
Overexpression of BmJHBPd2 Repressed Silk Synthesis by Inhibiting the JH/Kr-h1 Signaling Pathway in Bombyx mori

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
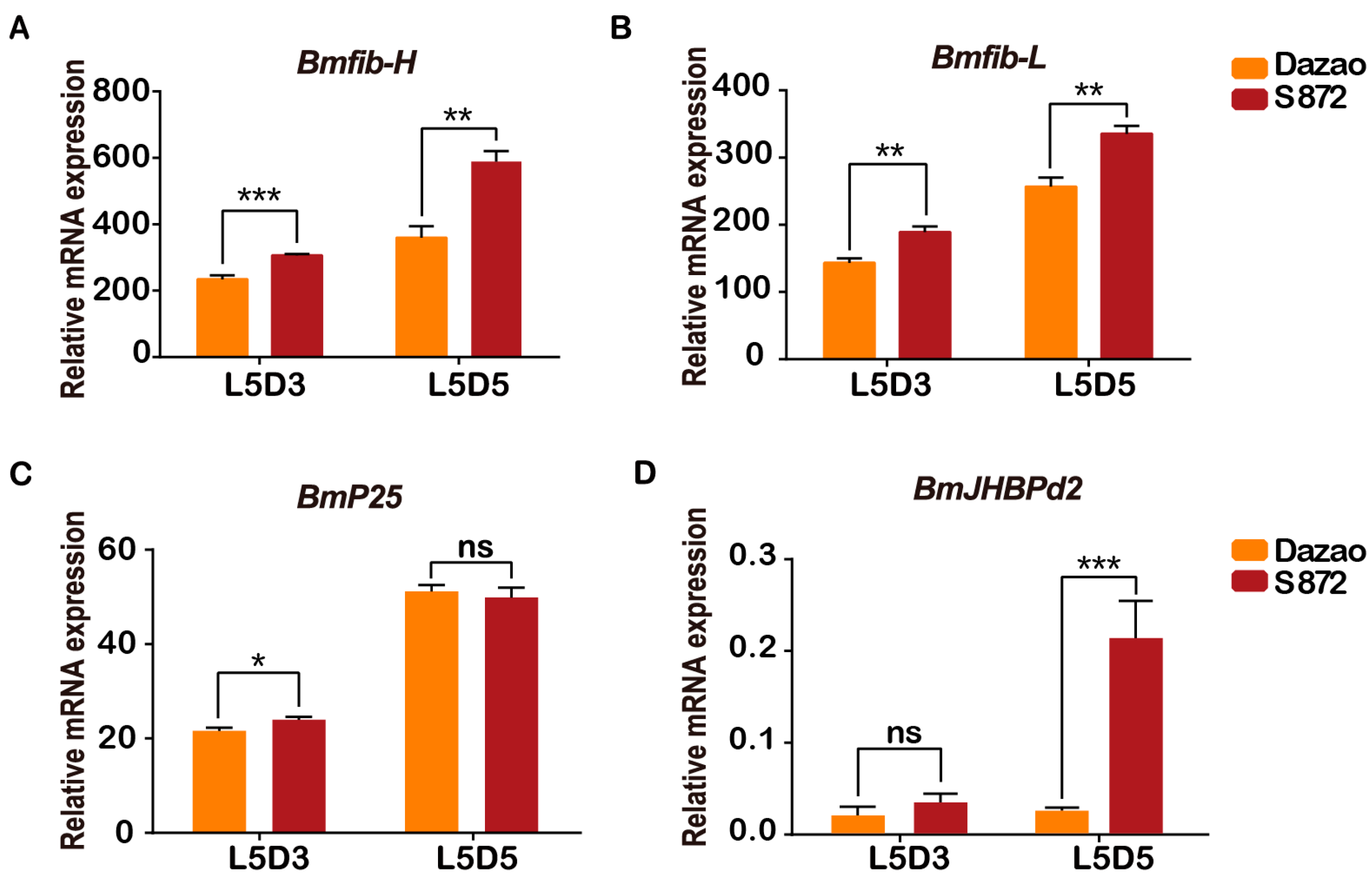
2.1. Expression of BmJHBPd2 in Different Silk-Producing Strains
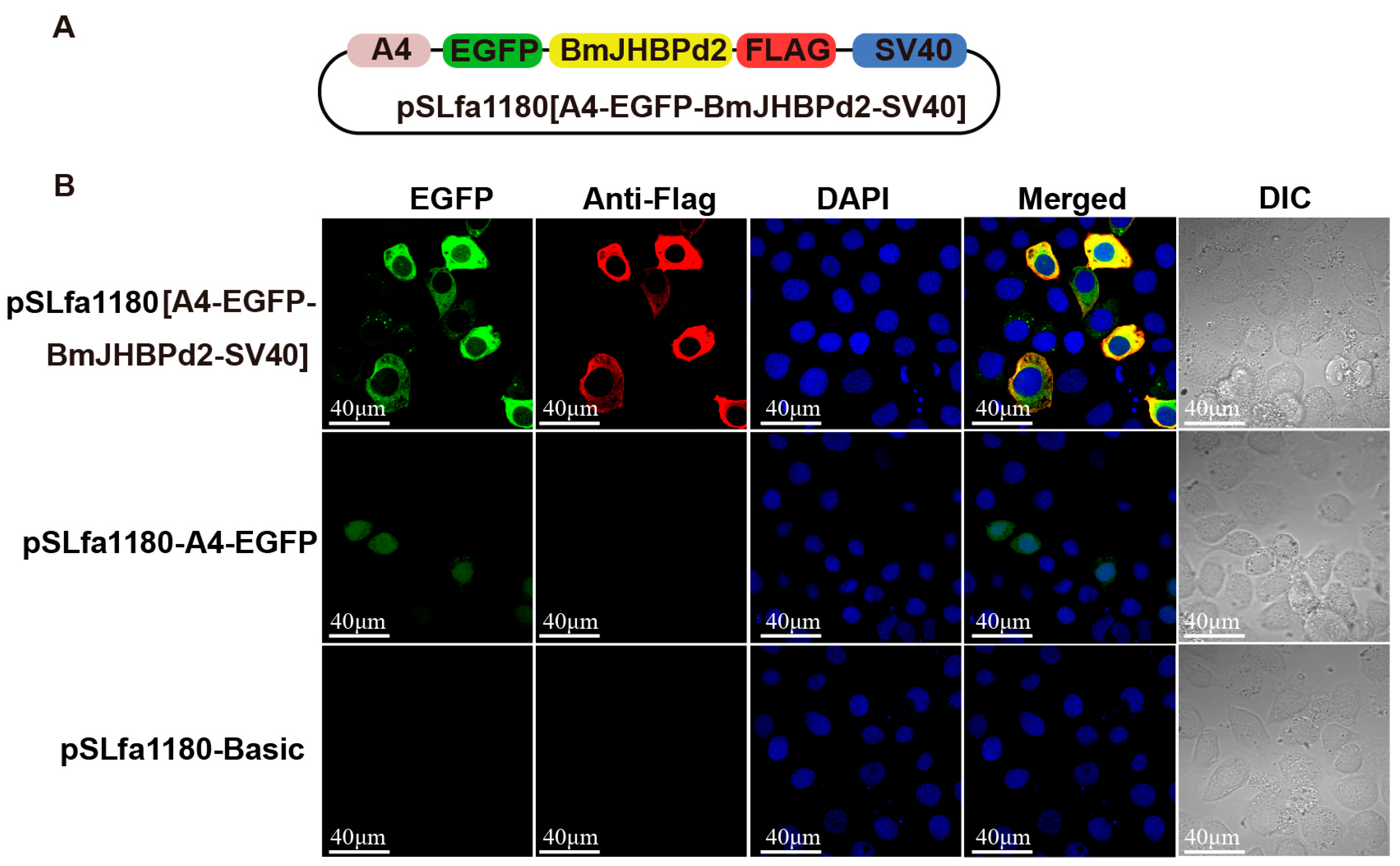
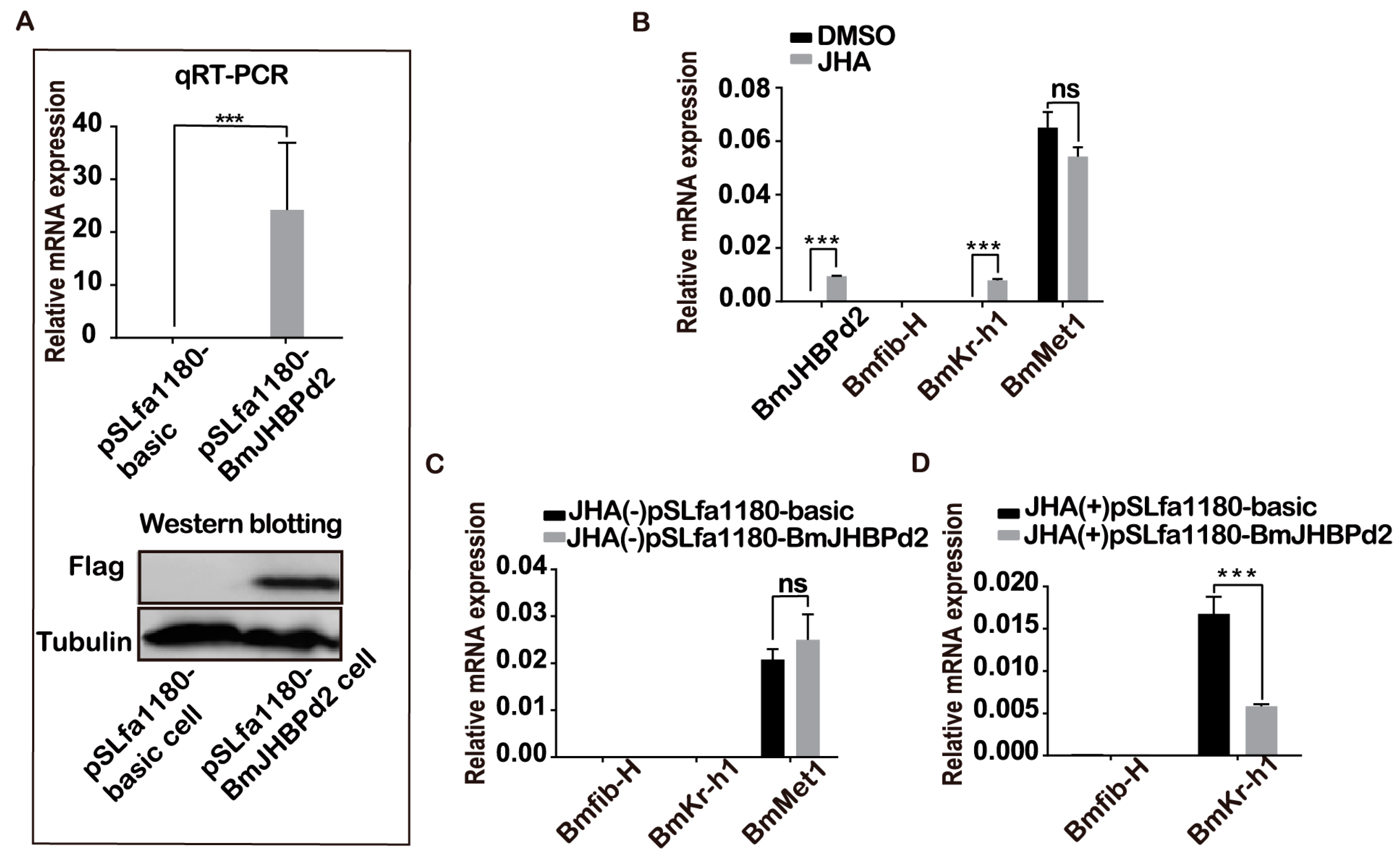
2.2. Overexpression of BmJHBPd2 at the Cellular Level
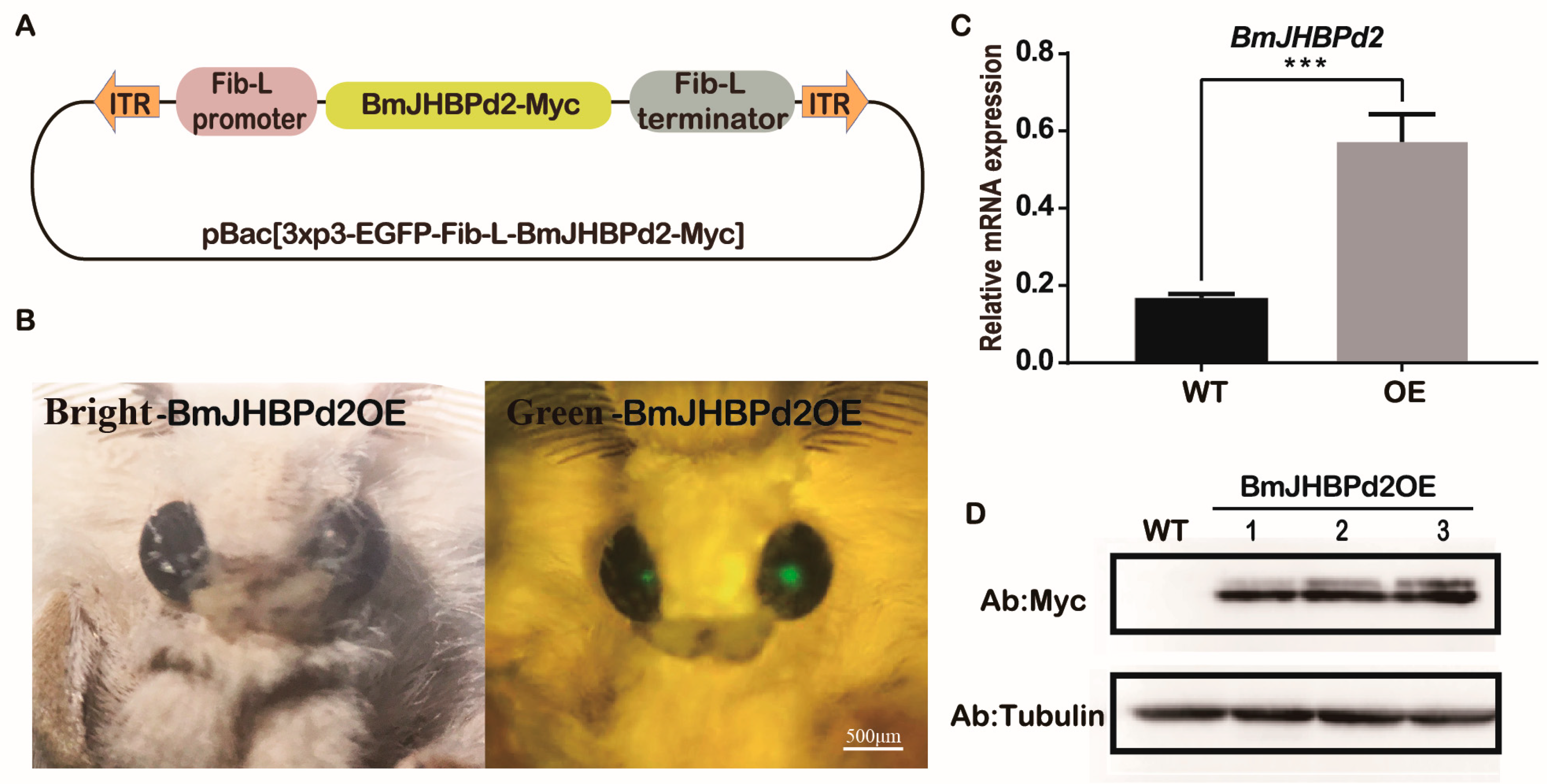
2.3. Transgenic Overexpression of BmJHBPd2 in the Silk Gland
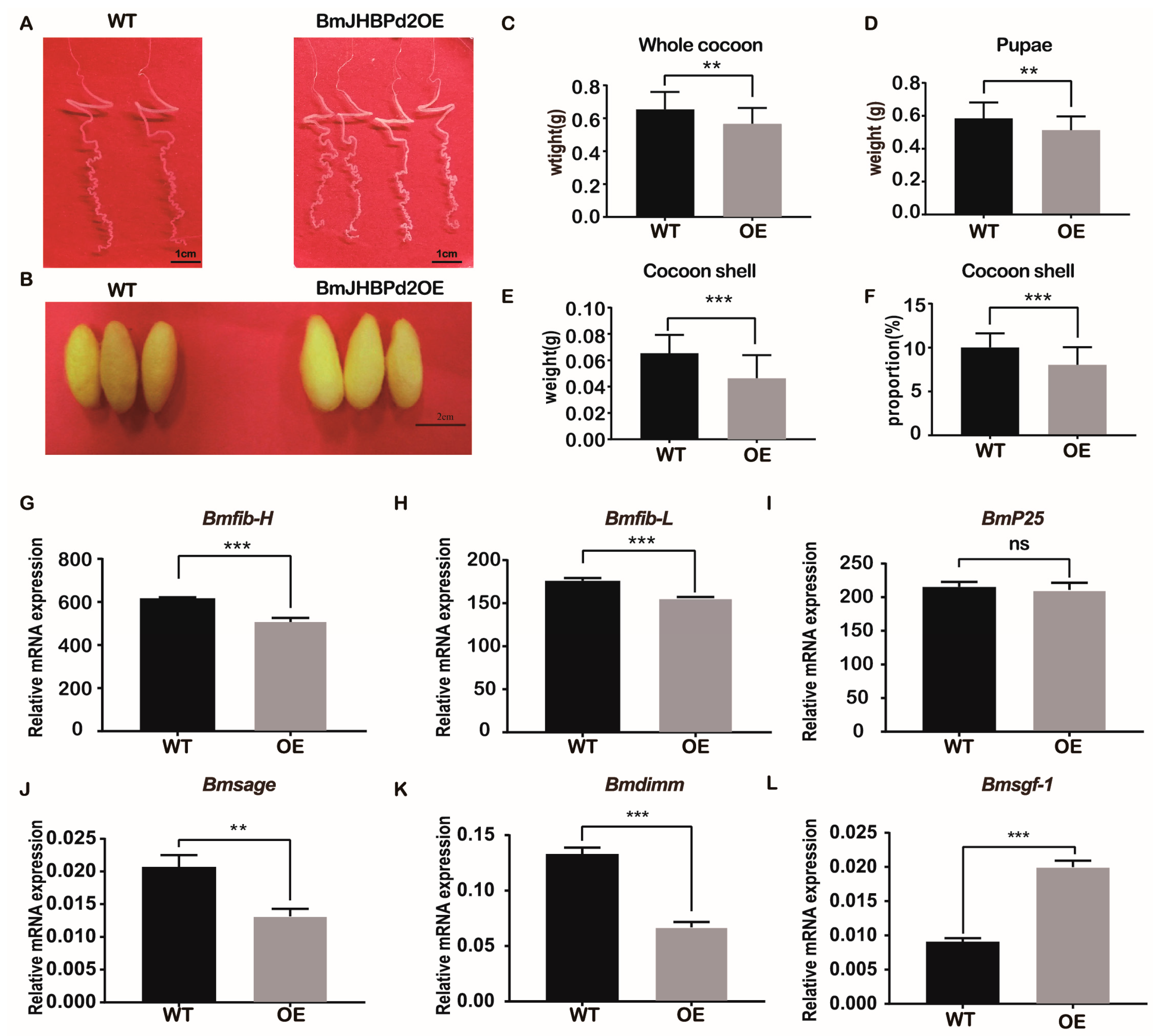
2.4. Overexpression of BmJHBPd2 Affects Silk Synthesis and Silk Yield
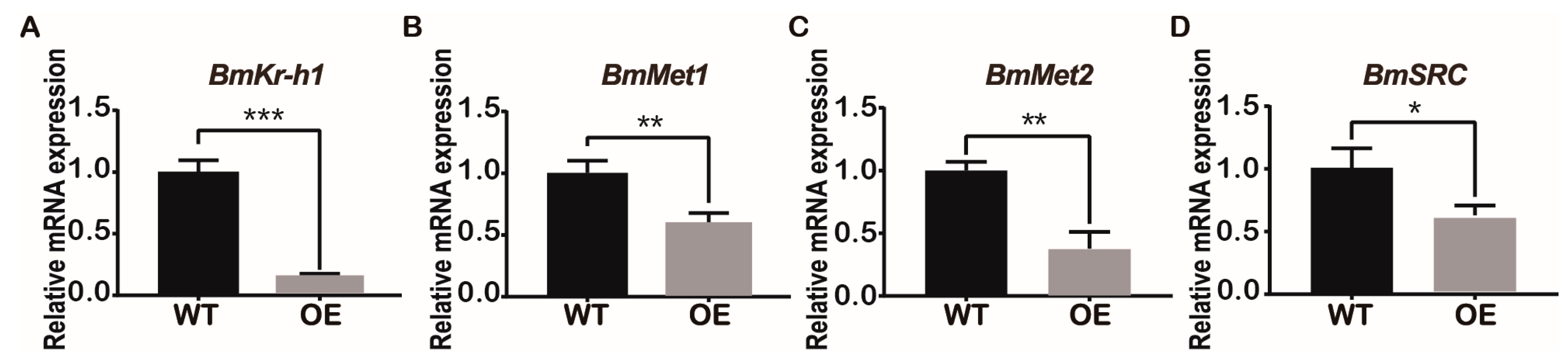
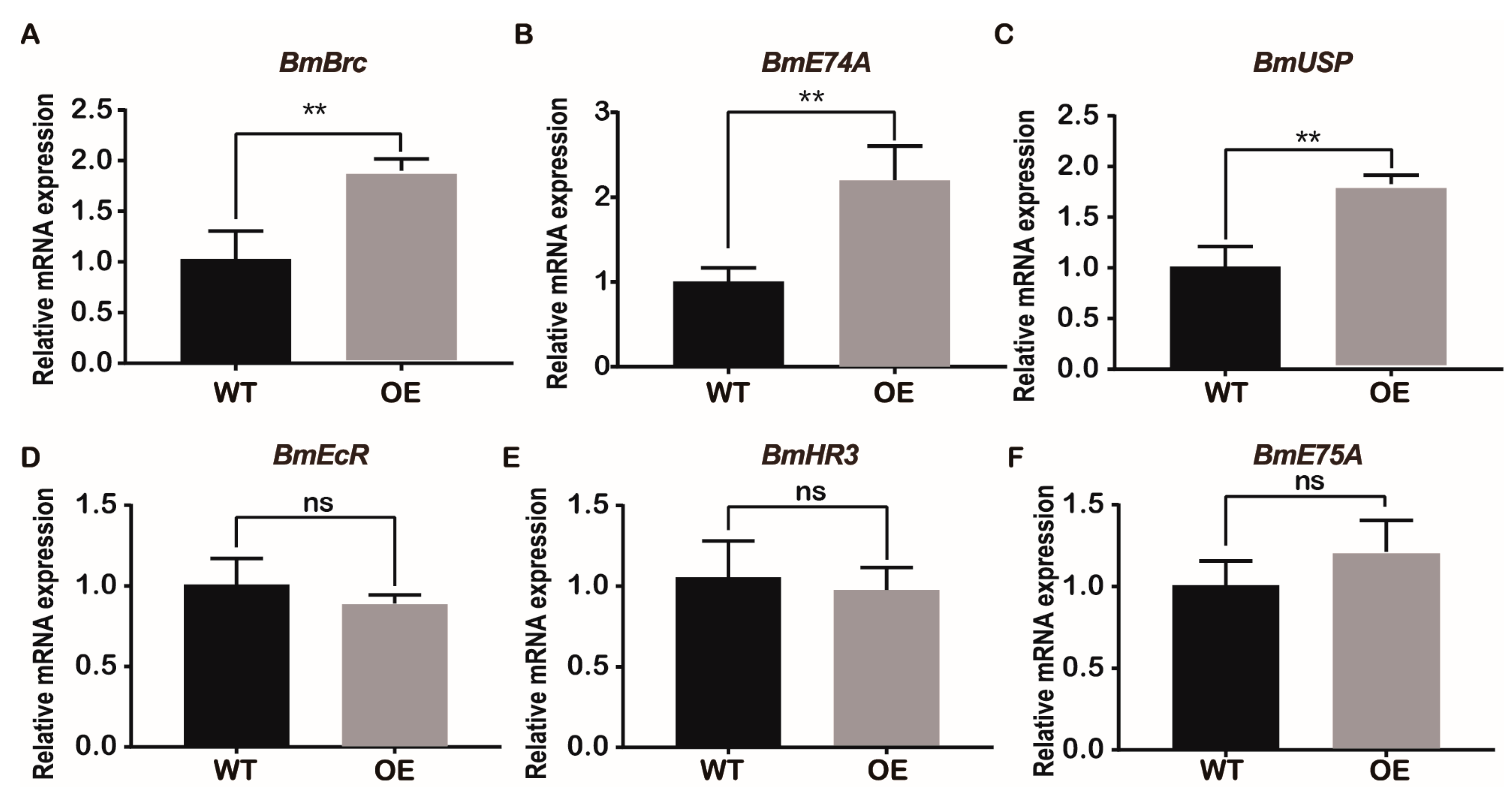
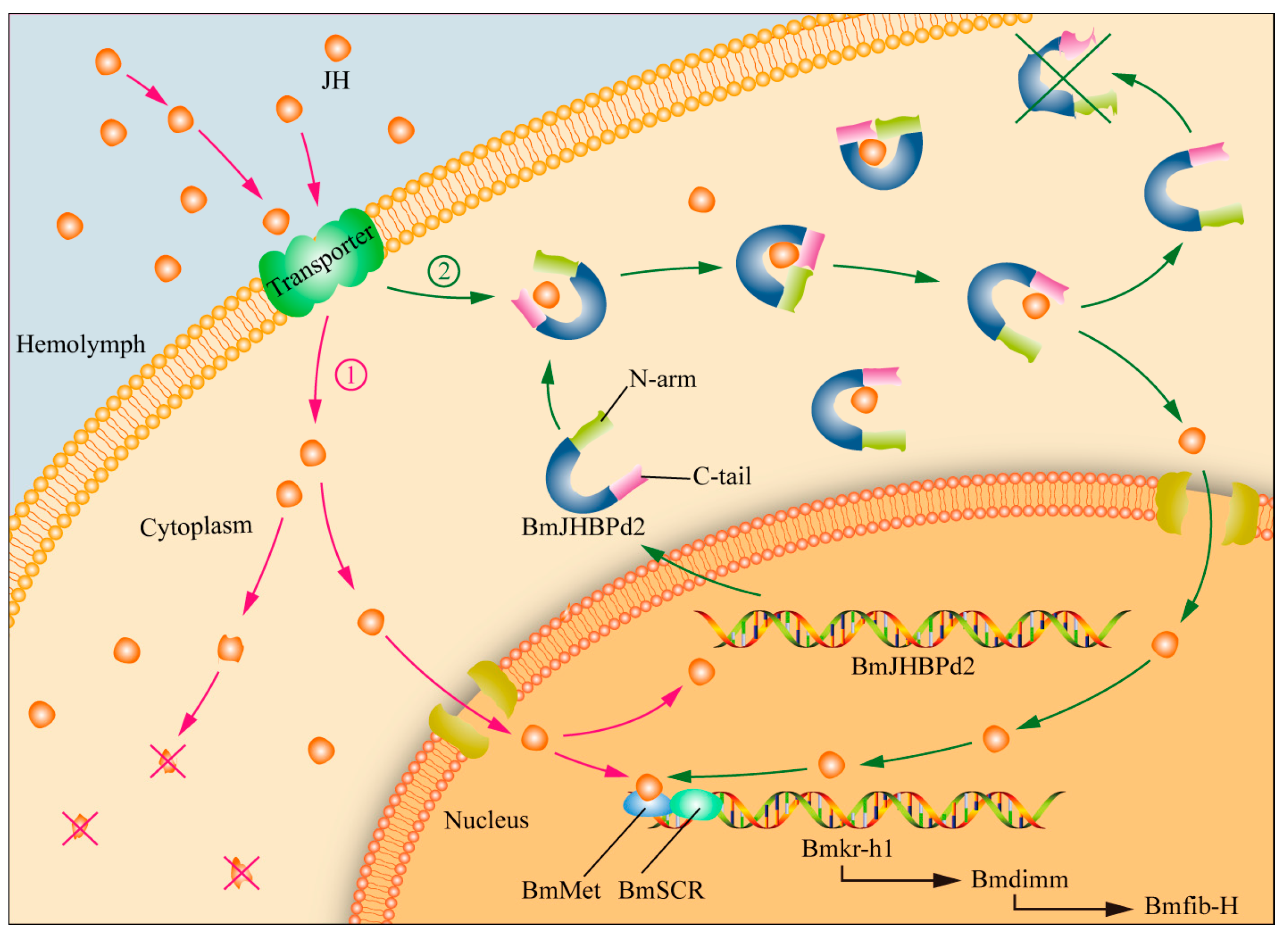
2.5. Overexpression of BmJHBPd2 Led to Repression of Silk Synthesis by Inhibiting Bmkr-h1 Expression in the Silk Glands
3. Discussion
4. Materials and Methods
4.1. Silkworm Strains and Cell Culture
4.2. RNA Preparation and Quantitative Real Time-PCR (qRT-PCR)
4.3. Subcellular Localization
4.4. Western Blotting
4.5. Statistical Method for Cocoon Layer Proportion
4.6. Plasmid Construction to Obtain Transgenic Silkworms
4.7. Silkworm Germline Transformation
4.8. Statistical Analysis
4.9. JHA Treatment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Truman, J.W.; Riddiford, L.M. The origins of insect metamorphosis. Nature 1999, 401, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.W.; Zou, Z.; Saha, T.T.; Raikhel, A.S. bHLH-PAS heterodimer of methoprene-tolerant and Cycle mediates circadian expression of juvenile hormone-induced mosquito genes. Proc. Natl. Acad. Sci. USA 2012, 109, 16576–16581. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, J.R.; Muszyńska-Pytel, M.; Grzelak, K.; Kochman, M. Biosynthesis and degradation of juvenile hormone in corpora allata and imaginal wing discs of Galleria mellonella (L.). Insect Biochem. 1987, 17, 249–254. [Google Scholar] [CrossRef]
- Sok, A.J.; Andruszewska, G.; Niewiadomska-Cimicka, A.; Grad, I.; Rymarczyk, G.; Pajdzik, D.; Orłowski, M.; Schmidt, M.T.; Grajek, W.; Ozyhar, A.; et al. Regulatory elements in the juvenile hormone binding protein gene from Galleria mellonella–topography of binding sites for Usp and EcRDBD. Biochim. Biophys. Acta 2008, 1779, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.I.; Granger, N.A.; Roe, R.M. The juvenile hormones: Historical facts and speculations on future research directions. Insect Biochem. Mol. Biol. 2000, 30, 617–644. [Google Scholar] [CrossRef]
- Hammock, B.; Nowock, J.; Goodman, W.; Stamoudis, V.; Gilbert, L.I. The influence of hemolymph-binding protein on juvenile hormone stability and distribution in Manduca sexta fat body and imaginal discs in vitro. Mol. Cell. Endocrinol. 1975, 3, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Sanburg, L.L.; Kramer, K.J.; Kezdy, F.J.; Law, J.H.; Oberlander, H. Role of juvenile hormone esterases and carrier proteins in insect development. Nature 1975, 253, 266–267. [Google Scholar] [CrossRef]
- Tauchman, S.J.; Lorch, J.M.; Orth, A.P.; Goodman, W.G. Effects of stress on the hemolymph juvenile hormone binding protein titers of Manduca sexta. Insect Biochem. Mol. Biol. 2007, 37, 847–854. [Google Scholar] [CrossRef]
