

Transcriptomic and Bioinformatic Analyses Identifying a Central Mif-Cop9-Nf-kB Signaling Network in Innate Immunity Response of Ciona robusta

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Results
2.1. Transcriptomic Analysis of C. robusta Pharynx
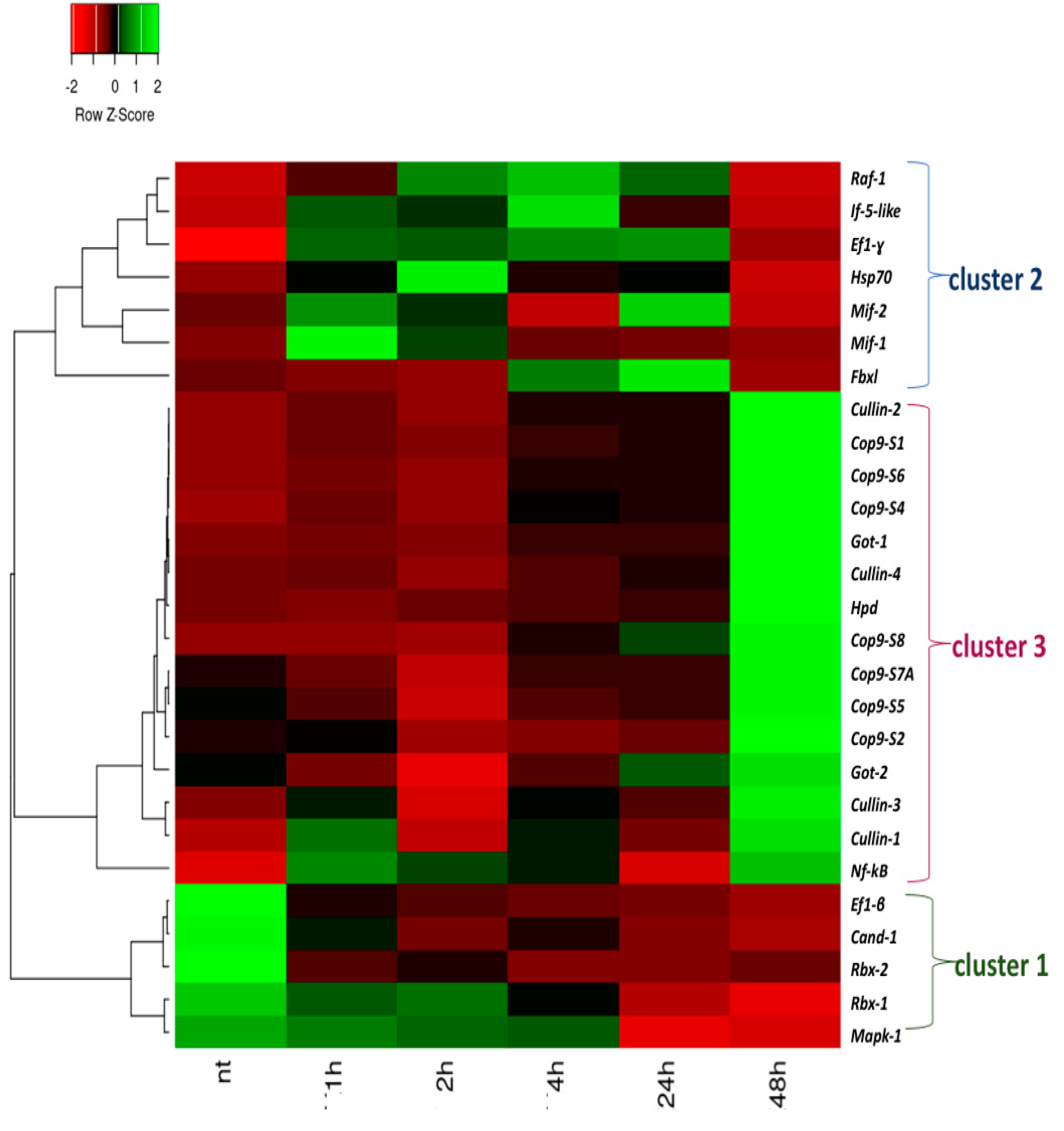
2.2. Analyses of the Expression of the Mif-Csn-Nf-κB Signaling Network Genes under LPS Exposure
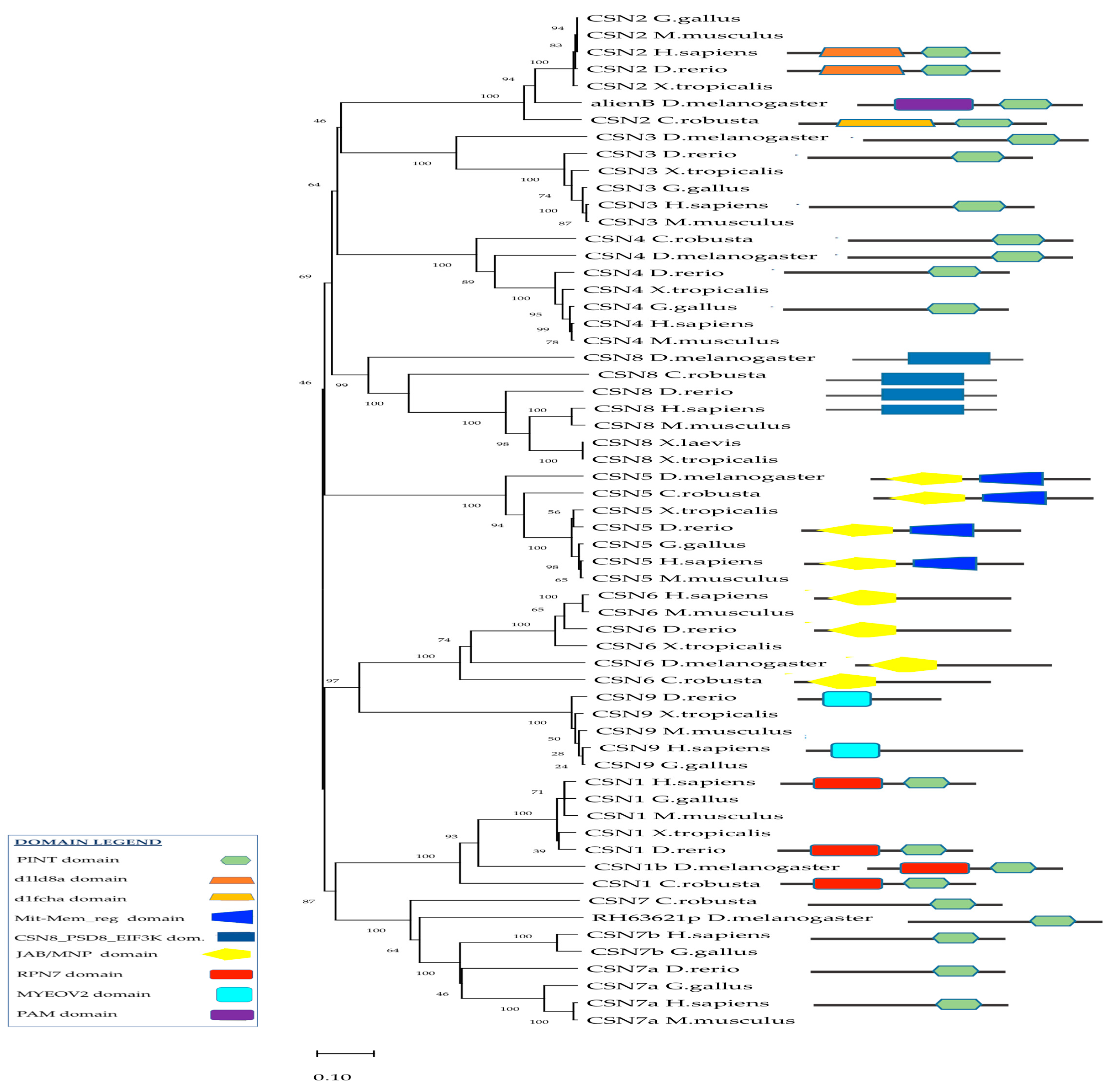
2.3. Phylogenetic Analysis of CSN and In-Silico Analyses of Csn Interactors
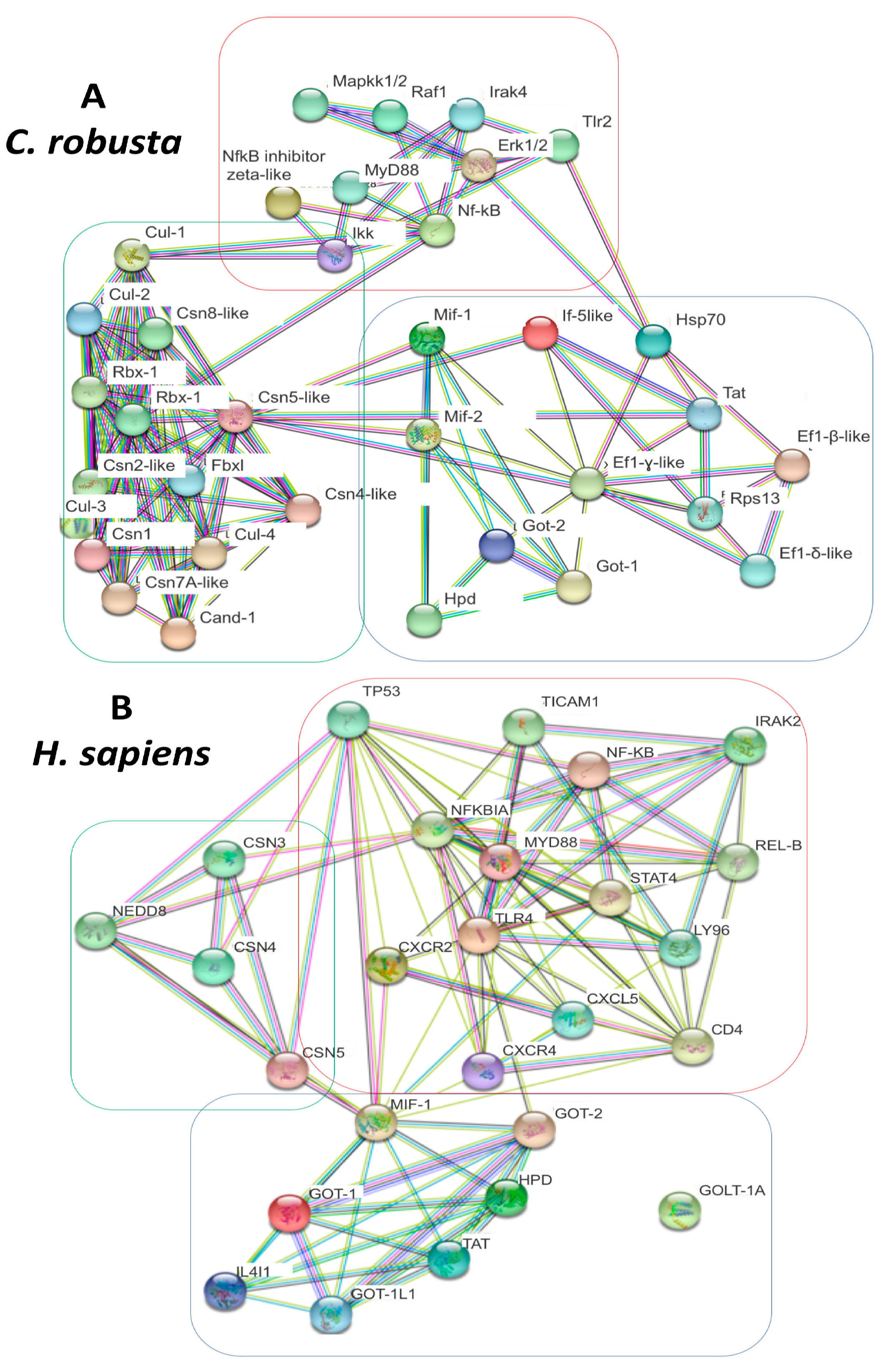
2.4. PPI analysis of C. robusta and H. sapiens Mif-Cop9-Nf-kB Signalosome Network
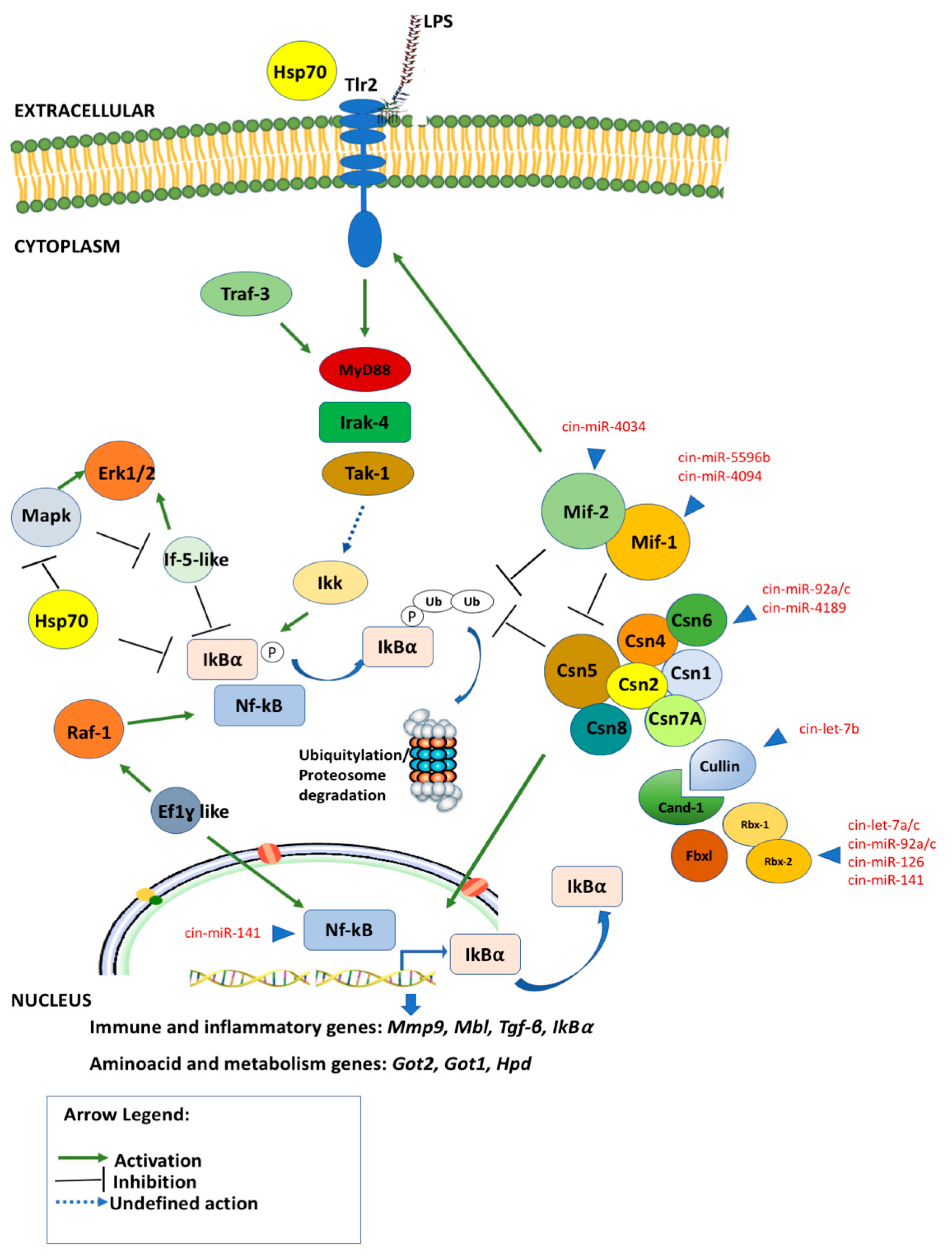
2.5. Post-Transcriptional Regulation of Mif-Cop9 Signalosome-Nf-kB Network
2.6. Network Reconstruction of Post-Transcriptional Regulation of Mif-Cop9 Signalosome-Nf-kB Network and Target Genes Interplay
3. Discussion
4. Materials and Methods
4.1. Tunicates and LPS Injection
4.2. Next-Generation Sequencing
4.3. qRT-PCR
4.4. Phylogenetic Analyses of Csn Complex and In-Silico Analysis of Csn Partners
4.5. STRING Analysis of C. robusta and H. sapiens Mif-Cop9 Interaction Network
4.6. miRNA-Target Prediction of Mif-Csn-Nf-kB Network
4.7. miRNA Analysis of Their Evolution Pattern
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bucala, R. Signal transduction. A most interesting factor. Nature 2000, 408, 146–147. [Google Scholar] [CrossRef]
- Flaster, H.; Bernhagen, J.; Calandra, T.; Bucala, R. The macrophage migration inhibitory factor-glucocorticoid dyad: Regulation of inflammation and immunity. Mol. Endocrinol. 2007, 21, 1267–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozza, M.T.; Martins, Y.C.; Carneiro, L.A.; Paiva, C.N. Macrophage migration inhibitory factor in protozoan infections. J. Parasitol. Res. 2012, 2012, 413052. [Google Scholar] [CrossRef] [Green Version]
- Leng, L.; Bucala, R. Insight into the biology of macrophage migration inhibitory factor (MIF) revealed by the cloning of its cell surface receptor. Cell Res. 2006, 16, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Sparkesa, A.; De Baetseliera, P.; Roelantsc, K.; De Treza, C.; Mageza, S.; Van Ginderachtera, J.A.; Raesa, G.; Bucalaf, R.; Stijlemansa, B. The non-mammalian MIF superfamily. Immunobiology 2017, 222, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Parisi, M.G.; Di Falco, F.; Cardinale, L.; Cammarata, M.; Arizza, V. Identification of CPE and GAIT elements in 3′UTR of macrophage migration inhibitory factor (MIF) involved in the inflammatory response induced by LPS in Ciona robusta. Mol. Immunol. 2018, 99, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsagkogeorga, G.; Turon, X.; Hopcroft, R.R.; Tilak, M.K.; Feldstein, T.; Shenkar, N.; Loya, Y.; Huchon, D.; Douzery, E.J.P.; Delsuc, F. An updated 18S rRNA phylogeny of tunicates based on mixture and secondary structure models. BMC Evol. Biol. 2009, 9, 187. [Google Scholar] [CrossRef] [Green Version]
