

Betaxolol as a Potent Inhibitor of NDM-1-Positive E. coli That Synergistically Enhances the Anti-Inflammatory Effect in Combination with Meropenem


Abstract
:1. Introduction
2. Results
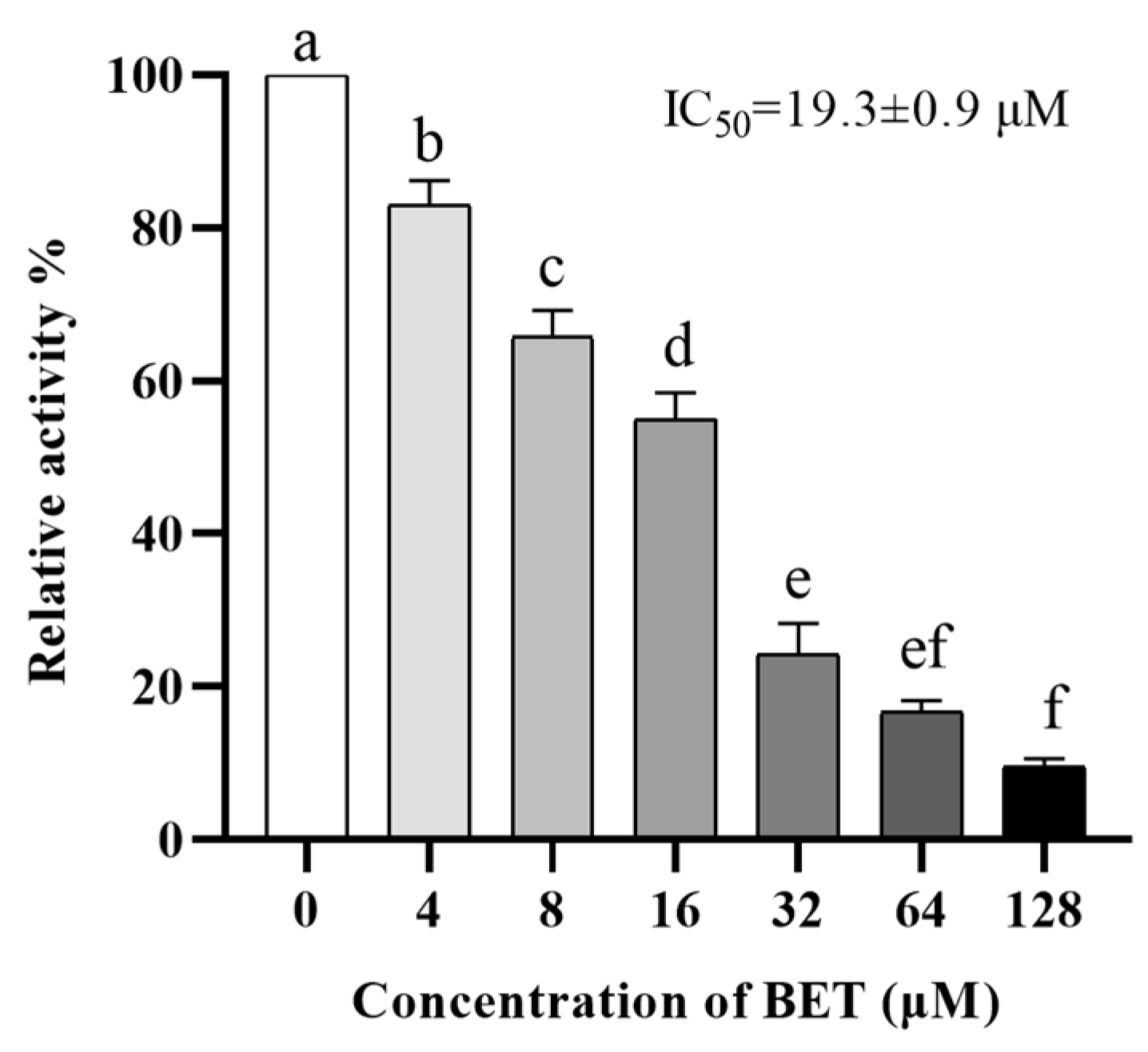
