
Adult and Larval Tracheal Systems Exhibit Different Molecular Architectures in Drosophila

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Genes That Are Specifically Expressed in Adult Trachea
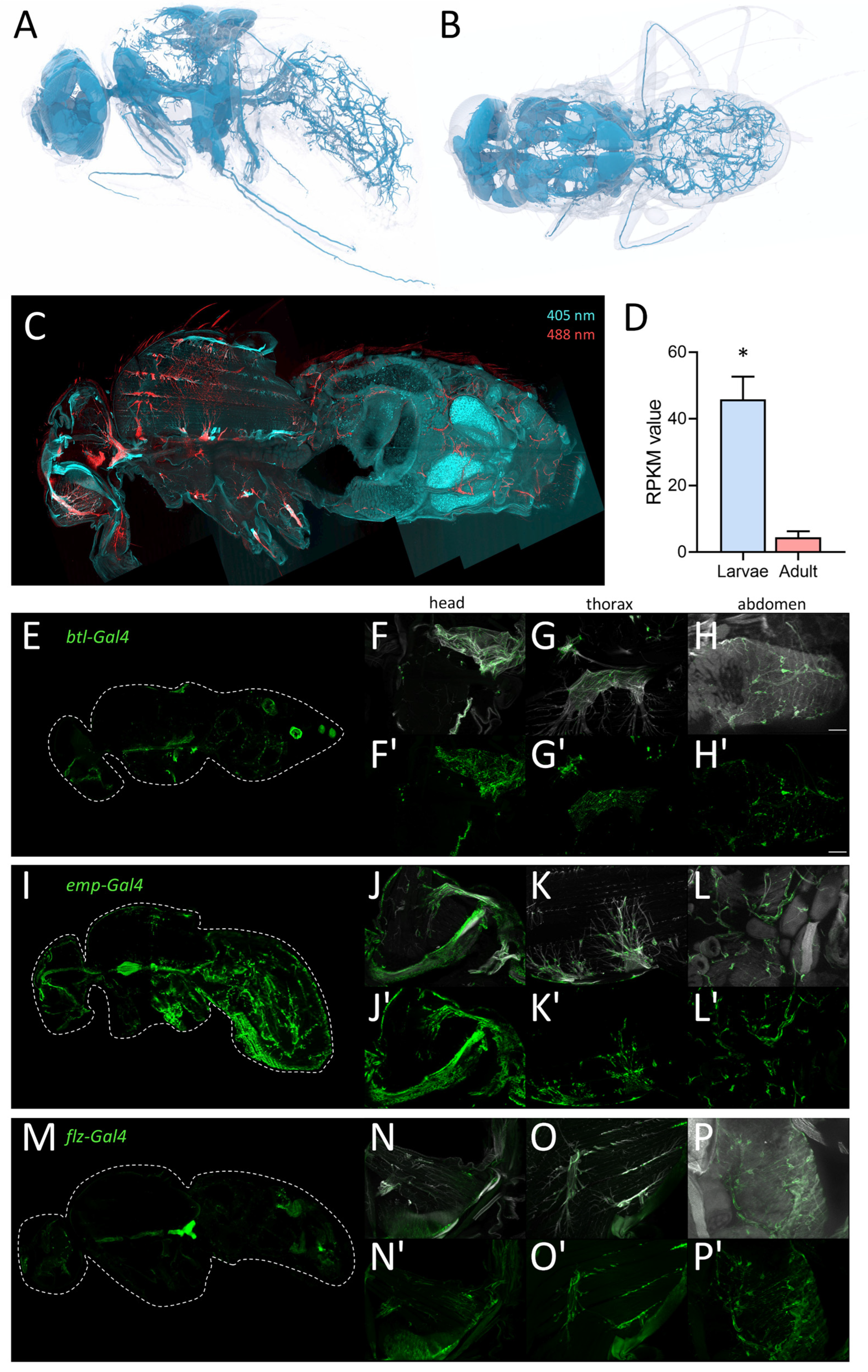
2.2. The Respiratory Systems from Adults and Larvae Are Clearly Distinct
2.3. ECM and Chitin-Associated Genes Are Overrepresented in Larval Trachea
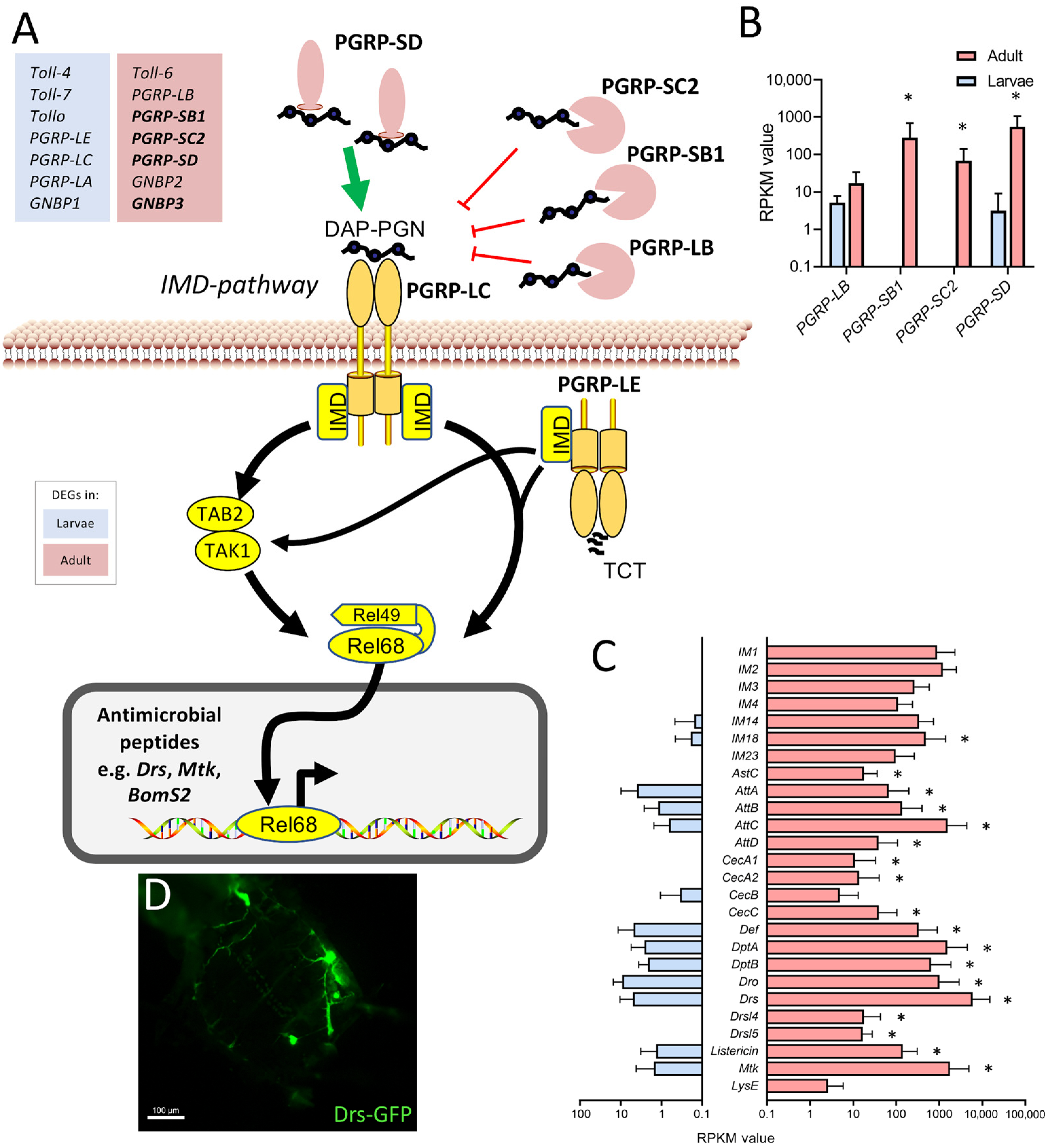
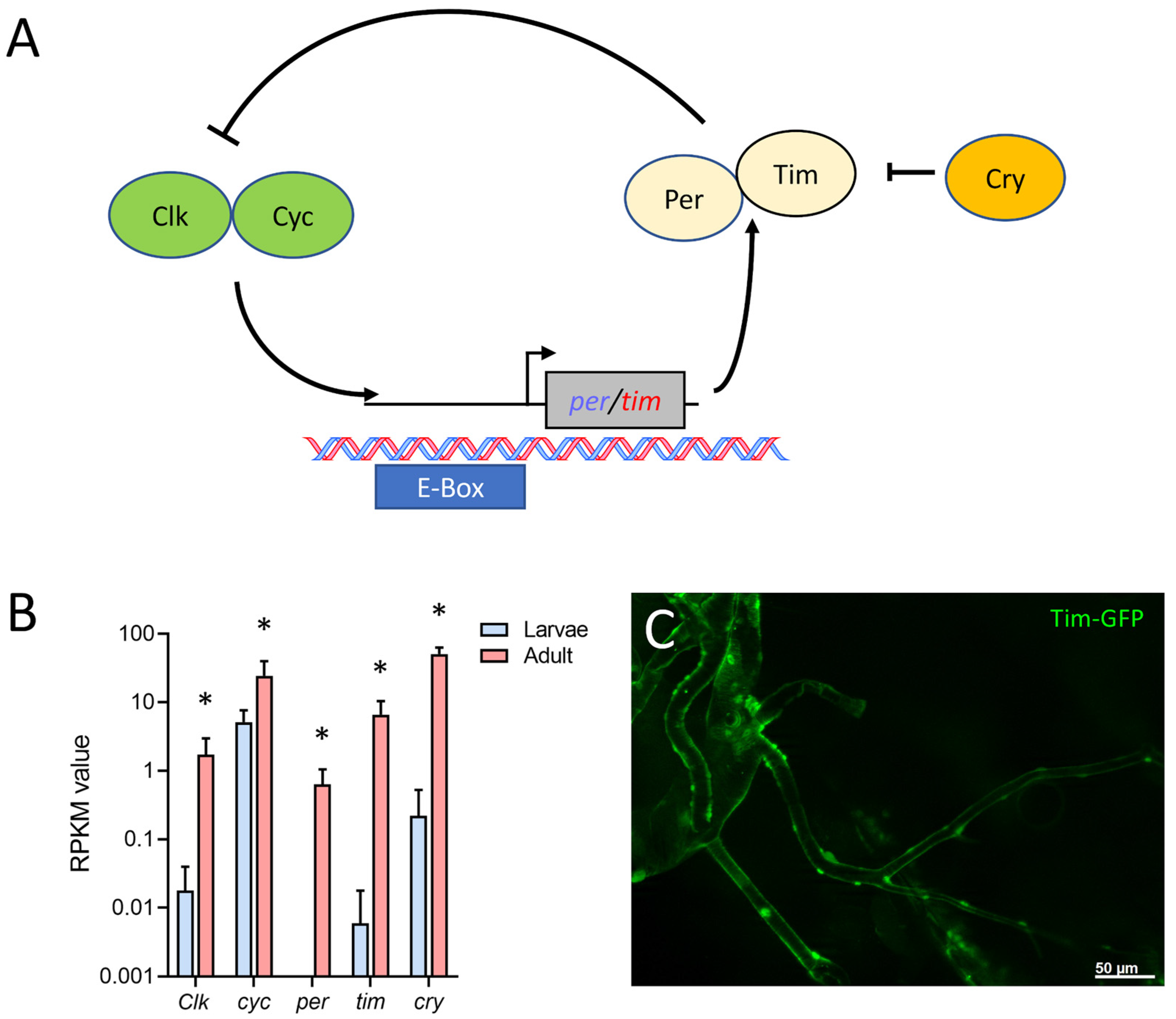
2.4. Overrepresented Processes in the Adult Tracheal System
2.5. Specific Driver Lines Targeting Cell Populations of Adult Trachea
3. Discussion
4. Materials and Methods
4.1. Fly Stocks and Husbandry
4.2. Immunohistochemistry and Fluorescence Microscopy
4.3. Dissection of Tracheae
4.4. RNA Extraction, cDNA Synthesis and PCR Amplification
4.5. Transcriptomic Analyses
4.6. Synchrotron-Radiation Based X-ray Computed Micro-Tomography
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roeder, T.; Isermann, K.; Kallsen, K.; Uliczka, K.; Wagner, C. A Drosophila Asthma Model—What the Fly Tells Us About Inflammatory Diseases of the Lung. Adv. Exp. Med. Biol. 2012, 710, 37–47. [Google Scholar] [CrossRef]
- Roeder, T.; Isermann, K.; Kabesch, M. Drosophila in asthma research. Am. J. Respir. Crit. Care Med. 2009, 179, 979–983. [Google Scholar] [CrossRef]
- Whitten, J.M. Comparative Anatomy of the Tracheal System. Annu. Rev. Èntomol. 1972, 17, 373–402. [Google Scholar] [CrossRef]
- Harrison, J.F.; Waters, J.S.; Cease, A.J.; Vandenbrooks, J.M.; Callier, V.; Klok, C.J.; Shaffer, K.; Socha, J.J. How locusts breathe. Physiology 2013, 28, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Wasserthal, L.T.; Fröhlich, A.S. Structure of the thoracic spiracular valves and their contribution to the unidirectional gas exchange in flying blowflies Calliphora vicina. J. Exp. Biol. 2017, 220 Pt 2, 208–219. [Google Scholar] [CrossRef]
- Lehmann, F.-O.; Heymann, N. Unconventional mechanisms control cyclic respiratory gas release in flying Drosophila. J. Exp. Biol. 2005, 208 Pt 19, 3645–3654. [Google Scholar] [CrossRef] [PubMed]
