
Long Non-Coding RNAs in Venous Thromboembolism: Where Do We Stand?
 , , ,
, , , 
Abstract
:
1. Introduction
2. LncRNAs: A Novel Epigenetic Regulator in VTE
2.1. ANRIL
2.2. GUSBP5-AS
2.3. MEG9
2.4. MALAT1
2.5. SIRT1-AS
2.6. LINC01123
2.7. TUG1
2.8. XIST
2.9. LINC00659 and UXT-AS1
2.10. CRNDE

{kind=link}
{kind=link}
{kind=link}
{kind=link}
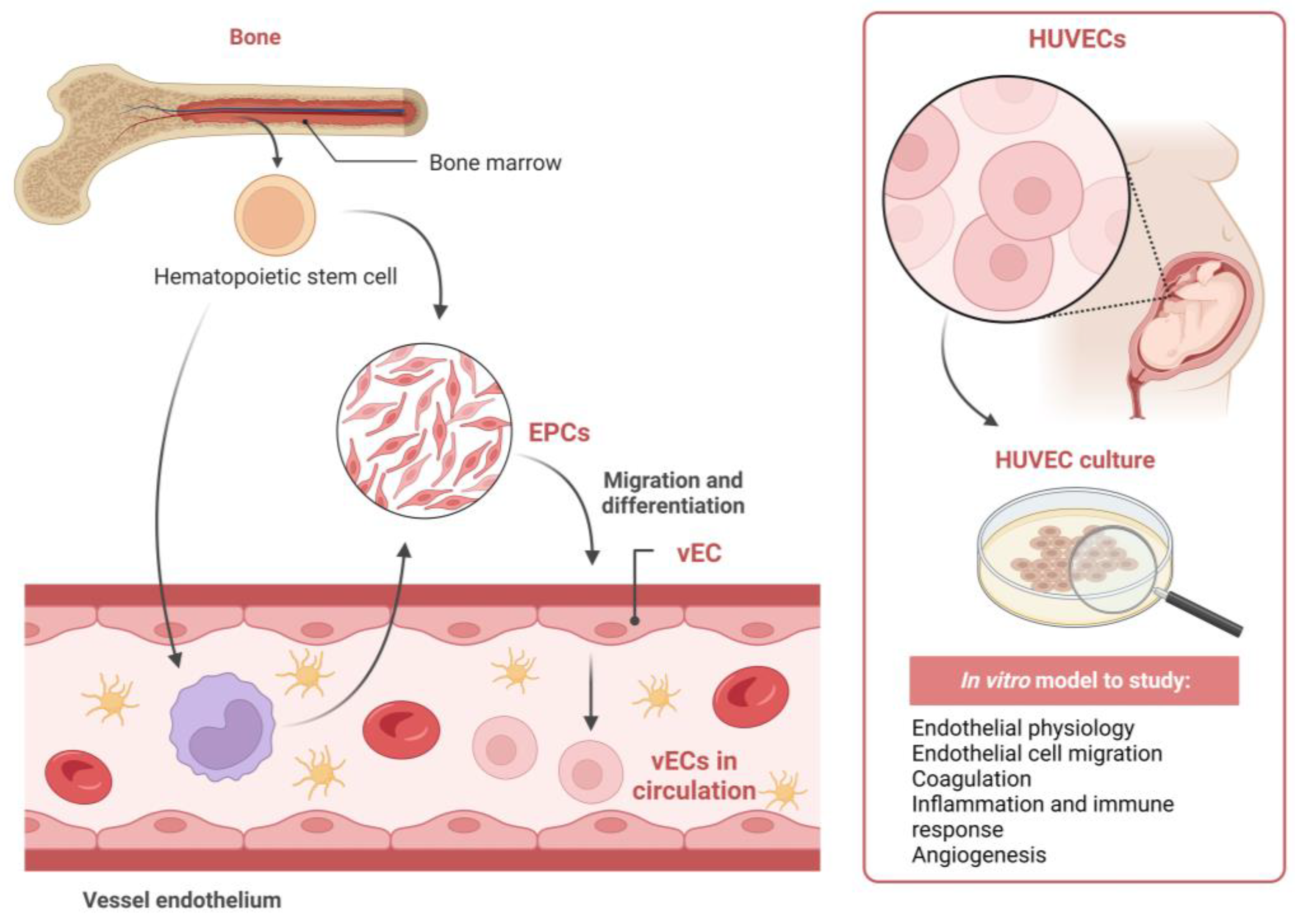{kind=link}
{kind=link}
| LncRNA | Study | LncRNA Location 1 | LncRNA Expression in VTE | Sample/Compartment or Study Model | LncRNA Target | Mechanism of Action | LncRNA Function in VTE |
|---|---|---|---|---|---|---|---|
| ANRIL | Zeng et al. (China, 2019) [67] | 9p21.3 | ↑ | HUVECs and rats | ↓miR-99a ↓miR-449a | Increases beclin-1 expression via miR-99a and miR-449a sponging and upregulates thrombomodulin | Promotes thrombosis |
| GUSBP5-AS | Sun et al. (China, 2020) [7] | 4q31.21 | ↑ | Human EPCs and mice | ↓miR-223-3p | Sponges miR-223-3p, which targets FOXO1 and activates the Akt pathway and enhances FGF2, MMP2/9 and F-actin expression | Promotes DVT resolution |
| MEG9 | Espinosa-Diez et al. (EUA, 2020) [73] | 14q32.31 | n.a | ECs | n.a | Inhibits fibrin formation | Possibly has a protective effect against thrombosis |
| MALAT1 | Du et al. (China, 2020) [8] | 11q13.1 | ↑ | Human EPCs | Wnt/β-catenin | Reduces endothelial function | Promotes DVT |
| Wang et al. (China, 2022) [9] | HUVECs | ↓miR-383-5p/BCL2L11 | |||||
| Sun et al. (China, 2022) [83] | CD34+ megakaryocytes and mice | PI3K/AKT/GSK-3β | Inhibits platelet activity and thrombus formation | Inhibits DVT | |||
| Sirt1-AS | Lou et al. (China, 2021) [39] | 10q21.3 | ↓ | Patients’ blood, mice and HUVECs | ↑Sirt1 | Suppresses the endothelial ageing and alleviates the thrombus formation through Sirt1/FOXO3a axis | Attenuates ageing-related DVT |
| LincRNA 1123 | Yang et al. (2022, China) [95] | 2q13 | ↑ | Rats | ↓miR-125a-3p | Sponges miR-125a-3p, which targets IL1R1, facilitating thrombus formation and inducing higher levels of IL-6 and IL-8 | Promotes LEDVT |
| TUG1 | Feng et al. (China, 2022) [58] | 22q12.2 | ↓ | Mice EPCs and mice | ↓miR-92a-3p | Accelerates proliferation, migration and tube-forming abilities and decreases apoptosis of EPCs and thrombus size through downregulation of miR-91a-3p and upregulation of HMGCR | Protects against DVT |
| XIST | Cao et al. (China, 2022) [102] | Xq13.2 | ↑ | Human plasma and HUVECs | ↓miR-103a-3p | Reduces ECs normal functions through miR-103a-3p sponging | Promotes DVT |
| LINC00659 | Zhang et al. (China, 2023) [121] | 20q13.33 | ↑ | Human IVC tissue samples, human EPCs and mice | ↑EIF4A3 | Upregulates EIF4A3 expression and exacerbates endothelial progenitor cell dysfunction | Promotes LEDVT |
| Zhu et al. (China, 2023) [120] | n.a | HUVECs | ↓miR-525-5p | Inhibits proliferation and cell viability in HUVECs through downregulation of miR-525-5p, which targets Bax | Possibly promotes DVT, given its role in HUVECs | ||
| CRNDE | He et al. (China, 2023) [129] | 16q12.2 | ↑ | Mice IVC | ↓miR-181a-5p | Competitively bound to and inhibits miR-181a-5p, promoting Pcyox1l expression and aggravating thrombus formation in DVT | Promotes DVT |
3. LncRNAs Targeting the Coagulation System
3.1. TFPI2AS1
3.2. Linc00473
3.3. AC003092.1
3.4. AGAP2-AS1
3.5. MEG8
| LncRNA | First Author (Country, Year) [Ref] | LncRNA Location 1 | Disease | Sample/Compartment or Study Model | LncRNA Expression in the Disease | LncRNA Targets | LncRNA Role in TFPI2 Expression |
|---|---|---|---|---|---|---|---|
| TFPI2AS1 | Gao et al. (China, 2017) [153] | 7q31-q32 | NSCLC | NSCLC tissue and cells | ↑ | TFPI2 | ↑ |
| Linc00473 | Wu et al. (China, 2018) [158] | 6q27 | Preeclampsia | Placenta tissues and trophoblast cell lines | ↓ | LSD1/TFPI2 | ↓ |
| AC003092.1 | Xu et al. (China, 2018) [160] | 7q21.3 | Glioblastoma | Glioblastoma tissue and cells and mice | ↓ | MiR-195/TFPI2 | ↑ |
| AGAP2-AS1 | Luo et al. (China, 2019) [164] | 12q14.1 | Glioblastoma | Glioblastoma tissue and cells and mice | ↑ | EZH2 and LSD1/TFPI2 | ↓ |
| MEG8 | Kremer et al. (2022, The Netherlands) [166] | 14q32.31 | Ischemic heart disease | Left ventricular tissues and HUVECs | ↓ | TFPI2 | ↑ |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naito, H.; Iba, T.; Takakura, N. Mechanisms of new blood-vessel formation and proliferative heterogeneity of endothelial cells. Int. Immunol. 2020, 32, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batty, P.; Smith, J.G. Haemostasis. Surgery 2010, 28, 530–535. [Google Scholar] [CrossRef]
- Tavares, V.; Pinto, R.; Assis, J.; Pereira, D.; Medeiros, R. Venous thromboembolism GWAS reported genetic makeup and the hallmarks of cancer: Linkage to ovarian tumour behaviour. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2020, 1873, 188331. [Google Scholar] [CrossRef]
- Zaidi, A.; Green, L. Physiology of Haemostasis. Anaesth. Intensive Care Med. 2019, 20, 152–158. [Google Scholar] [CrossRef]
- Kremers, R.; Peters, T.; Wagenvoord, R.; Hemker, H. The balance of pro-and anticoagulant processes underlying thrombin generation. J. Thromb. Haemost. 2015, 13, 437–447. [Google Scholar] [CrossRef]
- Neubauer, K.; Zieger, B. Endothelial cells and coagulation. Cell Tissue Res. 2022, 387, 391–398. [Google Scholar] [CrossRef]
- Sun, L.L.; Lei, F.R.; Jiang, X.D.; Du, X.L.; Xiao, L.; Li, W.D.; Li, X.Q. LncRNA GUSBP5-AS promotes EPC migration and angiogenesis and deep vein thrombosis resolution by regulating FGF2 and MMP2/9 through the miR-223-3p/FOXO1/Akt pathway. Aging 2020, 12, 4506–4526. [Google Scholar] [CrossRef]
