
Quercetins, Chlorogenic Acids and Their Colon Metabolites Inhibit Colon Cancer Cell Proliferation at Physiologically Relevant Concentrations




Abstract
:1. Introduction
2. Results
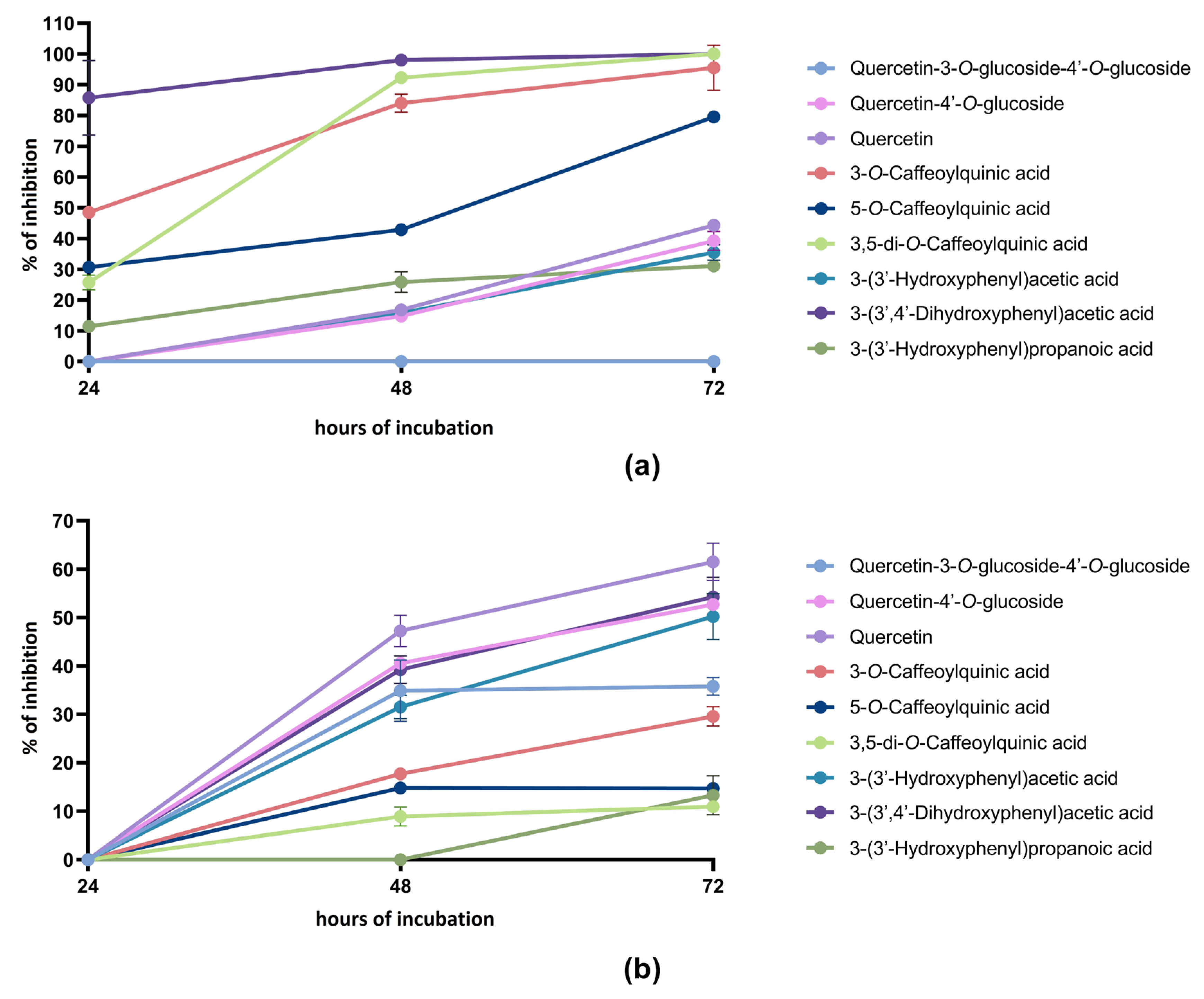
2.1. Anti-Proliferative Activity of Phenolic Compounds and Metabolites
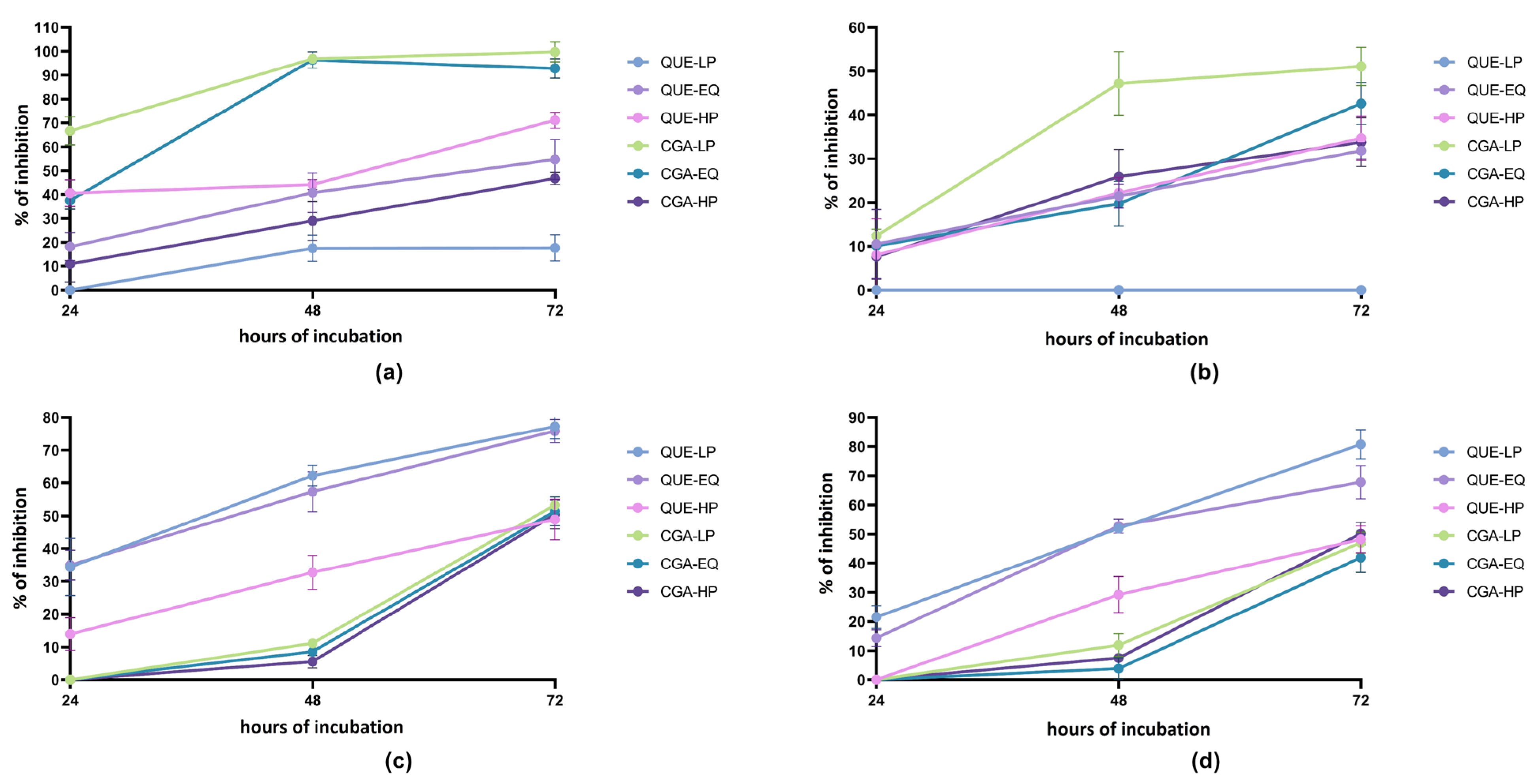
2.2. Anti-Proliferative Activity of Quercetin Derivatives/Metabolites and Chlorogenic Acid/Metabolite Mixes
2.3. Cell Cycle Analysis
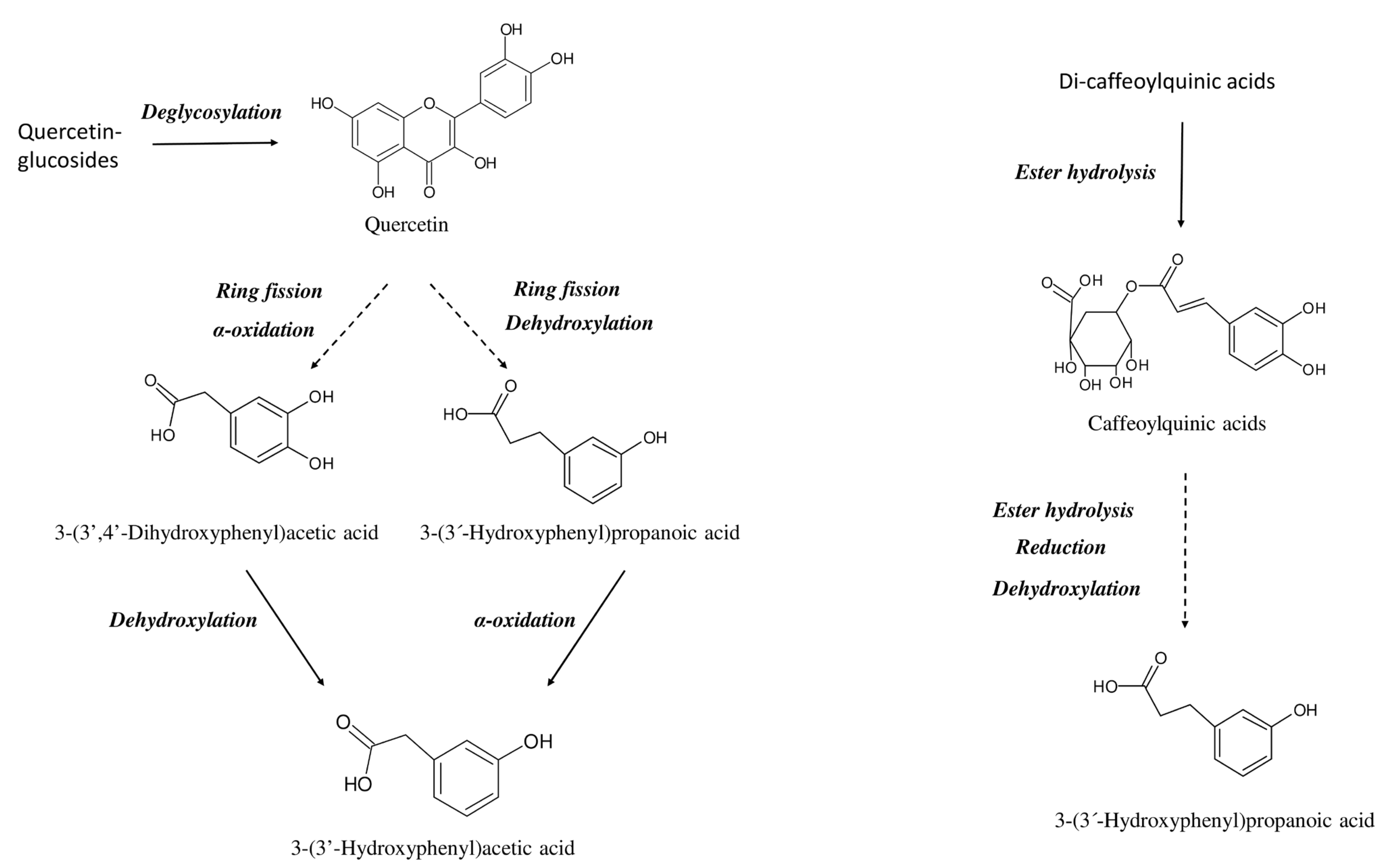
2.4. Cell Metabolism of Phenolic Compounds and Metabolites
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures, Phenolic Compound Preparation and Anti-Proliferative Assay
4.3. Cell Cycle Analysis
4.4. High-Resolution Mass Spectrometry Analysis of Cell Media
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Czene, K.; Lichtenstein, P.; Hemminki, K. Environmental and heritable causes of cancer among 9.6 million individuals in the Swedish Family-Cancer Database. Int. J. Cancer 2002, 99, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.T.; Giovannucci, E.L. Primary prevention of colorectal cancer. Gastroenterology 2010, 138, 2029–2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Wei, B.; Zhai, Z.; Zheng, Y.; Yao, J.; Wang, S.; Xiang, D.; Hu, J.; Ye, X.; Yang, S.; et al. Dietary Risk-Related Colorectal Cancer Burden: Estimates from 1990 to 2019. Front. Nutr. 2021, 8, 690663. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, N.; Bouras, E.; van den Brandt, P.A.; Muller, D.C.; Papadopoulou, A.; Heath, A.K.; Critselis, E.; Gunter, M.J.; Vineis, P.; Ferrari, P.; et al. A Prospective Diet-Wide Association Study for Risk of Colorectal Cancer in EPIC. Clin. Gastroenterol. Hepatol. 2022, 20, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, T.; Ruszkowska, M.; Danielewicz, A.; Niedźwiedzka, E.; Arłukowicz, T.; Przybyłowicz, K.E. A review of colorectal cancer in terms of epidemiology, risk factors, development, symptoms and diagnosis. Cancers 2021, 13, 2025. [Google Scholar] [CrossRef]
- Thanikachalam, K.; Khan, G. Colorectal cancer and nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef] [Green Version]
- Amintas, S.; Dupin, C.; Boutin, J.; Beaumont, P.; Moreau-Gaudry, F.; Bedel, A.; Krisa, S.; Vendrely, V.; Dabernat, S. Bioactive food components for colorectal cancer prevention and treatment: A good match. Crit. Rev. Food Sci. Nutr. 2022, in press. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Schwedhelm, C.; Hoffmann, G.; Knüppel, S.; Laure Preterre, A.; Iqbal, K.; Bechthold, A.; de Henauw, S.; Michels, N.; Devleesschauwer, B.; et al. Food groups and risk of colorectal cancer. Int. J. Cancer 2018, 142, 1748–1758. [Google Scholar] [CrossRef] [Green Version]
- Farinetti, A.; Zurlo, V.; Manenti, A.; Coppi, F.; Mattioli, A.V. Mediterranean diet and colorectal cancer: A systematic review. Nutrition 2017, 43–44, 83–88. [Google Scholar] [CrossRef]
- Finicelli, M.; Di Salle, A.; Galderisi, U.; Peluso, G. The Mediterranean Diet: An Update of the Clinical Trials. Nutrients 2022, 14, 2956. [Google Scholar] [CrossRef]
- Mahmod, A.I.; Haif, S.K.; Kamal, A.; Al-Ataby, I.A.; Talib, W.H. Chemoprevention effect of the Mediterranean diet on colorectal cancer: Current studies and future prospects. Front. Nutr. 2022, 9, 924192. [Google Scholar] [CrossRef]
