
Ellagic Acid from Hull Blackberries: Extraction, Purification, and Potential Anticancer Activity

Abstract
:1. Introduction
2. Results
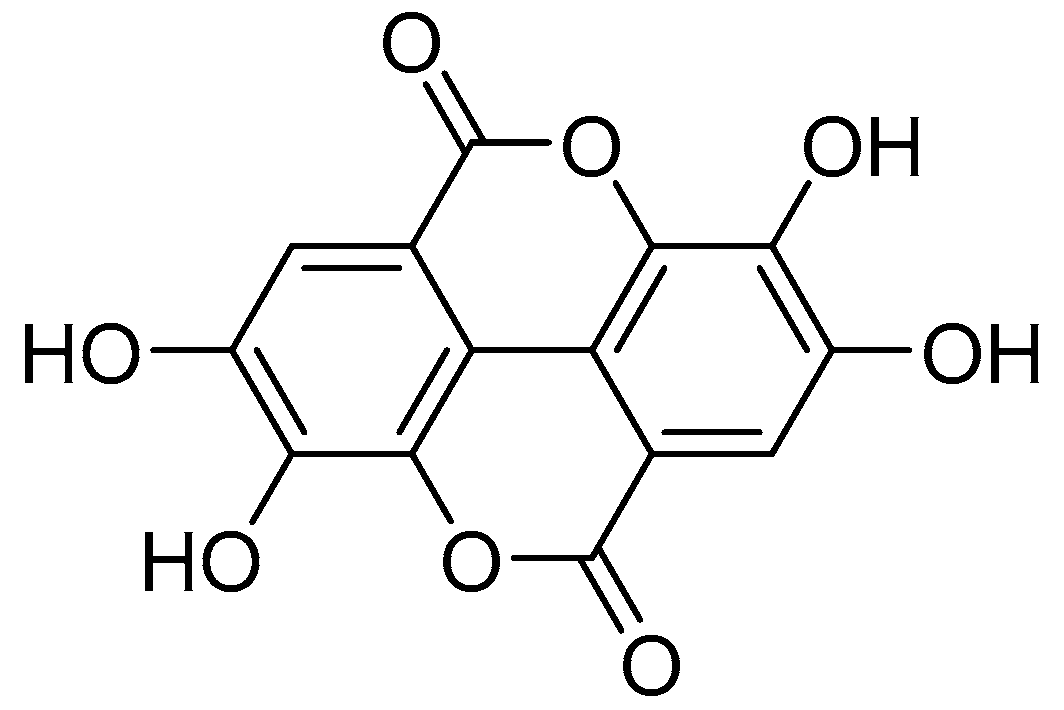
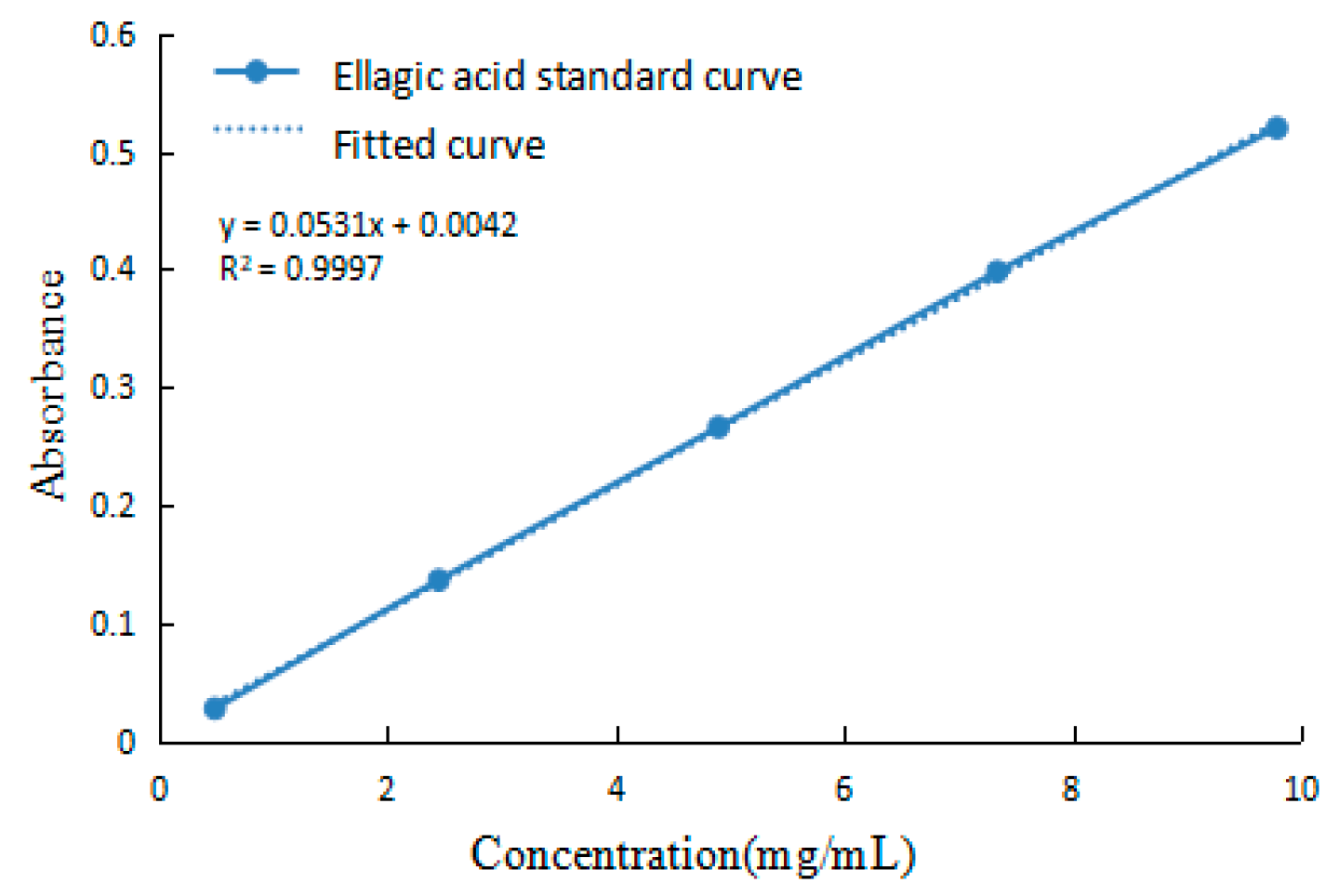
2.1. Determination of EA
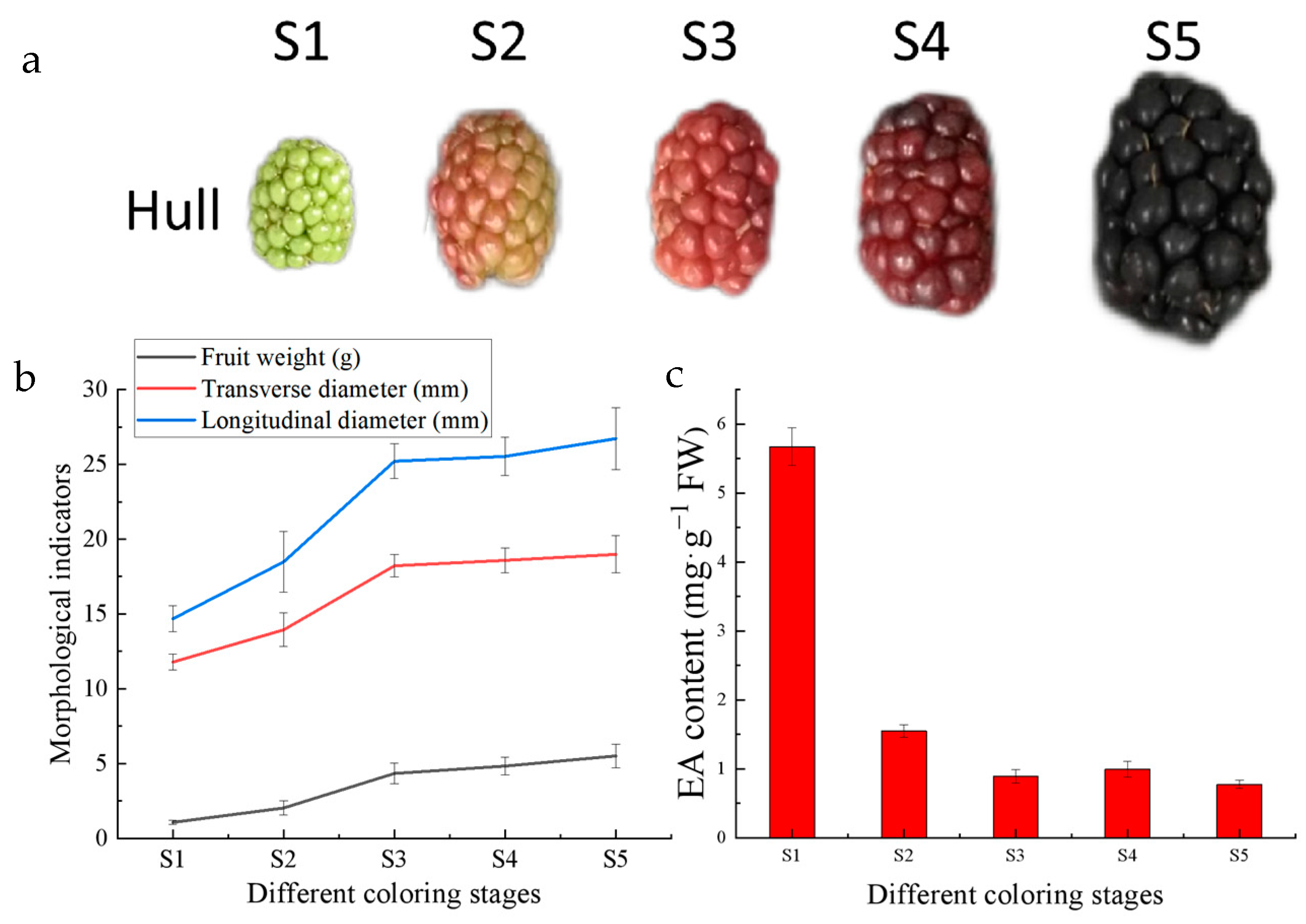
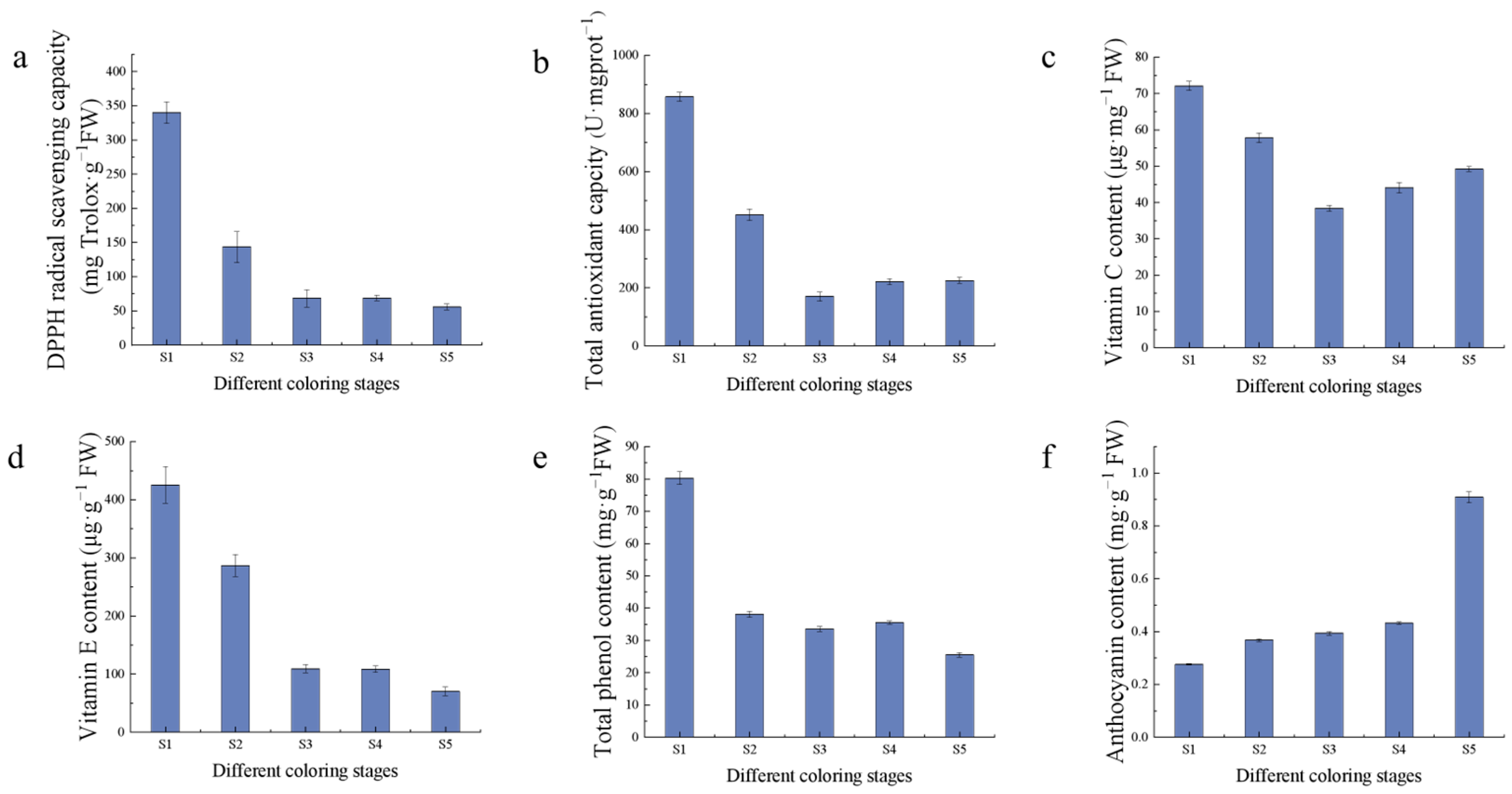
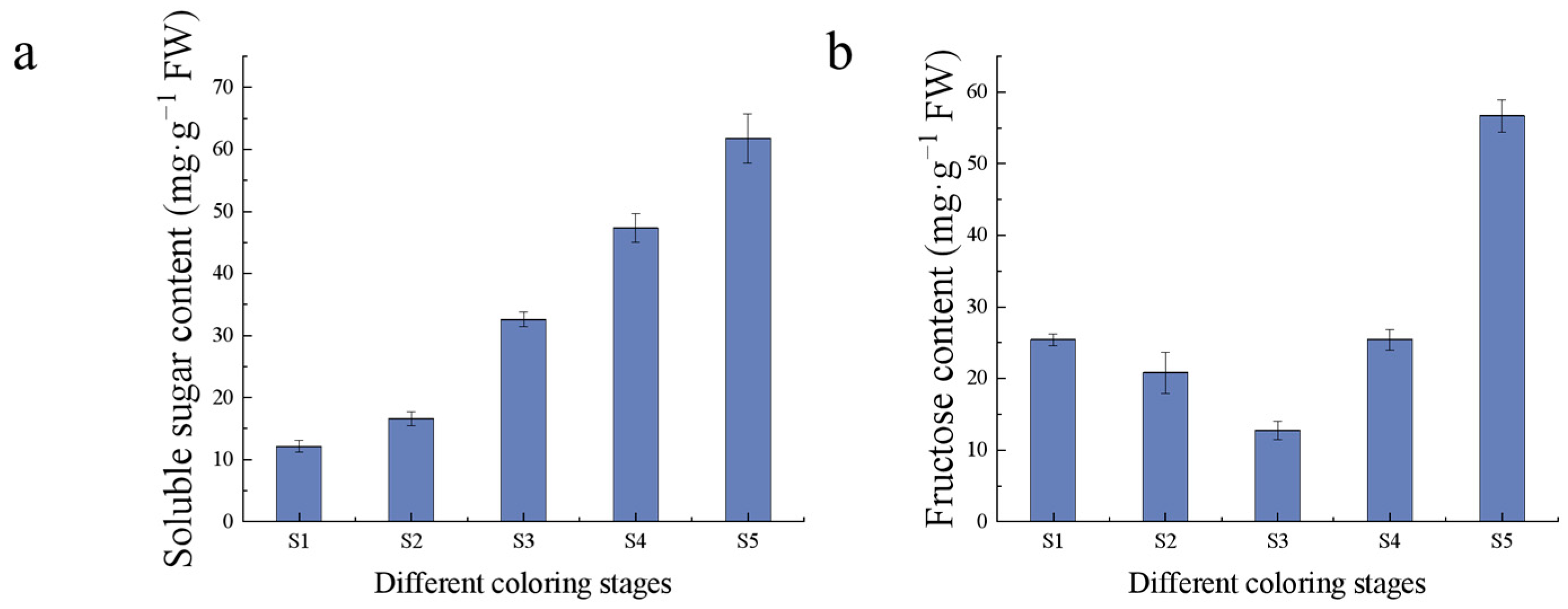
2.2. Measurement of Fruit Quality Indexes
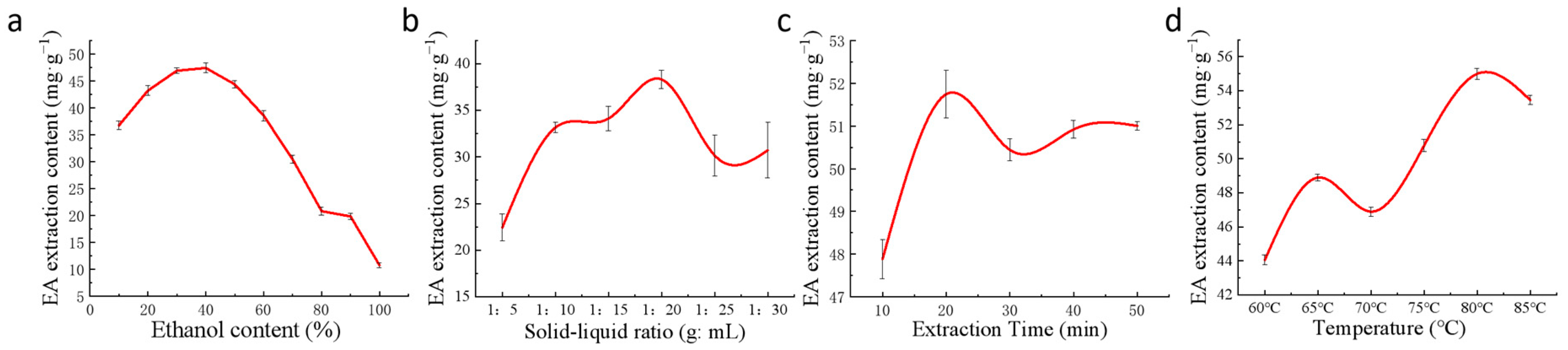
2.3. Single-Factor Tests for EA Extraction
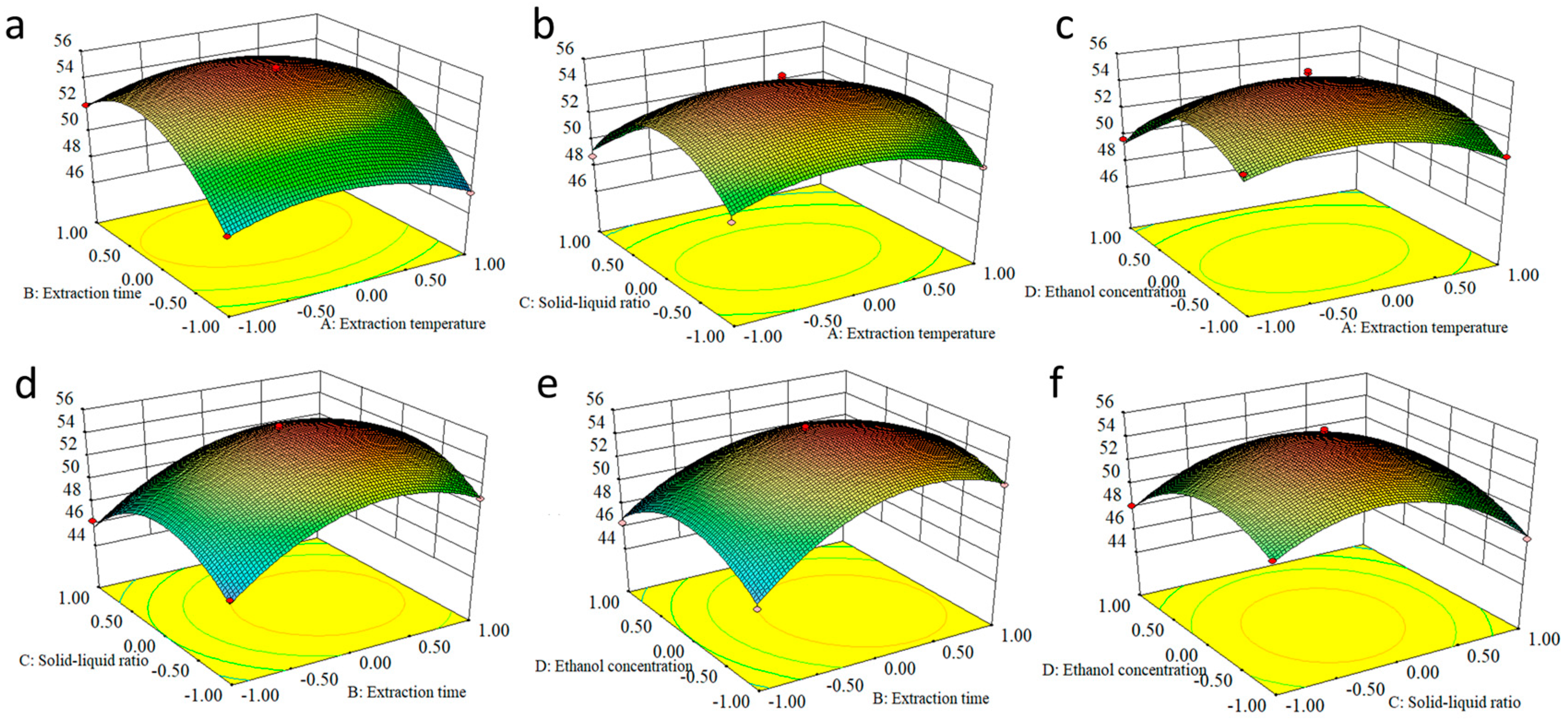
2.4. Response Surface Methodology to Optimize EA Extraction
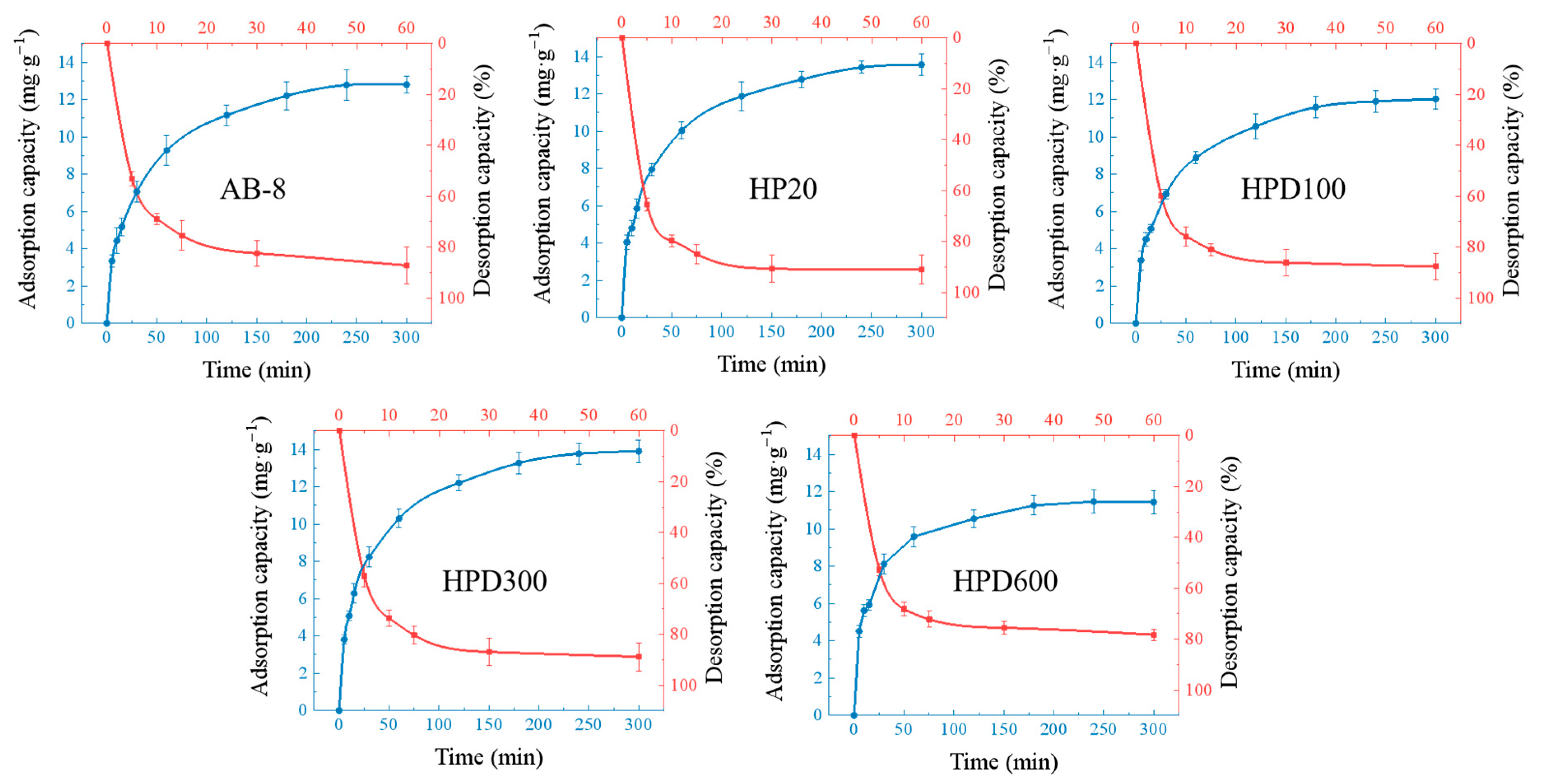
2.5. Purification of EA
2.6. Biological Evaluations
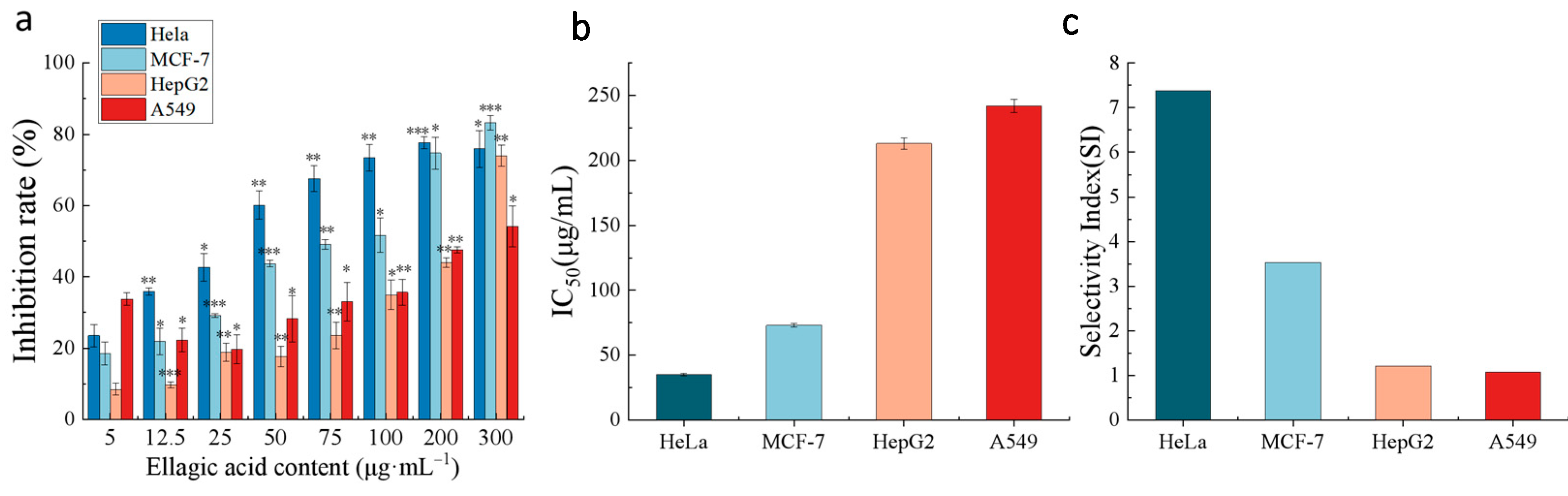
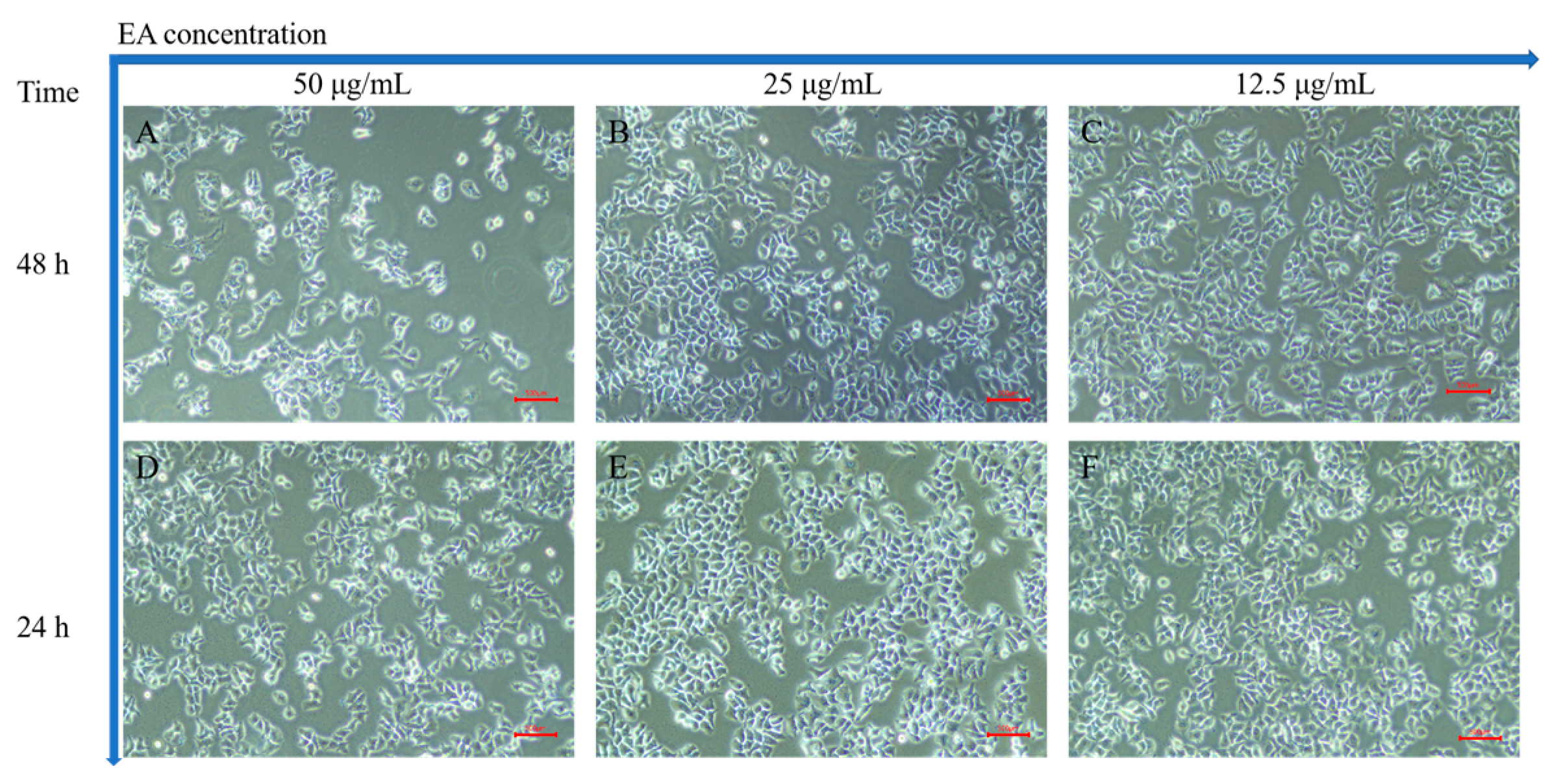
2.6.1. Anticancer Activity of EA In Vitro
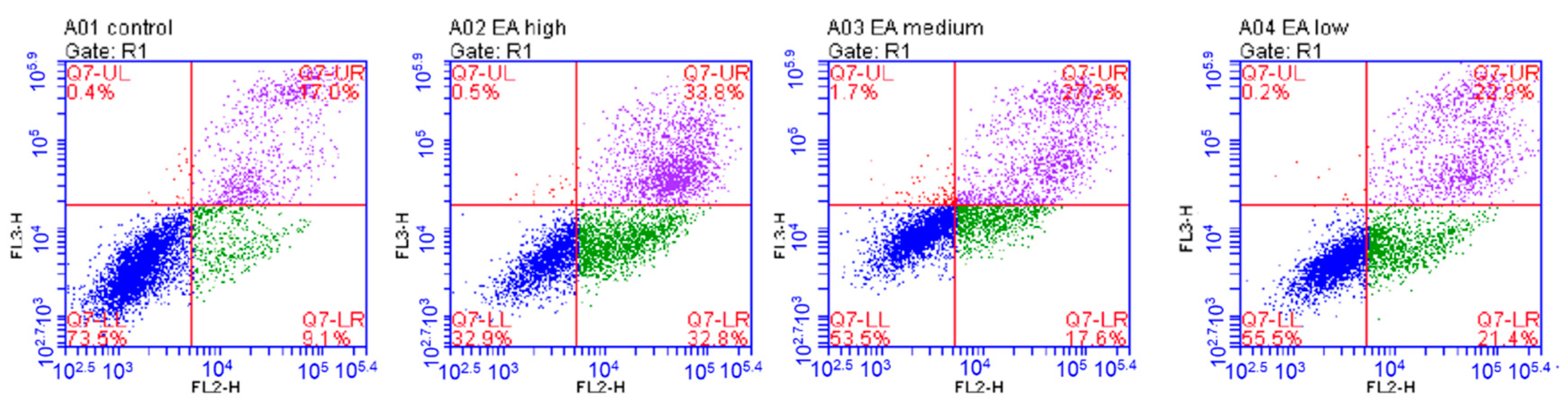
2.6.2. Induction of Apoptosis In Vitro
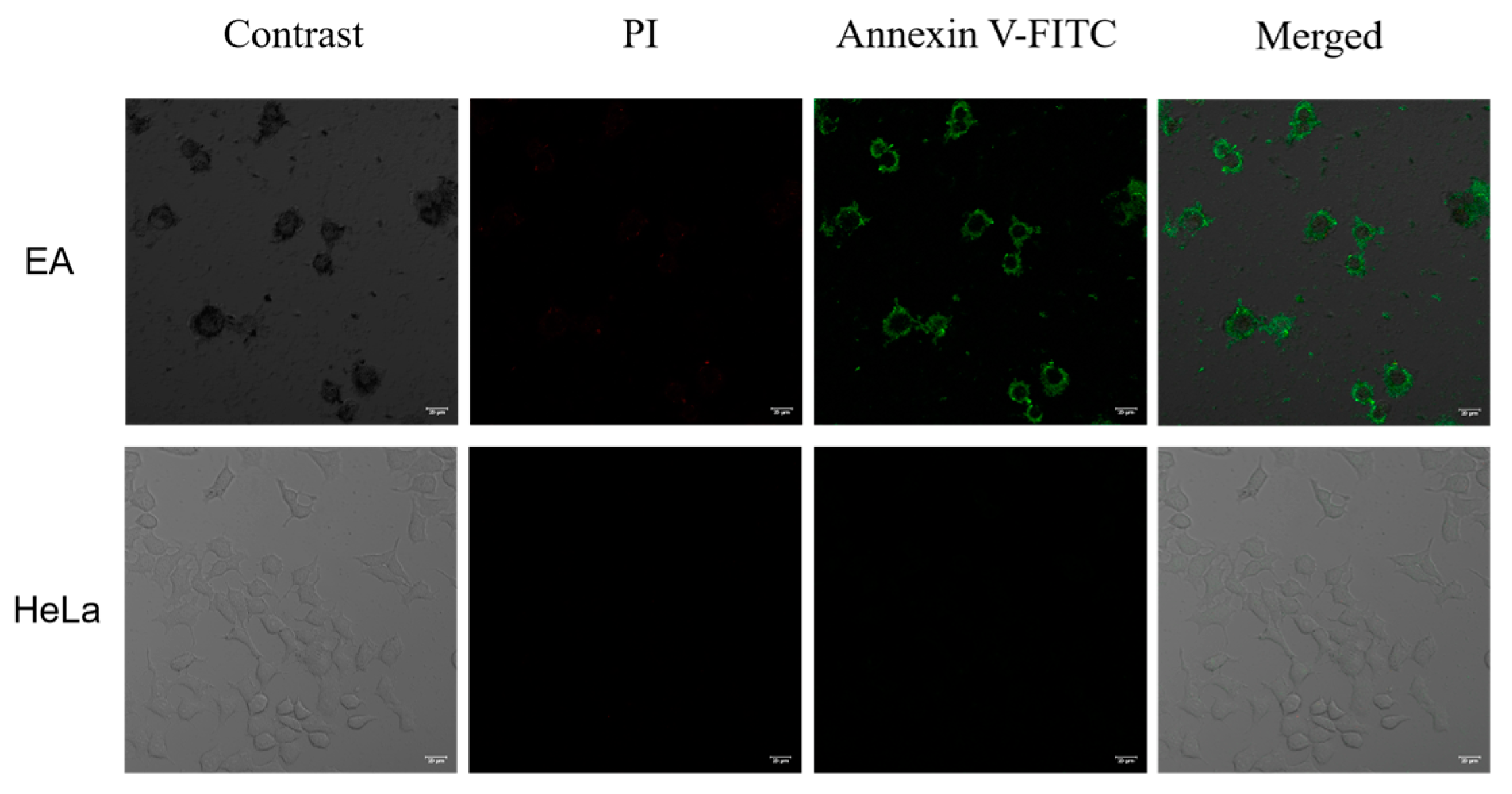
2.6.3. Confocal Fluorescence Imaging In Vitro
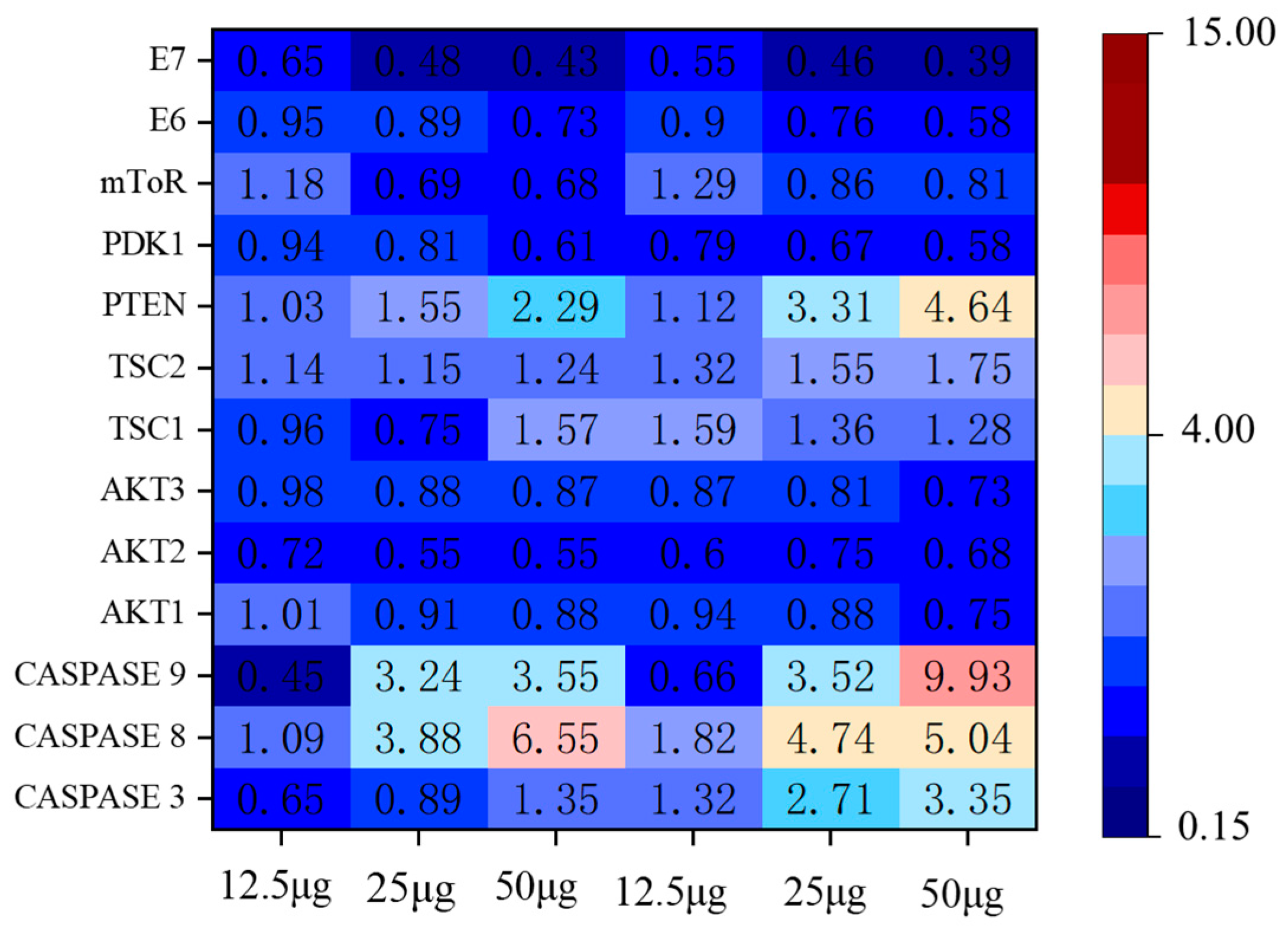
2.6.4. Apoptosis Pathway Assay
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Measurement Method of EA
4.2.1. UV Spectrophotometric Method
4.2.2. UHPLC Analysis
4.3. Measurement of Fruit Quality Indexes
4.4. Single-Factor Experiments of EA Extraction from Blackberries
4.5. Response Surface Factor Test
4.6. EA Purification Experiment
4.7. MTT Assay In Vitro
4.8. Induction of Apoptosis In Vitro
4.9. qRT–PCR Detection
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Morin, P.; Luke, R.H.; John, T.; Cindi, B.; Andy, M.; Renee, T.T.; John, R.C.; Margaret, W. Phenolics and volatiles in arkansas fresh-market blackberries (Rubus subgenus Rubus Watson). ACS Food Sci. Technol. 2022, 2, 1728–1737. [Google Scholar] [CrossRef]
- Moraes, D.P.; Machado, M.L.; Farias, C.A.A.; Barin, J.S.; Zabot, G.L.; Lozano, S.J.; Ferreira, D.F.; Vizzotto, M.; Leyva Jimenez, F.J.; Da Silveira, T.L.; et al. Effect of microwave hydrodiffusion and gravity on the extraction of phenolic compounds and antioxidant properties of blackberries (Rubus spp.): Scale-up extraction. Food Bioprocess. Technol. 2020, 13, 2200–2216. [Google Scholar] [CrossRef]
- Moghadam, D.; Zarei, R.; Tatar, M.; Khoshdel, Z.; Mashayekhi, F.J.; Naghibalhossaini, F. Antiproliferative and anti-telomerase effects of blackberry juice and berry-derived polyphenols on HepG2 liver cancer cells and normal human blood mononuclear cells. Anticancer Agents Med. Chem. 2022, 22, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Paczkowska-Walendowska, M.; Go’sciniak, A.; Szymanowska, D.; Szwajgier, D.; Baranowska-Wójcik, E.; Szulc, P.; Dreczka, D.; Simon, M.; Cielecka-Piontek, J. Blackberry leaves as new functional food screening antioxidant, anti-inflammatory and microbiological activities in correlation with phytochemical analysis. Antioxid 2021, 10, 1945. [Google Scholar] [CrossRef]