- Zeng, L.; Swalla, B.J. Molecular phylogeny of the protochordates: Chordate evolution. Can. J. Zool. 2005, 83, 24–33. [Google Scholar] [CrossRef]
- Satoh, N. The ascidian tadpole larva: Comparative molecular development and genomics. Nat. Rev. Genet. 2003, 4, 285–295. [Google Scholar] [CrossRef]
- Vizzini, A. Gene expression and regulation of molecules involved in pharynx inflammatory response induced by LPS in Ciona intestinalis. Invertebr. Surviv. J. 2017, 14, 119–128. [Google Scholar]
- Vizzini, A.; Dumas, F.; Di Falco, F.; Arizza, V. Evolutionary and transcriptional analyses of a pentraxin-like component family involved in the LPS inflammatory response of Ciona robusta. Fish Shellfish Immunol. 2021, 111, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Arizza, V.; Bonura, A.; La Paglia, L.; Urso, A.; Pinsino, A.; Vizzini, A. Transcriptional and in silico analyses of MIF cytokine and TLR signalling interplay in the LPS inflammatory response of Ciona robusta. Sci. Rep. 2020, 10, 11339. [Google Scholar] [CrossRef] [PubMed]
- Bech-Otschir, D.; Kraft, R.; Huang, X.; Henklein, P.; Kapelari, B.; Pollmann, C.; Dubiel, W. COP9 signalosome-specific phosphorylation targets p53 to degradation by the ubiquitin system. EMBO J. 2001, 20, 1630–1639. [Google Scholar] [CrossRef] [Green Version]
- Lue, H.; Kleemann, R.; Calandra, T.; Roger, T.; Bernhagen, J. Macrophage migration inhibitory factor (MIF): Mechanisms of action and role in disease. Microbes Infect. 2002, 4, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Xu, D.; Li, J.; Deng, X.W. COP9 signalosome: Discovery, conservation, activity, and function. JIPB 2020, 62, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Dubiel, W.; Chaithongyot, S.; Dubiel, D.; Naumann, M. The COP9 Signalosome: A Multi-DUB Complex. Biomolecule 2020, 10, 1082. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, K.; Naumann, M. Control of NF-kappaB activation by the COP9 signalosome. Biochem. Soc. Trans. 2010, 38, 156–161. [Google Scholar] [CrossRef]
- Milic, J.; Tian, Y.; Bernhagen, J. Role of the COP9 Signalosome (CSN) in Cardiovascular Diseases. Biomolecules 2019, 9, 217. [Google Scholar] [CrossRef] [Green Version]
- Jumpertz, S.; Bernhagen, J.; Schütz, A.K. Role of the COP9 Signalosome in Gastrointestinal Cancers. J. Carcinog. Mutagen. 2015, 6, 1. [Google Scholar]
- Schweitzer, K.; Bozko, P.M.; Dubiel, W.; Naumann, M. CSN controls NF-κB by deubiquitinylation of IκBα. EMBO J. 2007, 26, 1532–1541. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A.; Bonura, A.; La Paglia, L.; Fiannaca, A.; La Rosa, M.; Urso, A.; Arizza, V. ceRNA Network Regulation of TGF-β, WNT, FOXO, Hedgehog Pathways in the Pharynx of Ciona robusta. Int. J. Mol. Sci. 2021, 22, 3497. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Long, L.; Luo, Z.; Li, L.; You, J. Anti-inflammatory role of microRNA let-7c in LPS treated alveolar macrophages by targeting STAT3. Asian Pac. J. Trop. Med. 2016, 9, 1–72. [Google Scholar] [CrossRef] [Green Version]