- Morze, J.; Danielewicz, A.; Przybyłowicz, K.; Zeng, H.; Hoffmann, G.; Schwingshackl, L. An updated systematic review and meta-analysis on adherence to mediterranean diet and risk of cancer. Eur. J. Nutr. 2021, 60, 1561–1586. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Cayssials, V.; Jenab, M.; Rothwell, J.A.; Fedirko, V.; Aleksandrova, K.; Tjønneland, A.; Kyrø, C.; Overvad, K.; Boutron-Ruault, M.C.; et al. Dietary intake of total polyphenol and polyphenol classes and the risk of colorectal cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Eur. J. Epidemiol. 2018, 33, 1063–1075. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Lei, L.; Zhou, Y.; Ye, F.; Zhao, G. Dietary flavonoids and the risk of colorectal cancer: An updated meta-analysis of epidemiological studies. Nutrients 2018, 10, 950. [Google Scholar] [CrossRef] [Green Version]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic acids and the acyl-quinic acids: Discovery, biosynthesis, bioavailability and bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef] [Green Version]
- Bento-Silva, A.; Koistinen, V.M.; Mena, P.; Bronze, M.R.; Hanhineva, K.; Sahlstrøm, S.; Kitrytė, V.; Moco, S.; Aura, A.M. Factors affecting intake, metabolism and health benefits of phenolic acids: Do we understand individual variability? Eur. J. Nutr. 2020, 59, 1275–1293. [Google Scholar] [CrossRef] [Green Version]
- Godos, J.; Rapisarda, G.; Marventano, S.; Galvano, F.; Mistretta, A.; Grosso, G. Association between polyphenol intake and adherence to the Mediterranean diet in Sicily, southern Italy. NFS J. 2017, 8, 1–7. [Google Scholar] [CrossRef]
- Vázquez-Ruiz, Z.; Toledo, E.; Vitelli-Storelli, F.; Goni, L.; de la O, V.; Bes-Rastrollo, M.; Martínez-González, M.Á. Effect of Dietary Phenolic Compounds on Incidence of Cardiovascular Disease in the SUN Project; 10 Years of Follow-Up. Antioxidants 2022, 11, 783. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Cattivelli, A.; Tagliazucchi, D. Domestic cooking methods affect the stability and bioaccessibility of dark purple eggplant (Solanum melongena) phenolic compounds. Food Chem. 2021, 341, 128298. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Phenolic compounds profile and antioxidant properties of six sweet cherry (Prunus avium) cultivars. Food Res. Int. 2017, 97, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antiox. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattivelli, A.; Conte, A.; Martini, S.; Tagliazucchi, D. Influence of cooking methods on onion phenolic compounds bioaccessibility. Foods 2021, 10, 1023. [Google Scholar] [CrossRef]
- Coman, V.; Vodnar, D.C. Hydroxycinnamic acids and human health: Recent advances. J. Sci. Food Agric. 2020, 100, 483–499. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, T.; Chen, G.Y. Flavonoids and colorectal cancer prevention. Antioxidants 2018, 7, 187. [Google Scholar] [CrossRef] [Green Version]
- Mihanfar, A.; Darband, S.G.; Sadighparvar, S.; Kaviani, M.; Mirza-Aghazadeh-Attari, M.; Yousefi, B.; Majidinia, M. In vitro and in vivo anticancer effects of syringic acid on colorectal cancer: Possible mechanistic view. Chem. Biol. Interact. 2021, 337, 109337. [Google Scholar] [CrossRef]