- Du, B.; Wang, J.; Zang, S.; Mao, X.; Du, Y. Long non-coding RNA MALAT1 suppresses the proliferation and migration of endothelial progenitor cells in deep vein thrombosis by regulating the Wnt/β-catenin pathway. Exp. Ther. Med. 2020, 20, 3138–3146. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lin, S.; Yang, Y.; Zhao, M.; Li, X.; Zhang, L. Significant role of long non-coding RNA MALAT1 in deep vein thrombosis via the regulation of vascular endothelial cell physiology through the microRNA-383-5p/BCL2L11 axis. Bioengineered 2022, 13, 13728–13738. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Giménez, B.; Berastegui, C.; Loor, K.; López-Meseguer, M.; Monforte, V.; Bravo, C.; Santamaría, A.; Roman, A. Deep vein thrombosis and pulmonary embolism after solid organ transplantation: An unresolved problem. Transpl. Rev. 2015, 29, 85–92. [Google Scholar] [CrossRef]
- Khan, F.; Tritschler, T.; Kahn, S.R.; Rodger, M.A. Venous thromboembolism. Lancet 2021, 398, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Morelli, V.M.; Brækkan, S.K.; Hansen, J.B. Role of microRNAs in Venous Thromboembolism. Int. J. Mol. Sci. 2020, 21, 2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, A.-m.; Li, W.-D.; Kong, L.; Rong, J.-j.; Lei, F.-R.; Du, X.-l.; Li, C.-l.; Li, X.-Q. Long noncoding RNA expression profile of endothelial progenitor cells from deep vein thrombosis patients identified by microarray analysis. Int. J. Clin. Exp. Pathol. 2016, 9, 12275–12286. [Google Scholar]
- Wendelboe, A.M.; Raskob, G.E. Global Burden of Thrombosis. Circ. Res. 2016, 118, 1340–1347. [Google Scholar] [CrossRef]
- Engbers, M.J.; van Hylckama Vlieg, A.; Rosendaal, F.R. Venous thrombosis in the elderly: Incidence, risk factors and risk groups. J. Thromb. Haemost. 2010, 8, 2105–2112. [Google Scholar] [CrossRef] [PubMed]
- Kushner, A.; West, W.P.; Khan Suheb, M.Z.; Pillarisetty, L.S. Virchow Triad. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Wells, P.S.; Brill-Edwards, P.; Stevens, P.; Panju, A.; Patel, A.; Douketis, J.; Massicotte, M.P.; Hirsh, J.; Weitz, J.I.; Kearon, C.; et al. A Novel and Rapid Whole-Blood Assay for D-Dimer in Patients With Clinically Suspected Deep Vein Thrombosis. Circulation 1995, 91, 2184–2187. [Google Scholar] [CrossRef] [Green Version]
- Oi, M.; Yamashita, Y.; Toyofuku, M.; Morimoto, T.; Motohashi, Y.; Tamura, T.; Kaitani, K.; Amano, H.; Takase, T.; Hiramori, S.; et al. D-dimer levels at diagnosis and long-term clinical outcomes in venous thromboembolism: From the COMMAND VTE Registry. J. Thromb. Thrombolysis 2020, 49, 551–561. [Google Scholar] [CrossRef]
- Bates, S.M.; Jaeschke, R.; Stevens, S.M.; Goodacre, S.; Wells, P.S.; Stevenson, M.D.; Kearon, C.; Schunemann, H.J.; Crowther, M.; Pauker, S.G.; et al. Diagnosis of DVT: Antithrombotic Therapy and Prevention of Thrombosis, 9th ed: American College of Chest Physicians Evidence-Based Clinical Practice Guidelines. Chest 2012, 141 (Suppl. 2), e351S–e418S. [Google Scholar] [CrossRef] [Green Version]
- Stein, P.D.; Hull, R.D.; Patel, K.C.; Olson, R.E.; Ghali, W.A.; Brant, R.; Biel, R.K.; Bharadia, V.; Kalra, N.K. D-Dimer for the Exclusion of Acute Venous Thrombosis and Pulmonary Embolism. Ann. Intern. Med. 2004, 140, 589–602. [Google Scholar] [CrossRef]
- Mulder, F.I.; Horvàth-Puhó, E.; van Es, N.; van Laarhoven, H.; Pedersen, L.; Moik, F.; Ay, C.; Büller, H.; Sørensen, H.T. Venous thromboembolism in cancer patients: A population-based cohort study. Blood 2021, 137, 1959–1969. [Google Scholar] [CrossRef]
- Lim, W.; Le Gal, G.; Bates, S.M.; Righini, M.; Haramati, L.B.; Lang, E.; Kline, J.A.; Chasteen, S.; Snyder, M.; Patel, P. American Society of Hematology 2018 guidelines for management of venous thromboembolism: Diagnosis of venous thromboembolism. Blood Adv. 2018, 2, 3226–3256. [Google Scholar] [CrossRef] [Green Version]
- Hembrom, A.A.; Srivastava, S.; Garg, I.; Kumar, B. MicroRNAs in venous thrombo-embolism. Clin. Chim. Acta 2020, 504, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Grimnes, G.; Isaksen, T.; Tichelaar, Y.I.G.V.; Brox, J.; Brækkan, S.K.; Hansen, J.-B. C-reactive protein and risk of venous thromboembolism: Results from a population-based case-crossover study. Haematologica 2018, 103, 1245. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L. Linking Long Noncoding RNA Localization and Function. Trends Biochem. Sci. 2016, 41, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Weng, R.; Lu, C.; Liu, X.; Li, G.; Lan, Y.; Qiao, J.; Bai, M.; Wang, Z.; Guo, X.; Ye, D.; et al. Long Noncoding RNA-1604 Orchestrates Neural Differentiation through the miR-200c/ZEB Axis. Stem Cells 2018, 36, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Dias, T.R.; Santos, J.M.; da Costa, R.M.G.; Medeiros, R. Long non-coding RNAs regulate the hallmarks of cancer in HPV-induced malignancies. Crit. Rev. Oncol. Hematol. 2021, 161, 103310. [Google Scholar] [CrossRef]
- Tsagakis, I.; Douka, K.; Birds, I.; Aspden, J. Long non-coding RNAs in development and disease: Conservation to mechanisms: Long non-coding RNAs in development and disease. J. Pathol. 2020, 250, 480–495. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wang, J.; Li, Y.; Song, T.; Wu, Y.; Fang, S.; Bu, D.; Li, H.; Sun, L.; Pei, D. NONCODEV6: An updated database dedicated to long non-coding RNA annotation in both animals and plants. Nucleic Acids Res. 2021, 49, D165–D171. [Google Scholar] [CrossRef]