- Teng, G.; Wang, W.; Dai, Y.; Wang, S.; Chu, Y.; Li, J. Let-7b Is Involved in the Inflammation and Immune Responses Associated with Helicobacter pylor Infection by Targeting Toll-Like Receptor 4. PLoS ONE 2013, 8, 2. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Zhang, T.; Wu, H.; Jiang, K.; Qiu, C.; Deng, G. MicroRNA let-7c Improves LPS-Induced Outcomes of Endometritis by Suppressing NF-κB Signaling. Inflammation 2019, 42, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Lai, L.; Song, Y.; Liu, Y.; Chen, Q.; Han, Q.; Chen, W.; Pan, T.; Zhang, Y.; Cao, X.; Wang, Q. MicroRNA-92a Negatively Regulates Toll-like Receptor (TLR)-triggered Inflammatory Response in Macrophages by Targeting MKK4 Kinase. J. Biol. Chem. 2013, 288, 7956–7967. [Google Scholar] [CrossRef] [Green Version]
- Poissonnier, L.; Villain, G.; Sonci, F.; Mattot, V. miR126-5p repression of ALCAM and SetD5 in endothelial cells regulates leucocyte adhesion and transmigration. Cardiovasc. Res. 2014, 102, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Wang, Y. miR-141 is negatively correlated with TLR4 in neonatal sepsis and regulates LPS-induced inflammatory responses in monocytes. Braz. J. Med. Biol. Res. 2021, 54, 7. [Google Scholar]
- Kim, M.J.; Kim, W.S.; Kim, D.O.; Byun, J.E.; Huy, H.; Lee, S.Y.; Song, H.Y.; Park, Y.J.; Kim, T.D.; Yoon, S.R.; et al. Macrophage migration inhibitory factor interacts with thioredoxin–interacting protein and induces NF-κB activity. Cell. Signal. 2017, 34, 110–120. [Google Scholar] [CrossRef]
- Kapurniotu, A.; Gokce, O.; Bernhagen, J. The multitasking potential of alarmins and atypical chemokines. Front. Med. 2019, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Kato, J.; Yoneda-Kato, N. Mammalian COP9 signalosome. Genes Cells 2009, 14, 1209–1225. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal. Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Beinke, S.; Ley, S.C. Functions of NF-kappaB1 and NF-kappaB2 in immune cell biology. Biochem. J. 2004, 382, 393–409. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Q.; Wawrzyniuk, M.; Rutten, V.P.M.G.; Van Eden, W.; Sijts, A.J.A.M.; Broere, F. Hsp70 and NF-kB Mediated Control of Innate Inflammatory Responses in a Canine Macrophage. Cell Line IJMS 2020, 21, 6464. [Google Scholar] [CrossRef]
- Köller, M.; Hensler, T.; König, B.; Prévost, G.; Alouf, J.; Konig, W. Induction of heat-shock proteins by bacterial toxins, lipid mediators and cytokines in human leukocytes. Zent. Bakteriol. 1993, 278, 365–376. [Google Scholar] [CrossRef]
