
Distinct Clades of Protein Phosphatase 2A Regulatory B’/B56 Subunits Engage in Different Physiological Processes

Abstract
:1. Introduction
2. B’ Structure and Recognition of Slim Motifs

{kind=link}
{kind=link}
| Higher Plant B’ Clade | Each Clade Member Greek Letter/Number | AGI Locus Identifier and Protein UNIProtKB | Relative Expression in Seedlings% | Relative Expression in Rosette % | Relative Expression in Pollen % | Phosphorylation Predicted Number of Ser/Thr in Hot Spots | Matre 2009 Waadt 2015 Wang 2016 Main Localization (See Also Text) | SUBA Consensus Maximum Localization |
|---|---|---|---|---|---|---|---|---|
| α, β, ε/3,4,7 | B’α/B3 | At5g03470 O04375 | 21 | 52 | 20 | 2 | nucleus | nucleus 1.0 |
| B’β/B4 | At3g09880 O04376 | 42 | 45 | 15 | 7 | nucleus | nucleus 1.0 | |
| B’ε/B7 | At3g54930 Q9SV41 | 9 | 18 | 73 | 4 | cytosol nucleus | nucleus 0.76 | |
| γ, ζ/5,8 | B’γ/B5 | At4g15415 Q8RW96 | 39 | 37 | 16 | 0 | cytosol nucleus | cytosol 0.958 |
| B’ζ/B8 | At3g21650 Q9LVE2 | 21 | 65 | 6 | 11 | cytosol mitochondrion nuclear | mitochondrion 0.728 | |
| δ, η, θ/6,9,10 | B’δ/B6 | At3G26030 Q9ZQY6 | 100 | 100 | 0.3 | 0 | cytosol phragmoplast | golgi 1.000 |
| B’η/B9 | At3g26020 | 36 | 32 | 43 | 15 | cytosol nucleus | nucleus 1.000 | |
| B’θ/B10 | At1g13460 | 36 | 10 | 14 | 11 | cytosol peroxisome nuclear | nucleus 0.496 golgi 0.406 | |
| κ/11 | B’κ/B11 | At5g25510 Q93YV6 | 70 | 76 | 100 | 11 | cytosol nuclear | plastid 0.760 nucleus cytosol |
| ϕ | B’ϕ | not in Arabidopsis | - | - | - |
3. Expression Levels and Subcellular Localization Enable Diverse Functions of B’ Subunits
4. PP2A Is Involved in Various Hormone Signaling Pathways
4.1. Auxin and Ethylene
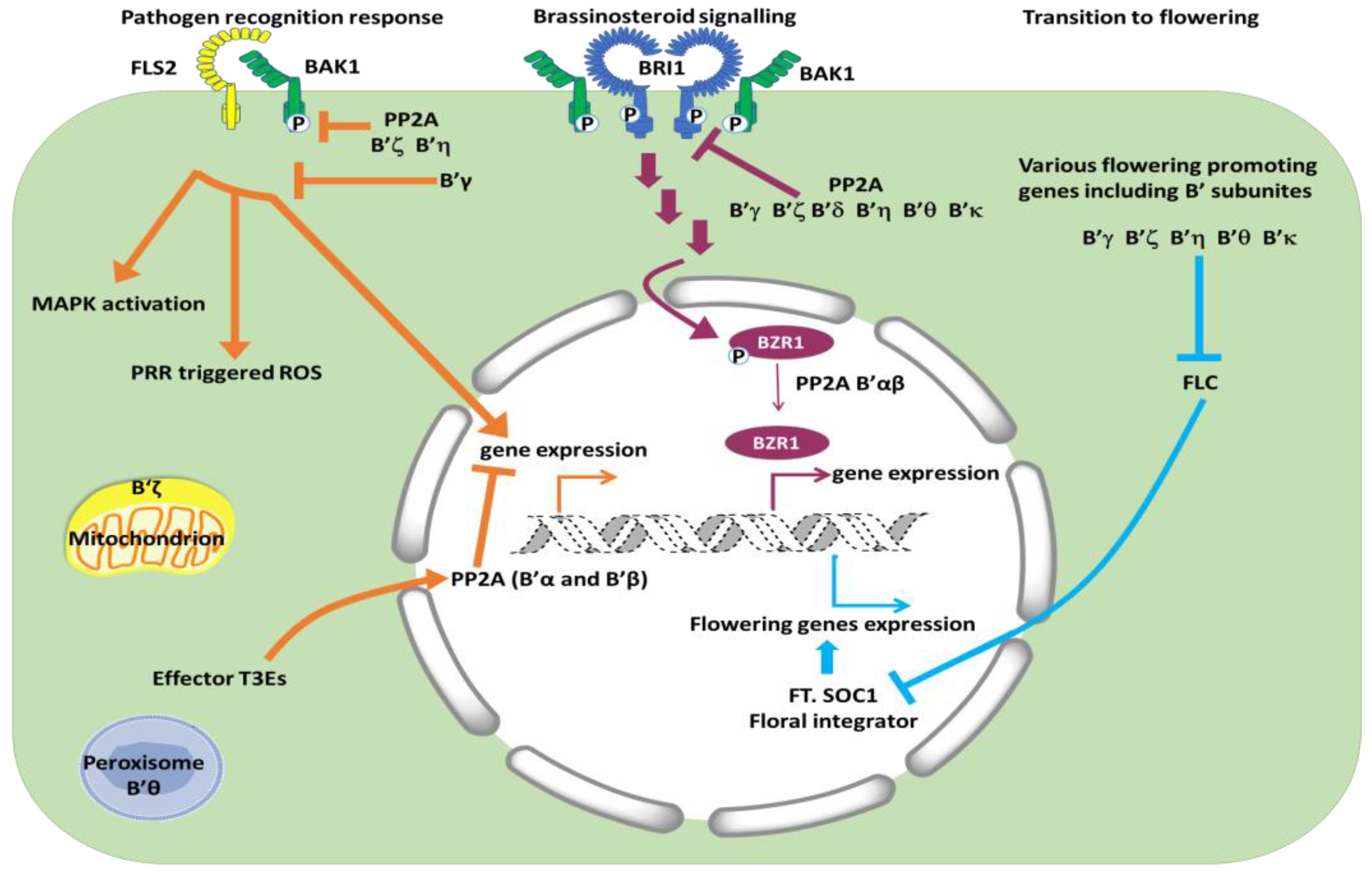
4.2. Brassinosteroid Signaling Is Regulated Positively by B’α and B’β and Negatively by Other B’ Subunits
4.3. Salicylic Acid Signaling—Involvement of B’γ
5. Interactions with Microbes
6. Transition to Reproductive Phase—Five b’ Single Mutants (γ, ζ, η, θ, κ) Are Late Flowering
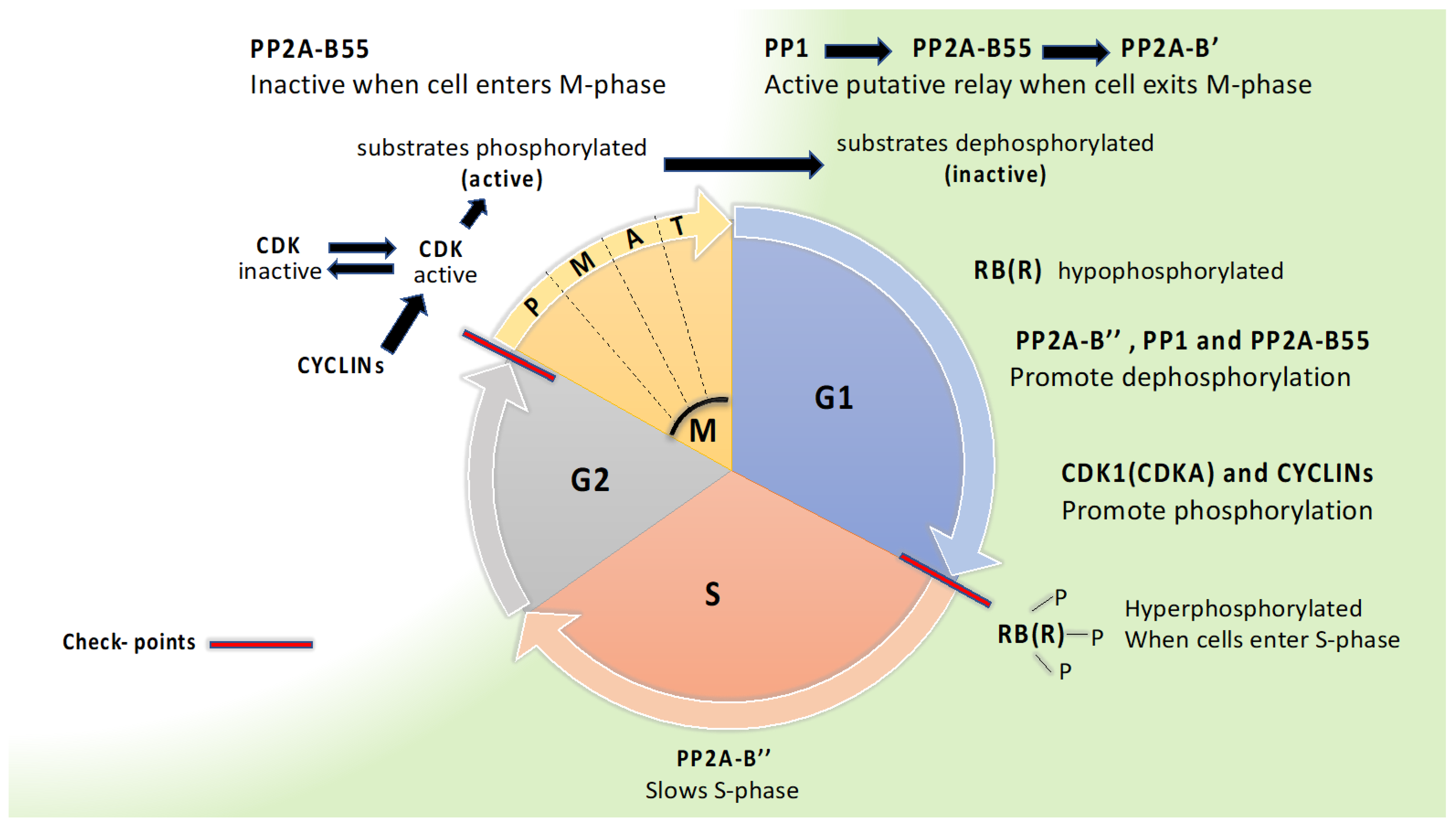
7. Cell Division
7.1. B’α and B’β Bind Shugoshin and Are Needed for Proper Meiosis in Plants
7.2. A Dephosphorylation Relay with PP1, PP2A-B55, and PP2A-B56 Controlling Mitosis Exit May Be Conserved in Plants
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mergner, J.; Frejno, M.; List, M.; Papacek, M.; Chen, X.; Chaudhary, A.; Samaras, P.; Richter, S.; Shikata, H.; Messerer, M. Mass-spectrometry-based draft of the Arabidopsis proteome. Nature 2020, 579, 409–414. [Google Scholar] [CrossRef]
- Kerk, D.; Templeton, G.; Moorhead, G.B.G. Evolutionary radiation pattern of novel protein phosphatases revealed by analysis of protein data from the completely sequenced genomes of humans, green algae, and higher plants. Plant Physiol. 2008, 146, 351. [Google Scholar] [CrossRef]