- Wu, H.; Di, Q.R.; Zhong, L.; Zhou, J.Z.; Shan, C.J.; Liu, X.L.; Ma, A.M. Enhancement on antioxidant, anti-hyperglycemic and antibacterial activities of blackberry anthocyanins by processes optimization involving extraction and purification. Front. Nutr. 2022, 9, 1007691. [Google Scholar] [CrossRef]
- Gil-Martínez, L.; Mut-Salud, N.; Ruiz-García, J.A.; Falcón-Piñeiro, A.; Maijó-Ferré, M.; Baños, A.; De la Torre-Ramírez, J.M.; Guillamón, E.; Verardo, V.; Gómez-Caravaca, A.M. Phytochemicals determination, and antioxidant, antimicrobial, anti-inflammatory and anticancer activities of blackberry fruits. Foods 2023, 12, 1505. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; De Masi, L.; et al. Ellagic acid: A review on its natural sources, chemical stability, and therapeutic potential. Oxid. Med. Cell Longev. 2022, 2, 3848084. [Google Scholar] [CrossRef]
- Wink, M. Modes of action of herbal medicines and plant secondary metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef]
- Pedro, A.Z.; Pedro, A.Z.; Jorge, E.W.; Juan, J.B.; Juan, A.A.; Juan, C.C.; Cristóbal, N.A. Ellagitannins: Bioavailability, purification and biotechnological degradation. Mini. Rev. Med. Chem. 2018, 18, 1244–1252. [Google Scholar] [CrossRef]
- Tošović, J.; Bren, U. Antioxidative action of ellagic acid-a kinetic DFT study. Antioxid 2020, 9, 587. [Google Scholar] [CrossRef]
- Ivan, S.M.; Emilija, J.; Nikolic Vesna, N.D.; Mirjana, P.M.; Srdjan, R.J.; Ivana, S.M. The effect of complexation with cyclodextrins on the antioxidant and antimicrobial activity of ellagic acid. Pharm. Dev. Technol. 2018, 24, 410–418. [Google Scholar] [CrossRef]
- Al-Mugdadi, S.F.H.; Al-Sudani, B.T.; Mohsin, R.A.; Mjali, A.J. Anticarcinogenic and antimicrobial activity effects of the ellagic acid extract. Int. J. Res. Pharm. Sci. 2019, 10, 1172–1180. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, F.; Li, B.; Song, Z.; Chen, P.; Ouyang, L. Ellagic acid exerts antitumor effects via the PI3K signaling pathway in endometrial cancer. J. Cancer 2019, 10, 3303–3314. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Li, Y.X.; Gao, H.H.; Yang, D.H.; He, X.; Fang, Y.L.; Zhou, G.B. Phenolic compound ellagic acid inhibits mitochondrial respiration and tumor growth in lung cancer. Food Funct. 2020, 11, 6332–6339. [Google Scholar] [CrossRef]
- Gil, T.Y.; Hong, C.H.; An, H.J. Anti-Inflammatory effects of ellagic acid on keratinocytes via MAPK and STAT pathways. Int. J. Mol. Sci. 2021, 22, 1277. [Google Scholar] [CrossRef] [PubMed]