- Marques, A.C.; Ponting, C.P. Intergenic lncRNAs and the evolution of gene expression. Curr. Opin. Genet. Dev. 2014, 27, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Villegas, V.E.; Zaphiropoulos, P.G. Neighboring gene regulation by antisense long non-coding RNAs. Int. J. Mol. Sci. 2015, 16, 3251–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, C.A.G.; Adachi, S.; Nong, Q.D.; Adhitama, N.; Matsuura, T.; Natsume, T.; Wada, T.; Kato, Y.; Watanabe, H. Sense-overlapping lncRNA as a decoy of translational repressor protein for dimorphic gene expression. PLoS Genet. 2021, 17, e1009683. [Google Scholar] [CrossRef]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.-L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef]
- Cheng, J.-T.; Wang, L.; Wang, H.; Tang, F.-R.; Cai, W.-Q.; Sethi, G.; Xin, H.-W.; Ma, Z. Insights into biological role of LncRNAs in epithelial-mesenchymal transition. Cells 2019, 8, 1178. [Google Scholar] [CrossRef] [Green Version]
- Sanchez Calle, A.; Kawamura, Y.; Yamamoto, Y.; Takeshita, F.; Ochiya, T. Emerging roles of long non-coding RNA in cancer. Cancer Sci. 2018, 109, 2093–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, L.; Ding, Y.; Lu, X.; Zhang, G.; Yang, J.; Zheng, H.; Wang, H.; Jiang, Y.; Xu, L. LncRNA Structural Characteristics in Epigenetic Regulation. Int. J. Mol. Sci. 2017, 18, 2659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Chen, Y.; Guo, R.; Dai, Y.; Tang, L.; Zhao, Y.; Wu, X.; Li, M.; Du, F.; Shen, J. Interaction of ncRNA and epigenetic modifications in gastric cancer: Focus on histone modification. Front. Oncol. 2022, 11, 822745. [Google Scholar] [CrossRef]
- Lou, Z.; Zhu, J.; Li, X.; Li, X.; Du, K.; Wang, B.; Zhang, F.; Zhang, X. LncRNA Sirt1-AS upregulates Sirt1 to attenuate aging related deep venous thrombosis. Aging 2021, 13, 6918–6935. [Google Scholar] [CrossRef]
- Holdt, L.M.; Sass, K.; Gäbel, G.; Bergert, H.; Thiery, J.; Teupser, D. Expression of Chr9p21 genes CDKN2B (p15INK4b), CDKN2A (p16INK4a, p14ARF) and MTAP in human atherosclerotic plaque. Atherosclerosis 2011, 214, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Dominguez, J.R.; Lodish, H.F. Emerging mechanisms of long noncoding RNA function during normal and malignant hematopoiesis. Blood J. Am. Soc. Hematol. 2017, 130, 1965–1975. [Google Scholar] [CrossRef] [Green Version]
- Sallam, T.; Sandhu, J.; Tontonoz, P. Long noncoding RNA discovery in cardiovascular disease: Decoding form to function. Circ. Res. 2018, 122, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Sallam, T.; Jones, M.; Thomas, B.J.; Wu, X.; Gilliland, T.; Qian, K.; Eskin, A.; Casero, D.; Zhang, Z.; Sandhu, J. Transcriptional regulation of macrophage cholesterol efflux and atherogenesis by a long noncoding RNA. Nat. Med. 2018, 24, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Boulberdaa, M.; Scott, E.; Ballantyne, M.; Garcia, R.; Descamps, B.; Angelini, G.D.; Brittan, M.; Hunter, A.; McBride, M.; McClure, J. A role for the long noncoding RNA SENCR in commitment and function of endothelial cells. Mol. Ther. 2016, 24, 978–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vausort, M.; Wagner, D.R.; Devaux, Y. Long noncoding RNAs in patients with acute myocardial infarction. Circ. Res. 2014, 115, 668–677. [Google Scholar] [CrossRef]
- Ishii, N.; Ozaki, K.; Sato, H.; Mizuno, H.; Saito, S.; Takahashi, A.; Miyamoto, Y.; Ikegawa, S.; Kamatani, N.; Hori, M. Identification of a novel non-coding RNA, MIAT, that confers risk of myocardial infarction. J. Hum. Genet. 2006, 51, 1087–1099. [Google Scholar] [CrossRef] [Green Version]
- Helgadottir, A.; Thorleifsson, G.; Manolescu, A.; Gretarsdottir, S.; Blondal, T.; Jonasdottir, A.; Jonasdottir, A.; Sigurdsson, A.; Baker, A.; Palsson, A. A common variant on chromosome 9p21 affects the risk of myocardial infarction. Science 2007, 316, 1491–1493. [Google Scholar] [CrossRef]
- McPherson, R.; Pertsemlidis, A.; Kavaslar, N.; Stewart, A.; Roberts, R.; Cox, D.R.; Hinds, D.A.; Pennacchio, L.A.; Tybjaerg-Hansen, A.; Folsom, A.R. A common allele on chromosome 9 associated with coronary heart disease. Science 2007, 316, 1488–1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, S.; Dimmeler, S. Long noncoding RNAs in cardiovascular diseases. Circ. Res. 2015, 116, 737–750. [Google Scholar] [CrossRef]
- Jerjes-Sanchez, C. Venous and Arterial Thrombosis: A Continuous Spectrum of the Same Disease? Oxford University Press: Oxford, UK, 2005; Volume 26, pp. 3–4. [Google Scholar]
- Kwaan, H.C.; Samama, M.M. The Significance of Endothelial Heterogeneity in Thrombosis and Hemostasis. Semin. Thromb. Hemost. 2010, 36, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Li, W.D.; Li, X.Q. Endothelial progenitor cells accelerate the resolution of deep vein thrombosis. Vasc. Pharmacol. 2016, 83, 10–16. [Google Scholar] [CrossRef]
- Anastasiou, G.; Gialeraki, A.; Merkouri, E.; Politou, M.; Travlou, A. Thrombomodulin as a regulator of the anticoagulant pathway: Implication in the development of thrombosis. Blood Coagul. Fibrinolysis 2012, 23, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Henke, P. Endothelial cell–mediated venous thrombosis. Blood 2022, 140, 1459–1460. [Google Scholar] [CrossRef]
- Wu, M.D.K.K.; Thiagarajan, M.D.P. Role of Endothelium in Thrombosis And Hemostasis. Annu. Rev. Med. 1996, 47, 315–331. [Google Scholar] [CrossRef]