- Lillo, C.; Kataya, A.R.A.; Heidari, B.; Creighton, M.T.; Nemie-Feyissa, D.; Ginbot, Z.; Jonassen, E.M. Protein phosphatases PP 2A, PP 4 and PP 6: Mediators and regulators in development and responses to environmental cues. Plant Cell Environ. 2014, 37, 2631–2648. [Google Scholar] [CrossRef]
- Wlodarchak, N.; Xing, Y. PP2A as a master regulator of the cell cycle. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 162–184. [Google Scholar] [CrossRef]
- Uhrig, R.G.; Labandera, A.-M.; Moorhead, G.B. Arabidopsis PPP family of serine/threonine protein phosphatases: Many targets but few engines. Trends Plant Sci. 2013, 18, 505–513. [Google Scholar] [CrossRef]
- Brautigan, D.L.; Shenolikar, S. Protein serine/threonine phosphatases: Keys to unlocking regulators and substrates. Annu. Rev. Biochem. 2018, 87, 921–964. [Google Scholar] [CrossRef]
- Bheri, M.; Pandey, G.K. PP2A phosphatases take a giant leap in the post-genomics era. Curr. Genom. 2019, 20, 154. [Google Scholar] [CrossRef]
- Mazhar, S.; Taylor, S.E.; Sangodkar, J.; Narla, G. Targeting PP2A in cancer: Combination therapies. Biochim. Biophys. Acta BBA Mol. Cell Res. 2019, 1866, 51–63. [Google Scholar] [CrossRef]
- Booker, M.A.; DeLong, A. Atypical protein phosphatase 2A gene families do not expand via Paleopolyploidization. Plant Physiol. 2017, 173, 1283–1300. [Google Scholar] [CrossRef]
- Bheri, M.; Mahiwal, S.; Sanyal, S.K.; Pandey, G.K. Plant protein phosphatases: What do we know about their mechanism of action? FEBS J. 2021, 288, 756–785. [Google Scholar] [CrossRef]
- Máthé, C.; Freytag, C.; Kelemen, A.; M-Hamvas, M.; Garda, T. “B” Regulatory Subunits of PP2A: Their Roles in Plant Development and Stress Reactions. Int. J. Mol. Sci. 2023, 24, 5147. [Google Scholar] [CrossRef]
- Hertz, E.P.T.; Kruse, T.; Davey, N.E.; Lopez-Mendez, B.; Sigurðsson, J.O.; Montoya, G.; Olsen, J.V.; Nilsson, J. A conserved motif provides binding specificity to the PP2A-B56 phosphatase. Mol. Cell 2016, 63, 686–695. [Google Scholar] [CrossRef]
- Kruse, T.; Gnosa, S.P.; Nasa, I.; Garvanska, D.H.; Hein, J.B.; Nguyen, H.; Samsøe-Petersen, J.; Lopez-Mendez, B.; Hertz, E.P.T.; Schwarz, J. Mechanisms of site-specific dephosphorylation and kinase opposition imposed by PP2A regulatory subunits. EMBO J. 2020, 39, e103695. [Google Scholar] [CrossRef]
- Charpentier, M.; Sun, J.; Wen, J.; Mysore, K.S.; Oldroyd, G.E.D. Abscisic acid promotion of arbuscular mycorrhizal colonization requires a component of the PROTEIN PHOSPHATASE 2A complex. Plant Physiol. 2014, 166, 2077–2090. [Google Scholar] [CrossRef]
- Averkina, I.O.; Harris, M.; Asare, E.O.; Hourdin, B.; Paponov, I.A.; Lillo, C. Pinpointing regulatory protein phosphatase 2A subunits involved in beneficial symbiosis between plants and microbes. BMC Plant Biol. 2021, 21, 183. [Google Scholar] [CrossRef]
- Davey, N.E.; Cyert, M.S.; Moses, A.M. Short linear motifs—Ex nihilo evolution of protein regulation. Cell Commun. Signal. 2015, 13, 43. [Google Scholar] [CrossRef]