- Ashutosh, G.; Ramesh, K.; Risha, G.; Singh, A.K.; Rana, H.K.; Pandey, A.K. Antioxidant, anti-inflammatory and hepatoprotective activities of Terminalia bellirica and its bioactive component ellagic acid against diclofenac induced oxidative stress and hepatotoxicity. Toxicol. Rep. 2021, 8, 44–52. [Google Scholar] [CrossRef]
- Zahin, M.; Ahmad, I.; Gupta, R.C.; Aqil, F. Punicalagin and ellagic acid demonstrate antimutagenic activity and inhibition of benzo[a]pyrene induced DNA adducts. BioMed Res. Int. 2014, 5, 467465. [Google Scholar] [CrossRef]
- Ramadan, D.T.; Ali, M.A.M.; Yahya, S.M.; El-Sayed, W.M. Correlation between Antioxidant/ antimutagenic and antiproliferative activity of some phytochemicals. Anticancer Agents Med. Chem. 2019, 19, 1481–1490. [Google Scholar] [CrossRef]
- Lin, C.; Wei, D.Z.; Xin, D.W.; Pan, J.L.; Huang, M.Y. Ellagic acid inhibits proliferation and migration of cardiac fibroblasts by down-regulating expression of HDAC1. J. Toxicol. Sci. 2019, 44, 425–433. [Google Scholar] [CrossRef]
- Javaid, N.; Shah, M.A.; Rasul, A.; Chauhdary, Z.; Saleem, U.; Khan, H.; Ahmed, N.; Uddin Md, S.; Mathew, B.; Behl, T.; et al. Neuroprotective effects of ellagic acid in alzheimer’s disease: Focus on underlying molecular mechanisms of therapeutic potential. Curr. Pharm. Des. 2021, 27, 3591–3601. [Google Scholar] [CrossRef]
- Antonio, J.A.; Carmen, G.G.; Emilio, O.; Aleix, S.V.; Iolanda, L. Ellagic acid as a tool to limit the diabetes burden: Updated evidence. Antioxid 2020, 9, 1226. [Google Scholar] [CrossRef]
- Harikrishnan, H.; Jantan, I.; Alagan, A.; Haque, M.A. Modulation of cell signaling pathways by Phyllanthus amarus and its major constituents: Potential role in the prevention and treatment of inflammation and cancer. Inflammopharmacology 2020, 28, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Shi, Y.; Wang, R.; Su, D.D.; Tang, M.F.; Liu, Y.D.; Li, Z.G. Antioxidant activity and healthy benefits of natural pigments in fruits: A review. Int. J. Mol. Sci. 2021, 22, 4945. [Google Scholar] [CrossRef]
- Syed, U.; Ponnuraj, N.; Ganapasam, S. Ellagic acid inhibits proliferation and induced apoptosis via the Akt signaling pathway in HCT-15 colon adenocarcinoma cells. Mol. Cell. Biochem. 2015, 399, 303–313. [Google Scholar] [CrossRef]
- Srigopalram, S.; Jayraaj, I.A.; Kaleeswaran, B.; Balamurugan, K.; Ranjithkumar, M.; Kumar, T.S.; Park, J.I.; Nou, I.S. Ellagic acid normalizes mitochondrial outer membrane permeabilization and attenuates inflammation-mediated cell proliferation in experimental liver cancer. Appl. Biochem. Biotechnol. 2014, 173, 2254–2266. [Google Scholar] [CrossRef]