- Teshima, S.; Rokutan, K.; Takahashi, M.; Nikawa, T.; Kishi, K. Induction of heat shock proteins and their possible roles in macrophages during activation by macrophage colony-stimulating factor. Biochem. J. 1996, 315, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Zininga, T.; Ramatsui, L.; Shonhai, A. Heat Shock Proteins as Immunomodulants. Molecules 2018, 23, 2846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukaj, S. Heat Shock Protein 70 as a Double Agent Acting Inside and Outside the Cell: Insights into Autoimmunity. Int. J. Mol. Sci. 2020, 21, 5298. [Google Scholar] [CrossRef] [PubMed]
- Ferat-Osorio, E.; Sánchez-Anaya, A.; Gutiérrez-Mendoza, M.; Boscó-Gárate, I.; Wong-Baeza, I.; Pastelin-Palacios, R.; Pedraza-Alva, G.; Bonifaz, L.C.; Cortés-Reynosa, P.; Pérez-Salazar, E.; et al. Heat shock protein 70 down-regulates the production of toll-like receptor-induced pro-inflammatory cytokines by a heat shock factor-1/constitutive heat shock element-binding factor-dependent mechanism. J. Inflamm. 2014, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Cope, G.A.; Deshaies, R.J. COP9 signalosome: A multifunctional regulator of SCF and other cullin-based ubiquitin ligases. Cell 2003, 114, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Chamovitz, D.A. Revisiting the COP9 signalosome as a transcriptional regulator. Embo. Rep. 2009, 10, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Le Floc’h, N.; Melchior, D.; Obled, C. Modifications of protein and amino acid metabolism during inflammation and immune system activation, Metab. Aminoacidic. Science 2004, 87, 37–45. [Google Scholar]
- Gringhuis, S.I.; Den Dunnen, J.; Litjens, M.; Van het Hof, B.; Van Kooyk, Y.; Geijtenbeek, T.B.H. C-Type Lectin DC-SIGN Modulates Toll-like Receptor Signaling via Raf-1 Kinase-Dependent Acetylation of Transcription Factor NF-kB. Immun. Cell Press 2007, 26, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, N.; Sanges, C. Raf kinases in signal transduction and interaction with translation machinery. BioMol. Concepts 2013, 4, 391–399. [Google Scholar] [CrossRef]
- Schulz, I.; Engel, C.; Niestroj, A.J.; Kehlen, A.; Rahfeld, J.; Kleinschmidt, M.; Lehmann, K.; Roßner, S.; Demuth, H. A non-canonical function of eukaryotic elongation factor 1A1: Regulation of interleukin-6 expression. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2014, 1843, 965–975. [Google Scholar] [CrossRef]
- Moore, C.C.; Martin, E.N.; Lee, G.; Taylor, C.; Dondero, R.; Reznikov, L.L.; Dinarello, C.; Thompson, J.; Scheld, W.M. Eukaryotic translation initiation factor 5A small interference RNA-liposome complexes reduce inflammation and increase survival in murine models of severe sepsis and acute lung injury. J. Infect. Dis. 2018, 198, 1407–1414. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.A.; Liu, Z.; Tang, T.C.; Zheng, Q.; Francis, S.; Wang, T.; Ye, B.; Lust, J.A.; Dondero, R.; Thompson, J.E. Modulation of eIF5A Expression Using SNS01 Nanoparticles Inhibits NF-KB Activity and Tumor Growth in Murine Models of Multiple Myeloma. Mol. Ther. 2012, 20, 1305–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiannaca, A.; La Rosa, M.; La Paglia, L.; Rizzo, R.; Urso, A. MiRNATIP: A SOM-based miRNA-target interactions predictor. BMC Bioinform. 