- Ueki, Y.; Hadders, M.A.; Weisser, M.B.; Nasa, I.; Sotelo-Parrilla, P.; Cressey, L.E.; Gupta, T.; Hertz, E.P.T.; Kruse, T.; Montoya, G. A highly conserved pocket on PP2A-B56 is required for hSgo1 binding and cohesion protection during mitosis. EMBO Rep. 2021, 22, e52295. [Google Scholar] [CrossRef]
- Tang, W.; Yuan, M.; Wang, R.; Yang, Y.; Wang, C.; Oses-Prieto, J.A.; Kim, T.-W.; Zhou, H.-W.; Deng, Z.; Gampala, S.S. PP2A activates brassinosteroid-responsive gene expression and plant growth by dephosphorylating BZR1. Nat. Cell Biol. 2011, 13, 124–131. [Google Scholar] [CrossRef]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef]
- Farkas, I.; Dombradi, V.; Miskei, M.; Szabados, L.; Koncz, C. Arabidopsis PPP family of serine/threonine phosphatases. Trends Plant Sci. 2007, 12, 169–176. [Google Scholar] [CrossRef]
- Creighton, M.T.; Kolton, A.; Kataya, A.R.A.; Maple-Grødem, J.; Averkina, I.O.; Heidari, B.; Lillo, C. Methylation of protein phosphatase 2A—Influence of regulators and environmental stress factors. Plant Cell Environ. 2017, 40, 2347–2358. [Google Scholar] [CrossRef] [PubMed]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Durek, P.; Schmidt, R.; Heazlewood, J.L.; Jones, A.; MacLean, D.; Nagel, A.; Kersten, B.; Schulze, W.X. PhosPhAt: The Arabidopsis thaliana phosphorylation site database. An update. Nucleic Acids Res. 2010, 38, D828–D834. [Google Scholar] [CrossRef]
- Matre, P.; Meyer, C.; Lillo, C. Diversity in subcellular targeting of the PP2A B’ η subfamily members. Planta 2009, 230, 935–945. [Google Scholar] [CrossRef]
- Waadt, R.; Manalansan, B.; Rauniyar, N.; Munemasa, S.; Booker, M.A.; Brandt, B.; Waadt, C.; Nusinow, D.A.; Kay, S.A.; Kunz, H.-H. Identification of OST1-interacting proteins reveals interactions with SnRK2-type protein kinases and with PP2A-type protein phosphatases that function in ABA responses. Plant Physiol. 2015, 169, 760–779. [Google Scholar] [CrossRef]
- Wang, R.; Liu, M.; Yuan, M.; Oses-Prieto, J.A.; Cai, X.; Sun, Y.; Burlingame, A.L.; Wang, Z.-Y.; Tang, W. The brassinosteroid-activated BRI1 receptor kinase is switched off by dephosphorylation mediated by cytoplasm-localized PP2A B’ subunits. Mol. Plant 2016, 9, 148–157. [Google Scholar] [CrossRef]
- Hooper, C.M.; Castleden, I.R.; Tanz, S.K.; Aryamanesh, N.; Millar, A.H. SUBA4: The interactive data analysis centre for Arabidopsis subcellular protein locations. Nucleic Acids Res. 2017, 45, D1064–D1074. [Google Scholar] [CrossRef]
- Grallert, A.; Boke, E.; Hagting, A.; Hodgson, B.; Connolly, Y.; Griffiths, J.R.; Smith, D.L.; Pines, J.; Hagan, I.M. A PP1–PP2A phosphatase relay controls mitotic progression. Nature 2015, 517, 94–98. [Google Scholar] [CrossRef]
- Lambrecht, C.; Haesen, D.; Sents, W.; Ivanova, E.; Janssens, V. Structure, regulation, and pharmacological modulation of PP2A phosphatases. In Phosphatase Modulators; Springer: Berlin/Heidelberg, Germany, 2013; pp. 283–305. [Google Scholar]