- Socha, J.J.; Lee, W.-K.; Harrison, J.F.; Waters, J.S.; Fezzaa, K.; Westneat, M.W. Correlated patterns of tracheal compression and convective gas exchange in a carabid beetle. J. Exp. Biol. 2008, 211 Pt 21, 3409–3420. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, E.C.; McHenry, M.J.; Bradley, T.J. Coordinated ventilation and spiracle activity produce unidirectional airflow in the hissing cockroach, Gromphadorhina portentosa. J. Exp. Biol. 2013, 216 Pt 23, 4473–4482. [Google Scholar] [CrossRef]
- Westneat, M.W.; Betz, O.; Blob, R.W.; Fezzaa, K.; Cooper, W.J.; Lee, W.-K. Tracheal Respiration in Insects Visualized with Synchrotron X-ray Imaging. Science 2003, 299, 558–560. [Google Scholar] [CrossRef]
- Ghabrial, A.; Luschnig, S.; Metzstein, M.M.; Krasnow, M.A. Branching Morphogenesis of the Drosophila Tracheal System. Annu. Rev. Cell Dev. Biol. 2003, 19, 623–647. [Google Scholar] [CrossRef]
- Samakovlis, C.; Hacohen, N.; Manning, G.; Sutherland, D.C.; Guillemin, K.; Krasnow, M.A. Development of the Drosophila tracheal system occurs by a series of morphologically distinct but genetically coupled branching events. Development 1996, 122, 1395–1407. [Google Scholar] [CrossRef]
- Hanna, L.; Popadić, A. A hemipteran insect reveals new genetic mechanisms and evolutionary insights into tracheal system development. Proc. Natl. Acad. Sci. USA 2020, 117, 4252–4261. [Google Scholar] [CrossRef]
- de Miguel, C.; Linsler, F.; Casanova, J.; Franch-Marro, X. Genetic basis for the evolution of organ morphogenesis. The case of spalt and cut in development of insect trachea. Development 2016, 143, 3615–3622. [Google Scholar] [CrossRef]
- Hayashi, S.; Kondo, T. Development and Function of the Drosophila Tracheal System. Genetics 2018, 209, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Kornberg, T.B. FGF Is an Essential Mitogen and Chemoattractant for the Air Sacs of the Drosophila Tracheal System. Dev. Cell 2002, 3, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Pitsouli, C.; Perrimon, N. Embryonic multipotent progenitors remodel the Drosophila airways during metamorphosis. Development 2010, 137, 3615–3624. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.R.; Lin, L.; Huang, H.; Guha, A.; Roy, S.; Kornberg, T.B. Developmental compartments in the larval trachea of Drosophila. eLife 2015, 4, e08666. [Google Scholar] [CrossRef] [PubMed]
