
In Silico Characterisation of the Aedes aegypti Gustatory Receptors

 , , and
, , and
Abstract
:1. Introduction
2. Results
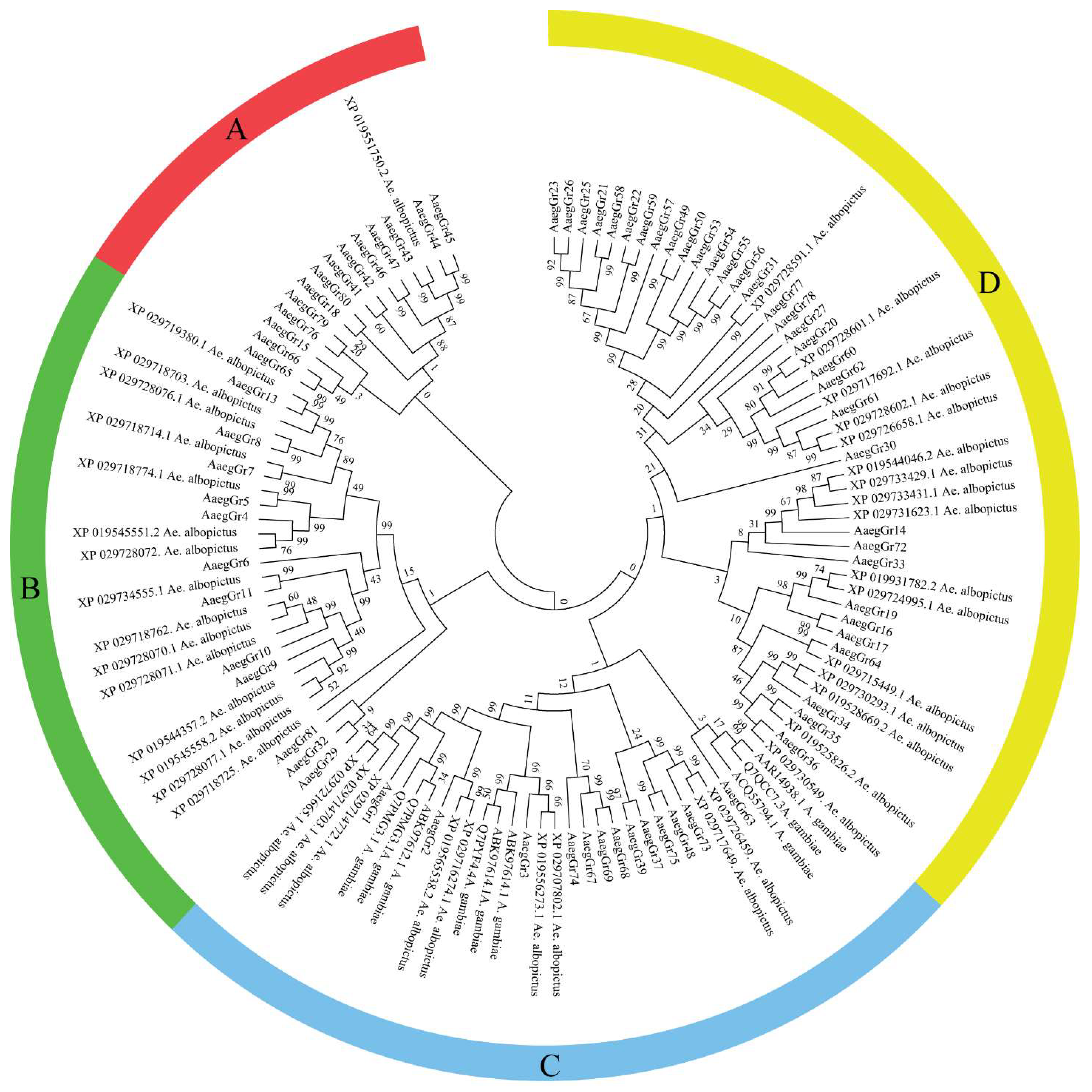
2.1. Identification of A. aegypti Gustatory Receptors and Phylogenetic Analysis
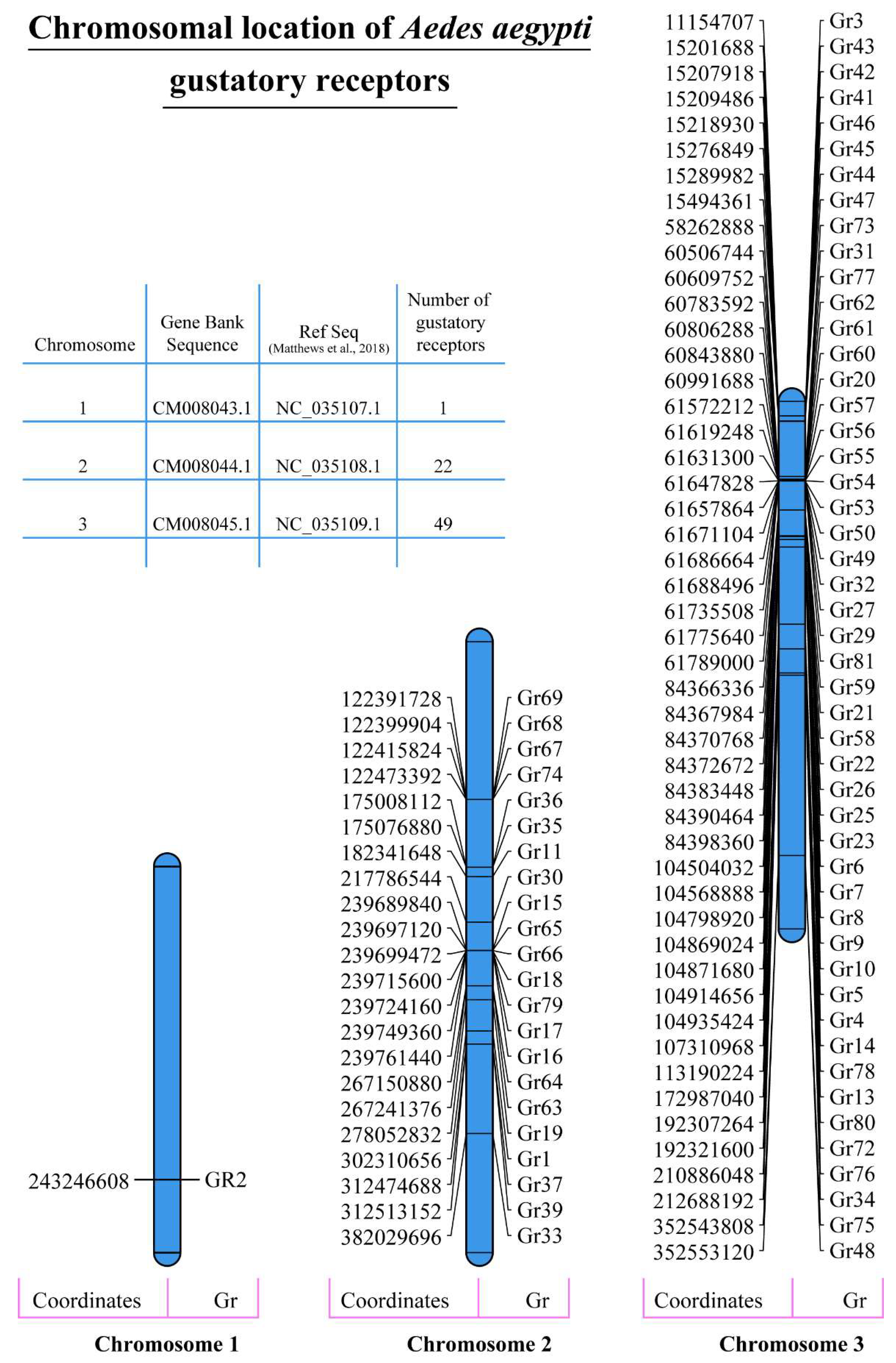
2.2. Physiochemical Characterisation and Chromosomal Distribution of A. aegypti Gustatory Receptors
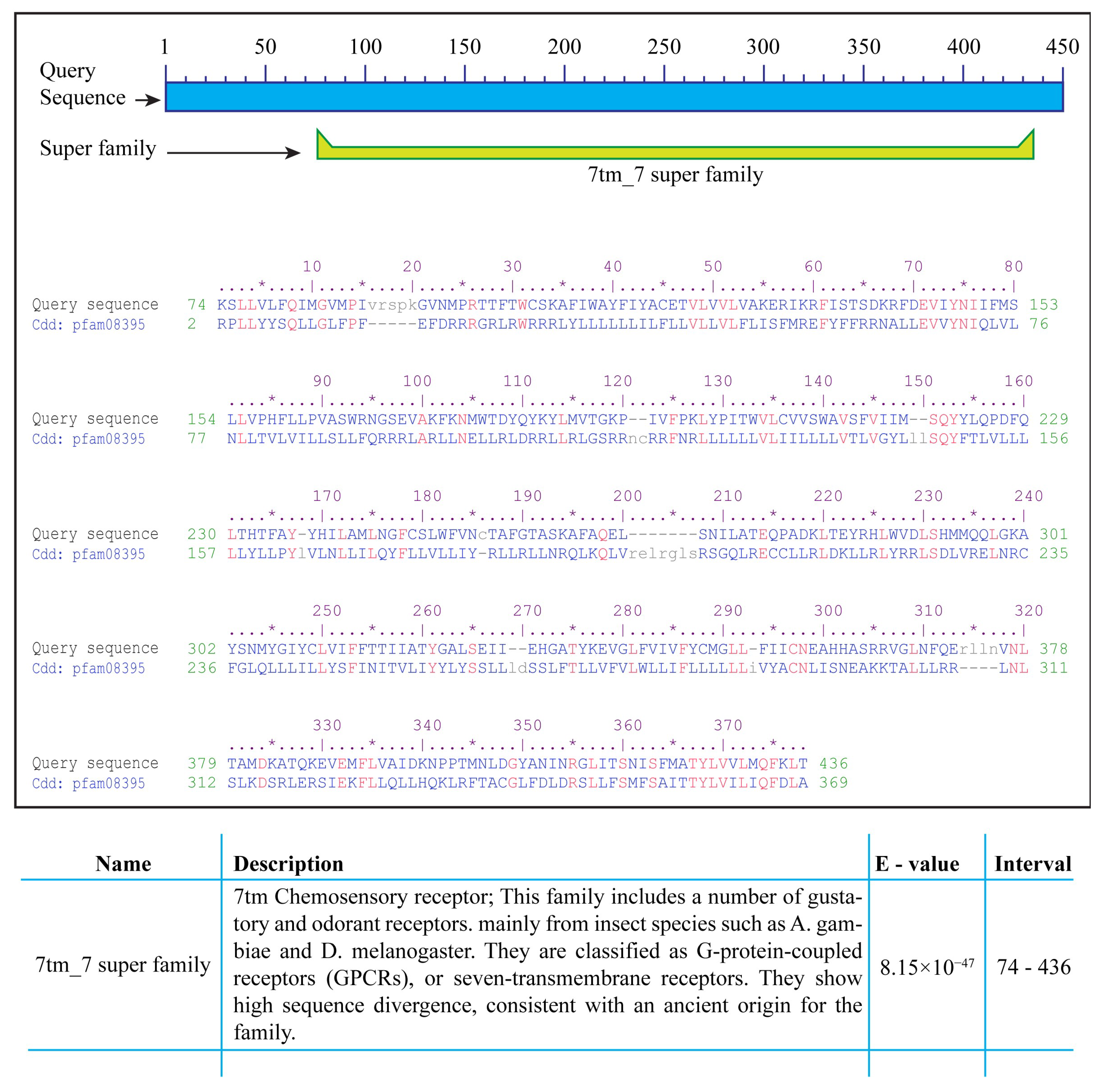
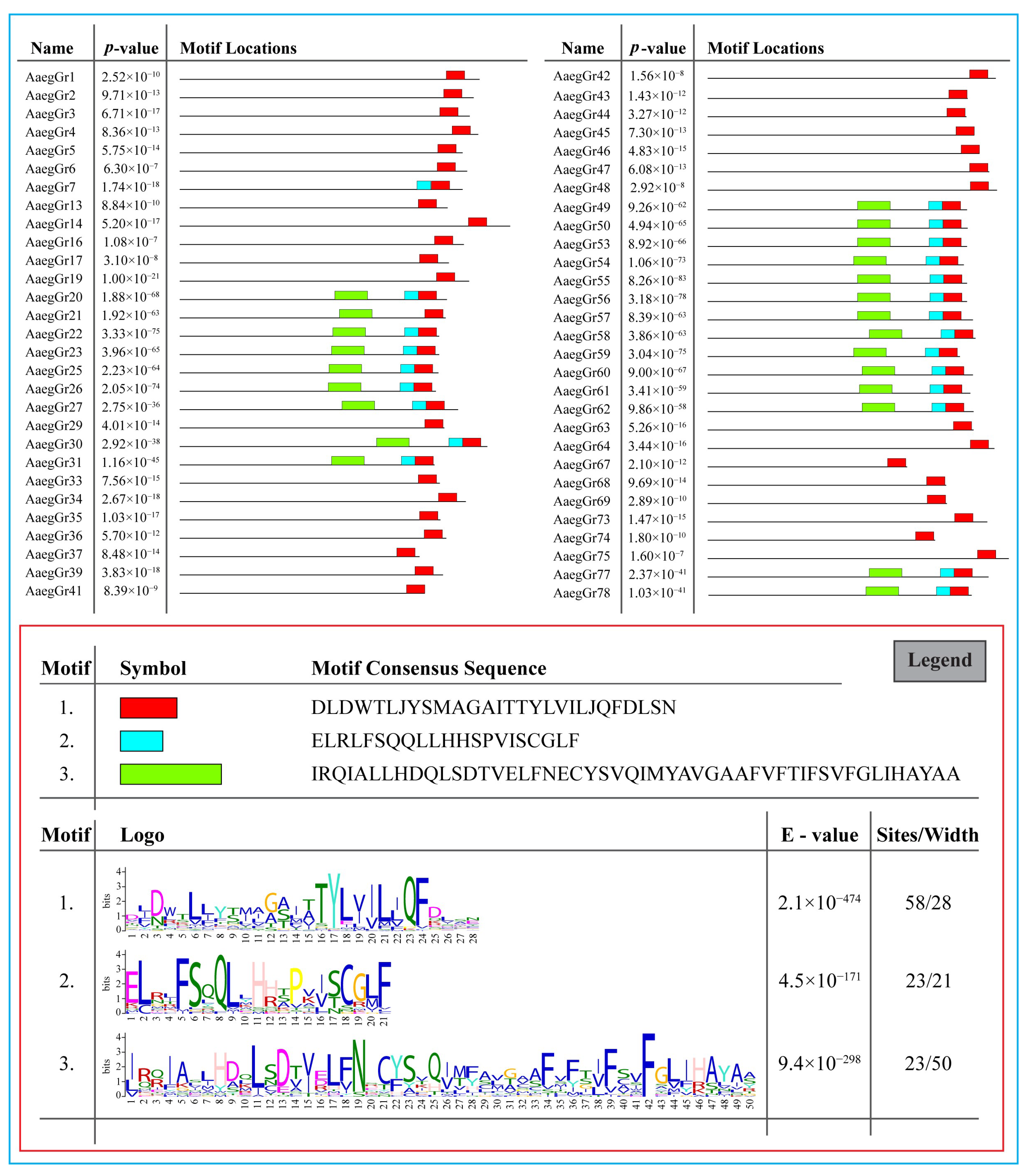
2.3. Identification of Conserved Domains and Motifs
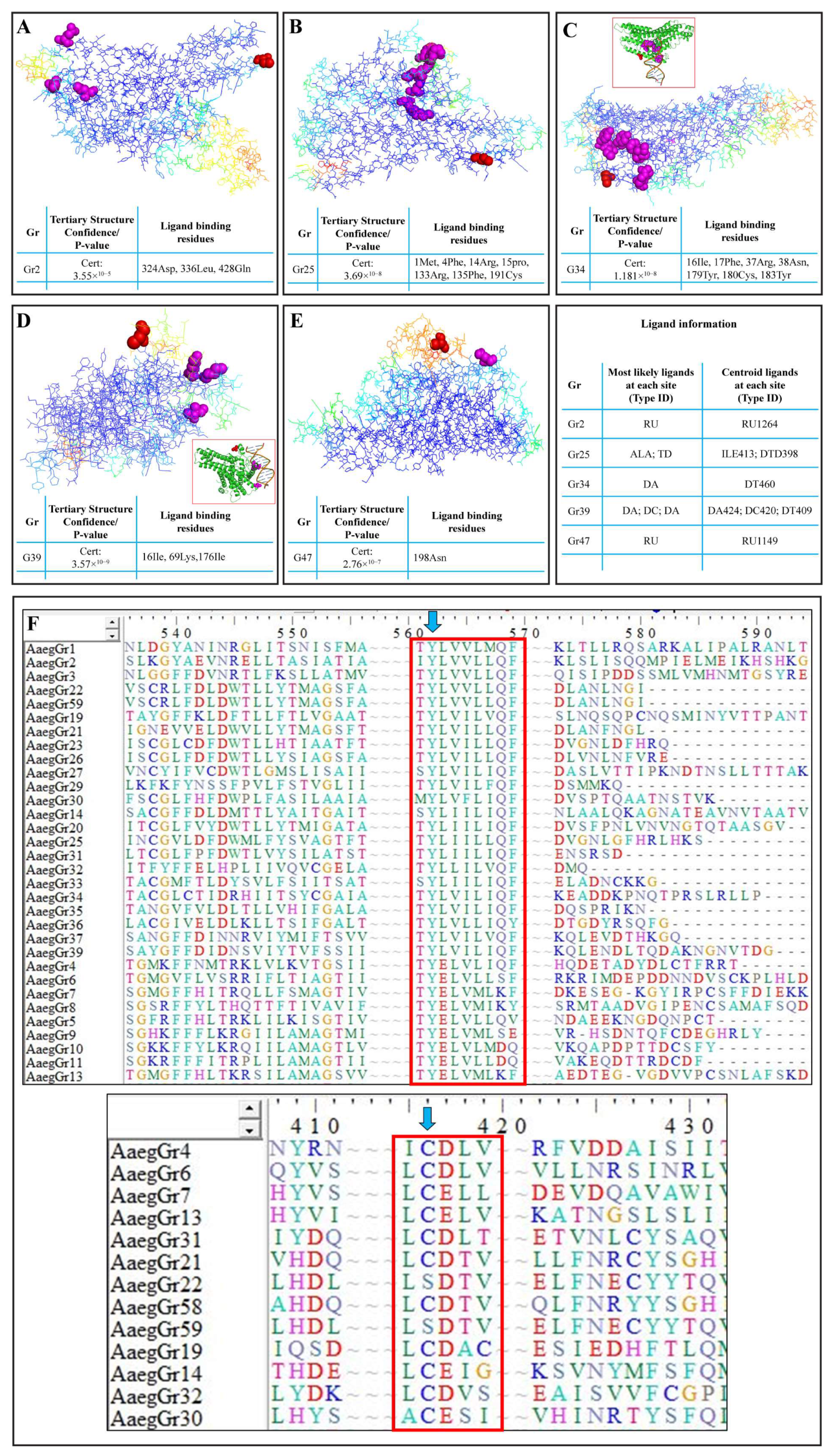
2.4. Homology Modelling and Prediction of S-Nitrosylation Sites in GR Proteins
3. Discussion
4. Materials and Methods
4.1. Identification of A. aegypti Gustatory Receptors and Phylogenetic Analysis
4.2. Physiochemical Characterisation
4.3. Chromosomal Distribution of A. aegypti Gustatory Receptors
4.4. Identification of Conserved Domains in A. aegypti Gustatory Receptors
4.5. Motif Composition Analysis
4.6. Homology Modelling, Tertiary Structure and Prediction of Ligand-Binding and S-Nitrosylation Sites in GR Proteins
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andrew, J.; Bar, A. Morphology and morphometry of Aedes aegypti adult mosquito. Annu. Res. Rev. Biol. 2013, 3, 52–69. [Google Scholar]
- Holeva-Eklund, W.M.; Young, S.J.; Will, J.; Busser, N.; Townsend, J.; Hepp, C.M. Species Distribution Modeling of Aedes aegypti in Maricopa County, Arizona from 2014–2020. Front. Environ. Sci. 2022, 10, 2107. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Munstermann, L.E.; Powell, J.R. Genetic distinctness of sympatric forms of Aedes aegypti in East Africa. Evolution 1979, 33, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Barrera, R. Competition and resistance to starvation in larvae of container-inhabiting Aedes mosquitoes. Ecol. Entomol. 1996, 21, 117–127. [Google Scholar] [CrossRef]
- Zettel, C.; Kaufman, P. Yellow fever mosquito Aedes aegypti (Linnaeus) (Insecta: Diptera: Culicidae). EDIS 2009, 2009, 1–8. [Google Scholar] [CrossRef]
- Alberts, B.; Bray, D.; Hopkin, K.; Johnson, A.D.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Essential Cell Biology; Garland Science: New York, NY, USA, 2015. [Google Scholar]
- Pask, G.M.; Ray, A. Insect Olfactory Receptors: An Interface between Chemistry and Biology. In Chemosensory Transduction; Zufall, F., Munger, S.D., Eds.; Academic Press: Amsterdam, The Netherlands, 2016; pp. 101–122. [Google Scholar]
- Clyne, P.J.; Warr, C.G.; Carlson, J.R. Candidate taste receptors in Drosophila. Science 2000, 287, 1830–1834. [Google Scholar] [CrossRef]
- McIver, S.; Siemicki, R. Fine structure of tarsal sensilla of Aedes aegypti (L.) (Diptera: Culicidae). J. Morphol. 1978, 155, 137–155. [Google Scholar] [CrossRef]
- McIver, S.B. Sensilla of mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1982, 19, 489–535. [Google Scholar] [CrossRef]