- Serra, A.; MacIà, A.; Romero, M.P.; Reguant, J.; Ortega, N.; Motilva, M.J. Metabolic pathways of the colonic metabolism of flavonoids (flavonols, flavones and flavanones) and phenolic acids. Food Chem. 2012, 130, 383–393. [Google Scholar] [CrossRef]
- Cattivelli, A.; Nissen, L.; Casciano, F.; Tagliazucchi, D.; Gianotti, A. Impact of cooking methods of red-skinned onion on in vitro colon metabolic transformation of phenolic compounds and gut microbiota changes. Food Funct. 2023, 14, 3509–3525. [Google Scholar] [CrossRef]
- Juániz, I.; Ludwig, I.A.; Bresciani, L.; Dall’Asta, M.; Mena, P.; del Rio, D.; Cid, C.; de Peña, M.P. Bioaccessibility of (poly)phenolic compounds of raw and cooked cardoon (Cynara cardunculus L.) after simulated gastrointestinal digestion and fermentation by human colonic microbiota. J. Funct. Foods 2017, 32, 195–207. [Google Scholar] [CrossRef]
- Nissen, L.; Cattivelli, A.; Casciano, F.; Gianotti, A.; Tagliazucchi, D. Roasting and frying modulate the phenolic profile of dark purple eggplant and differently change the colon microbiota and phenolic metabolites after in vitro digestion and fermentation in a gut model. Food Res. Int. 2022, 160, 111702. [Google Scholar] [CrossRef]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés-Martín, A.; Selma, M.V.; Tomás-Barberán, F.A.; González-Sarrías, A.; Espín, J.C. Where to Look into the Puzzle of Polyphenols and Health? The Postbiotics and Gut Microbiota Associated with Human Metabotypes. Mol. Nutr. Food Res. 2020, 64, 1900952. [Google Scholar] [CrossRef] [PubMed]
- Iglesias-Aguirre, C.E.; Cortés-Martín, A.; Ávila-Gálvez, M.A.; Giménez-Bastida, J.A.; Selma, M.V.; González-Sarrías, A.; Espín, J.C. Main drivers of (poly)phenol effects on human health: Metabolite production and/or gut microbiota-associated metabotypes? Food Funct. 2021, 12, 10324–10355. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Fernández, M.; Young Tie Yang, P.; Ludwig, I.A.; Clifford, M.N.; Cid, C.; Rodriguez-Mateos, A. In vivo study of the bioavailability and metabolic profile of (poly)phenols after sous-vide artichoke consumption. Food Chem. 2022, 367, 130620. [Google Scholar] [CrossRef]
- Feng, S.; Yi, J.; Li, X.; Wu, X.; Zhao, Y.; Ma, Y.; Bi, J. Systematic Review of Phenolic Compounds in Apple Fruits: Compositions, Distribution, Absorption, Metabolism, and Processing Stability. J. Agric. Food Chem. 2021, 69, 7–27. [Google Scholar] [CrossRef]
- Hai, Y.; Zhang, Y.; Liang, Y.; Ma, X.; Qi, X.; Xiao, J.; Xue, W.; Luo, Y.; Yue, T. Advance on the absorption, metabolism, and efficacy exertion of quercetin and its important derivatives. Food Front. 2020, 1, 420–434. [Google Scholar] [CrossRef]
- Gómez-Juaristi, M.; Martínez-López, S.; Sarria, B.; Bravo, L.; Mateos, R. Absorption and metabolism of yerba mate phenolic compounds in humans. Food Chem. 2018, 240, 1028–1038. [Google Scholar] [CrossRef] [Green Version]
- Carregosa, D.; Pinto, C.; Ávila-Gálvez, M.Á.; Bastos, P.; Berry, D.; Santos, C.N. A look beyond dietary (poly)phenols: The low molecular weight phenolic metabolites and their concentrations in human circulation. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3931–3962. [Google Scholar] [CrossRef]