- Djabrayan, N.J.-V.; Cruz, J.; de Miguel, C.; Franch-Marro, X.; Casanova, J. Specification of Differentiated Adult Progenitors via Inhibition of Endocycle Entry in the Drosophila Trachea. Cell Rep. 2014, 9, 859–865. [Google Scholar] [CrossRef]
- Peterson, S.J.; Krasnow, M.A. Subcellular Trafficking of FGF Controls Tracheal Invasion of Drosophila Flight Muscle. Cell 2015, 160, 313–323. [Google Scholar] [CrossRef]
- Wagner, C.; Isermann, K.; Fehrenbach, H.; Roeder, T. Molecular architecture of the fruit fly’s airway epithelial immune system. BMC Genom. 2008, 9, 446. [Google Scholar] [CrossRef]
- Faisal, M.N.; Hoffmann, J.; El-Kholy, S.; Kallsen, K.; Wagner, C.; Bruchhaus, I.; Fink, C.; Roeder, T. Transcriptional Regionalization of the Fruit Fly’s Airway Epithelium. PLoS ONE 2014, 9, e102534. [Google Scholar] [CrossRef] [PubMed]
- Leader, D.P.; Krause, S.A.; Pandit, A.; Davies, S.A.; Dow, J.A.T. FlyAtlas 2: A new version of the Drosophila melanogaster expression atlas with RNA-Seq, miRNA-Seq and sex-specific data. Nucleic Acids Res. 2018, 46, D809–D815. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.F.; Waters, J.S.; Biddulph, T.A.; Kovacevic, A.; Klok, C.J.; Socha, J.J. Developmental plasticity and stability in the tracheal networks supplying Drosophila flight muscle in response to rearing oxygen level. J. Insect Physiol. 2018, 106 Pt 3, 189–198. [Google Scholar] [CrossRef]
- Liu, L.; Johnson, W.A.; Welsh, M.J. Drosophila DEG/ENaC pickpocket genes are expressed in the tracheal system, where they may be involved in liquid clearance. Proc. Natl. Acad. Sci. USA 2003, 100, 2128–2133. [Google Scholar] [CrossRef]
- Prange, R.; Thiedmann, M.; Bhandari, A.; Mishra, N.; Sinha, A.; Häsler, R.; Rosenstiel, P.; Uliczka, K.; Wagner, C.; Yildirim, A.Ö.; et al. A Drosophila model of cigarette smoke induced COPD identifies Nrf2 signaling as an expedient target for intervention. Aging 2018, 10, 2122–2135. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Han, L.; Shi, L.; Lin, X. Trachea-Derived Dpp Controls Adult Midgut Homeostasis in Drosophila. Dev. Cell 2013, 24, 133–143. [Google Scholar] [CrossRef]