- Shao, Z.; Zhao, B.; Kotla, P.; Tran, J.; Chen, X.; Browning, K.; Qiao, H. Phosphorylation Status of B beta Subunit Acts as a Switch to Regulate the Function of Phosphatase PP2A in Ethylene-mediated Root Growth Inhibition. New Phytol. 2022, 236, 1762–1778. [Google Scholar] [CrossRef]
- Carroll, A.J.; Heazlewood, J.L.; Ito, J.; Millar, A.H. Analysis of the Arabidopsis cytosolic ribosome proteome provides detailed insights into its components and their post-translational modification. Mol. Cell. Proteom. 2008, 7, 347–369. [Google Scholar] [CrossRef]
- Yuan, G.; Ahootapeh, B.H.; Komaki, S.; Schnittger, A.; Lillo, C.; De Storme, N.; Geelen, D. PROTEIN PHOSHATASE 2A B’α and β maintain centromeric sister chromatid cohesion during meiosis in Arabidopsis. Plant Physiol. 2018, 178, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Trotta, A.; Wrzaczek, M.; Scharte, J.; Tikkanen, M.; Konert, G.; Rahikainen, M.; Holmström, M.; Hiltunen, H.-M.; Rips, S.; Sipari, N. Regulatory subunit B’ γ of protein phosphatase 2A prevents unnecessary defense reactions under low light in Arabidopsis. Plant Physiol. 2011, 156, 1464–1480. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, O.A.; Tomlinson, M.L.; Leader, D.; Shaw, P.; Doonan, J.H. High-throughput protein localization in Arabidopsis using Agrobacterium-mediated transient expression of GFP-ORF fusions. Plant J. 2005, 41, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Kataya, A.R.A.; Heidari, B.; Hagen, L.; Kommedal, R.; Slupphaug, G.; Lillo, C. Protein phosphatase 2A holoenzyme is targeted to peroxisomes by piggybacking and positively affects peroxisomal β-oxidation. Plant Physiol. 2015, 167, 493–506. [Google Scholar] [CrossRef]
- Kataya, A.; Mitchell, S.; Etman, R.; Samuel, M.; Moorhead, G.B. Peroxisomal protein phosphatase PP2A-B′theta interacts with and piggybacks SINA-like 10 E3 ligase into peroxisomes. Biochem. Biophys. Res. Commun. 2022, 644, 34–39. [Google Scholar] [CrossRef]
- MacKintosh, C.; Coggins, J.; Cohen, P. Plant protein phosphatases. Subcellular distribution, detection of protein phosphatase 2C and identification of protein phosphatase 2A as the major quinate dehydrogenase phosphatase. Biochem. J. 1991, 273, 733–738. [Google Scholar] [CrossRef]
- Blakeslee, J.J.; Zhou, H.-W.; Heath, J.T.; Skottke, K.R.; Barrios, J.A.R.; Liu, S.-Y.; DeLong, A. Specificity of RCN1-mediated protein phosphatase 2A regulation in meristem organization and stress response in roots. Plant Physiol. 2008, 146, 539–553. [Google Scholar] [CrossRef]
- White-Gloria, C.; Johnson, J.J.; Marritt, K.; Kataya, A.; Vahab, A.; Moorhead, G.B. Protein kinases and phosphatases of the plastid and their potential role in starch metabolism. Front. Plant Sci. 2018, 9, 1032. [Google Scholar] [CrossRef]
- Uhrig, R.G.; Moorhead, G.B. Two ancient bacterial-like PPP family phosphatases from Arabidopsis are highly conserved plant proteins that possess unique properties. Plant Physiol. 2011, 157, 1778–1792. [Google Scholar] [CrossRef]