2.1. BET Has Enzymatic Inhibitory Activity against NDM-1
2.2. MIC and FICI of BET+MEM against NDM-1 E. coli
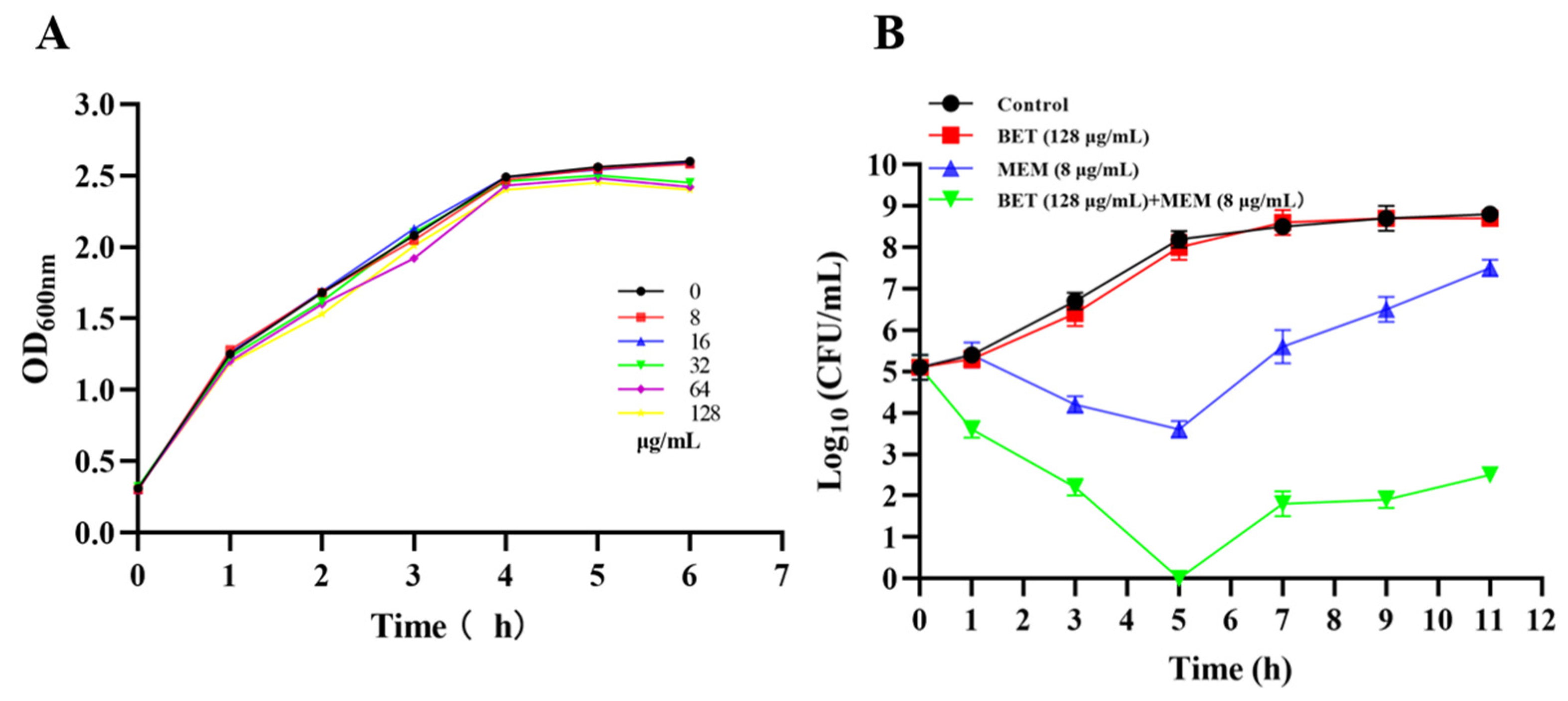
2.3. Growth Effect of NDM-1 E. coli Treated with BET+MEM
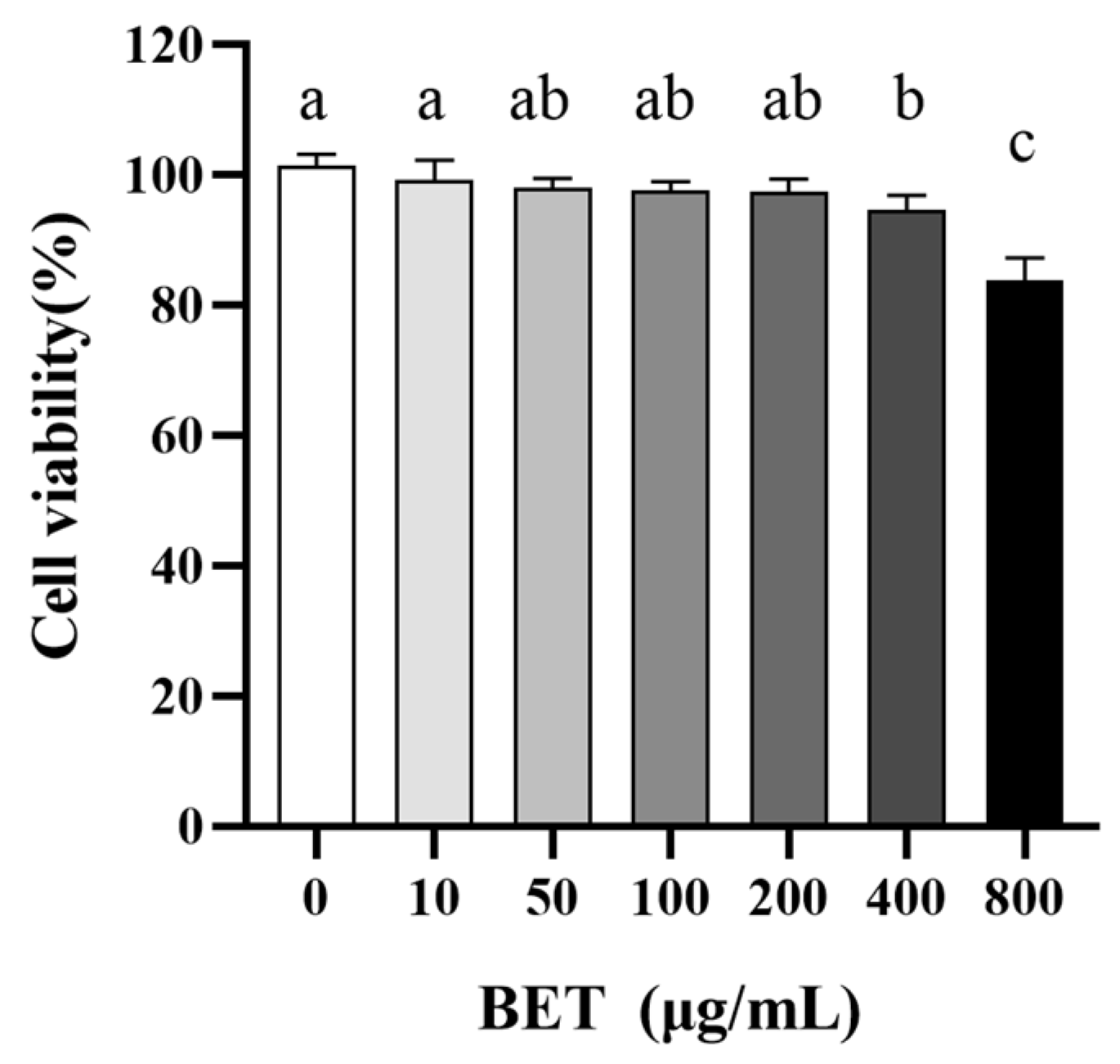