- Bosch, P.S.; Makhijani, K.; Herboso, L.; Gold, K.S.; Baginsky, R.; Woodcock, K.J.; Alexander, B.; Kukar, K.; Corcoran, S.; Jacobs, T.; et al. Adult Drosophila Lack Hematopoiesis but Rely on a Blood Cell Reservoir at the Respiratory Epithelia to Relay Infection Signals to Surrounding Tissues. Dev. Cell 2019, 51, 787–803.e5. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Janssens, J.; De Waegeneer, M.; Kolluru, S.S.; Davie, K.; Gardeux, V.; Saelens, W.; David, F.P.A.; Brbić, M.; Spanier, K.; et al. Fly Cell Atlas: A single-nucleus transcriptomic atlas of the adult fruit fly. Science 2022, 375, eabk2432. [Google Scholar] [CrossRef]
- Tang, H.; Kambris, Z.; Lemaitre, B.; Hashimoto, C. A Serpin that Regulates Immune Melanization in the Respiratory System of Drosophila. Dev. Cell 2008, 15, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Hattrup, C.L.; Gendler, S.J. Structure and Function of the Cell Surface (Tethered) Mucins. Annu. Rev. Physiol. 2008, 70, 431–457. [Google Scholar] [CrossRef]
- Ridley, C.; Thornton, D.J. Mucins: The frontline defence of the lung. Biochem. Soc. Trans. 2018, 46, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Denes, A.S.; Kanca, O.; Affolter, M. A cellular process that includes asymmetric cytokinesis remodels the dorsal tracheal branches in Drosophila larvae. Development 2015, 142, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Sauerwald, J.; Backer, W.; Matzat, T.; Schnorrer, F.; Luschnig, S. Matrix metalloproteinase 1 modulates invasive behavior of tracheal branches during entry into Drosophila flight muscles. eLife 2019, 8, e48857. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.-T.; Zirin, J.; Kanca, O.; Lin, W.-W.; Schulze, K.L.; Li-Kroeger, D.; Tao, R.; Devereaux, C.; Hu, Y.; Chung, V.; et al. A gene-specific T2A-GAL4 library for Drosophila. eLife 2018, 7, e35574. [Google Scholar] [CrossRef]
- Raś, M.; Iwan, D.; Kamiński, M.J. The tracheal system in post-embryonic development of holometabolous insects: A case study using the mealworm beetle. J. Anat. 2018, 232, 997–1015. [Google Scholar] [CrossRef]
- Hauser, F.; Cazzamali, G.; Williamson, M.; Blenau, W.; Grimmelikhuijzen, C.J. A review of neurohormone GPCRs present in the fruitfly Drosophila melanogaster and the honey bee Apis mellifera. Prog. Neurobiol. 2006, 80, 1–19. [Google Scholar] [CrossRef]