2016, 17, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vizzini, A.; Bonura, A.; La Paglia, L.; Fiannaca, A.; La Rosa, M.; Urso, A.; Mauro, M.; Vazzana, M.; Arizza, V. Transcriptomic Analyses Reveal 2 and 4 Family Members of Cytochromes P450 (CYP) Involved in LPS Inflammatory Response in Pharynx of Ciona robusta. Int. J. Mol. Sci. 2021, 22, 11141. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Ge, J.; Wang, Z.; Ren, J.; Wang, X.; Xiong, H.; Gao, J.; Zhang, Y.; Zhang, Q. Let-7e modulates the inflammatory response in vascular endothelial cells through ceRNA crosstalk. Sci. Rep. 2017, 7, 42498. [Google Scholar] [CrossRef]
- Sun, H.; Tian, J.; Li, J. MiR-92b-3p ameliorates inflammation and autophagy by targeting TRAF3 and suppressing MKK3-p38 pathway in caerulein-induced AR42J cells. Int. Immunopharmacol. 2020, 88, 106691. [Google Scholar] [CrossRef] [PubMed]
- Krasowski, M.D.; Reschly, E.J.; Ekins, S. Intrinsic disorder in nuclear hormone receptors. J. Proteome Res. 2008, 7, 4359–4372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, R.; Gissi, C.; Pennati, R.; Manni, L. Morphological evidence that the molecularly determined Ciona intestinalis type A and type B are different species: Ciona robusta and Ciona intestinalis. J. Zool. Syst. Evol. Res. 2015, 53, 186–193. [Google Scholar] [CrossRef]
- Pennati, R.; Ficetola, G.F.; Brunetti, R.; Caicci, F.; Gasparini, F.; Griggio, F.; Sato, A.; Stach, T.; Kaul-Strehlow, S.; Gissi, C.; et al. Morphological differences between larvae of the Ciona intestinalis species complex: Hints for a valid taxonomic definition of distinct species. PLoS ONE 2015, 10, e0122879. [Google Scholar] [CrossRef] [Green Version]
- Caputi, L.; Andreakis, N.; Mastrototaro, F.; Cirino, P.; Vassillo, M.; Sordino, P. Cryptic speciation in a model invertebrate chordate. Proc. Natl. Acad. Sci. USA 2007, 104, 9364–9369. [Google Scholar] [CrossRef] [Green Version]
- Iannelli, F.; Pesole, G.; Sordino, P.; Gissi, C. Mitogenomics reveals two cryptic species in Ciona intestinalis. Trends Genet. 2007, 23, 419–422. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S. HISAT: A fast spliced aligner with low memory requirements. Nat Methods. 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Falco, F.D.; Parrinello, D.; Sanfratello, M.A.; Mazzarella, C.; Parrinello, N.; Cammarata, M. Ciona intestinalis interleukin17-like genes expression is upregulated by LPS challenge. Dev. Comp. Immunol. 2015, 48, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, A.; Di Falco, F.; Parrinello, D.; Sanfratello, M.A.; Cammarata, M. Transforming growth factor b (CiTGF-b) gene expression is induced in the inflammatory reaction of Ciona intestinalis. Dev. Comp. Immunol. 2016, 55, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Yu, X.; Wu, C.; Zhu, T.; Wang, W.; Zheng, X.; Jin, H. RNA sequencing-based longitudinal transcriptomic profiling gives novel insights into the disease mechanism of generalized pustular psoriasis. BMC Med. Genom. 2018, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 97–166. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ensemble ID | Gene Name | Log Fold Change | Down (-)/up (+) Regulation | p-Value |