2.4. Potential Toxicity of BET to IECs
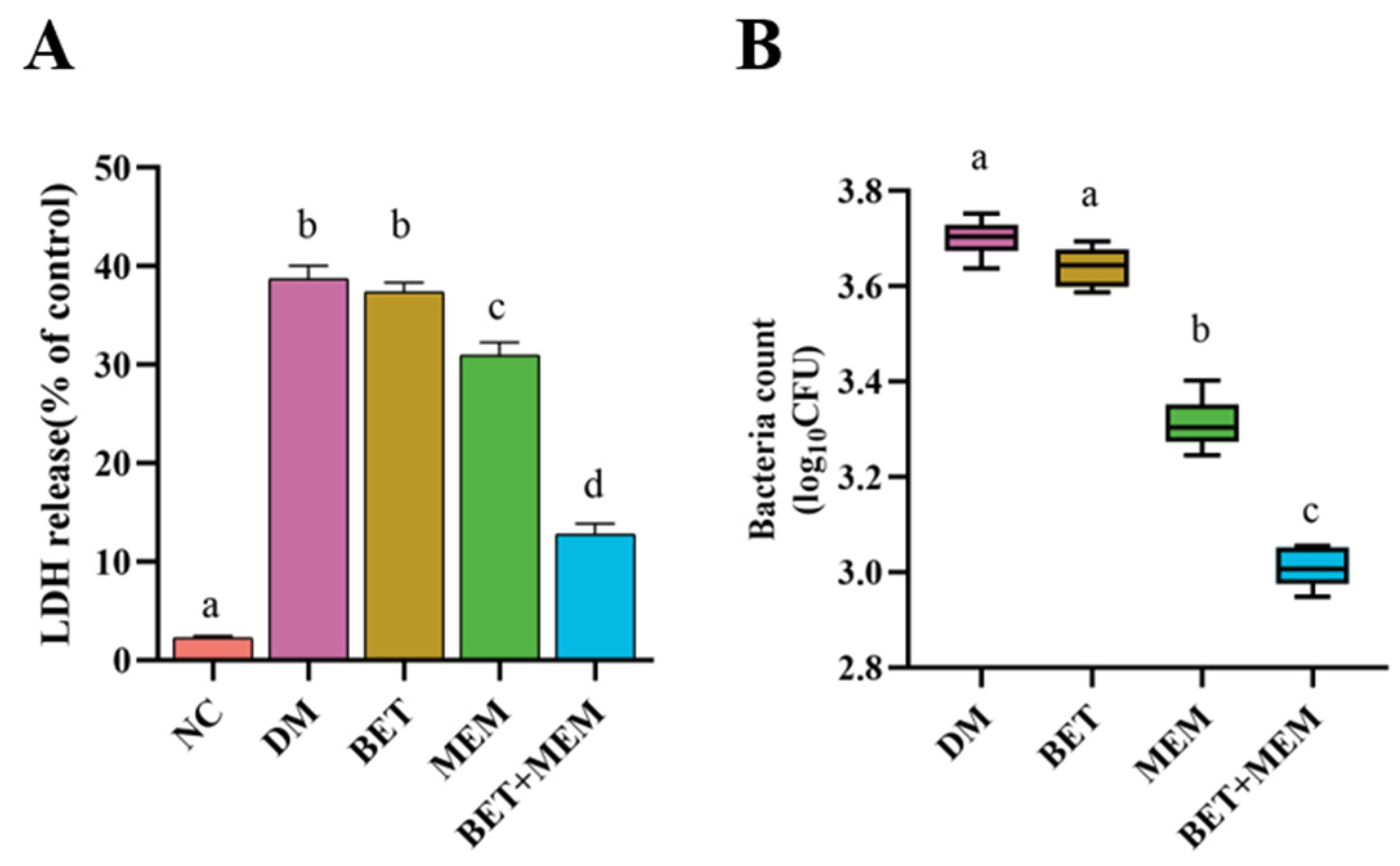