- McIver, S.; Siemicki, R. Innervation of cibarial sensilla of Aedes aegypti (L.) (Diptera: Culicidae). Int. J. Insect Morphol. Embryol. 1981, 10, 355–357. [Google Scholar] [CrossRef]
- Lee, R.M.K.W.; Davies, D.M. Cibarial sensilla of Toxorhynchites mosquitoes (Diptera: Culicidae). Int. J. Insect Morphol. Embryol. 1978, 7, 189–194. [Google Scholar] [CrossRef]
- Robertson, H.M.; Warr, C.G.; Carlson, J.R. Molecular evolution of the insect chemoreceptor gene superfamily in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2003, 100, 14537–14542. [Google Scholar] [CrossRef] [PubMed]
- Matthews, B.J.; Dudchenko, O.; Kingan, S.B.; Koren, S.; Antoshechkin, I.; Crawford, J.E.; Glassford, W.J.; Herre, M.; Redmond, S.N.; Rose, N.H.; et al. Improved reference genome of Aedes aegypti informs arbovirus vector control. Nature 2018, 563, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Arensburger, P.; Megy, K.; Waterhouse, R.M.; Abrudan, J.; Amedeo, P.; Antelo, B.; Bartholomay, L.; Bidwell, S.; Caler, E.; Camara, F.; et al. Sequencing of Culex quinquefasciatus establishes a platform for mosquito comparative genomics. Science 2010, 330, 86–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, J.T.; Vinyard, B.T.; Dickens, J.C. Gustatory receptor expression in the labella and tarsi of Aedes aegypti. Insect Biochem. Mol. Biol. 2013, 43, 1161–1171. [Google Scholar] [CrossRef]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.J.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome sequence of Aedes aegypti, a major arbovirus vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef] [Green Version]
- Timoshevskiy, V.A.; Severson, D.W.; Debruyn, B.S.; Black, W.C.; Sharakhov, I.V.; Sharakhova, M.V. An integrated linkage, chromosome, and genome map for the yellow fever mosquito Aedes aegypti. PLoS Neglected Trop. Dis. 2013, 7, e2052. [Google Scholar] [CrossRef] [Green Version]
- Dudchenko, O.; Batra, S.S.; Omer, A.D.; Nyquist, S.K.; Hoeger, M.; Durand, N.C.; Shamim, M.S.; Machol, I.; Lander, E.S.; Aiden, A.P.; et al. De novo assembly of the Aedes aegypti genome using Hi-C yields chromosome-length scaffolds. Science 2017, 356, 92–95. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.C.; Hazzard, J.M.H.; Nussenzveig, R.H.; Champagne, D.E.; Walker, F.A. Reversible binding of Nitric-Oxide by a salivary heme protein from a bloodsucking insect. Science 1993, 260, 539–541. [Google Scholar] [CrossRef]
- Walker, F.A. Nitric oxide interaction with insect nitrophorins and thoughts on the electron configuration of the {FeNO} 6 complex. J. Inorg. Biochem. 2005, 99, 216–236. [Google Scholar] [CrossRef]
- Damhus, T.; Hartshorn, R.; Hutton, A. Nomenclature of inorganic chemistry: IUPAC recommendations 2005. Chem. Int. 2005, 27, 25–26. [Google Scholar] [CrossRef]