- Poredos, P.; Jezovnik, M.K. Endothelial Dysfunction and Venous Thrombosis. Angiology 2018, 69, 564–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yau, J.W.; Teoh, H.; Verma, S. Endothelial cell control of thrombosis. BMC Cardiovasc. Disord. 2015, 15, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Lei, B.; Zhang, H.; Niu, L.; Li, X.; Luo, X.; Zhang, F. Long noncoding RNA TUG1 induces angiogenesis of endothelial progenitor cells and dissolution of deep vein thrombosis. Thromb. J. 2022, 20, 54. [Google Scholar] [CrossRef] [PubMed]
- Real, C.; Caiado, F.; Dias, S. Endothelial progenitors in vascular repair and angiogenesis: How many are needed and what to do? Cardiovasc. Haematol. Disord. -Drug Targets (Former. Curr. Drug Targets-Cardiovasc. Hematol. Disord.) 2008, 8, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Abou-Saleh, H.; Yacoub, D.; Théorêt, J.-F.; Gillis, M.-A.; Neagoe, P.-E.; Labarthe, B.; Théroux, P.; Sirois, M.G.; Tabrizian, M.; Thorin, E.; et al. Endothelial Progenitor Cells Bind and Inhibit Platelet Function and Thrombus Formation. Circulation 2009, 120, 2230–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.-J.; Zhang, Y.; Georgescu, S.P.; Johnson, K.L.; Kong, D.; Galper, J.B. Human umbilical vein endothelial cells and human dermal microvascular endothelial cells offer new insights into the relationship between lipid metabolism and angiogenesis. Stem Cell Rev. 2006, 2, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Medina-Leyte, D.J.; Domínguez-Pérez, M.; Mercado, I.; Villarreal-Molina, M.T.; Jacobo-Albavera, L. Use of human umbilical vein endothelial cells (HUVEC) as a model to study cardiovascular disease: A review. Appl. Sci. 2020, 10, 938. [Google Scholar] [CrossRef] [Green Version]
- Pasmant, E.; Laurendeau, I.; Héron, D.; Vidaud, M.; Vidaud, D.; Bièche, I. Characterization of a germ-line deletion, including the entire INK4/ARF locus, in a melanoma-neural system tumor family: Identification of ANRIL, an antisense noncoding RNA whose expression coclusters with ARF. Cancer Res. 2007, 67, 3963–3969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Huang, X.; Hu, F.; Fu, X.; Jiang, Z.; Chen, K. LncRNA ANRIL knockdown relieves myocardial cell apoptosis in acute myocardial infarction by regulating IL-33/ST2. Cell Cycle 2019, 18, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Lou, N.; Liu, G.; Pan, Y. Long noncoding RNA ANRIL as a novel biomarker in human cancer. Future Oncol. 2020, 16, 2981–2995. [Google Scholar] [CrossRef] [PubMed]
- Razeghian-Jahromi, I.; Karimi Akhormeh, A.; Zibaeenezhad, M.J. The Role of ANRIL in Atherosclerosis. Dis. Markers 2022, 2022, 8859677. [Google Scholar] [CrossRef]
- Zeng, R.; Song, X.J.; Liu, C.W.; Ye, W. LncRNA ANRIL promotes angiogenesis and thrombosis by modulating microRNA-99a and microRNA-449a in the autophagy pathway. Am. J. Transl. Res. 2019, 11, 7441–7448. [Google Scholar] [PubMed]
- Okamoto, T.; Tanigami, H.; Suzuki, K.; Shimaoka, M. Thrombomodulin: A bifunctional modulator of inflammation and coagulation in sepsis. Crit. Care Res. Pract. 2012, 2012, 614545. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Qi, M.; Yang, Y.; Xu, P.; Zhua, Y.; Zhang, J. Circulating lncRNA ANRIL in the Serum of Patients with Ischemic Stroke. Clin. Lab. 2019, 65, 1459–1465. [Google Scholar] [CrossRef]
- Zhou, L.; Zheng, D.; Song, X.; Zhu, J.; Qi, W.; Ding, S.; Zhang, Y.; Xu, Q.; Han, X.; Zhao, Y. Alternated mRNA expression of the genes in chromosome 9p21 is associated with coronary heart disease and genetic variants in chromosome 9p21. Thromb. Res. 2019, 178, 17–19. [Google Scholar] [CrossRef]
- Espinosa-Diez, C.; Fraile-Bethencourt, E.; Hudson, C.; Ruhl, R.; Anand, S. Abstract 1423: A lncRNA MEG9 protects vasculature from DNA damage. Cancer Res. 2020, 80 (Suppl. 16), 1423. [Google Scholar] [CrossRef]
- Anand, S.; Espinosa Diez, M.; Chatterjee, N.; Ruhl, R. Dna Methylation of Lncrna Meg9 Inhibits Angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2019, 39 (Suppl. 1), A200. [Google Scholar]
- Espinosa-Diez, C.; Wilson, R.; Mukherjee, R.; Feltham, M.; Hudson, C.; Ruhl, R.; Anand, S. DNA damage dependent hypomethylation regulates the pro-angiogenic LncRNA MEG9. BioRxiv 2018, 442699. [Google Scholar] [CrossRef]
- Fraile-Bethencourt, E.; Khou, S.; Wilson, R.; Baris, A.; Ruhl, R.; Espinosa-Diez, C.; Anand, S. DNA damage-induced lncRNA MEG9 impacts angiogenesis. BioRxiv 2022. [Google Scholar] [CrossRef]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chi, Z.; Jin, H.; Yang, W. MicroRNA miR-188-5p as a mediator of long non-coding RNA MALAT1 regulates cell proliferation and apoptosis in multiple myeloma. Bioengineered 2021, 12, 1611–1626. [Google Scholar] [CrossRef]
- Chen, D.; Liu, L.; Wang, K.; Yu, H.; Wang, Y.; Liu, J.; Guo, Y.; Zhang, H. The role of MALAT-1 in the invasion and metastasis of gastric cancer. Scand. J. Gastroenterol. 2017, 52, 790–796. [Google Scholar] [CrossRef]
- Cheng, Y.; Imanirad, P.; Jutooru, I.; Hedrick, E.; Jin, U.H.; Rodrigues Hoffman, A.; Leal de Araujo, J.; Morpurgo, B.; Golovko, A.; Safe, S. Role of metastasis-associated lung adenocarcinoma transcript-1 (MALAT-1) in pancreatic cancer. PLoS ONE 2018, 13, e0192264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Wang, Y. Long noncoding RNA MALAT1 regulates apoptosis in ischemic stroke by sponging miR-205-3p and modulating PTEN expression. Am. J. Transl. Res. 2020, 12, 2738–2748. [Google Scholar]
