Li, C.; Tian, Y.; Wang, N.; Zhou, D.; et al. Tea Polyphenols Protects Tracheal Epithelial Tight Junctions in Lung during Actinobacillus pleuropneumoniae Infection via Suppressing TLR-4/MAPK/PKC-MLCK Signaling. Int. J. Mol. Sci. 2023, 24, 11842. https://doi.org/10.3390/ijms241411842
Li X, Liu Z, Gao T, Liu W, Yang K, Guo R, Li C, Tian Y, Wang N, Zhou D, et al. Tea Polyphenols Protects Tracheal Epithelial Tight Junctions in Lung during Actinobacillus pleuropneumoniae Infection via Suppressing TLR-4/MAPK/PKC-MLCK Signaling. International Journal of Molecular Sciences. 2023; 24(14):11842. https://doi.org/10.3390/ijms241411842
Chicago/Turabian StyleLi, Xiaoyue, Zewen Liu, Ting Gao, Wei Liu, Keli Yang, Rui Guo, Chang Li, Yongxiang Tian, Ningning Wang, Danna Zhou, and et al. 2023. "Tea Polyphenols Protects Tracheal Epithelial Tight Junctions in Lung during Actinobacillus pleuropneumoniae Infection via Suppressing TLR-4/MAPK/PKC-MLCK Signaling" International Journal of Molecular Sciences 24, no. 14: 11842. https://doi.org/10.3390/ijms241411842
APA StyleLi, X., Liu, Z., Gao, T., Liu, W., Yang, K., Guo, R., Li, C., Tian, Y., Wang, N., Zhou, D., Bei, W., & Yuan, F. (2023). Tea Polyphenols Protects Tracheal Epithelial Tight Junctions in Lung during Actinobacillus pleuropneumoniae Infection via Suppressing TLR-4/MAPK/PKC-MLCK Signaling. International Journal of Molecular Sciences, 24(14), 11842. https://doi.org/10.3390/ijms241411842