- Shi, L.; Gao, X.; Li, X.; Jiang, N.; Luo, F.; Gu, C.; Chen, M.; Cheng, H.; Liu, P. Ellagic acid enhances the efficacy of PI3K inhibitor GDC-0941 in breast cancer cells. Curr. Mol. Med. 2015, 15, 478–486. [Google Scholar] [CrossRef]
- Mouad, E.; Irina, O.; Izumi, O.; Ilya, G.; Liang, W.G.V.; Stephen, P.; Anna, S.G. Ellagic acid induces apoptosis through inhibition of nuclear factor κB in pancreatic cancer cells. World J. Gastroenterol. 2019, 14, 3672–3680. [Google Scholar] [CrossRef]
- Tan, Y.H.; Shudo, T.; Tomoki, Y.; Yuma, S.; Ying, S.J.; Chihiro, T.; Takemoto, Y.; Akira, K. Ellagic acid, extracted from Sanguisorba officinalis, induces G1 arrest by modulating PTEN activity in B16F10 melanoma cells. Genes Cell Devot. Mol. Cell Mech. 2019, 24, 688–704. [Google Scholar] [CrossRef]
- Li, L.W.; Na, C.; Tian, S.Y.; Chen, J.; Ma, R.; Gao, Y.; Lou, G. Ellagic acid induces HeLa cell apoptosis via regulating signal transducer and activator of transcription 3 signaling. Exp. Ther. Med. 2018, 16, 29–36. [Google Scholar] [CrossRef]
- Li, K.; Yin, R.; Wang, D.Q.; Li, Q.L. Human papillomavirus subtypes distribution among 2309 cervical cancer patients in West China. Oncotarget 2017, 8, 28502–28509. [Google Scholar] [CrossRef]
- Zhang, J.J.; Cao, X.C.; Zheng, X.Y.; Wang, H.Y.; Li, Y.W. Feasibility study of a human papillomavirus E6 and E7 oncoprotein test for the diagnosis of cervical precancer and cancer. J. Int. Med. Res. 2018, 46, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, A.; Zhang, X.; Hu, S.Y.; Bao, Z.H.; Zhang, Y.H.; Jiang, X.A.; He, H.P.; Zhang, T.C. ERa-36 instead of ERa mediates the stimulatory effects of estrogen on the expression of viral oncogenes HPV E6/E7 and the malignant phenotypes in cervical cancer cells. Virus Res. 2021, 306, 198602. [Google Scholar] [CrossRef]
- Negi, S.S.; Sharma, K.; Sharma, D.; Singh, P.; Agarwala, P.; Hussain, N.; Bhargava, A.; Das, P.; Agarwal, S. Genetic analysis of human papilloma virus 16 E6/E7 variants obtained from cervical cancer cases in Chhattisgarh, a central state of India. Virusdisease 2021, 32, 492–503. [Google Scholar] [CrossRef]
- Wen, N.; Bian, L.H.; Gon, J.; Meng, Y.G. RPRD1B is a potentially molecular target for diagnosis and prevention of human papillomavirus E6/E7 infection-induced cervical cancer: A case-control study. Asia-Pac. J. Clin. Oncol. 2020, 17, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Butz, K.; Geisen, C.; Ullmann, A.; Spitkovsky, D.; Hoppe-Seyler, F. Cellular responses of HPV-positive cancer cells to genotoxic anti-cancer agents: Repression of E6/E7-oncogene expression and induction of apoptosis. Int. J. Cancer 1996, 68, 506–513. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Wang, J.L.; Wang, W.F.; Lyu, L.F.; Wu, W.L.; Li, W.L. The extraction and high antiproliferative effect of anthocyanin from gardenblue blueberry. Molecules 2023, 28, 2850. [Google Scholar] [CrossRef]