- Davies, S.-A. Nitric oxide signalling in insects. Insect Biochem. Mol. Biol. 2000, 30, 1123–1138. [Google Scholar] [CrossRef] [PubMed]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.-G.; Yun, B.-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Hussain, A.; Mun, B.-G.; Imran, Q.M.; Lee, S.-U.; Adamu, T.A.; Shahid, M.; Kim, K.-M.; Yun, B.-W. Nitric oxide mediated transcriptome profiling reveals activation of multiple regulatory pathways in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Yun, B.-W.; Kim, J.H.; Gupta, K.J.; Hyung, N.-I.; Loake, G.J. Novel and conserved functions of S-nitrosoglutathione reductase in tomato. J. Exp. Bot. 2019, 70, 4877–4886. [Google Scholar] [CrossRef] [PubMed]
- Imran, Q.M.; Falak, N.; Hussain, A.; Mun, B.G.; Sharma, A.; Lee, S.U.; Kim, K.M.; Yun, B.W. Nitric Oxide Responsive Heavy Metal-Associated Gene AtHMAD1 Contributes to Development and Disease Resistance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pande, A.; Mun, B.-G.; Lee, D.-S.; Khan, M.; Lee, G.-M.; Hussain, A.; Yun, B.-W. NO network for plant–microbe communication underground: A review. Front. Plant Sci. 2021, 12, 658–679. [Google Scholar] [CrossRef]
- Roberts Jr, J.D.; Lang, P.; Bigatello, L.M.; Vlahakes, G.J.; Zapol, W.M. Inhaled nitric oxide in congenital heart disease. Circulation 1993, 87, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Gusarov, I.; Gautier, L.; Smolentseva, O.; Shamovsky, I.; Eremina, S.; Mironov, A.; Nudler, E. Bacterial nitric oxide extends the lifespan of C. elegans. Cell 2013, 152, 818–830. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Jia, F.; Shao, S.; Zhang, H.; Li, G.; Xia, X.; Zhou, Y.; Yu, J.; Shi, K. Involvement of nitric oxide in the jasmonate-dependent basal defense against root-knot nematode in tomato plants. Front. Plant Sci. 2015, 6, 193. [Google Scholar] [CrossRef] [Green Version]
- Sadekuzzaman, M.; Stanley, D.; Kim, Y. Nitric oxide mediates insect cellular immunity via phospholipase A2 activation. J. Innate Immun. 2018, 10, 70–81. [Google Scholar] [CrossRef]
- Müller, U. The molecular signalling processes underlying olfactory learning and memory formation in honeybees. Apidologie 2012, 43, 322–333. [Google Scholar] [CrossRef] [Green Version]
- Trimmer, B.A.; Aprille, J.R.; Dudzinski, D.M.; Lagace, C.J.; Lewis, S.M.; Michel, T.; Qazi, S.; Zayas, R.M. Nitric oxide and the control of firefly flashing. Science 2001, 292, 2486–2488. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, H.; Yokoyama, J.; Ohba, N.; Ohmiya, Y.; Kawata, M.; Shelly, T. Expression of the NOS gene and firefly flashing: A test of the nitric-oxide-mediated flash control model. J. Insect Sci. 2014, 14, 56. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.A.; Rillich, J. Adding up the odds—Nitric oxide signaling underlies the decision to flee and post-conflict depression of aggression. Sci. Adv. 2015, 1, e1500060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-B. Nitric oxide fumigation for control of western flower thrips and its safety to postharvest quality of fresh fruit and vegetables. J. Asia-Pac. Entomol. 2016, 19, 1191–1195. [Google Scholar] [CrossRef]