|---|---|---|---|---|
| ENSCING00000021369 | Macrophage migration inhibitory factor-1 (Mif-1) | 1.337426682 | + | 0.005771886 |
| ENSCING00000006933 | Nuclear factor kappa-light-chain-enhancer of activated B cells (Nf-kB) | −1.171509012 | - | 0.011439532 |
| ENSCING00000009525 | Eukaryotic translation initiation factor 5-like (If5-like) | −1.041407587 | - | 0.024225277 |
| ENSCING00000020171 | Complement component C6 | −1.31525517 | - | 0.004785769 |
| ENSCING00000006967 | Apoptosis-inducing factor 2 | −4.983320748 | - | 5.75 × 10−20 |
| ENSCING00000005269 | Ci-META4 (meta-4) | −4.382519803 | - | 1.97 × 10−16 |
| ENSCING00000014006 | Collectin-46-like | −3.936841979 | - | 3.62 × 10−9 |
| ENSCING00000005686 | Complement component C6-like | −3.712293984 | - | 3.00 × 10−13 |
| ENSCING00000013919 | Cytochrome P450 2C13, male-specific-like | −3.458407214 | - | 6.28 × 10−12 |
| ENSCING00000001811 | E-selectin-like | −1.841338282 | - | 9.61 × 10−5 |
| ENSCING00000002203 | Heat shock 70 kDa protein 4L | −1.682141547 | - | 0.000361039 |
| ENSCING00000008644 | T-box transcription factor Ci-mT (mT) | 2.266940329 | + | 0.000455334 |
| ENSCING00000023704 | TNF receptor-associated factor 3-like | 2.326276709 | + | 7.85 × 10−5 |
| ENSCING00000007413 | L-rhamnose-binding lectin CSL3-like | 1.770298473 | + | 0.000345541 |
| ENSCING00000019232 | Interleukin 17-1 (il-17-1) | 1.611522116 | + | 0.000760084 |
| ENSCING00000005903 | Homeobox transcription factor Hox1 (hox1) | 1.511849854 | + | 0.002721531 |
| ENSCING00000003820 | Immunoglobulin superfamily containing leucine-rich repeat protein 2-like | 1.57489592 | + | 0.001058269 |
| ENSCING00000016336 | Integrin alpha-V like | 1.591865774 | + | 0.000862848 |
| ENSCINGENE | Gene Name | Protein ID |
|---|---|---|
| ENSCING00000015215 | 4-hydroxyphenylpyruvate dioxygenase(LOC100185199) | Hpd |
| ENSCING00000005686 | heat shock protein 70(hsp70) | Hsp70 |
| ENSCING00000013244 | Toll-like receptor 2(ci-tlr2) | Tlr2 |
| ENSCING00000017616 | myeloid differentiation primary response protein MyD88(LOC100179208) | Myd88 |
| ENSCING00000004105 | mitogen-activated protein kinase kinase(mapkk1/2) | Mapk1/2 |
| ENSCING00000007067 | mitogen-activated protein kinase(erk1/2) | Erk1/2 |
| ENSCING00000005640 | IkappaB kinase(ikk-epsilon) | Ikk-epsilon |
| ENSCING00000016280 | interleukin-1 receptor-associated kinase 1-binding protein 1-like(LOC100177072) | Irak4 |
| ENSCING00000003721 | B-raf proto-oncogene serine/threonine-protein kinase | Raf-1 |
| ENSCING00000002308 | elongation factor 1-gamma-A-like(LOC100187007) | Ef1-ɣ-A |