- Di Pede, G.; Bresciani, L.; Calani, L.; Petrangolini, G.; Riva, A.; Allegrini, P.; Del Rio, D.; Mena, P. The human microbial metabolism of quercetin in different formulations: An in vitro evaluation. Foods 2020, 9, 1121. [Google Scholar] [CrossRef]
- Juániz, I.; Ludwig, I.A.; Bresciani, L.; Dall’Asta, M.; Mena, P.; Del Rio, D.; Cid, C.; De Peña, M.P. Catabolism of raw and cooked green pepper (Capsicum annuum) (poly)phenolic compounds after simulated gastrointestinal digestion and faecal fermentation. J. Funct. Foods 2016, 27, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Jaganath, I.B.; Mullen, W.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. In vitro catabolism of rutin by human fecal bacteria and the antioxidant capacity of its catabolites. Free Radic. Biol. Med. 2009, 47, 1180–1189. [Google Scholar] [CrossRef]
- Aura, A.M.; O’Leary, K.A.; Williamson, G.; Ojala, M.; Bailey, M.; Puupponen-Pimiä, R.; Nuutila, A.M.; Oksman-Caldentey, K.M.; Poutanen, K. Quercetin derivatives are deconjugated and converted to hydroxyphenylacetic acids but not methylated by human fecal flora in vitro. J. Agric. Food Chem. 2002, 50, 1725–1730. [Google Scholar] [CrossRef]
- Gonthier, M.P.; Remesy, C.; Scalbert, A.; Cheynier, V.; Souquet, J.M.; Poutanen, K.; Aura, A.M. Microbial metabolism of caffeic acid and its esters chlorogenic and caftaric acids by human faecal microbiota in vitro. Biomed. Pharmacother. 2006, 60, 536–540. [Google Scholar] [CrossRef]
- Ludwig, I.A.; de Peña, M.P.; Cid, C.; Crozier, A. Catabolism of coffee chlorogenic acids by human colonic microbiota. Biofactors 2013, 39, 623–632. [Google Scholar] [CrossRef]
- Mills, C.E.; Tzounis, X.; Oruna-Concha, M.J.; Mottram, D.S.; Gibson, G.R.; Spencer, J.P.E. In vitro colonic metabolism of coffee and chlorogenic acid results in selective changes in human faecal microbiota growth. Br. J. Nutr. 2015, 113, 1220–1227. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, P.C.; Huss, U.; Jenner, A.; Halliwell, B.; Bohlin, L.; Rafter, J.J. Human fecal water inhibits COX-2 in colonic HT-29 cells: Role of phenolic compounds. J. Nutr. 2005, 135, 2343–2349. [Google Scholar] [CrossRef] [Green Version]
- Jenner, A.M.; Rafter, J.; Halliwell, B. Human fecal water content of phenolics: The extent of colonic exposure to aromatic compounds. Free Radic. Biol. Med. 2005, 38, 763–772. [Google Scholar] [CrossRef]
- Knust, U.; Erben, G.; Spiegelhalder, B.; Bartsch, H.; Owen, R.W. Identification and quantitation of phenolic compounds in faecal matrix by capillary gas chromatography and nano-electrospray mass spectrometry. Rapid Commun. Mass. Spectrom. 2006, 20, 3119–3129. [Google Scholar] [CrossRef]
- Gill, C.I.R.; Mcdougall, G.J.; Glidewell, S.; Stewart, D.; Shen, Q.; Tuohy, K.; Dobbin, A.; Boyd, A.; Brown, E.; Haldar, S.; et al. Profiling of phenols in human fecal water after raspberry supplementation. J. Agric. Food Chem. 2010, 58, 10389–10395. [Google Scholar] [CrossRef]
- Ekbatan, S.S.; Li, X.Q.; Ghorbani, M.; Azadi, B.; Kubow, S. Chlorogenic acid and its microbial metabolites exert anti-proliferative effects, S-phase cell-cycle arrest and apoptosis in human colon cancer caco-2 cells. Int. J. Mol. Sci. 2018, 19, 723. [Google Scholar] [CrossRef] [Green Version]