- Hussain, A.; Imran, Q.M.; Shahid, M.; Yun, B.-W. Nitric oxide synthase in the plant kingdom. In Nitric Oxide in Plant Biology; Pratap Singh, V., Singh, S., Tripathi, D.K., Romero-Puertas, M.C., Sandalio, L.M., Eds.; Academic Press: Amsterdam, The Netherlands, 2022; pp. 43–52. [Google Scholar]
- Hess, D.T.; Matsumoto, A.; Kim, S.-O.; Marshall, H.E.; Stamler, J.S. Protein S-nitrosylation: Purview and parameters. Nat. Rev. Mol. Cell Biol. 2005, 6, 150–166. [Google Scholar] [CrossRef]
- Elphick, M.R.; Green, I.C.; O’Shea, M. Nitric oxide synthesis and action in an invertebrate brain. Brain Res. 1993, 619, 344–346. [Google Scholar] [CrossRef]
- Bredt, D.S.; Snyder, S.H. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 682–685. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: Amsterdam, The Netherlands, 1965; pp. 97–166. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guruprasad, K.; Reddy, B.V.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. Des. Sel. 1990, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Ikai, A. Thermostability and aliphatic index of globular proteins. J. Biochem. 1980, 88, 1895–1898. [Google Scholar] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Tan, P.L.; Chrystal, M.A.; Cravchik, A.; Collins, F.H.; Robertson, H.M.; Zwiebel, L.J. G protein-coupled receptors in Anopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the International Conference on Intelligent Systems for Molecular Biology, Madison, WI, USA, 14−17 August 1994; Volume 2, pp. 28–36. [Google Scholar]
- Rockman, M.V.; Skrovanek, S.S.; Kruglyak, L. Selection at linked sites shapes heritable phenotypic variation in C. elegans. Science 2010, 330, 372–376. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.L.; Dickson, L.B.; Lozano-Fuentes, S.; Juneja, P.; Jiggins, F.M.; Black, W.C. Alternative patterns of sex chromosome differentiation in Aedes aegypti (L). BMC Genom. 2017, 18, 943. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, M.J.; Terrile, M.C.; Correa-Aragunde, N.; Colman, S.L.; Izquierdo-Álvarez, A.; Fiol, D.F.; París, R.; Sánchez-López, N.; Marina, A.; Calderón Villalobos, L.I.A.; et al. Regulation of SCF(TIR1/AFBs) E3 ligase assembly by S-nitrosylation of Arabidopsis SKP1-like1 impacts on auxin signaling. Redox Biol. 2018, 18, 200–210. [Google Scholar] [CrossRef]
- Terrile, M.C.; París, R.; Calderón-Villalobos, L.I.; Iglesias, M.J.; Lamattina, L.; Estelle, M.; Casalongué, C.A. Nitric oxide influences auxin signaling through S-nitrosylation of the Arabidopsis TRANSPORT INHIBITOR RESPONSE 1 auxin receptor. Plant J. Cell Mol. Biol. 2012, 70, 492–500. [Google Scholar] [CrossRef] [Green Version]