- Li, Q.; DeBeaubien, N.A.; Sokabe, T.; Montell, C. Temperature and Sweet Taste Integration in Drosophila. Curr. Biol. 2020, 30, 2051–2067.e5. [Google Scholar] [CrossRef]
- Wagner, C.; Isermann, K.; Roeder, T. Infection induces a survival program and local remodeling in the airway epithelium of the fly. FASEB J. 2009, 23, 2045–2054. [Google Scholar] [CrossRef]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef]
- Wagner, C.; Uliczka, K.; Bossen, J.; Niu, X.; Fink, C.; Thiedmann, M.; Knop, M.; Vock, C.; Abdelsadik, A.; Zissler, U.M.; et al. Constitutive immune activity promotes JNK- and FoxO-dependent remodeling of Drosophila airways. Cell Rep. 2021, 35, 108956. [Google Scholar] [CrossRef]
- Tang, H. Regulation and function of the melanization reaction in Drosophila. Fly 2009, 3, 105–111. [Google Scholar] [CrossRef]
- Dudzic, J.P.; Hanson, M.A.; Iatsenko, I.; Kondo, S.; Lemaitre, B. More Than Black or White: Melanization and Toll Share Regulatory Serine Proteases in Drosophila. Cell Rep. 2019, 27, 1050–1061.e3. [Google Scholar] [CrossRef]
- Iatsenko, I.; Kondo, S.; Mengin-Lecreulx, D.; Lemaitre, B. PGRP-SD, an Extracellular Pattern-Recognition Receptor, Enhances Peptidoglycan-Mediated Activation of the Drosophila Imd Pathway. Immunity 2016, 45, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Akhouayri, I.; Turc, C.; Royet, J.; Charroux, B. Toll-8/Tollo negatively regulates antimicrobial response in the Drosophila respiratory epithelium. PLoS Pathog. 2011, 7, e1002319. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.C. Mucus and mucins in diseases of the intestinal and respiratory tracts. J. Intern. Med. 2019, 285, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Syed, Z.A.; Härd, T.; Uv, A.; Van Dijk-Härd, I.F. A Potential Role for Drosophila Mucins in Development and Physiology. PLoS ONE 2008, 3, e3041. [Google Scholar] [CrossRef]
- Bermbach, S.; Weinhold, K.; Roeder, T.; Petersen, F.; Kugler, C.; Goldmann, T.; Rupp, J.; König, P. Mechanisms of Cilia-Driven Transport in the Airways in the Absence of Mucus. Am. J. Respir. Cell Mol. Biol. 2014, 51, 56–67. [Google Scholar] [CrossRef]