2.5. Intervention of IECs with BET+MEM in NDM-1 E. coli Infection
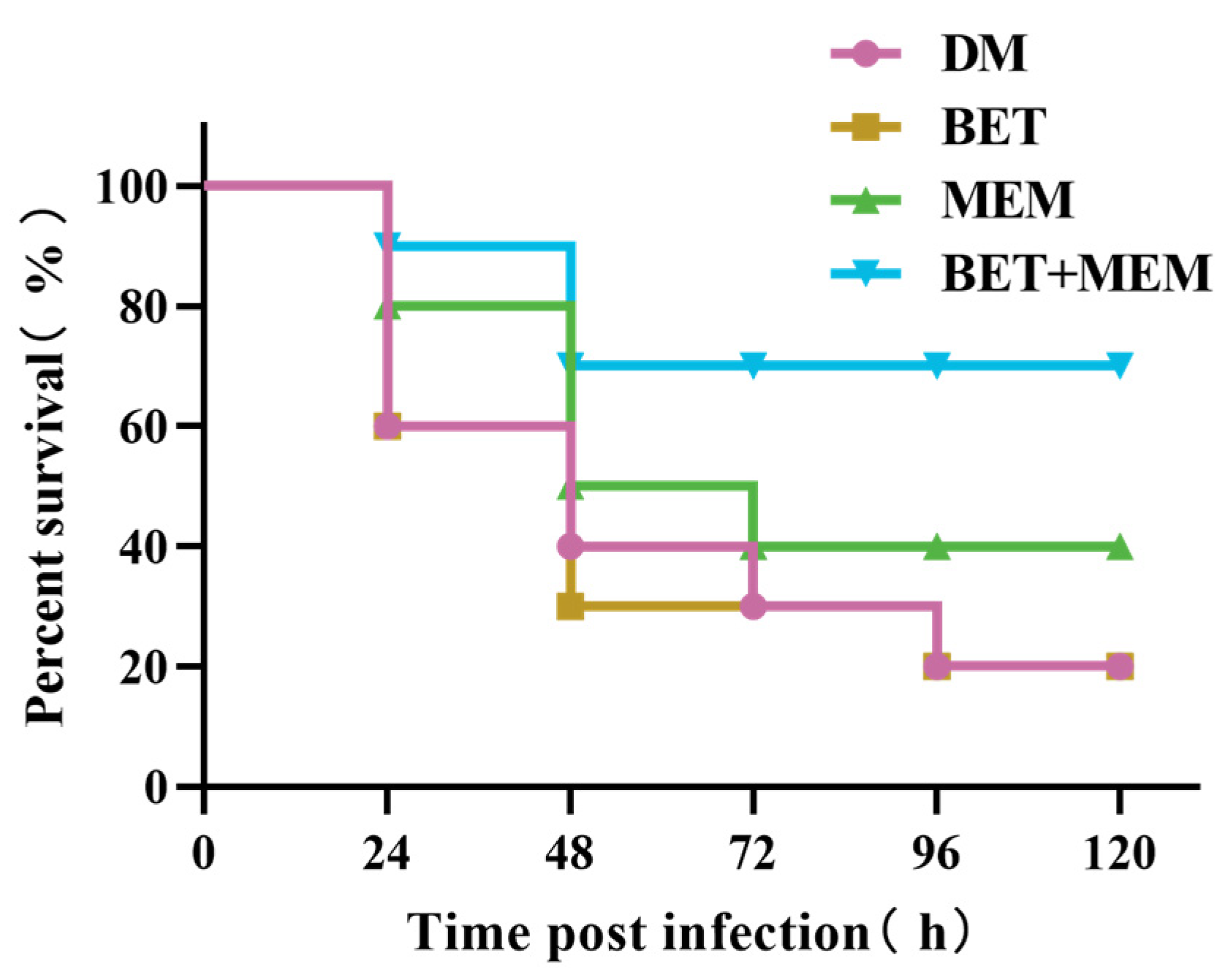
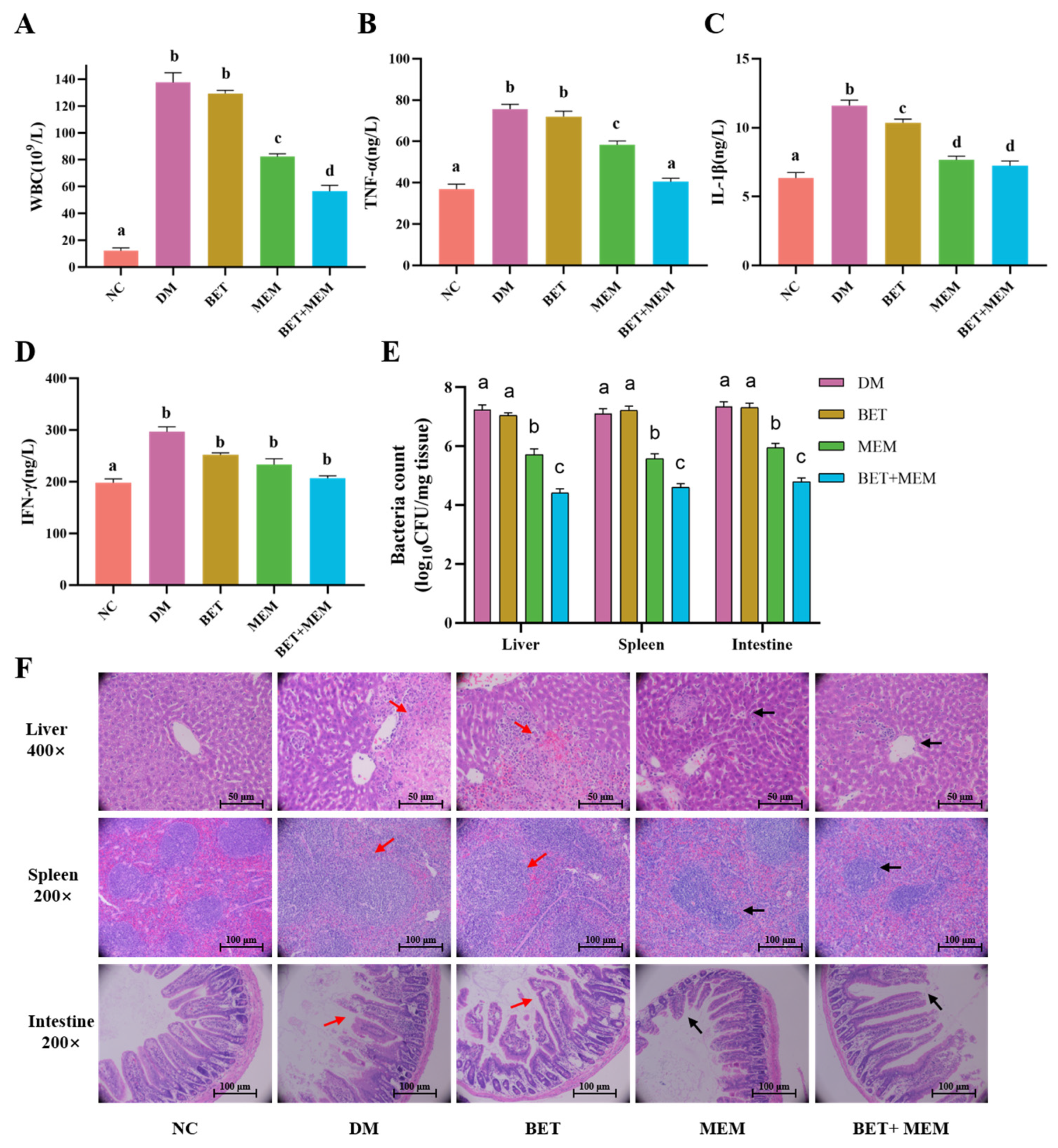