- Albertos, P.; Romero-Puertas, M.C.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Nambara, E.; Lorenzo, O. S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth. Nat. Commun. 2015, 6, 8669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada, Y.; Spoel, S.H.; Pajerowska-Mukhtar, K.; Mou, Z.L.; Song, J.Q.; Wang, C.; Zuo, J.R.; Dong, X.N. Plant immunity requires conformational changes of NPR1 via S-nitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayyar, P.V. Uncovering the Role of S-Nitrosylation in Jasmonic Acid Signalling During the Plant Immune Response; IMPS; The University of Edinburgh: Edinburgh, UK, 2016. [Google Scholar]
- Weichsel, A.; Maes, E.M.; Andersen, J.F.; Valenzuela, J.G.; Shokhireva, T.; Walker, F.A.; Montfort, W.R. Heme-assisted S-nitrosation of a proximal thiolate in a nitric oxide transport protein. Proc. Natl. Acad. Sci. USA 2005, 102, 594–599. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program For Windows 95/98/NT; Nucleic Acids Symposium Series, 1999; Oxford Academic: Oxford, UK, 1999; pp. 95–98. [Google Scholar]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- McGuffin, L.J.; Shuid, A.N.; Kempster, R.; Maghrabi, A.H.A.; Nealon, J.O.; Salehe, B.R.; Atkins, J.D.; Roche, D.B. Accurate template-based modeling in CASP12 using the IntFOLD4-TS, ModFOLD6, and ReFOLD methods. Proteins 2018, 86, 335–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuffin, L.J.; Adiyaman, R.; Maghrabi, A.H.A.; Shuid, A.N.; Brackenridge, D.A.; Nealon, J.O.; Philomina, L.S. IntFOLD: An integrated web resource for high performance protein structure and function prediction. Nucleic Acids Res. 2019, 47, W408–W413. [Google Scholar] [CrossRef] [PubMed]
- Roche, D.B.; Tetchner, S.J.; McGuffin, L.J. FunFOLD: An improved automated method for the prediction of ligand binding residues using 3D models of proteins. BMC Bioinform. 2011, 12, 160. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Liu, Z.X.; Gao, X.J.; Jin, C.J.; Wen, L.P.; Yao, X.B.; Ren, J.A. GPS-SNO: Computational prediction of protein S-Nitrosylation sites with a modified GPS algorithm. PLoS ONE 2010, 5, e11290. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptors | Physicochemical Properties | Subcellular Localization | ||||||
|---|---|---|---|---|---|---|---|---|
| Gustatory Receptor | M Weight (da) | Instability Index | Aliphatic Index | Hydropathicity (GRAVY) | Length | pI | Extracellular Matrix | Plasma Membrane |
| AaegGr1 | 53,444.62 | 41.52 | 98.35 | 0.101 | 460 | 9.04 | 18 | 12 |
| AaegGr2 | 51,563.44 | 32.05 | 113.08 | 0.394 | 451 | 6.63 | 20 | 10 |
| AaegGr3 | 50,713.64 | 41.93 | 101.98 | 0.411 | 445 | 8.28 | 23 | 7 |
| AaegGr4 | 53,284.49 | 36.36 | 95.94 | 0.17 | 458 | 9.59 | 16 | 11 |
| AaegGr5 | 50,232.77 | 35.02 | 101.01 | 0.225 | 434 | 9.43 | 19 | 8 |
| AaegGr6 | 51,286.81 | 50.57 | 104.29 | 0.254 | 441 | 8.93 | 18 | 9 |
| AaegGr7 | 50,049.73 | 34.02 | 109.84 | 0.301 | 434 | 8.81 | 18 | 12 |
| AaegGr8 | 48,289.27 | 41.6 | 99.13 | 0.392 | 412 | 8.74 | 12 | 10 |
| AaegGr9 | 50,837.81 | 42.43 | 105.76 | 0.312 | 434 | 9.16 | 20 | 10 |