- Kim, K.; Lane, E.A.; Saftien, A.; Wang, H.; Xu, Y.; Wirtz-Peitz, F.; Perrimon, N. Drosophila as a model for studying cystic fibrosis pathophysiology of the gastrointestinal system. Proc. Natl. Acad. Sci. USA 2020, 117, 10357–10367. [Google Scholar] [CrossRef]
- Kanamori, Y.; Saito, A.; Hagiwara-Komoda, Y.; Tanaka, D.; Mitsumasu, K.; Kikuta, S.; Watanabe, M.; Cornette, R.; Kikawada, T.; Okuda, T. The trehalose transporter 1 gene sequence is conserved in insects and encodes proteins with different kinetic properties involved in trehalose import into peripheral tissues. Insect Biochem. Mol. Biol. 2010, 40, 30–37. [Google Scholar] [CrossRef]
- Volkenhoff, A.; Weiler, A.; Letzel, M.; Stehling, M.; Klämbt, C.; Schirmeier, S. Glial Glycolysis Is Essential for Neuronal Survival in Drosophila. Cell Metab. 2015, 22, 437–447. [Google Scholar] [CrossRef]
- Inomata, N.; Takahasi, K.R.; Koga, N. Association between duplicated maltase genes and the transcriptional regulation for the carbohydrate changes in Drosophila melanogaster. Gene 2019, 686, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Buch, S.; Melcher, C.; Bauer, M.; Katzenberger, J.; Pankratz, M.J. Opposing Effects of Dietary Protein and Sugar Regulate a Transcriptional Target of Drosophila Insulin-like Peptide Signaling. Cell Metab. 2008, 7, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Hasygar, K.; Deniz, O.; Liu, Y.; Gullmets, J.; Hynynen, R.; Ruhanen, H.; Kokki, K.; Käkelä, R.; Hietakangas, V. Coordinated control of adiposity and growth by anti-anabolic kinase ERK7. EMBO Rep. 2021, 22, e49602. [Google Scholar] [CrossRef] [PubMed]
- Grönke, S.; Müller, G.; Hirsch, J.; Fellert, S.; Andreou, A.; Haase, T.; Jäckle, H.; Kühnlein, R.P. Dual Lipolytic Control of Body Fat Storage and Mobilization in Drosophila. PLOS Biol. 2007, 5, e137. [Google Scholar] [CrossRef] [PubMed]
- Vihervaara, T.; Puig, O. dFOXO Regulates Transcription of a Drosophila Acid Lipase. J. Mol. Biol. 2008, 376, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Beenakkers, A.; Van der Horst, D.; Van Marrewijk, W. Insect flight muscle metabolism. Insect Biochem. 1984, 14, 243–260. [Google Scholar] [CrossRef]