- Chang, E.S.; Bruce, M.J.; Prestwich, G.D. Further characterization of the juvenile hormone binding protein from the cytosol of a Drosophila cell line: Use of a photoaffinity label. Insect Biochem. 1985, 15, 197–204. [Google Scholar] [CrossRef]
- Shemshedini, L.; Lanoue, M.; Wilson, T.G. Evidence for a juvenile hormone receptor involved in protein synthesis in Drosophila melanogaster. J. Biol. Chem. 1990, 265, 1913–1918. [Google Scholar] [CrossRef]
- Ritdachyeng, E.; Manaboon, M.; Tobe, S.S.; Singtripop, T. Molecular characterization and gene expression of juvenile hormone binding protein in the bamboo borer, Omphisa fuscidentalis. J. Insect Physiol. 2012, 58, 1493–1501. [Google Scholar] [CrossRef]
- Rebijith, K.B.; Asokan, R.; Ranjitha, H.H.; Rajendra Prasad, B.S.; Krishna, V.; Krishna Kumar, N.K. Diet-Delivered dsRNAs for Juvenile Hormone-Binding Protein and Vacuolar ATPase-H Implied Their Potential in the Management of the Melon Aphid (Hemiptera: Aphididae). Environ. Entomol. 2016, 45, 268–275. [Google Scholar] [CrossRef]
- Wei, D.; Li, H.M.; Tian, C.B.; Smagghe, G.; Jia, F.X.; Jiang, H.B.; Dou, W.; Wang, J.J. Proteome analysis of male accessory gland secretions in oriental fruit flies reveals juvenile hormone-binding protein, suggesting impact on female reproduction. Sci. Rep. 2015, 5, 16845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Su, Z.H.; Emi, A.; Mita, K.; Takeda, M.; Fujiwara, Y. Cloning and expression analysis of takeout/JHBP family genes of silkworm, Bombyx mori. Insect Mol. Biol. 2006, 15, 245–251. [Google Scholar] [CrossRef]
- Whitmore, E.; Gilbert, L.I. Haemolymph lipoprotein transport of juvenile hormone. J. Insect Physiol. 1972, 18, 1153–1167. [Google Scholar] [CrossRef]