| AaegGr10 | 53,308.31 | 37.88 | 97.93 | 0.274 | 455 | 9.16 | 18 | 10 |
| AaegGr11 | 54,851.61 | 30.85 | 105.37 | 0.395 | 471 | 9.18 | 21 | 9 |
| AaegGr13 | 47,425.52 | 41.39 | 109.05 | 0.387 | 412 | 7.6 | 19 | 9 |
| AaegGr14 | 57,732.89 | 40 | 110.53 | 0.351 | 507 | 7.21 | 19 | 9 |
| AaegGr15 | 45,520.29 | 35.96 | 112.76 | 0.458 | 381 | 8.61 | 17 | 7 |
| AaegGr16 | 51,633.77 | 38.72 | 106.58 | 0.324 | 436 | 8.86 | 19 | 11 |
| AaegGr17 | 48,875.03 | 33.83 | 107.8 | 0.3 | 413 | 9.44 | 20 | 10 |
| AaegGr18 | 43,743.17 | 39.96 | 120.45 | 0.488 | 374 | 9.1 | 17 | 10 |
| AaegGr19 | 50,495.17 | 40.54 | 110.43 | 0.446 | 444 | 8.53 | 19 | 10 |
| AaegGr20 | 47,285.68 | 39.29 | 109.22 | 0.547 | 410 | 8.32 | 7 | 11 |
| AaegGr21 | 47,288.6 | 31.81 | 114.14 | 0.395 | 408 | 8.8 | 22 | 8 |
| AaegGr22 | 45,861.16 | 29.16 | 101.41 | 0.279 | 398 | 7.69 | 20 | 10 |
| AaegGr23 | 46,417.3 | 37.96 | 106.26 | 0.356 | 398 | 9.08 | 21 | 9 |
| AaegGr25 | 45,282.08 | 48.52 | 114.33 | 0.475 | 397 | 8.51 | 18 | 11 |
| AaegGr26 | 45,332.87 | 32.87 | 103.94 | 0.316 | 393 | 9.1 | 22 | 8 |
| AaegGr27 | 48,633.48 | 38.66 | 111.41 | 0.451 | 427 | 9.4 | 20 | 10 |
| AaegGr29 | 47,496.89 | 40.28 | 107.22 | 0.291 | 406 | 9.17 | 16 | 11 |
| AaegGr30 | 54,471.95 | 46.85 | 112.33 | 0.393 | 472 | 9.14 | 20 | 10 |
| AaegGr31 | 44,257.42 | 33.02 | 124.4 | 0.732 | 391 | 6.43 | 22 | 8 |
| AaegGr32 | 46,508.67 | 43.07 | 125.2 | 0.481 | 404 | 6.15 | 19 | 11 |
| AaegGr33 | 45,832.22 | 43.61 | 125.06 | 0.441 | 399 | 8.63 | 22 | 8 |
| AaegGr34 | 49,575.45 | 37.58 | 110.3 | 0.351 | 439 | 9.11 | 14 | 13 |
| AaegGr35 | 46,145.76 | 35.65 | 119.1 | 0.467 | 400 | 9.51 | 22 | 8 |
| AaegGr36 | 47,530.12 | 28.35 | 116.19 | 0.452 | 409 | 8.27 | 19 | 9 |
| AaegGr37 | 43,274.2 | 43.18 | 114.05 | 0.476 | 368 | 8.61 | 19 | 9 |
| AaegGr39 | 46,630.56 | 32.32 | 109.95 | 0.449 | 404 | 9.34 | 19 | 10 |
| AaegGr41 | 44,001.3 | 25.44 | 121.25 | 0.705 | 376 | 8.5 | 19 | 8 |
| AaegGr42 | 51,572.17 | 39.14 | 112.76 | 0.459 | 442 | 9.33 | 18 | 9 |
| AaegGr43 | 46,364.08 | 36.59 | 121.13 | 0.607 | 399 | 8.78 | 19 | 8 |
| AaegGr44 | 45,955.25 | 24.94 | 118.04 | 0.472 | 397 | 8.43 | 6 | 11 |
| AaegGr45 | 47,807.09 | 42.35 | 98.66 | 0.253 | 410 | 6.18 | 19 | 10 |
| AaegGr46 | 48,429.06 | 26.81 | 109.4 | 0.323 | 417 | 9.5 | 21 | 9 |
| AaegGr47 | 50,471.36 | 32.11 | 111.2 | 0.263 | 432 | 9.43 | 20 | 10 |
| AaegGr48 | 51,570.11 | 25.41 | 111.08 | 0.436 | 444 | 9.28 | 18 | 9 |
| AaegGr49 | 46,189.63 | 36.04 | 101.61 | 0.273 | 398 | 9.3 | 8 | 11 |
| AaegGr50 | 46,375.8 | 36.45 | 97.39 | 0.299 | 399 | 8.9 | 18 | 9 |
| AaegGr53 | 45,703.47 | 37.67 | 112.86 | 0.573 | 398 | 9.29 | 15 | 12 |
| AaegGr54 | 45,563.19 | 42.34 | 97.25 | 0.344 | 393 | 8.97 | 12 | 11 |
| AaegGr55 | 46,453.84 | 41.32 | 96.26 | 0.352 | 398 | 9.25 | 19 | 8 |
| AaegGr56 | 46,242.62 | 43.95 | 103.39 | 0.432 | 398 | 9.06 | 17 | 10 |
| AaegGr57 | 46,338.85 | 39.62 | 117.59 | 0.531 | 407 | 8.74 | 23 | 7 |