- Hanis, N.A.A.; Aini, N.A.; Suzana, Y.; Mohd, Y.M.H. Ultrasonic extraction of 2-acetyl-1-pyrroline (2AP) from pandanus amaryllifolius roxb. Using ethanol as solvent. Molecules 2022, 27, 4906. [Google Scholar] [CrossRef]
- Li, X.P. Study on Extraction, Refining and Oxidationresistant and Antimicrobiall Effects of Ellagic Acid from Red Raspberry. Ph.D. Thesis, Gansu Agriculture University, Lanzhou China, 2010. [Google Scholar]
- Wang, J.H.; Yu, X.Y.; Liao, S.L.; Chen, S.K.; Zhao, Y.; Fu, Q. Optimization of extraction process of ellagic acid from raspberry. For. By-Prod. Spéc. China 2017, 27, 14–17. [Google Scholar] [CrossRef]
- Wang, J.N. Separation and Purification of Ellagic Acid of and Its Effect on Lipid Peroxidation in Lonicera caerulea L. Ph.D Thesis, Harbin University Commerce, Harbin, China, 2018. [Google Scholar]
- Paulo Isaac, D.A.; Edemilson, C.C.; Leonardo, L.B.; Joelma, A.M.P. Development and validation of a HPLC-UV method for the evaluation of ellagic acid in liquid extracts of Eugenia uniflora L. (Myrtaceae) leaves and its ultrasound-assisted extraction optimization. evidence-based complement. Altern. Med. 2017, 2017, 1501038. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Du, L.L.; Wang, J.L.; Liu, H.X.; Zhao, H.F.; Lyu, L.F.; Wang, W.F.; Wu, W.L.; Li, W.L. Polyphenols from Prunus mume: Extraction, purification and anticancer activity. Food Funct. 2023, 14, 4380–4391. [Google Scholar] [CrossRef]
- Kumar, D.; Basu, S.; Parija, L.; Rout, D.; Manna, S.; Dandapat, J.; Debata, P.R. Curcumin and ellagic acid synergistically induce ROS generation, DNA damage, p53 accumulation and apoptosis in HeLa cervical carcinoma cells. Biomed. Pharmacother. 2016, 6, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Mark, S.D. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Feng, Y.Q.; Zou, W.; Hu, C.H.; Li, G.Y.; Zhou, S.H.; He, Y.; Ma, F.; Deng, C.; Sun, L.L. Modulation of CASC2/miR-21/PTEN pathway sensitizes cervical cancer to cisplatin. Arch. Biochem. Biophys. 2017, 6, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Du, G.H.; Cao, D.M.; Meng, L.Z. miR-21 inhibitor suppresses cell proliferation and colony formation through regulating the PTEN/AKT pathway and improves paclitaxel sensitivity in cervical cancer cells. Mol. Med. Rep. 2017, 15, 2713–2719. [Google Scholar] [CrossRef]