- Kramer, K.J.; Sanburg, L.L.; Kézdy, F.J.; Law, J.H. The Juvenile Hormone Binding Protein in the Hemolymph of Manduca sexta Johannson (Lepidoptera: Sphingidae). Proc. Natl. Acad. Sci. USA 1974, 71, 493–497. [Google Scholar] [CrossRef]
- Lerro, K.A.; Prestwich, G.D. Cloning and sequencing of a cDNA for the hemolymph juvenile hormone binding protein of larval Manduca sexta. J. Biol. Chem. 1990, 265, 19800–19806. [Google Scholar] [CrossRef]
- Wojtasek, H.; Prestwich, G.D. Key disulfide bonds in an insect hormone binding protein: cDNA cloning of a juvenile hormone binding protein of Heliothis virescens and ligand binding by native and mutant forms. Biochemistry 1995, 34, 5234–5241. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez Parkitna, J.M.; Ozyhar, A.; Wiśniewski, J.R.; Kochman, M. Cloning and sequence analysis of Galleria mellonella juvenile hormone binding protein—A search for ancestors and relatives. Biol. Chem. 2002, 383, 1343–1355. [Google Scholar] [CrossRef]
- Vermunt, A.M.; Kamimura, M.; Hirai, M.; Kiuchi, M.; Shiotsuki, T. The juvenile hormone binding protein of silkworm haemolymph: Gene and functional analysis. Insect Mol. Biol. 2001, 10, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Cheng, T.; Hu, W.; Peng, Z.; Liu, C.; Xia, Q. Genome-wide identification and analysis of JHBP-domain family members in the silkworm Bombyx mori. Mol. Genet. Genom. MGG 2016, 291, 2159–2171. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef] [PubMed]