| AaegGr58 | 47,868.28 | 32.1 | 111.41 | 0.334 | 411 | 8.83 | 22 | 8 |
| AaegGr59 | 44,534.76 | 29.95 | 101.27 | 0.251 | 387 | 7.72 | 20 | 10 |
| AaegGr60 | 46,826.52 | 33.15 | 111.99 | 0.578 | 407 | 8.66 | 19 | 9 |
| AaegGr61 | 45,627.76 | 28.62 | 116 | 0.599 | 403 | 7.49 | 6 | 11 |
| AaegGr62 | 46,596.22 | 37.12 | 118.85 | 0.512 | 408 | 8.97 | 19 | 9 |
| AaegGr63 | 46,553.49 | 47.49 | 116.89 | 0.314 | 408 | 9.24 | 16 | 9 |
| AaegGr64 | 50,213.48 | 33.39 | 110.95 | 0.184 | 440 | 8.59 | 15 | 8 |
| AaegGr65 | 46,745.81 | 46.41 | 107.04 | 0.371 | 395 | 8.83 | 20 | 10 |
| AaegGr66 | 45,292.62 | 40.56 | 120.2 | 0.372 | 380 | 9.36 | 19 | 9 |
| AaegGr67 | 35,207.96 | 31.8 | 121.37 | 0.495 | 306 | 9.33 | 19 | 9 |
| AaegGr68 | 42,279.62 | 33.99 | 124.13 | 0.606 | 366 | 5.54 | 17 | 10 |
| AaegGr69 | 42,413.22 | 35.1 | 114.47 | 0.597 | 367 | 7.58 | 21 | 9 |
| AaegGr72 | 44,179.17 | 40.55 | 109.5 | 0.371 | 381 | 8.62 | 22 | 8 |
| AaegGr73 | 48,550.67 | 33.11 | 100.91 | 0.321 | 429 | 8.74 | 22 | 8 |
| AaegGr74 | 41,027.28 | 42.18 | 102.26 | 0.282 | 349 | 9.16 | 19 | 11 |
| AaegGr75 | 53,206.08 | 28.21 | 113.92 | 0.496 | 462 | 9.36 | 19 | 10 |
| AaegGr76 | 45,984.17 | 36.07 | 117.47 | 0.573 | 395 | 5.88 | 19 | 9 |
| AaegGr77 | 49,365.32 | 45.77 | 114.48 | 0.559 | 431 | 7.13 | 11 | 10 |
| AaegGr78 | 47,010.94 | 28.17 | 107.08 | 0.296 | 404 | 8.67 | 16 | 10 |
| AaegGr79 | 46,939.44 | 36.47 | 125.07 | 0.417 | 406 | 8.86 | 20 | 8 |
| AaegGr80 | 43,910.87 | 34.06 | 124.89 | 0.438 | 380 | 8.67 | 18 | 9 |
| AaegGr81 | 44,339.37 | 32.21 | 104.97 | 0.338 | 390 | 9.96 | 19 | 11 |
| GR | Position | Peptide | Score | Cutoff | Cluster |
|---|---|---|---|---|---|
| AaegGr2 | 270 | RKDVAIECTAAMISQ | 4.842 | 2.443 | Cluster B |
| AaegGr3 | 206 | ILLPVLSCLAVIITH | 3.478 | 2.443 | Cluster B |
| AaegGr4 | 371 | RTLAVSMCTAAVNDE | 2.815 | 2.443 | Cluster B |
| AaegGr7 | 158 | ALLLGLACCEHLLAT | 2.658 | 2.443 | Cluster B |
| AaegGr8 | 235 | FWRIEVACNGTVLPT | 3.408 | 2.443 | Cluster B |
| AaegGr9 | 8 | MQAPNQHCLAQLRKW | 3.147 | 2.443 | Cluster B |
| AaegGr10 | 326 | GHLILLSCANDMYFI | 2.63 | 2.443 | Cluster B |
| AaegGr11 | 285 | YIDVFIICVSLVLQR | 2.804 | 2.443 | Cluster B |
| AaegGr13 | 404 | GVGDVVPCSNLAFSK | 3.25 | 2.443 | Cluster B |
| AaegGr14 | 95 | YMEPLMMCIDMLAAM | 2.946 | 2.443 | Cluster B |
| 111 | NQKRLIECVERLDKV | 3.163 | 2.443 | Cluster B | |
| AaegGr15 | 6 | MAKISCLYRHVLK | 2.668 | 2.443 | Cluster B |
| AaegGr16 | 84 | ILLTLSVCSAEILIA | 3.582 | 2.443 | Cluster B |
| AaegGr20 | 337 | LVHKAINCASSSAVI | 3.168 | 2.443 | Cluster B |
| AaegGr23 | 116 | ILANINDCDRKLGKL | 2.652 | 2.443 | Cluster B |
| AaegGr25 | 343 | RVLKELRCFSQQLQH | 2.5 | 2.443 | Cluster B |
| AaegGr26 | 145 | LSTGVWMCFSVIITL | 2.495 | 2.443 | Cluster B |
| AaegGr32 | 221 | RLQLLNRCLEEMLLE | 2.793 | 2.443 | Cluster B |
| AaegGr33 | 396 | QFELADNCKKG | 1.738 | 1.484 | Cluster A |
| AaegGr34 | 51 | YGLGIVFCLAGLTYK | 3.299 | 2.443 | Cluster B |