- Selcho, M.; Millán, C.; Palacios-Muñoz, A.; Ruf, F.; Ubillo, L.; Chen, J.; Bergmann, G.; Ito, C.; Silva, V.; Wegener, C.; et al. Central and peripheral clocks are coupled by a neuropeptide pathway in Drosophila. Nat. Commun. 2017, 8, 15563. [Google Scholar] [CrossRef]
- Ito, C.; Etomioka, K. Heterogeneity of the Peripheral Circadian Systems in Drosophila melanogaster: A Review. Front. Physiol. 2016, 7, 8. [Google Scholar] [CrossRef]
- Parasram, K.; Bernardon, N.; Hammoud, M.; Chang, H.; He, L.; Perrimon, N.; Karpowicz, P. Intestinal Stem Cells Exhibit Conditional Circadian Clock Function. Stem Cell Rep. 2018, 11, 1287–1301. [Google Scholar] [CrossRef]
- Sundar, I.K.; Yao, H.; Sellix, M.T.; Rahman, I. Circadian Clock–Coupled Lung Cellular and Molecular Functions in Chronic Airway Diseases. Am. J. Respir. Cell Mol. Biol. 2015, 53, 285–290. [Google Scholar] [CrossRef]
- Sundar, I.K.; Yao, H.; Sellix, M.T.; Rahman, I. Circadian molecular clock in lung pathophysiology. Am. J. Physiol. Cell. Mol. Physiol. 2015, 309, L1056–L1075. [Google Scholar] [CrossRef] [PubMed]
- Schramm, G.; Bruchhaus, I.; Roeder, T. A simple and reliable 5′-RACE approach. Nucleic Acids Res. 2000, 28, e96. [Google Scholar] [CrossRef] [PubMed]
- Franz, O.; Bruchhaus, I.; Roeder, T. Verification of differential gene transcription using virtual northern blotting. Nucleic Acids Res. 1999, 27, e3. [Google Scholar] [CrossRef]
- Picelli, S.; Faridani, O.R.; Björklund, K.; Winberg, G.; Sagasser, S.; Sandberg, R. Full-length RNA-seq from single cells using Smart-seq2. Nat. Protoc. 2014, 9, 171–181. [Google Scholar] [CrossRef]
- Stephano, F.; Nolte, S.; Hoffmann, J.; El-Kholy, S.; von Frieling, J.; Bruchhaus, I.; Fink, C.; Roeder, T. Impaired Wnt signaling in dopamine containing neurons is associated with pathogenesis in a rotenone triggered Drosophila Parkinson’s disease model. Sci. Rep. 2018, 8, 2372. [Google Scholar] [CrossRef] [PubMed]
- Hagemann-Jensen, M.; Ziegenhain, C.; Chen, P.; Ramsköld, D.; Hendriks, G.-J.; Larsson, A.J.M.; Faridani, O.R.; Sandberg, R. Single-cell RNA counting at allele and isoform resolution using Smart-seq3. Nat. Biotechnol. 2020, 38, 708–714. [Google Scholar] [CrossRef]