- Kraus, T.F.J.; Haider, M.; Spanner, J.; Steinmaurer, M.; Dietinger, V.; Kretzschmar, H.A. Altered Long Noncoding RNA Expression Precedes the Course of Parkinson’s Disease-a Preliminary Report. Mol. Neurobiol. 2017, 54, 2869–2877. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Thomas, A.A.; Chen, S.; Aref-Eshghi, E.; Feng, B.; Gonder, J.; Sadikovic, B.; Chakrabarti, S. MALAT1: An epigenetic regulator of inflammation in diabetic retinopathy. Sci. Rep. 2018, 8, 6526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arslan, S.; Berkan, Ö.; Lalem, T.; Özbilüm, N.; Göksel, S.; Korkmaz, Ö.; Çetin, N.; Devaux, Y. Long non-coding RNAs in the atherosclerotic plaque. Atherosclerosis 2017, 266, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, T.; Lv, Y.; Li, J.; Jiang, X.; Jiang, J.; Zhang, D.; Bian, W.; Zhang, C. MALAT1 promotes platelet activity and thrombus formation through PI3k/Akt/GSK-3β signalling pathway. Stroke Vasc. Neurol. 2022, 8, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Pang, W.-J.; Wei, N.; Xiong, Y.; Wu, W.-J.; Zhao, C.-Z.; Shen, Q.-W.; Yang, G.-S. Identification, stability and expression of Sirt1 antisense long non-coding RNA. Gene 2014, 539, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.L.; Yin, W.G.; Huang, B.S.; Wu, L.X. LncRNA SNHG12 inhibits miR-199a to upregulate SIRT1 to attenuate cerebral ischemia/reperfusion injury through activating AMPK signaling pathway. Neurosci. Lett. 2019, 690, 188–195. [Google Scholar] [CrossRef]
- Qian, W.; Cai, X.; Qian, Q. Sirt1 antisense long non-coding RNA attenuates pulmonary fibrosis through sirt1-mediated epithelial-mesenchymal transition. Aging 2020, 12, 4322–4336. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.C.; Guarente, L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol. Metab. 2014, 25, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhou, M.; Ge, Y.; Wang, X. SIRT1 and aging related signaling pathways. Mech. Ageing Dev. 2020, 187, 111215. [Google Scholar] [CrossRef]
- Ding, X.H.; Han, J.; Liu, Y.; Jin, Y.; Ye, P. D-4F decreases the expression of Aβ protein through up-regulating long non coding RNA sirt1-as in SAMP8 mice. Saudi Pharm. J. 2017, 25, 517–522. [Google Scholar] [CrossRef]
- Li, B.; Hu, Y.; Li, X.; Jin, G.; Chen, X.; Chen, G.; Chen, Y.; Huang, S.; Liao, W.; Liao, Y.; et al. Sirt1 Antisense Long Noncoding RNA Promotes Cardiomyocyte Proliferation by Enhancing the Stability of Sirt1. J. Am. Heart Assoc. 2018, 7, e009700. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Liu, Y.; Zhao, J.; Li, L.; Hu, L.; Lu, Q.; Zeng, Z.; Liu, X.; Huang, D.; Yang, W.; et al. Long noncoding RNA LINC01123 promotes the proliferation and invasion of hepatocellular carcinoma cells by modulating the miR-34a-5p/TUFT1 axis. Int. J. Biol. Sci. 2020, 16, 2296–2305. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Zhou, H.; Liu, W.; Wang, J.; Zhang, Q. STAT1-induced upregulation of lncRNA LINC01123 predicts poor prognosis and promotes the progression of endometrial cancer through miR-516b/KIF4A. Cell Cycle 2020, 19, 1502–1516. [Google Scholar] [CrossRef]
- Ye, S.; Sun, B.; Wu, W.; Yu, C.; Tian, T.; Lian, Z.; Liang, Q.; Zhou, Y. LINC01123 facilitates proliferation, invasion and chemoresistance of colon cancer cells. Biosci. Rep. 2020, 40, BSR20194062. [Google Scholar] [CrossRef] [PubMed]
- Weng, G.; Gu, M.; Zhang, Y.; Zhao, G.; Gu, Y. LINC01123 promotes cell proliferation and migration via regulating miR-1277-5p/KLF5 axis in ox-LDL-induced vascular smooth muscle cells. J. Mol. Histol. 2021, 52, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhang, Z. Suppression of long intergenic non-protein coding RNA 1123 constrains lower extremity deep vein thrombosis via microRNA-125a-3p to target interleukin 1 receptor type 1. Bioengineered 2022, 13, 13452–13461. [Google Scholar] [CrossRef] [PubMed]
- Young, T.L.; Matsuda, T.; Cepko, C.L. The noncoding RNA taurine upregulated gene 1 is required for differentiation of the murine retina. Curr. Biol. 2005, 15, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Xue, M.; Xia, F.; Wang, Y.; Zhu, L.; Li, Y.; Jia, D.; Gao, Y.; Shi, Y.; Zhang, C.; He, Y.; et al. The Role of LncRNA TUG1 in Obesity-related Diseases. Mini-Rev. Med. Chem. 2022, 22, 1305–1313. [Google Scholar] [CrossRef]
- Azizidoost, S.; Nasrolahi, A.; Ghaedrahmati, F.; Kempisty, B.; Mozdziak, P.; Radoszkiewicz, K.; Farzaneh, M. The pathogenic roles of lncRNA-Taurine upregulated 1 (TUG1) in colorectal cancer. Cancer Cell Int. 2022, 22, 335. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.N.; Sattler, M. Challenges and perspectives for structural biology of lncRNAs—The example of the Xist lncRNA A-repeats. J. Mol. Cell Biol. 2019, 11, 845–859. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Gao, Y.; Liu, S. lncRNA XIST is associated with preeclampsia and mediates trophoblast cell invasion via miR-340-5p/KCNJ16 signaling pathway. Transpl. Immunol. 2022, 74, 101666. [Google Scholar] [CrossRef]
- Chen, Y.K.; Yen, Y. The Ambivalent Role of lncRNA Xist in Carcinogenesis. Stem Cell Rev. Rep. 2019, 15, 314–323. [Google Scholar] [CrossRef]
- Cao, G.; Zhou, H.; Wang, D.; Xu, L. Knockdown of lncRNA XIST Ameliorates IL-1β-Induced Apoptosis of HUVECs and Change of Tissue Factor Level via miR-103a-3p/HMGB1 Axis in Deep Venous Thrombosis by Regulating the ROS/NF-κB Signaling Pathway. Cardiovasc. Ther. 2022, 2022, 6256384. [Google Scholar] [CrossRef]
- Starikova, I.; Jamaly, S.; Sorrentino, A.; Blondal, T.; Latysheva, N.; Sovershaev, M.; Hansen, J.B. Differential expression of plasma miRNAs in patients with unprovoked venous thromboembolism and healthy control individuals. Thromb. Res. 2015, 136, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhao, Q.; Gong, K.; Long, Y.; Zhang, J.; Li, Y.; Guo, X. Downregulation of miR-103a-3p Contributes to Endothelial Progenitor Cell Dysfunction in Deep Vein Thrombosis Through PTEN Targeting. Ann. Vasc. Surg. 2020, 64, 339–346. [Google Scholar] [CrossRef]