- Gamo, T.; Inokuchi, T.; Laufer, H. Polypeptides of fibroin and sericin secreted from the different sections of the silk gland in Bombyx mori. Insect Biochem. 1977, 7, 285–295. [Google Scholar] [CrossRef]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6:6:1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akai, H.; Kiguchi, K.; Mori, K. Increased Accumulation of Silk Protein Accompanying JH-Induced Prolongation of Larval Life in Bombyx mori L.(Lepidoptera: Bombycidae). Appl. Entomol. Zool. 1971, 6, 218–220. [Google Scholar] [CrossRef] [Green Version]
- Akai, H.; Kobayashi, M. Induction of Prolonged Larval Instar by the Juvenile Hormone in Bombyx mori L.: Lepidoptera: Bombycidae. Appl. Entomol. Zool. 1971, 6, 138–139. [Google Scholar] [CrossRef] [Green Version]
- Kurata, S.; Koga, K.; Sakaguchi, B. Nucleolar size in parallel with ribosomal RNA synthesis at diapause termination in the eggs of Bombyx mori. Chromosoma 1978, 68, 313–317. [Google Scholar] [CrossRef]
- Kurata, K. Effect of a juvenile hormon analogue given at various ages of 5th instar larvae on RNA synthesis in the posterior silk gland of the silkworm, Bombyx mori. J. Insect Biotechnol. Sericol. 2010, 53, 421–426. [Google Scholar] [CrossRef]
- Kayukawa, T.; Murata, M.; Kobayashi, I.; Muramatsu, D.; Okada, C.; Uchino, K.; Sezutsu, H.; Kiuchi, M.; Tamura, T.; Hiruma, K.; et al. Hormonal regulation and developmental role of Krüppel homolog 1, a repressor of metamorphosis, in the silkworm Bombyx mori. Dev. Biol. 2014, 388, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.M.; Liu, C.; Jiang, L.J.; Li, Q.Y.; Zhou, M.T.; Cheng, T.C.; Mita, K.; Xia, Q.Y. A juvenile hormone transcription factor Bmdimm-fibroin H chain pathway is involved in the synthesis of silk protein in silkworm, Bombyx mori. J. Biol. Chem. 2015, 290, 972–986. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Song, W.; Li, Z.; Qian, W.; Wei, L.; Yang, Y.; Wang, W.; Zhou, X.; Meng, M.; Peng, J.; et al. Krüppel homolog 1 represses insect ecdysone biosynthesis by directly inhibiting the transcription of steroidogenic enzymes. Proc. Natl. Acad. Sci. USA 2018, 115, 3960–3965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Xu, H.; Zhu, J.; Ma, S.; Liu, Y.; Jiang, R.J.; Xia, Q.; Li, S. Ras1(CA) overexpression in the posterior silk gland improves silk yield. Cell Res. 2011, 21, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Furuta, K.; Ichikawa, A.; Murata, M.; Kuwano, E.; Shinoda, T.; Shiotsuki, T. Determination by LC-MS of juvenile hormone titers in hemolymph of the silkworm, Bombyx mori. Biosci. Biotechnol. Biochem. 2013, 77, 988–991. [Google Scholar] [CrossRef]
- Kayukawa, T.; Nagamine, K.; Ito, Y.; Nishita, Y.; Ishikawa, Y.; Shinoda, T. Krüppel Homolog 1 Inhibits Insect Metamorphosis via Direct Transcriptional Repression of Broad-Complex, a Pupal Specifier Gene. J. Biol. Chem. 2016, 291, 1751–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayukawa, T.; Jouraku, A.; Ito, Y.; Shinoda, T. Molecular mechanism underlying juvenile hormone-mediated repression of precocious larval-adult metamorphosis. Proc. Natl. Acad. Sci. USA 2017, 114, 1057–1062. [Google Scholar] [CrossRef]