- Glei, M.; Kirmse, A.; Habermann, N.; Persin, C.; Pool-Zobel, B.L. Bread enriched with green coffee extract has chemoprotective and antigenotoxic activities in human cells. Nutr. Cancer 2006, 56, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.d.S.; Silva, N.J.A.; Soares, N.C.P.; Monteiro, M.C.; Teodoro, A.J. Anticancer properties of phenolic acids in colon cancer—A review. J. Nutr. Food Sci. 2016, 6, 1000468. [Google Scholar]
- Gao, K.; Xu, A.; Krul, C.; Venema, K.; Liu, Y.; Niu, Y.; Lu, J.; Bensoussan, L.; Seeram, N.P.; Heber, D.; et al. Of the major phenolic acids formed during human microbial fermentation of tea, citrus, and soy flavonoid supplements, only 3,4-dihydroxyphenylacetic acid has antiproliferative activity. J. Nutr. 2006, 136, 52–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Kim, S.Y.; Park, S.Y.; Han, M.J. Metabolism of quercitrin by human intestinal bacteria and its relation to some biological activities. Biol. Pharm. Bull. 1999, 22, 749–751. [Google Scholar] [CrossRef] [PubMed]
- Dihal, A.A.; Woutersen, R.A.; Van Ommen, B.; Rietjens, I.M.C.M.; Stierum, R.H. Modulatory effects of quercetin on proliferation and differentiation of the human colorectal cell line Caco-2. Cancer Lett. 2006, 238, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Neamtu, A.A.; Maghiar, T.A.; Alaya, A.; Olah, N.K.; Turcus, V.; Pelea, D.; Totolici, B.D.; Neamtu, C.; Maghiar, A.M.; Mathe, E. A Comprehensive View on the Quercetin Impact on Colorectal Cancer. Molecules 2022, 27, 1873. [Google Scholar] [CrossRef] [PubMed]
- Darband, S.G.; Kaviani, M.; Yousefi, B.; Sadighparvar, S.; Pakdel, F.G.; Attari, J.A.; Mohebbi, I.; Naderi, S.; Majidinia, M. Quercetin: A functional dietary flavonoid with potential chemo-preventive properties in colorectal cancer. J. Cell Physiol. 2018, 233, 6544–6560. [Google Scholar] [CrossRef]
- Rosa, L.S.; Jordão, N.A.; da Costa Pereira Soares, N.; deMesquita, J.F.; Monteiro, M.; Teodoro, A.J. Pharmacokinetic, Antiproliferative and Apoptotic Effects of Phenolic Acids in Human Colon Adenocarcinoma Cells Using In Vitro and In Silico Approaches. Molecules 2018, 23, 2569. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hassan, Y.I.; Liu, R.; Mats, L.; Yang, C.; Liu, C.; Tsao, R. Molecular mechanisms underlying the absorption of aglycone and glycosidic flavonoids in a Caco-2 BBe1 cell model. ACS Omega 2020, 5, 10782–10793. [Google Scholar] [CrossRef]
- Maiyo, F.C.; Moodley, R.; Singh, M. Cytotoxicity, Antioxidant and Apoptosis Studies of Quercetin-3-O Glucoside and 4-(β-D-Glucopyranosyl-1→4-α-L-Rhamnopyranosyloxy)-Benzyl Isothiocyanate from Moringa oleifera. Anticancer. Agents Med. Chem. 2016, 16, 648–656. [Google Scholar] [CrossRef]
- Gómez-Juaristi, M.; Sarria, B.; Goya, L.; Bravo-Clemente, L.; Mateos, R. Experimental confounding factors affecting stability, transport, and metabolism of flavanols and hydroxycinnamic acids in Caco-2 cells. Food Res. Int. 2020, 129, 108797. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Antiproliferative Activity and Cell Metabolism of Hydroxycinnamic Acids in Human Colon Adenocarcinoma Cell Lines. J. Agric. Food Chem. 2019, 67, 3919–3931. [Google Scholar] [CrossRef]