- Sun, S.; Chai, S.; Zhang, F.; Lu, L. Overexpressed microRNA-103a-3p inhibits acute lower-extremity deep venous thrombosis via inhibition of CXCL12. IUBMB Life 2020, 72, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Dyer, M.R.; Chen, Q.; Haldeman, S.; Yazdani, H.; Hoffman, R.; Loughran, P.; Tsung, A.; Zuckerbraun, B.S.; Simmons, R.L.; Neal, M.D. Deep vein thrombosis in mice is regulated by platelet HMGB1 through release of neutrophil-extracellular traps and DNA. Sci. Rep. 2018, 8, 2068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golino, P.; Ragni, M.; Cirillo, P.; Avvedimento, V.E.; Feliciello, A.; Esposito, N.; Scognamiglio, A.; Trimarco, B.; Iaccarino, G.; Condorelli, M.; et al. Effects of tissue factor induced by oxygen free radicals on coronary flow during reperfusion. Nat. Med. 1996, 2, 35–40. [Google Scholar] [CrossRef]
- Guan, G.; Han, H.; Yang, Y.; Jin, Y.; Wang, X.; Liu, X. Neferine prevented hyperglycemia-induced endothelial cell apoptosis through suppressing ROS/Akt/NF-κB signal. Endocrine 2014, 47, 764–771. [Google Scholar] [CrossRef]
- Gutmann, C.; Siow, R.; Gwozdz, A.M.; Saha, P.; Smith, A. Reactive Oxygen Species in Venous Thrombosis. Int. J. Mol. Sci. 2020, 21, 1918. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.W.; Lo, Y.H.; Liu, H.; Yeh, C.Y.; Chen, Y.Z.; Hsu, C.W.; Chen, W.S.; Wang, J.H. Linc00659, a long noncoding RNA, acts as novel oncogene in regulating cancer cell growth in colorectal cancer. Mol. Cancer 2018, 17, 72. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Guo, Y.; Zhuang, T.; Xu, T.; Ji, M. SP1-Induced Upregulation of lncRNA LINC00659 Promotes Tumour Progression in Gastric Cancer by Regulating miR-370/AQP3 Axis. Front. Endocrinol. 2022, 13, 936037. [Google Scholar] [CrossRef]
- Fang, Y.; Wu, X.; Gu, Y.; Shi, R.; Yu, T.; Pan, Y.; Zhang, J.; Jing, X.; Ma, P.; Shu, Y. LINC00659 cooperated with ALKBH5 to accelerate gastric cancer progression by stabilising JAK1 mRNA in an m(6) A-YTHDF2-dependent manner. Clin. Transl. Med. 2023, 13, e1205. [Google Scholar] [CrossRef]
- Jha, P.K.; Sahu, A.; Prabhakar, A.; Tyagi, T.; Chatterjee, T.; Arvind, P.; Nair, J.; Gupta, N.; Kumari, B.; Nair, V.; et al. Genome-Wide Expression Analysis Suggests Hypoxia-Triggered Hyper-Coagulation Leading to Venous Thrombosis at High Altitude. Thromb. Haemost. 2018, 118, 1279–1295. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.K.; Vijay, A.; Prabhakar, A.; Chatterjee, T.; Nair, V.; Bajaj, N.; Kumar, B.; Sharma, M.; Ashraf, M.Z. Transcriptome Profiling Reveals the Endogenous Sponging Role of LINC00659 and UST-AS1 in High-Altitude Induced Thrombosis. Thromb. Haemost. 2021, 121, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, A.; Chatterjee, T.; Bajaj, N.; Tyagi, T.; Sahu, A.; Gupta, N.; Kumari, B.; Nair, V.; Kumar, B.; Ashraf, M.Z. Venous thrombosis at altitude presents with distinct biochemical profiles: A comparative study from the Himalayas to the plains. Blood Adv. 2019, 3, 3713–3723. [Google Scholar] [CrossRef]
- Gupta, N.; Sahu, A.; Prabhakar, A.; Chatterjee, T.; Tyagi, T.; Kumari, B.; Khan, N.; Nair, V.; Bajaj, N.; Sharma, M.; et al. Activation of NLRP3 inflammasome complex potentiates venous thrombosis in response to hypoxia. Proc. Natl. Acad. Sci. USA 2017, 114, 4763–4768. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Lohani, B.; Murphy, H. Pulmonary Embolism Masquerading as High Altitude Pulmonary Edema at High Altitude. High Alt. Med. Biol. 2016, 17, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Fricke-Galindo, I.; Buendia-Roldan, I.; Chavez-Galan, L.; Pérez-Rubio, G.; Hernández-Zenteno, R.d.J.; Ramos-Martinez, E.; Zazueta-Márquez, A.; Reyes-Melendres, F.; Alarcón-Dionet, A.; Guzmán-Vargas, J. SERPINE1 rs6092 Variant Is Related to Plasma Coagulation Proteins in Patients with Severe COVID-19 from a Tertiary Care Hospital. Biology 2022, 11, 595. [Google Scholar] [CrossRef]
- Evans, C.E.; Humphries, J.; Mattock, K.; Saha, P.; Smith, A. HIF1 signalling regulates venous thrombus resolution. Thromb. Res. 2012, 130, 971–973. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, B.; Xu, H. By modulating miR-525-5p/Bax axis, LINC00659 promotes vascular endothelial cell apoptosis. Immun. Inflamm. Dis. 2023, 11, e764. [Google Scholar] [CrossRef]
- Zhang, B.; Qin, J. LINC00659 exacerbates endothelial progenitor cell dysfunction in deep vein thrombosis of the lower extremities by activating DNMT3A-mediated FGF1 promoter methylation. Thromb. J. 2023, 21, 24. [Google Scholar] [CrossRef]
- Huang, L.; Wang, F.; Wang, Y.; Cao, Q.; Sang, T.; Liu, F.; Chen, S. Acidic Fibroblast Growth Factor Promotes Endothelial Progenitor Cells Function via Akt/FOXO3a Pathway. PLoS ONE 2015, 10, e0129665. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yang, B.; Chen, J.; Xiong, H.; Li, Y.; Pan, Z.; Cao, Y.; Chen, J.; Li, T.; Zhou, S.; et al. Upregulation of long non-coding RNA RAB1A-2 induces FGF1 expression worsening lung cancer prognosis. Cancer Lett. 2018, 438, 116–125. [Google Scholar] [CrossRef]