- van Veelen, W.; Korsse, S.E.; van de Laar, L.; Peppelenbosch, M.P. The long and winding road to rational treatment of cancer associated with LKB1/AMPK/TSC/mTORC1 signaling. Oncog 2011, 30, 2289–2303. [Google Scholar] [CrossRef]
- Ken, I.; Li, Y.; Xu, T.; Guan, K.L. Rheb GTPase is a direct target of TSC2 GAP activity and regulates mTOR signaling. Genes Dev. 2003, 17, 1829–1834. [Google Scholar] [CrossRef]
- Wu, X.; Schauss, A.G. Mitigation of inflammation with foods. J. Agric. Food Chem. 2012, 60, 6703–6717. [Google Scholar] [CrossRef]
- Jean-Gilles, D.; Li, L.; Ma, H.; Yuan, T.; Chichester, C.O.; Seeram, N.P. Anti-inflammatory effects of polyphenolic-enriched red raspberry extract in an antigen-induced arthritis rat model. J. Agric. Food Chem. 2012, 60, 5755–5762. [Google Scholar] [CrossRef]
- Chen, Z.; Jiang, J.Y.; Shu, L.Z.; Li, X.B.; Huang, J.; Qian, B.Y.; Wang, X.D.; Li, X.; Chen, J.X.; Xu, H.D. Combined transcriptomic and metabolic analyses reveal potential mechanism for fruit development and quality control of Chinese raspberry (Rubus chingii Hu). Plant Cell Rep. 2021, 40, 1–24. [Google Scholar] [CrossRef]
- Lei, F.; Xing, D.M.; Xiang, L.; Zhao, L.X.; Wang, W.; Zhang, L.J. Pharmacokinetic study of ellagic acid in rat after oral administration of pomegranate leaf extract. Drug Evaluat. 2005, 2, 38–41. [Google Scholar] [CrossRef]
- Zhou, J.; Zhou, B.X.; Tu, J.; Zhang, C.; Luo, Y.; Liu, Y. Pharmacokinetic study of ellagic acid and gallic acid of Punica granatum L. husk extract in rats. China Med. Herald. 2015, 12, 19–23. [Google Scholar]
- Zhao, H.F.; Lyu, L.F.; Zhang, C.H.; Wang, X.M.; Li, W.L.; Wu, W.L. Rubus‘Ningzhi 3′: A new blackberry cultivar. J. Nanjing For. Univ. 2020, 44, 231–232. [Google Scholar] [CrossRef]
- Zhang, C.H.; Lyu, L.F.; Zhao, H.F.; Wang, X.M.; Li, W.L.; Wu, W.L. Rubus‘Shuofeng’: A new blackberry cultivar. J. Nanjing For. Univ. 2020, 44, 245–246. [Google Scholar] [CrossRef]
- Chen, G.; Huang, L.S.; Xu, J.; Jian, S.P.; Wang, H.L. Ultrasonic-assisted extraction and antioxidant activity of antho cyanins from blackberry fruits. Food Sci. 2012, 33, 117–121. [Google Scholar]
- Li, J.; Nie, J.Y.; Li, H.F.; Xu, G.F.; Wang, X.D.; Wu, Y.L.; Wang, Z.X. On determination conditions for total polyphenols in fruits and its derived products by Folin- phenol methods. J. Fruit Sci. 2008, 25, 126–131. [Google Scholar] [CrossRef]