- Zhang, Y.; Giese, J.; Kerbler, S.M.; Siemiatkowska, B.; de Souza, L.P.; Alpers, J.; Medeiros, D.B.; Hincha, D.K.; Daloso, D.M.; Stitt, M. Two mitochondrial phosphatases, PP2c63 and Sal2, are required for posttranslational regulation of the TCA cycle in Arabidopsis. Mol. Plant 2021, 14, 1104–1118. [Google Scholar] [CrossRef]
- Lucero, M.; Suarez, A.E.; Chambers, J.W. Phosphoregulation on mitochondria: Integration of cell and organelle responses. CNS Neurosci. Ther. 2019, 25, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Law, Y.-S.; Ngan, L.; Yan, J.; Kwok, L.Y.; Sun, Y.; Cheng, S.; Schwenkert, S.; Lim, B.L. Multiple kinases can phosphorylate the N-terminal sequences of mitochondrial proteins in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 982. [Google Scholar] [CrossRef] [PubMed]
- Garbers, C.; DeLong, A.; Deruere, J.; Bernasconi, P.; Söll, D. A mutation in protein phosphatase 2A regulatory subunit A affects auxin transport in Arabidopsis. EMBO J. 1996, 15, 2115–2124. [Google Scholar] [CrossRef] [PubMed]
- Michniewicz, M.; Zago, M.K.; Abas, L.; Weijers, D.; Schweighofer, A.; Meskiene, I.; Heisler, M.G.; Ohno, C.; Zhang, J.; Huang, F. Antagonistic regulation of PIN phosphorylation by PP2A and PINOID directs auxin flux. Cell 2007, 130, 1044–1056. [Google Scholar] [CrossRef] [PubMed]
- Máthé, C.; M-Hamvas, M.; Freytag, C.; Garda, T. The protein phosphatase PP2A plays multiple roles in plant development by regulation of vesicle traffic—Facts and questions. Int. J. Mol. Sci. 2021, 22, 975. [Google Scholar] [CrossRef]
- Ballesteros, I.; Domínguez, T.; Sauer, M.; Paredes, P.; Duprat, A.; Rojo, E.; Sanmartín, M.; Sánchez-Serrano, J.J. Specialized functions of the PP 2A subfamily II catalytic subunits PP 2A-C3 and PP 2A-C4 in the distribution of auxin fluxes and development in A rabidopsis. Plant J. 2013, 73, 862–872. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, G.; Wijewardene, I.; Cai, Y.; Esmaeili, N.; Sun, L.; Zhang, H. The B’ζ subunit of protein phosphatase 2A negatively regulates ethylene signaling in Arabidopsis. Plant Physiol. Biochem. 2021, 169, 81–91. [Google Scholar] [CrossRef]
- Jonassen, E.M.; Heidari, B.; Nemie-Feyissa, D.; Matre, P.; Lillo, C. Protein phosphatase 2A regulatory subunits are starting to reveal their functions in plant metabolism and development. Plant Signal. Behav. 2011, 6, 1216–1218. [Google Scholar] [CrossRef]
- Zhang, Y.-L.; Zhang, H.; Gao, Y.-J.; Yan, L.-L.; Yu, X.-Y.; Yang, Y.-H.; Xu, W.-Y.; Pu, C.-X.; Sun, Y. Protein Phosphatase 2A B’α and B’β Protect Centromeric Cohesion during Meiosis I. Plant Physiol. 2019, 179, 1556–1568. [Google Scholar] [CrossRef]
- Zhou, H.; Ge, H.; Chen, J.; Li, X.; Yang, L.; Zhang, H.; Wang, Y. Salicylic Acid Regulates Root Gravitropic Growth via Clathrin-Independent Endocytic Trafficking of PIN2 Auxin Transporter in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 9379. [Google Scholar] [CrossRef]