- Konishi, Y.; Kobayashi, S. Transepithelial transport of chlorogenic acid, caffeic acid, and their colonic metabolites in intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2004, 52, 2518–2526. [Google Scholar] [CrossRef]
- Konishi, Y. Transepithelial transport of microbial metabolites of quercetin in intestinal Caco-2 cell monolayers. J. Agric. Food Chem. 2005, 53, 601–607. [Google Scholar] [CrossRef]
- Nicoletti, I.; Migliorati, G.; Pagliacci, M.C.; Grignani, F.; Riccardi, C. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J. Immunol. Methods 1991, 139, 271–279. [Google Scholar] [CrossRef]
- Sala, R.; Mena, P.; Savi, M.; Brighenti, F.; Crozier, A.; Miragoli, M.; Stilli, D.; Del Rio, D. Urolithins at physiological concentrations affect the levels of pro-inflammatory cytokines and growth factor in cultured cardiac cells in hyperglucidic conditions. J. Funct. Foods 2015, 15, 97–105. [Google Scholar] [CrossRef]
- Martini, S.; Tagliazucchi, D.; Minelli, G.; Lo Fiego, D.P. Influence of linseed and antioxidant-rich diets in pig nutrition on lipid oxidation during cooking and in vitro digestion of pork. Food Res. Int. 2020, 137, 109528. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Caco-2 (μmol/L) | SW 480 (μmol/L) | ||||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| Quercetin-3-O-glucoside-4′-O-glucoside | n.a. | n.a. | n.a. | n.a. | >200 | ~200 |
| Quercetin-4′-O-glucoside | n.a. | >200 | >200 | n.a. | 144.3 ± 6.6 | 102.2 ± 5.3 # |
| Quercetin | n.a. | >200 | 116.1 ± 4.9 | n.a. | 89.2 ± 3.1 | 58.2 ± 2.7 |
| 3-O-Caffeoylquinic acid | 97.3 ± 2.9 # | 42.8 ± 1.7 * | 40.4 ± 2.0 * | n.a. | >200 | >200 |
| 5-O-Caffeoylquinic acid | 169.5 ± 8.3 | 98.8 ± 3.0 # | 31.3 ± 2.1 | n.a. | >200 | >200 |
| 3,5-di-O-Caffeoylquinic acid | >200 | 27.5 ± 1.0 | 8.8 ± 0.2 | n.a. | >200 | >200 |
| 3-(3′-Hydroxyphenyl)acetic acid | n.a. | > 200 | > 200 | n.a. | >200 | 99.6 ± 4.8 # |
| 3-(3′,4′-Dihydroxyphenyl)acetic acid | 79.5 ± 1.4 | 14.7 ± 0.8 | 3.0 ± 0.1 | n.a. | >200 | 92.2 ± 6.9 # |
| 3-(3′-Hydroxyphenyl)propanoic acid | >200 | >200 | >200 | n.a. | n.a. | >200 |
| Compound | SW 480 (μmol/L) | ||||
|---|---|---|---|---|---|
| 0 h | 24 h | 48 h | 72 h | ||
| Substrate | Quercetin-3-O-glucoside-4′-O-glucoside | 50 | 42.77 ± 0.10 | 37.04 ± 0.35 | 23.22 ± 0.22 |
| Metabolite | Quercetin-3-O-glucoside | 3.35 ± 0.01 | 8.94 ± 0.03 | 9.34 ± 0.05 | |
| Quercetin | n.d. | 0.04 ± 0.00 | 0.07 ± 0.00 | ||
| Isorhamnetin | n.d. | 0.01 ± 0.00 | 0.01 ± 0.00 | ||
| 4′-O-methylquercetin | n.d. | 0.01 ± 0.00 | 0.01 ± 0.00 | ||
| Substrate | Quercetin-4′-O-glucoside | 50 | 2.98 ± 0.07 | 0.07 ± 0.00 | 0.01 ± 0.00 |
| Metabolite | Quercetin | 52.70 ± 2.24 | 36.89 ± 0.52 | 12.78 ± 0.41 | |
| Isorhamnetin | n.d. | 0.79 ± 0.03 | 1.03 ± 0.04 | ||
| 4′-O-methylquercetin | n.d. | 0.49 ± 0.01 | 0.71 ± 0.04 | ||
| Quercetin-O-sulphate | n.d. | 0.06 ± 0.00 | 0.12 ± 0.01 | ||
| Substrate | Quercetin | 50 | 47.39 ± 1.22 | 40.37 ± 1.20 | 14.83 ± 0.84 |
| Metabolite | Isorhamnetin | 1.61 ± 0.01 | 3.46 ± 0.08 | 3.76 ± 0.11 | |
| 4′-O-methylquercetin | 0.97 ± 0.04 | 1.65 ± 0.04 | 1.83 ± 0.25 | ||