- Wu, W.L.; Sun, Z.J.; Cai, J.H. The five varieties of black poison are “Hull” and “Chester” and their cultivation techniques. China Fruit 1995, 4, 16–18. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Wang, L.; Zhang, J.J.; Xiong, X.M.; Zhang, D.; Tang, X.M.; Luo, X.J.; Ma, Q.L. Vascular peroxide 1 promotes ox-LDL-induced programmed necrosis in endothelial cells through a mechanism involving β-catenin signaling. Atheroscler 2018, 274, 128–138. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Zhao, H.F.; Wu, W.L.; Wang, W.F.; Li, W.L. Research on anthocyanins from Rubus “Shuofeng” as potential antiproliferative and apoptosis-inducing agents. Foods 2023, 12, 1216. [Google Scholar] [CrossRef]
- Croge, P.C.; Cuquel, L.F.; Pintro, T.M.P.; Biasi, L.A.; Claudine, M.B. Antioxidant capacity and polyphenolic compounds of blackberries produced in different climates. HortScience 2019, 54, 2209–2213. [Google Scholar] [CrossRef]
- Neth, B.J.; Bauer, B.A.; Benarroch, E.E. The role of oxidative stress in Parkinson’s disease. Antioxyd 2020, 9, 461–491. [Google Scholar] [CrossRef]
- Mar, L.; Antonio, G.S.; Teresa, M.G.; Francisco, A.T.; Juan, C.E. Urolithins, ellagic acid-derived metabolites produced by human colonic microflora, exhibit estrogenic and antiestrogenic activities. J. Agric. Food Chem. 2006, 54, 1611–1620. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | A (°C) | B (min) | C (mL/g) | D/% | Ellagic Acid Content (mg/g) |
|---|---|---|---|---|---|
| 1 | 1 | 0 | 1 | 0 | 48.28 |
| 2 | 0 | 0 | 0 | 0 | 54.84 |
| 3 | 1 | 1 | 0 | 0 | 51.24 |
| 4 | 0 | −1 | 1 | 0 | 46.26 |
| 5 | 0 | 0 | −1 | 1 | 48.12 |
| 6 | −1 | 0 | −1 | 0 | 50.21 |
| 7 | 0 | 1 | 1 | 0 | 49.06 |
| 8 | −1 | 0 | 1 | 0 | 48.73 |
| 9 | 0 | −1 | −1 | 0 | 47.22 |
| 10 | 0 | 1 | 0 | 1 | 48.82 |
| 11 | 0 | 1 | −1 | 0 | 50.83 |
| 12 | 1 | 0 | 0 | −1 | 50.75 |
| 13 | 0 | 0 | 0 | 0 | 54.86 |
| 14 | −1 | −1 | 0 | 0 | 48.62 |
| 15 | 1 | 0 | 0 | 1 | 49.51 |
| 16 | 0 | −1 | 0 | 1 | 46.36 |
| 17 | 0 | 0 | 1 | 1 | 47.11 |
| 18 | 0 | 0 | 1 | −1 | 47.95 |
| 19 | −1 | 0 | 0 | 1 | 49.70 |
| 20 | 1 | 0 | −1 | 0 | 50.10 |
| 21 | −1 | 1 | 0 | 0 | 52.04 |
| 22 | 0 | 0 | 0 | 0 | 55.55 |
| 23 | 0 | 1 | 0 | −1 | 52.18 |
| 24 | −1 | 0 | 0 | −1 | 52.50 |
| 25 | 1 | −1 | 0 | 0 | 47.52 |
| 26 | 0 | 0 | 0 | 0 | 55.72 |
| 27 | 0 | −1 | 0 | −1 | 46.87 |
| 28 | 0 | 0 | −1 | −1 | 50.95 |
| 29 | 0 | 0 | 0 | 0 | 54.71 |
| Source | Sum of Squares | DF | Mean Square | F Value | p Value |
|---|---|---|---|---|---|
| Model | 223.41 | 14 | 15.96 | 80.66 | <0.0001 |
| A | 1.61 | 1 | 1.61 | 8.16 | 0.0127 |
| B | 37.85 | 1 | 37.85 | 191.3 | <0.0001 |
| C | 8.42 | 1 | 8.42 | 42.55 | <0.0001 |
| D | 11.16 | 1 | 11.16 | 56.42 | <0.0001 |
| AB | 0.023 | 1 | 0.023 | 0.12 | 0.7385 |
| AC | 0.029 | 1 | 0.029 | 0.15 | 0.708 |
| AD | 0.62 | 1 | 0.62 | 3.12 | 0.099 |
| BC | 0.17 | 1 | 0.17 | 0.85 | 0.3734 |
| BD | 2.04 | 1 | 2.04 | 10.34 | 0.0062 |
| CD | 0.99 | 1 | 0.99 | 5 | 0.0422 |
| A2 | 22.71 | 1 | 22.71 | 114.81 | <0.0001 |