- Tan, S.; Abas, M.; Verstraeten, I.; Glanc, M.; Molnár, G.; Hajný, J.; Lasák, P.; Petřík, I.; Russinova, E.; Petrášek, J. Salicylic acid targets protein phosphatase 2A to attenuate growth in plants. Curr. Biol. 2020, 30, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Durian, G.; Jeschke, V.; Rahikainen, M.; Vuorinen, K.; Gollan, P.J.; Brosché, M.; Salojärvi, J.; Glawischnig, E.; Winter, Z.; Li, S. PROTEIN PHOSPHATASE 2A-B′ γ controls Botrytis cinerea resistance and developmental leaf senescence. Plant Physiol. 2020, 182, 1161–1181. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Anderson, J.C.; Pozo, O.d.; Gu, Y.Q.; Tang, X.; Martin, G.B. Silencing of subfamily I of protein phosphatase 2A catalytic subunits results in activation of plant defense responses and localized cell death. Plant J. 2004, 38, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, Y.; Su, Z.; Lv, L.; Zhang, Z. Silencing of the wheat protein phosphatase 2A catalytic subunit TaPP2Ac enhances host resistance to the necrotrophic pathogen Rhizoctonia cerealis. Front. Plant Sci. 2018, 9, 1437. [Google Scholar] [CrossRef]
- Segonzac, C.; Macho, A.P.; Sanmartín, M.; Ntoukakis, V.; Sánchez-Serrano, J.J.; Zipfel, C. Negative control of BAK 1 by protein phosphatase 2A during plant innate immunity. EMBO J. 2014, 33, 2069–2079. [Google Scholar] [CrossRef]
- Erickson, J.; Weckwerth, P.; Romeis, T.; Lee, J. What’s new in protein kinase/phosphatase signalling in the control of plant immunity? Essays Biochem. 2022, 66, 621–634. [Google Scholar]
- Kataya, A.R.A.; Heidari, B.; Lillo, C. Protein phosphatase 2A regulatory subunits affecting plant innate immunity, energy metabolism, and flowering time–joint functions among B’η subfamily members. Plant Signal. Behav. 2015, 10, e1026024. [Google Scholar] [CrossRef]
- Kang, Y.; Barry, K.; Cao, F.; Zhou, M. Genome-wide association mapping for adult resistance to powdery mildew in common wheat. Mol. Biol. Rep. 2020, 47, 1241–1256. [Google Scholar] [CrossRef]
- Jin, L.; Ham, J.H.; Hage, R.; Zhao, W.; Soto-Hernández, J.; Lee, S.Y.; Paek, S.-M.; Kim, M.G.; Boone, C.; Coplin, D.L. Direct and indirect targeting of PP2A by conserved bacterial type-III effector proteins. PLoS Pathog. 2016, 12, e1005609. [Google Scholar] [CrossRef]
- Degrave, A.; Siamer, S.; Boureau, T.; Barny, M.A. The AvrE superfamily: Ancestral type III effectors involved in suppression of pathogen-associated molecular pattern-triggered immunity. Mol. Plant Pathol. 2015, 16, 899–905. [Google Scholar] [CrossRef]
- Stassen, M.J.J.; Hsu, S.-H.; Pieterse, C.M.J.; Stringlis, I.A. Coumarin communication along the microbiome–root–shoot axis. Trends Plant Sci. 2021, 26, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Batista, B.D.; Bazany, K.E.; Singh, B.K. Plant–microbiome interactions under a changing world: Responses, consequences and perspectives. New Phytol. 2022, 234, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Averkina, I.O.; Paponov, I.A.; Sánchez-Serrano, J.J.; Lillo, C. Specific PP2A Catalytic Subunits Are a Prerequisite for Positive Growth Effects in Arabidopsis Co-Cultivated with Azospirillum brasilense and Pseudomonas simiae. Plants 2020, 10, 66. [Google Scholar] [CrossRef]