| ENSCING00000012434 | elongation factor 1-beta-like(LOC100176941) | Ef1-β-A |
| ENSCING00000012434 | elongation factor 1-beta/delta-like | Ef1-β/δ like |
| ENSCING00000009525 | translation initiation factor(if5-1) | If-5-like |
| ENSCING00000021423 | macrophage migration inhibitory factor-like(LOC100175627) | Mif-2 |
| ENSCING00000021369 | macrophage migration inhibitory factor-like(LOC100183461) | Mif-1 |
| ENSCING00000018967 | NF-kappa-B inhibitor zeta-like(LOC100182428) | Nf-kB inhib. zeta like |
| ENSCING00000006933 | Nf-kB protein(nfkbp105) | Nf-kB |
| ENSCING00000008364 | aspartate aminotransferase, cytoplasmic-like(LOC100184965) | Got-1 |
| ENSCING00000003669 | aspartate aminotransferase, mitochondrial(LOC100179315) | Got-2 |
| ENSCING00000006501 | cullin-1-like(LOC100177902) | Cul-1 |
| ENSCING00000002219 | cullin-2(LOC100183130) | Cul-2 |
| ENSCING00000002955 | cullin-3(LOC100177000) | Cul-3 |
| ENSCING00000001234 | cullin-4A(LOC100175879) | Cul-4A |
| ENSCING00000019895 | F-box/LRR-repeat protein 8-like(LOC100187209) | Fbxl |
| ENSCING00000007306 | cullin-associated NEDD8-dissociated protein 1(LOC100177977) | Cand-1 |
| ENSCING00000022721 | RING-box protein 1(LOC100182516) | Rbx-1 |
| ENSCING00000001372 | RING-box protein 2-like(LOC100181702) | Rbx-2 |
| ENSCING00000009475 | COP9 signalosome complex subunit 1(LOC100176942) | Csn1 |
| ENSCING00000008702 | COP9 signalosome complex subunit 2-like(LOC100177051) | Csn2 |
| ENSCING00000002866 | COP9 signalosome complex subunit 4-like(LOC100183592) | Csn4 |
| ENSCING00000006436 | COP9 signalosome complex subunit 5-like(LOC100187322) | Csn5 |
| ENSCING00000000624 | COP9 signalosome complex subunit 6(LOC100176116) | Csn6 |
| ENSCING00000021617 | COP9 signalosome complex subunit 7a-like(LOC100184522) | Csn7A-like |
| ENSCING00000011567 | COP9 signalosome complex subunit 8-like(LOC100182035) | Csn8-like |
| Protein Name | C. robusta Protein ID | H. sapiens Protein ID | Sequence Identity | Protein Domain |
|---|---|---|---|---|
| Cul-1 | F6PW28 | Q13616 | 72.9% | CULLIN-DOMAIN |
| Cul-2 | F6Z726 | Q13617 | 46.7% | CULLIN-DOMAIN |
| Cul-3 | F6R472 | Q13618 | 71.2% | CULLIN-DOMAIN |
| Cul-4A | F6SBC7 | Q13619 | 62.7% | CULLIN-DOMAIN |
| Csn-1 | F7AEY7 | Q13098 | 59.3% | PCI-DOMAIN |
| Csn-2 | F6WYI9 | P61201 | 80.9% | PCI-DOMAIN |
| Csn-4 | F6TMN3 | Q9BT78 | 68.5% | PCI-DOMAIN |
| Csn-5 | F6QLM7 | Q92905 | 80.2% | MPN-DOMAIN |
| Csn-6 | F6RRF2 | Q7L5N1 | 58.2% | MPN-DOMAIN |
| Csn7a-like | F6Z0V4 | Q9UBW8 | 45.8% | PCI-DOMAIN |
| Csn8-like | F6TBA1 | Q99627 | 35.4% | PCI-DOMAIN |
| Rbx-1 | H2XNZ7 | P62877 | 92.2% | RING-TYPE DOMAIN |
| Rbx-2 | F6SAA4 | Q9UBF6 | 82.6% | RING-TYPE DOMAIN |
| Cand-1 | F6Z3T2 | Q86VP6 | 56.3% | HEAT REPEAT |
| Fbxl | H2XKE5 | Q96CD0 | 24.6% | F-BOX DOMAIN |
| miRNA ID | Conserved miRNAs Evidenced by C. robusta NGS | Authors | Reference |
|---|---|---|---|