- Graham, L.D.; Pedersen, S.K.; Brown, G.S.; Ho, T.; Kassir, Z.; Moynihan, A.T.; Vizgoft, E.K.; Dunne, R.; Pimlott, L.; Young, G.P.; et al. Colorectal Neoplasia Differentially Expressed (CRNDE), a Novel Gene with Elevated Expression in Colorectal Adenomas and Adenocarcinomas. Genes Cancer 2011, 2, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Li, S.; Qin, X. Colorectal neoplasia differentially expressed: A long noncoding RNA with an imperative role in cancer. Onco Targets Ther. 2018, 11, 3755–3763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Ma, B.; Gao, Q.; Zhan, H.; Liu, Y.; Chen, Z.; Ye, S.; Li, J.; Yao, L.; Huang, W. Long non-coding RNA CRNDE in cancer prognosis: Review and meta-analysis. Clin. Chim. Acta 2018, 485, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Ren, L.; Liu, Y.; Liu, L. Long non-coding RNA CRNDE as potential biomarkers facilitate inflammation and apoptosis in alcoholic liver disease. Aging 2021, 13, 23233–23244. [Google Scholar] [CrossRef]
- Zhou, Y.; He, X.; Liu, R.; Qin, Y.; Wang, S.; Yao, X.; Li, C.; Hu, Z. LncRNA CRNDE regulates the proliferation and migration of vascular smooth muscle cells. J. Cell. Physiol. 2019, 234, 16205–16214. [Google Scholar] [CrossRef]
- He, X.; Liu, Y.; Li, Y.; Wu, K. Long non-coding RNA crnde promotes deep vein thrombosis by sequestering miR-181a-5p away from thrombogenic Pcyox1l. Thromb. J. 2023, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Marcos, L.V.; Lou-Bonafonte, J.M.; Martinez-Gracia, M.V.; Arnal, C.; Navarro, M.A.; Osada, J. Prenylcysteine oxidase 1, a pro-oxidant enzyme of low density lipoproteins. Front. Biosci. 2018, 23, 1020–1037. [Google Scholar] [CrossRef] [Green Version]
- Banfi, C.; Amadio, P.; Zarà, M.; Brioschi, M.; Sandrini, L.; Barbieri, S.S. Prenylcysteine Oxidase 1 (PCYOX1), a New Player in Thrombosis. Int. J. Mol. Sci. 2022, 23, 2831. [Google Scholar] [CrossRef]
- Nakamura, M.; Yamada, N.; Ito, M. Novel anticoagulant therapy of venous thromboembolism: Current status and future directions. Ann. Vasc. Dis. 2017, 10, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Bartholomew, J.R. Update on the management of venous thromboembolism. Cleve Clin. J. Med. 2017, 84 (Suppl. 3), 39–46. [Google Scholar] [CrossRef] [Green Version]
- Austin, S.K. Haemostasis. Medicine 2017, 45, 204–208. [Google Scholar] [CrossRef]
- Mackman, N.; Tilley, R.E.; Key, N.S. Role of the extrinsic pathway of blood coagulation in hemostasis and thrombosis. Arter. Thromb. Vasc. Biol. 2007, 27, 1687–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapin, J.C.; Hajjar, K.A. Fibrinolysis and the control of blood coagulation. Blood Rev. 2015, 29, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Grover, S.P.; Mackman, N. Intrinsic Pathway of Coagulation and Thrombosis. Arter. Thromb. Vasc. Biol. 2019, 39, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Al-Amer, O.M. The role of thrombin in haemostasis. Blood Coagul. Fibrinolysis 2022, 33, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Mast, A.E. Tissue factor pathway inhibitor: Multiple anticoagulant activities for a single protein. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.P.; Bunce, M.W.; Maroney, S.A.; Tracy, P.B.; Camire, R.M.; Mast, A.E. Tissue factor pathway inhibitor-alpha inhibits prothrombinase during the initiation of blood coagulation. Proc. Natl. Acad. Sci. USA 2013, 110, 17838–17843. [Google Scholar] [CrossRef]
- Chowdary, P. Inhibition of tissue factor pathway inhibitor (TFPI) as a treatment for haemophilia: Rationale with focus on concizumab. Drugs 2018, 78, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Vlodavsky, I.; Singh, P.; Boyango, I.; Gutter-Kapon, L.; Elkin, M.; Sanderson, R.D.; Ilan, N. Heparanase: From basic research to therapeutic applications in cancer and inflammation. Drug Resist. Updat. 2016, 29, 54–75. [Google Scholar] [CrossRef] [Green Version]
- Al-Mugeiren, M.M.; Abdel Gader, A.G.; Al-Rasheed, S.A.; Al-Salloum, A.A. Tissue factor pathway inhibitor in childhood nephrotic syndrome. Pediatr. Nephrol. 2006, 21, 771–777. [Google Scholar] [CrossRef]
- Kamikura, Y.; Wada, H.; Yamada, A.; Shimura, M.; Hiyoyama, K.; Shiku, H.; Tanigawa, M.; Nishikawa, H.; Yamada, N.; Isaka, N.; et al. Increased tissue factor pathway inhibitor in patients with acute myocardial infarction. Am. J. Hematol. 1997, 55, 183–187. [Google Scholar] [CrossRef]
- Leurs, P.B.; Stolk, R.P.; Hamulyak, K.; Van Oerle, R.; Grobbee, D.E.; Wolffenbuttel, B.H. Tissue factor pathway inhibitor and other endothelium-dependent hemostatic factors in elderly individuals with normal or impaired glucose tolerance and type 2 diabetes. Diabetes Care 2002, 25, 1340–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, N.; Lindahl, A.K.; Abildgaard, U. Elevated TFPI in malignant disease: Relation to cancer type and hypercoagulation. Br. J. Haematol. 1998, 102, 889–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iversen, N.; Lindahl, A.K.; Abildgaard, U. Elevated plasma levels of the factor Xa-TFPI complex in cancer patients. Thromb. Res. 2002, 105, 33–36. [Google Scholar] [CrossRef]
- Yamamuro, M.; Wada, H.; Kumeda, K.; Inoue, A.; Tsuji, I.; Nakasaki, T.; Shimura, M.; Hiyoyama, K.; Gabazza, E.C.; Nishikawa, M.; et al. Changes in plasma tissue factor pathway inhibitor levels during the clinical course of disseminated intravascular coagulation. Blood Coagul. Fibrinolysis 1998, 9, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Sprecher, C.A.; Kisiel, W.; Mathewes, S.; Foster, D.C. Molecular cloning, expression, and partial characterization of a second human tissue-factor-pathway inhibitor. Proc. Natl. Acad. Sci. USA 1994, 91, 3353–3357. [Google Scholar] [CrossRef] [PubMed]
- Lwaleed, B.A.; Bass, P.S. Tissue factor pathway inhibitor: Structure, biology and involvement in disease. J. Pathol. 2006, 208, 327–339. [Google Scholar] [CrossRef]