| Quercetin-O-sulphate | 0.26 ± 0.01 | 0.48 ± 0.01 | 0.61 ± 0.01 | ||
| Substrate | 3-O-Caffeoylquinic acid trans | 50 | 23.34 ± 0.07 | 19.54 ± 0.07 | 19.83 ± 0.05 |
| Metabolite | 5-O-Caffeoylquinic acid trans | 4.95 ± 0.04 | 7.37 ± 0.07 | 11.32 ± 0.43 | |
| 4-O-Caffeoylquinic acid trans | 6.89 ± 0.06 | 8.07 ± 0.24 | 12.02 ± 0.24 | ||
| 3-O-Caffeoylquinic acid cis | 4.72 ± 0.04 | 2.51 ± 0.01 | 3.27 ± 0.17 | ||
| Caffeic acid | 0.44 ± 0.01 | 0.43 ± 0.01 | 0.57 ± 0.01 | ||
| 3-O-Feruloylquinic acid | 0.08 ± 0.00 | 0.12 ± 0.00 | 0.17 ± 0.00 | ||
| 4-O-Feruloylquinic acid | 0.02 ± 0.00 | 0.04 ± 0.00 | 0.07 ± 0.00 | ||
| 5-O-Feruloylquinic acid | 0.03 ± 0.00 | 0.03 ± 0.00 | 0.05 ± 0.00 | ||
| Substrate | 5-O-Caffeoylquinic acid trans | 50 | 16.76 ± 0.01 | 17.52 ± 0.08 | 13.96 ± 0.03 |
| Metabolite | 5-O-Caffeoylquinic acid cis | 2.50 ± 0.01 | 0.97 ± 0.05 | 1.42 ± 0.01 | |
| 3-O-Caffeoylquinic acid trans | 2.21 ± 0.05 | 5.14 ± 0.07 | 5.55 ± 0.09 | ||
| 4-O-Caffeoylquinic acid trans | 12.69 ± 0.12 | 14.80 ± 0.09 | 11.93 ± 0.03 | ||
| 3-O-Caffeoylquinic acid cis | 0.41 ± 0.05 | 0.36 ± 0.04 | 0.72 ± 0.03 | ||
| Caffeic acid | n.d. | 0.08 ± 0.00 | 0.09 ± 0.01 | ||
| 3-O-Feruloylquinic acid | 0.06 ± 0.00 | 0.07 ± 0.00 | 0.05 ± 0.00 | ||
| 4-O-Feruloylquinic acid | 0.04 ± 0.00 | 0.05 ± 0.00 | 0.04 ± 0.00 | ||
| 5-O-Feruloylquinic acid | 0.01 ± 0.00 | 0.03 ± 0.00 | 0.03 ± 0.00 | ||
| Ferulic acid | 0.06 ± 0.02 | 0.33 ± 0.01 | 0.37 ± 0.01 | ||
| Substrate | 3,5-di-O-Caffeoylquinic acid | 50 | 7.13 ± 0.13 | 5.41 ± 0.07 | 6.17 ± 0.05 |
| Metabolite | 1,3-di-O-Caffeoylquinic acid | 8.97 ± 0.04 | 8.87 ± 0.05 | 10.98 ± 0.02 | |
| 3,4-di-O-Caffeoylquinic acid | 10.12 ± 0.10 | 8.04 ± 0.14 | 9.05 ± 0.15 | ||
| 4,5-di-O-Caffeoylquinic acid | 3.30 ± 0.05 | 2.20 ± 0.02 | 1.71 ± 0.03 | ||
| 1,4-di-O-Caffeoylquinic acid | 0.91 ± 0.03 | 0.66 ± 0.03 | 0.55 ± 0.01 | ||
| 3-O-Caffeoylquinic acid trans | 0.09 ± 0.01 | 0.13 ± 0.01 | 0.27 ± 0.01 | ||
| 4-O-Caffeoylquinic acid trans | 0.07 ± 0.01 | 0.12 ± 0.01 | 0.25 ± 0.01 | ||
| 5-O-Caffeoylquinic acid trans | 0.08 ± 0.01 | 0.14 ± 0.01 | 0.29 ± 0.01 | ||
| Caffeic acid | n.d. | n.d. | 1.07 ± 0.01 | ||
| Substrate | 3-(3′-hydroxyphenyl)acetic acid | 50 | 42.23 ± 0.55 | 42.81 ± 0.10 | 39.28 ± 0.85 |
| Substrate | 3-(3′,4′-dihydroxyphenyl)acetic acid | 50 | 21.62 ± 0.48 | 15.84 ± 0.09 | 10.30 ± 0.36 |
| Substrate | 3-(3′-hydroxyphenyl)propanoic acid | 50 | 52.74 ± 0.44 | 50.70 ± 0.11 | 49.95 ± 0.37 |
| Compound | Caco-2 (μmol/L) | ||||
|---|---|---|---|---|---|
| 0 h | 24 h | 48 h | 72 h | ||
| Substrate | Quercetin-3-O-glucoside-4′-O-glucoside | 50 | 53.05 ± 0.44 | 50.05 ± 0.32 | 50.18 ± 0.66 |
| Substrate | Quercetin-4′-O-glucoside | 50 | n.d. | n.d. | n.d. |
| Substrate | Quercetin | 50 | 0.61 ± 0.01 | 0.29 ± 0.01 | 0.52 ± 0.02 |
| Substrate | 3-O-Caffeoylquinic acid trans | 50 | 2.80 ± 0.16 | 0.04 ± 0.00 | 0.02 ± 0.00 |
| Metabolite | 5-O-Caffeoylquinic acid trans | 2.71 ± 0.09 | 0.02 ± 0.00 | 0.01 ± 0.00 | |
| 4-O-Caffeoylquinic acid trans | 2.93 ± 0.10 | 0.03 ± 0.00 | 0.02 ± 0.00 | ||
| 3-O-Caffeoylquinic acid cis | 0.26 ± 0.01 | 0.02 ± 0.00 | 0.02 ± 0.00 | ||
| Substrate | 5-O-Caffeoylquinic acid trans | 50 | 2.70 ± 0.01 | 0.02 ± 0.00 | 0.02 ± 0.00 |