| B2 | 74.74 | 1 | 74.74 | 377.81 | <0.0001 |
| C2 | 87.38 | 1 | 87.38 | 441.69 | <0.0001 |
| D2 | 55.37 | 1 | 55.37 | 279.86 | <0.0001 |
| Residual | 2.77 | 14 | 0.2 | ||
| Lack of fit | 1.91 | 10 | 0.19 | 0.88 | 0.6044 |
| Pure error | 0.86 | 4 | 0.22 | ||
| Cor total | 226.18 | 28 | |||
| R-squared | 0.9878 | ||||
| Pred R-squared | 0.9455 | ||||
| Adj R-squared | 0.9755 |
| Factors | Code | Coding Level | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| Ethanol concentration | A | 30% | 40% | 50% |
| Solid–liquid ratio | B | 1:15 | 1:20 | 1:25 |
| Extraction time | C | 10 min | 20 min | 30 min |
| Extraction temperature | D | 75 °C | 80 °C | 85 °C |
| Gene Name | Forward (5′-3′) | Reverse (5′-3′) | Amplification Length (bp) |
|---|---|---|---|
| Caspase 3 | ACCAGTGGAGGCCGACTTCT | GCATGGCACAAAGCGACTGG | 107 |
| Caspase 8 | ACCGAAACCCTGCAGAGGGA | CATCGCCTCGAGGACATCGC | 78 |
| Caspase 9 | GATGCCCTGTGTCGGTCGAG | GTGGAGGCCACCTCAAACCC | 139 |
| AKT1 | GCGGCACACCTGAGTACCTG | CAGGCGACCGCACATCATCT | 113 |
| AKT2 | CAGAACACCAGGCACCCGTT | GCCCGCTCCTCTGTGAAGAC | 143 |
| AKT3 | TGTCGAGAGAGCGGGTGTTC | TGGTGGCTGCATCTGTGATCC | 198 |
| PTEN | TCCCAGTCAGAGGCGCTATGT | CCGTCGTGTGGGTCCTGAAT | 199 |
| TSC1 | AAGCTTGGGCCTGACACACC | CTGTCTCCCGCAGGGCTTTC | 86 |
| TSC2 | ATCTGCAGCGTGGAGATGCC | GTGTACGGCAGGGAGATGGC | 200 |
| mTOR | GCCTTTCCTGCGCAAGATGC | GCGGGCACTCTGCTCTTTGA | 85 |
| PI3K | AGGAGATCGCTCTGGCCTCA | TGGCTCGGTCCAGGTCATCC | 161 |
| E7 | GGACGGGCCAGATGGACAAG | GGGTTCGTACGTCGGTTGCT | 122 |
| E6 | TGTGTCAGGCGTTGGAGACAT | ACCTCAGATCGCTGCAAAGT | 82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhao, F.; Wu, W.; Lyu, L.; Li, W.; Zhang, C. Ellagic Acid from Hull Blackberries: Extraction, Purification, and Potential Anticancer Activity. Int. J. Mol. Sci. 2023, 24, 15228. https://doi.org/10.3390/ijms242015228
Wang J, Zhao F, Wu W, Lyu L, Li W, Zhang C. Ellagic Acid from Hull Blackberries: Extraction, Purification, and Potential Anticancer Activity. International Journal of Molecular Sciences. 2023; 24(20):15228. https://doi.org/10.3390/ijms242015228
Chicago/Turabian StyleWang, Jialuan, Fengyi Zhao, Wenlong Wu, Lianfei Lyu, Weilin Li, and Chunhong Zhang. 2023. "Ellagic Acid from Hull Blackberries: Extraction, Purification, and Potential Anticancer Activity" International Journal of Molecular Sciences 24, no. 20: 15228. https://doi.org/10.3390/ijms242015228
APA StyleWang, J., Zhao, F., Wu, W., Lyu, L., Li, W., & Zhang, C. (2023). Ellagic Acid from Hull Blackberries: Extraction, Purification, and Potential Anticancer Activity. International Journal of Molecular Sciences, 24(20), 15228. https://doi.org/10.3390/ijms242015228