- Qiao, L.; Liang, N.; Zhang, J.; Xie, J.; Liu, F.; Xu, D.; Yu, X.; Tian, Y. Advanced research on vasculogenic mimicry in cancer. J. Cell. Mol. Med. 2015, 19, 315–326. [Google Scholar] [CrossRef]
- Chand, H.S.; Foster, D.C.; Kisiel, W. Structure, function and biology of tissue factor pathway inhibitor-2. Thromb. Haemost. 2005, 94, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Lin, Z.; Li, C.; Wang, Y.; Yang, L.; Zou, B.; Chen, J.; Li, J.; Song, Z.; Liu, G. TFPI2AS1, a novel lncRNA that inhibits cell proliferation and migration in lung cancer. Cell Cycle 2017, 16, 2249–2258. [Google Scholar] [CrossRef] [PubMed]
- Hassan, N.; Efing, J.; Kiesel, L.; Bendas, G.; Götte, M. The Tissue Factor Pathway in Cancer: Overview and Role of Heparan Sulfate Proteoglycans. Cancers 2023, 15, 1524. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Fu, W.; Zhang, L.; Fu, K.; Hu, J.; Jia, W.; Liu, G. LINC 00473 antagonizes the tumour suppressor miR-195 to mediate the pathogenesis of Wilms tumour via IKK α. Cell Prolif. 2018, 51, e12416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Song, Y. LINC00473 predicts poor prognosis and regulates cell migration and invasion in gastric cancer. Biomed. Pharmacother. 2018, 107, 1–6. [Google Scholar] [CrossRef]
- Vidaeff, A.C.; Saade, G.R.; Sibai, B.M. Preeclampsia: The need for a biological definition and diagnosis. Am. J. Perinatol. 2021, 38, 976–982. [Google Scholar] [CrossRef]
- Wu, D.; Xu, Y.; Zou, Y.; Zuo, Q.; Huang, S.; Wang, S.; Lu, X.; He, X.; Wang, J.; Wang, T.; et al. Long Noncoding RNA 00473 is Involved in Preeclampsia by LSD1 Binding-Regulated TFPI2 Transcription in Trophoblast Cells. Mol. Ther. Nucleic Acids 2018, 12, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Matsubara, S.; Yoshimoto, C.; Shigetomi, H.; Imanaka, S. Tissue Factor Pathway Inhibitors as Potential Targets for Understanding the Pathophysiology of Preeclampsia. Biomedicines 2023, 11, 1237. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Liu, B.; Lian, C.; Doycheva, D.M.; Fu, Z.; Liu, Y.; Zhou, J.; He, Z.; Yang, Z.; Huang, Q.; et al. Long noncoding RNA AC003092.1 promotes temozolomide chemosensitivity through miR-195/TFPI-2 signaling modulation in glioblastoma. Cell Death Dis. 2018, 9, 1139. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sun, M.; Zang, C.; Ma, P.; He, J.; Zhang, M.; Huang, Z.; Ding, Y.; Shu, Y. Upregulated long non-coding RNA AGAP2-AS1 represses LATS2 and KLF2 expression through interacting with EZH2 and LSD1 in non-small-cell lung cancer cells. Cell Death Dis. 2016, 7, e2225. [Google Scholar] [CrossRef]
- Hong, S.; Yan, Z.; Song, Y.; Bi, M.; Li, S. LncRNA AGAP2-AS1 augments cell viability and mobility, and confers gemcitabine resistance by inhibiting miR-497 in colorectal cancer. Aging 2020, 12, 5183–5194. [Google Scholar] [CrossRef]
- An, L.; Huang, J.; Ge, S.; Zhang, X.; Wang, J. lncRNA AGAP2-AS1 Facilitates Tumorigenesis and Ferroptosis Resistance through SLC7A11 by IGF2BP2 Pathway in Melanoma. Comput. Math. Methods Med. 2022, 2022, 1972516. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Li, X.; Song, Z.; Zhu, X.; Zhao, S. Long non-coding RNA AGAP2-AS1 exerts oncogenic properties in glioblastoma by epigenetically silencing TFPI2 through EZH2 and LSD1. Aging 2019, 11, 3811–3823. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Taheri, M.; Shojaei, S. A review on the role of MEG8 lncRNA in human disorders. Cancer Cell Int. 2022, 22, 285. [Google Scholar] [CrossRef] [PubMed]
- Kremer, V.; Bink, D.I.; Stanicek, L.; van Ingen, E.; Gimbel, T.; Hilderink, S.; Günther, S.; Nossent, A.Y.; Boon, R.A. MEG8 regulates Tissue Factor Pathway Inhibitor 2 (TFPI2) expression in the endothelium. Sci. Rep. 2022, 12, 843. [Google Scholar] [CrossRef]
- Volders, P.-J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. LNCipedia 5: Towards a reference set of human long non-coding RNAs. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef] [Green Version]
- Fathi Dizaji, B. Strategies to target long non-coding RNAs in cancer treatment: Progress and challenges. Egypt. J. Med. Hum. Genet. 2020, 21, 41. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—challenges and potential solutions. Nature Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Samad, A.F.A.; Kamaroddin, M.F. Innovative approaches in transforming microRNAs into therapeutic tools. WIREs RNA 2023, 14, e1768. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, I.S.; Tavares, V.; Neto, B.V.; Mota, I.N.R.; Pereira, D.; Medeiros, R. Long Non-Coding RNAs in Venous Thromboembolism: Where Do We Stand? Int. J. Mol. Sci. 2023, 24, 12103. https://doi.org/10.3390/ijms241512103
Marques IS, Tavares V, Neto BV, Mota INR, Pereira D, Medeiros R. Long Non-Coding RNAs in Venous Thromboembolism: Where Do We Stand? International Journal of Molecular Sciences. 2023; 24(15):12103. https://doi.org/10.3390/ijms241512103
Chicago/Turabian StyleMarques, Inês Soares, Valéria Tavares, Beatriz Vieira Neto, Inês N. R. Mota, Deolinda Pereira, and Rui Medeiros. 2023. "Long Non-Coding RNAs in Venous Thromboembolism: Where Do We Stand?" International Journal of Molecular Sciences 24, no. 15: 12103. https://doi.org/10.3390/ijms241512103
APA StyleMarques, I. S., Tavares, V., Neto, B. V., Mota, I. N. R., Pereira, D., & Medeiros, R. (2023). Long Non-Coding RNAs in Venous Thromboembolism: Where Do We Stand? International Journal of Molecular Sciences, 24(15), 12103. https://doi.org/10.3390/ijms241512103