- Wilde, F.; Ogurreck, M.; Greving, I.; Hammel, J.U.; Beckmann, F.; Hipp, A.; Lottermoser, L.; Khokhriakov, I.; Lytaev, P.; Dose, T.; et al. Micro-CT at the imaging beamline P05 at PETRA III. In AIP Conference Proceedings; AIP Publishing LLC.: Melville, NY, USA, 2016; Volume 1741, p. 030035. [Google Scholar] [CrossRef]
- Moosmann, J.; Ershov, A.; Weinhardt, V.; Baumbach, T.; Prasad, M.S.; LaBonne, C.; Xiao, X.; Kashef, J.; Hofmann, R. Time-lapse X-ray phase-contrast microtomography for in vivo imaging and analysis of morphogenesis. Nat. Protoc. 2014, 9, 294–304. [Google Scholar] [CrossRef]
- van Aarle, W.; Palenstijn, W.J.; De Beenhouwer, J.; Altantzis, T.; Bals, S.; Batenburg, K.J.; Sijbers, J. The ASTRA Toolbox: A platform for advanced algorithm development in electron tomography. Ultramicroscopy 2015, 157, 35–47. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bossen, J.; Prange, R.; Kühle, J.-P.; Künzel, S.; Niu, X.; Hammel, J.U.; Krieger, L.; Knop, M.; Ehrhardt, B.; Uliczka, K.; et al. Adult and Larval Tracheal Systems Exhibit Different Molecular Architectures in Drosophila. Int. J. Mol. Sci. 2023, 24, 5628. https://doi.org/10.3390/ijms24065628
Bossen J, Prange R, Kühle J-P, Künzel S, Niu X, Hammel JU, Krieger L, Knop M, Ehrhardt B, Uliczka K, et al. Adult and Larval Tracheal Systems Exhibit Different Molecular Architectures in Drosophila. International Journal of Molecular Sciences. 2023; 24(6):5628. https://doi.org/10.3390/ijms24065628
Chicago/Turabian StyleBossen, Judith, Ruben Prange, Jan-Philip Kühle, Sven Künzel, Xiao Niu, Jörg U. Hammel, Laura Krieger, Mirjam Knop, Birte Ehrhardt, Karin Uliczka, and et al. 2023. "Adult and Larval Tracheal Systems Exhibit Different Molecular Architectures in Drosophila" International Journal of Molecular Sciences 24, no. 6: 5628. https://doi.org/10.3390/ijms24065628
APA StyleBossen, J., Prange, R., Kühle, J. -P., Künzel, S., Niu, X., Hammel, J. U., Krieger, L., Knop, M., Ehrhardt, B., Uliczka, K., Krauss-Etschmann, S., & Roeder, T. (2023). Adult and Larval Tracheal Systems Exhibit Different Molecular Architectures in Drosophila. International Journal of Molecular Sciences, 24(6), 5628. https://doi.org/10.3390/ijms24065628