| 368 | VCNLMRTCKDSLTKE | 4.049 | 2.443 | Cluster B | |
| AaegGr39 | 7 | MLSFRPCRNKYIQQ | 3.337 | 2.443 | Cluster B |
| AaegGr43 | 5 | MHTTCRTVFRLK | 4.874 | 1.484 | Cluster A |
| 345 | FYNDAGRCVEQSIEM | 3.424 | 2.443 | Cluster B | |
| AaegGr46 | 4 | MFHCSQNPLLS | 3.033 | 2.443 | Cluster B |
| AaegGr47 | 12 | KTSHKKACIHDKTYQ | 1.743 | 1.484 | Cluster A |
| 313 | IMGVFIACVTTVNDI | 2.696 | 2.443 | Cluster B | |
| AaegGr49 | 54 | LVMGVFMCVGAMYYS | 2.663 | 2.443 | Cluster B |
| AaegGr53 | 251 | VVVLFNKCFSKLVMF | 3.495 | 2.443 | Cluster B |
| AaegGr54 | 184 | AFSWVMGCYQTLAST | 2.685 | 2.443 | Cluster B |
| AaegGr55 | 184 | AFWWVMSCYQTMTSI | 3.136 | 2.443 | Cluster B |
| AaegGr57 | 300 | NMVYNIFCSGFIIQL | 2.522 | 2.443 | Cluster B |
| AaegGr60 | 2 | MCYRAVNIY | 5.093 | 1.484 | Cluster A |
| 336 | LVHKAINCSTSSVVI | 4.049 | 2.443 | Cluster B | |
| AaegGr62 | 53 | WLNLLGNCISYLLVV | 2.5 | 2.443 | Cluster B |
| 337 | VYKGITNCPSSAVKN | 4.842 | 2.443 | Cluster B | |
| AaegGr63 | 6 | MIGNICLFSSKPF | 3.448 | 1.484 | Cluster A |
| AaegGr64 | 72 | STLGILQCVAACVGY | 3.027 | 2.443 | Cluster B |
| AaegGr66 | 333 | LHQLSISCINQFRAL | 2.495 | 2.443 | Cluster B |
| AaegGr67 | 207 | NLGFFVQCLDEIDEL | 3.337 | 2.443 | Cluster B |
| AaegGr69 | 110 | KLLHFDQCYNAMINS | 3.87 | 2.443 | Cluster B |
| AaegGr73 | 104 | ILSIFTVCDEKMRTM | 3.56 | 2.443 | Cluster B |
| 137 | ACIVTLTCGGTLGGL | 2.875 | 2.443 | Cluster B | |
| AaegGr76 | 107 | IYWRQFMCNERIQQL | 3.212 | 2.443 | Cluster B |
| 341 | LQRIGIVCLDSRLTE | 2.788 | 2.443 | Cluster B | |
| 349 | LDSRLTECIYGLSKV | 2.549 | 2.443 | Cluster B | |
| 359 | GLSKVVQCMQEETVM | 3.06 | 2.443 | Cluster B | |
| 385 | TTIIAASCSYLILLI | 1.874 | 1.484 | Cluster A | |
| AaegGr77 | 291 | FSIFSMFCIGALVSY | 3.402 | 2.443 | Cluster B |
| AaegGr79 | 221 | LHVQILSCYMALINV | 2.484 | 2.443 | Cluster B |
| AaegGr80 | 7 | MAILVACHYIMEKH | 2.495 | 2.443 | Cluster B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bibi, M.; Hussain, A.; Ali, F.; Ali, A.; Said, F.; Tariq, K.; Yun, B.-W. In Silico Characterisation of the Aedes aegypti Gustatory Receptors. Int. J. Mol. Sci. 2023, 24, 12263. https://doi.org/10.3390/ijms241512263
Bibi M, Hussain A, Ali F, Ali A, Said F, Tariq K, Yun B-W. In Silico Characterisation of the Aedes aegypti Gustatory Receptors. International Journal of Molecular Sciences. 2023; 24(15):12263. https://doi.org/10.3390/ijms241512263
Chicago/Turabian StyleBibi, Maria, Adil Hussain, Farman Ali, Asad Ali, Fazal Said, Kaleem Tariq, and Byung-Wook Yun. 2023. "In Silico Characterisation of the Aedes aegypti Gustatory Receptors" International Journal of Molecular Sciences 24, no. 15: 12263. https://doi.org/10.3390/ijms241512263
APA StyleBibi, M., Hussain, A., Ali, F., Ali, A., Said, F., Tariq, K., & Yun, B. -W. (2023). In Silico Characterisation of the Aedes aegypti Gustatory Receptors. International Journal of Molecular Sciences, 24(15), 12263. https://doi.org/10.3390/ijms241512263