- Li, K.; Tian, L.; Guo, Z.; Guo, S.; Zhang, J.; Gu, S.H.; Palli, S.R.; Cao, Y.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E75 Isoforms Mediate Steroidogenesis Autoregulation and Regulate Developmental Timing in Bombyx. J. Biol. Chem. 2016, 291, 18163–18175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S.R.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. J. Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [Green Version]
- Cong, J.; Tao, C.; Zhang, X.; Zhang, H.; Cheng, T.; Liu, C. Transgenic Ectopic Overexpression of Broad Complex (BrC-Z2) in the Silk Gland Inhibits the Expression of Silk Fibroin Genes of Bombyx mori. Insects 2020, 11, 374. [Google Scholar] [CrossRef]
- Li, J.Y.; Yang, H.J.; Lan, T.Y.; Wei, H.; Zhang, H.R.; Chen, M.; Fan, W.; Ma, Y.Y.; Zhong, B.X. Expression profiling and regulation of genes related to silkworm posterior silk gland development and fibroin synthesis. J. Proteome Res. 2011, 10, 3551–3564. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Huang, Y.; Xu, J.; Shiotsuki, T.; Bai, H.; Palli, S.R.; Huang, Y.; Tan, A. The FOXO transcription factor controls insect growth and development by regulating juvenile hormone degradation in the silkworm, Bombyx mori. J. Biol. Chem. 2017, 292, 11659–11669. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, X.; Shiotsuki, T.; Wang, Z.; Xu, X.; Huang, Y.; Li, M.; Li, K.; Tan, A. Depletion of juvenile hormone esterase extends larval growth in Bombyx mori. Insect Biochem. Mol. Biol. 2017, 81, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayukawa, T.; Minakuchi, C.; Namiki, T.; Togawa, T.; Yoshiyama, M.; Kamimura, M.; Mita, K.; Imanishi, S.; Kiuchi, M.; Ishikawa, Y. Transcriptional regulation of juvenile hormone-mediated induction of Krüppel homolog 1, a repressor of insect metamorphosis. Proc. Natl. Acad. Sci. USA 2012, 109, 11729–11734. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Xiao, S.Q.; Chen, M.; Hong, X.J.; Lu, C. Establishment and characterization of two embryonic cell lines of Bombyx mori. In Vitro Cell. Dev. Biol. Anim. 2007, 43, 101–104. [Google Scholar] [CrossRef]
- Tamura, T.; Thibert, C.; Royer, C.; Kanda, T.; Abraham, E.; Kamba, M.; Komoto, N.; Thomas, J.L.; Mauchamp, B.; Chavancy, G.; et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotechnol. 2000, 18, 81–84. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, X.; Zhang, H.; Li, J.; Li, W.; Liu, C. Overexpression of BmJHBPd2 Repressed Silk Synthesis by Inhibiting the JH/Kr-h1 Signaling Pathway in Bombyx mori. Int. J. Mol. Sci. 2023, 24, 12650. https://doi.org/10.3390/ijms241612650
Zhang J, Zhang X, Zhang H, Li J, Li W, Liu C. Overexpression of BmJHBPd2 Repressed Silk Synthesis by Inhibiting the JH/Kr-h1 Signaling Pathway in Bombyx mori. International Journal of Molecular Sciences. 2023; 24(16):12650. https://doi.org/10.3390/ijms241612650
Chicago/Turabian StyleZhang, Jikailang, Xia Zhang, Hui Zhang, Jiaojiao Li, Wei Li, and Chun Liu. 2023. "Overexpression of BmJHBPd2 Repressed Silk Synthesis by Inhibiting the JH/Kr-h1 Signaling Pathway in Bombyx mori" International Journal of Molecular Sciences 24, no. 16: 12650. https://doi.org/10.3390/ijms241612650
APA StyleZhang, J., Zhang, X., Zhang, H., Li, J., Li, W., & Liu, C. (2023). Overexpression of BmJHBPd2 Repressed Silk Synthesis by Inhibiting the JH/Kr-h1 Signaling Pathway in Bombyx mori. International Journal of Molecular Sciences, 24(16), 12650. https://doi.org/10.3390/ijms241612650