| let-7a | ENSCING00000017776 | Yu et al. | [23] |
| let-7b | ENSCING00000017757 | Teng et al. | [24] |
| let-7c | ENSCING00000017755 | Zhao et al. | [25] |
| miR-92a | ENSCING00000017761 | Li et al. | [26] |
| miR-92c | ENSCING00000017777 | None | None |
| miR-126 | ENSCING00000017730 | Poissonnier et al. | [27] |
| miR-141 | ENSCING00000017782 | Lin et al. | [28] |
| miRNA ID | Species-Specific miRNAs Evidenced by C. robusta NGS | miRNA ID | Species-Specific miRNAs Evidenced by C. robusta NGS |
|---|---|---|---|
| cin-miR-4011-a | ENSCING00000021193 | cin-miR-4163 | ENSCING00000020782 |
| cin-miR-4019 | ENSCING00000025043 | cin-miR-4183 | ENSCING00000021879 |
| cin-miR-4024 | ENSCING00000021882 | cin-miR-4186 | ENSCING00000020139 |
| cin-miR-4034 | ENSCING00000018691 | cin-miR-4187 | ENSCING00000024515 |
| cin-miR-4053 | ENSCING00000020571 | cin-miR-4189 | ENSCING00000018460 |
| cin-miR-4056 | ENSCING00000021074 | cin-miR-4197 | ENSCING00000023707 |
| cin-miR-4069 | ENSCING00000021942 | cin-miR-4200 | ENSCING00000021921 |
| cin-miR-4075 | ENSCING00000025167 | cin-miR-5596b | ENSCING00000020725 |
| cin-miR-4089 | ENSCING00000020134 | cin-miR-5598 | ENSCING00000021178 |
| cin-miR-4094 | ENSCING00000022222 | cin-miR-5600 | ENSCING00000017949 |
| cin-miR-4098 | ENSCING00000021981 | cin-miR-5605 | ENSCING00000023578 |
| cin-miR-4109 | ENSCING00000022889 | cin-miR-5609 | ENSCING00000024461 |
| cin-miR-4144 | ENSCING00000019473 | cin-miR-5611 | ENSCING00000023047 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Paglia, L.; Vazzana, M.; Mauro, M.; Dumas, F.; Fiannaca, A.; Urso, A.; Arizza, V.; Vizzini, A. Transcriptomic and Bioinformatic Analyses Identifying a Central Mif-Cop9-Nf-kB Signaling Network in Innate Immunity Response of Ciona robusta. Int. J. Mol. Sci. 2023, 24, 4112. https://doi.org/10.3390/ijms24044112
La Paglia L, Vazzana M, Mauro M, Dumas F, Fiannaca A, Urso A, Arizza V, Vizzini A. Transcriptomic and Bioinformatic Analyses Identifying a Central Mif-Cop9-Nf-kB Signaling Network in Innate Immunity Response of Ciona robusta. International Journal of Molecular Sciences. 2023; 24(4):4112. https://doi.org/10.3390/ijms24044112
Chicago/Turabian StyleLa Paglia, Laura, Mirella Vazzana, Manuela Mauro, Francesca Dumas, Antonino Fiannaca, Alfonso Urso, Vincenzo Arizza, and Aiti Vizzini. 2023. "Transcriptomic and Bioinformatic Analyses Identifying a Central Mif-Cop9-Nf-kB Signaling Network in Innate Immunity Response of Ciona robusta" International Journal of Molecular Sciences 24, no. 4: 4112. https://doi.org/10.3390/ijms24044112
APA StyleLa Paglia, L., Vazzana, M., Mauro, M., Dumas, F., Fiannaca, A., Urso, A., Arizza, V., & Vizzini, A. (2023). Transcriptomic and Bioinformatic Analyses Identifying a Central Mif-Cop9-Nf-kB Signaling Network in Innate Immunity Response of Ciona robusta. International Journal of Molecular Sciences, 24(4), 4112. https://doi.org/10.3390/ijms24044112