- Quiroz, S.; Yustis, J.C.; Chávez-Hernández, E.C.; Martínez, T.; Sanchez, M.d.l.P.; Garay-Arroyo, A.; Álvarez-Buylla, E.R.; García-Ponce, B. Beyond the genetic pathways, flowering regulation complexity in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 5716. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.-Z.; Zhou, Y.-P.; Lv, T.-X.; Xie, C.-P.; Tian, C.-E. Research progress on the autonomous flowering time pathway in Arabidopsis. Physiol. Mol. Biol. Plants 2017, 23, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Heidari, B.; Nemie-Feyissa, D.; Kangasjärvi, S.; Lillo, C. Antagonistic regulation of flowering time through distinct regulatory subunits of protein phosphatase 2A. PLoS ONE 2013, 8, e67987. [Google Scholar] [CrossRef]
- Konert, G.; Rahikainen, M.; Trotta, A.; Durian, G.; Salojärvi, J.; Khorobrykh, S.; Tyystjärvi, E.; Kangasjärvi, S. Subunits B’ γ and B’ ζ of protein phosphatase 2A regulate photo-oxidative stress responses and growth in Arabidopsis thaliana. Plant Cell Environ. 2015, 38, 2641–2651. [Google Scholar] [CrossRef]
- Spinner, L.; Gadeyne, A.; Belcram, K.; Goussot, M.; Moison, M.; Duroc, Y.; Eeckhout, D.; De Winne, N.; Schaefer, E.; Van De Slijke, E. A protein phosphatase 2A complex spatially controls plant cell division. Nat. Commun. 2013, 4, 1863. [Google Scholar] [CrossRef]
- Zamariola, L.; De Storme, N.; Vannerum, K.; Vandepoele, K.; Armstrong, S.J.; Franklin, F.C.H.; Geelen, D. SHUGOSHIN s and PATRONUS protect meiotic centromere cohesion in A rabidopsis thaliana. Plant J. 2014, 77, 782–794. [Google Scholar] [CrossRef]
- Sablowski, R.; Gutierrez, C. Cycling in a crowd: Coordination of plant cell division, growth, and cell fate. Plant Cell 2022, 34, 193–208. [Google Scholar] [CrossRef]
- Desvoyes, B.; Gutierrez, C. Roles of plant retinoblastoma protein: Cell cycle and beyond. EMBO J. 2020, 39, e105802. [Google Scholar] [CrossRef] [PubMed]
- Ábrahám, E.; Yu, P.; Farkas, I.; Darula, Z.; Varga, E.; Lukács, N.; Ayaydin, F.; Medzihradszky, K.F.; Dombrádi, V.; Dudits, D. The B″regulatory subunit of protein phosphatase 2A mediates the dephosphorylation of rice retinoblastoma-related protein-1. Plant Mol. Biol. 2015, 87, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Shimotohno, A.; Aki, S.S.; Takahashi, N.; Umeda, M. Regulation of the plant cell cycle in response to hormones and the environment. Annu. Rev. Plant Biol. 2021, 72, 273–296. [Google Scholar] [CrossRef]