| Metabolite | 5-O-Caffeoylquinic acid cis | 0.12 ± 0.01 | 0.01 ± 0.00 | 0.01 ± 0.00 | |
| 3-O-Caffeoylquinic acid trans | 2.35 ± 0.02 | 0.21 ± 0.00 | 0.19 ± 0.00 | ||
| 4-O-Caffeoylquinic acid trans | 3.46 ± 0.03 | 0.09 ± 0.00 | 0.09 ± 0.00 | ||
| 3-O-Caffeoylquinic acid cis | 0.15 ± 0.01 | 0.02 ± 0.00 | 0.02 ± 0.00 | ||
| Substrate | 3,5-di-O-Caffeoylquinic acid | 50 | 0.15 ± 0.00 | 0.08 ± 0.00 | 0.18 ± 0.00 |
| Metabolite | 1,3-di-O-Caffeoylquinic acid | 0.33 ± 0.01 | 0.16 ± 0.00 | 0.39 ± 0.00 | |
| 3,4-di-O-Caffeoylquinic acid | 0.32 ± 0.00 | 0.18 ± 0.00 | 0.36 ± 0.00 | ||
| 4,5-di-O-Caffeoylquinic acid | 0.06 ± 0.00 | 0.03 ± 0.00 | 0.03 ± 0.00 | ||
| 1,4-di-O-Caffeoylquinic acid | n.d. | n.d. | 0.10 ± 0.00 | ||
| Substrate | 3-(3′-hydroxyphenyl)acetic acid | 50 | 48.83 ± 0.85 | 47.02 ± 0.39 | 43.17 ± 0.06 |
| Substrate | 3-(3′,4′-dihydroxyphenyl)acetic acid | 50 | n.d. | n.d. | n.d. |
| Hydroxybenzoic acid | n.d. | 3.51 ± 0.07 | 15.73 ± 0.37 | ||
| Substrate | 3-(3′-hydroxyphenyl)propanoic acid | 50 | 45.51 ± 0.41 | 46.27 ± 0.58 | 46.28 ± 0.76 |
| Hydroxybenzoic acid | n.d. | 0.31 ± 0.01 | 0.30 ± 0.01 | ||
| Compound | QUE-LP | QUE-EQ | QUE-HP | CGA-LP | CGA-EQ | CGA-HP |
|---|---|---|---|---|---|---|
| Quercetin-3-O-glucoside-4′-O-glucoside | 60 μmol/L | 33.3 μmol/L | 6.7 μmol/L | / | / | / |
| Quercetin-4′-O-glucoside | 60 μmol/L | 33.3 μmol/L | 6.7 μmol/L | / | / | / |
| Quercetin | 60 μmol/L | 33.3 μmol/L | 6.7 μmol/L | / | / | / |
| 3-O-Caffeoylquinic acid | / | / | / | 60 μmol/L | 50 μmol/L | 6.7 μmol/L |
| 5-O-Caffeoylquinic acid | / | / | / | 60 μmol/L | 50 μmol/L | 6.7 μmol/L |
| 3,5-di-O-Caffeoylquinic acid | / | / | / | 60 μmol/L | 50 μmol/L | 6.7 μmol/L |
| 3-(3′-Hydroxyphenyl)acetic acid | 6.7 μmol/L | 33.3 μmol/L | 60 μmol/L | / | / | / |
| 3-(3′,4′-Dihydroxyphenyl)acetic acid | 6.7 μmol/L | 33.3 μmol/L | 60 μmol/L | / | / | / |
| 3-(3′-Hydroxyphenyl)propanoic acid | 6.7 μmol/L | 33.3 μmol/L | 60 μmol/L | 20 μmol/L | 50 μmol/L | 180 μmol/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cattivelli, A.; Conte, A.; Tagliazucchi, D. Quercetins, Chlorogenic Acids and Their Colon Metabolites Inhibit Colon Cancer Cell Proliferation at Physiologically Relevant Concentrations. Int. J. Mol. Sci. 2023, 24, 12265. https://doi.org/10.3390/ijms241512265
Cattivelli A, Conte A, Tagliazucchi D. Quercetins, Chlorogenic Acids and Their Colon Metabolites Inhibit Colon Cancer Cell Proliferation at Physiologically Relevant Concentrations. International Journal of Molecular Sciences. 2023; 24(15):12265. https://doi.org/10.3390/ijms241512265
Chicago/Turabian StyleCattivelli, Alice, Angela Conte, and Davide Tagliazucchi. 2023. "Quercetins, Chlorogenic Acids and Their Colon Metabolites Inhibit Colon Cancer Cell Proliferation at Physiologically Relevant Concentrations" International Journal of Molecular Sciences 24, no. 15: 12265. https://doi.org/10.3390/ijms241512265
APA StyleCattivelli, A., Conte, A., & Tagliazucchi, D. (2023). Quercetins, Chlorogenic Acids and Their Colon Metabolites Inhibit Colon Cancer Cell Proliferation at Physiologically Relevant Concentrations. International Journal of Molecular Sciences, 24(15), 12265. https://doi.org/10.3390/ijms241512265