- Labandera, A.-M.; Vahab, A.R.; Chaudhuri, S.; Kerk, D.; Moorhead, G.B.G. The mitotic PP2A regulator ENSA/ARPP-19 is remarkably conserved across plants and most eukaryotes. Biochem. Biophys. Res. Commun. 2015, 458, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, J. Protein phosphatases in the regulation of mitosis. J. Cell Biol. 2019, 218, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-W.; Park, C.H.; Hsu, C.-C.; Kim, Y.-W.; Ko, Y.-W.; Zhang, Z.; Zhu, J.-Y.; Hsiao, Y.-C.; Branon, T.; Kaasik, K. Mapping the signaling network of BIN2 kinase using TurboID-mediated biotin labeling and phosphoproteomics. Plant Cell 2023, 35, 975–993. [Google Scholar] [CrossRef]


| Flowering Time Marker Genes | AGI Number | Flowering Pathway | B’56 Docking Motif |
|---|---|---|---|
| FLK (FLOWERING LOCUS K) | At3g04610 | Autonomous | IKKIVEETR |
| FLD (FLOWERING LOCUS D) | At3g10390 | Autonomous | LRGIYEPQG LYYLCETLG |
| FLC (FLOWERING LOCUS C) | At5g10140 | Autonomous | LVQLEEHLE |
| SVP (SHORT VEGETATIVE PHASE) | At2g22540 | Ambient temperature | MSEISELQK LTRVIETKS |
| SPL3 (SQUAMOSA PROMOTER BINDING PROTEIN-LIKE3) | At2g33810 | Age | LRELSEEEE |
| GI (GIGANTEA) | At1g22770 | Photoperiodic | LLGLLEAPP ICTIWEAAY LLKVLEYLP ILAILEALF |
| FKF1 (FLAVIN-BINDING KELCH REPEAT F-BOX 1) | At1g68050 | Photoperiodic | LNELHELCL |
| VRN2 (VERNALIZATION 2) | At4g16845 | Vernalization | CNTILENCR |
| MYB33 (MYB DOMAIN PROTEIN 33) | At5g06100 | Gibberellin | LGIVKETGS |
| Subunit | AGI Number | PP1 Docking Motifs and POLO Kinase Consensus |
|---|---|---|
| B’α | At5g03470 | TVIRGLLKFWPVTNCTKEVLFLGELEEVLE |
| B’ε | At3g54930 | TVIRGLLKFWPLTNCQKEVLFLGELEEVLD |
| B’γ | At4g15415 | TVIRGLLKYWPVTNSSKEVMFLGELEEVLE |
| B’ζ | At3g21650 | TVIRGLLKYWPVTNSSKEVMFLGELEEVLE |
| B’η | At3g26020 | TVIRGLLKYWPVTNSSKEVMFLNELEEVVI |
| B’θ | At1g13460 | TVIRGLLKSWPVTNSSKEVMFLNELEEVLE |
| B’κ | At5g25510 | VVIKGLLKFWPITNSQKEVMFLGEVEEIVE |
| B55α | At1g51690 | RERERGNHLATGDRGGRVVLFERTDTNNSS |
| B55β | At1g17720 | EFDKSGDHLATGDRGGRVVLFERTDTKDHG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heidari, B.; Nemie-Feyissa, D.; Lillo, C. Distinct Clades of Protein Phosphatase 2A Regulatory B’/B56 Subunits Engage in Different Physiological Processes. Int. J. Mol. Sci. 2023, 24, 12255. https://doi.org/10.3390/ijms241512255
Heidari B, Nemie-Feyissa D, Lillo C. Distinct Clades of Protein Phosphatase 2A Regulatory B’/B56 Subunits Engage in Different Physiological Processes. International Journal of Molecular Sciences. 2023; 24(15):12255. https://doi.org/10.3390/ijms241512255
Chicago/Turabian StyleHeidari, Behzad, Dugassa Nemie-Feyissa, and Cathrine Lillo. 2023. "Distinct Clades of Protein Phosphatase 2A Regulatory B’/B56 Subunits Engage in Different Physiological Processes" International Journal of Molecular Sciences 24, no. 15: 12255. https://doi.org/10.3390/ijms241512255
APA StyleHeidari, B., Nemie-Feyissa, D., & Lillo, C. (2023). Distinct Clades of Protein Phosphatase 2A Regulatory B’/B56 Subunits Engage in Different Physiological Processes. International Journal of Molecular Sciences, 24(15), 12255. https://doi.org/10.3390/ijms241512255