2.6. BET+MEM Treatment of NDM-1 E. coli-Infected Mice
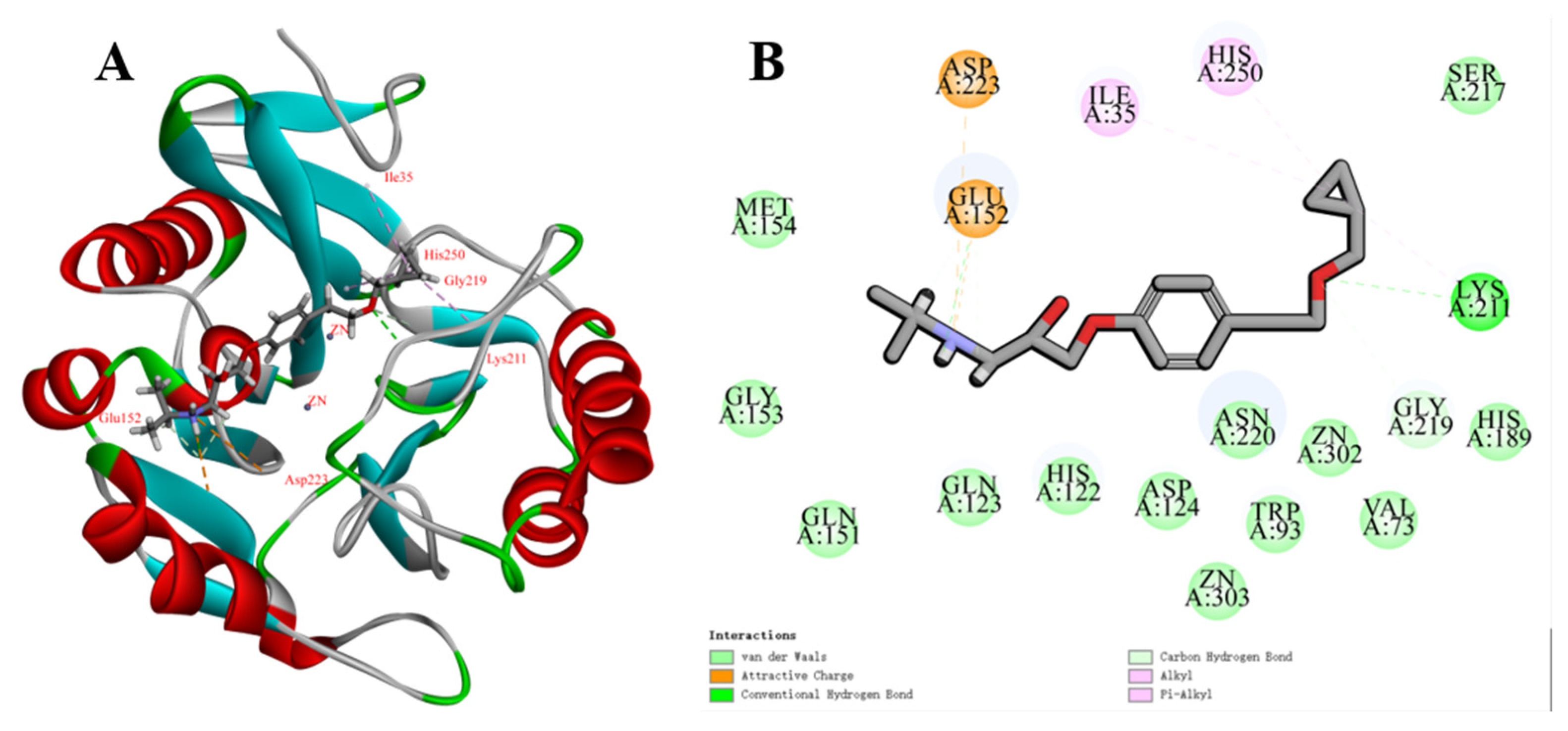
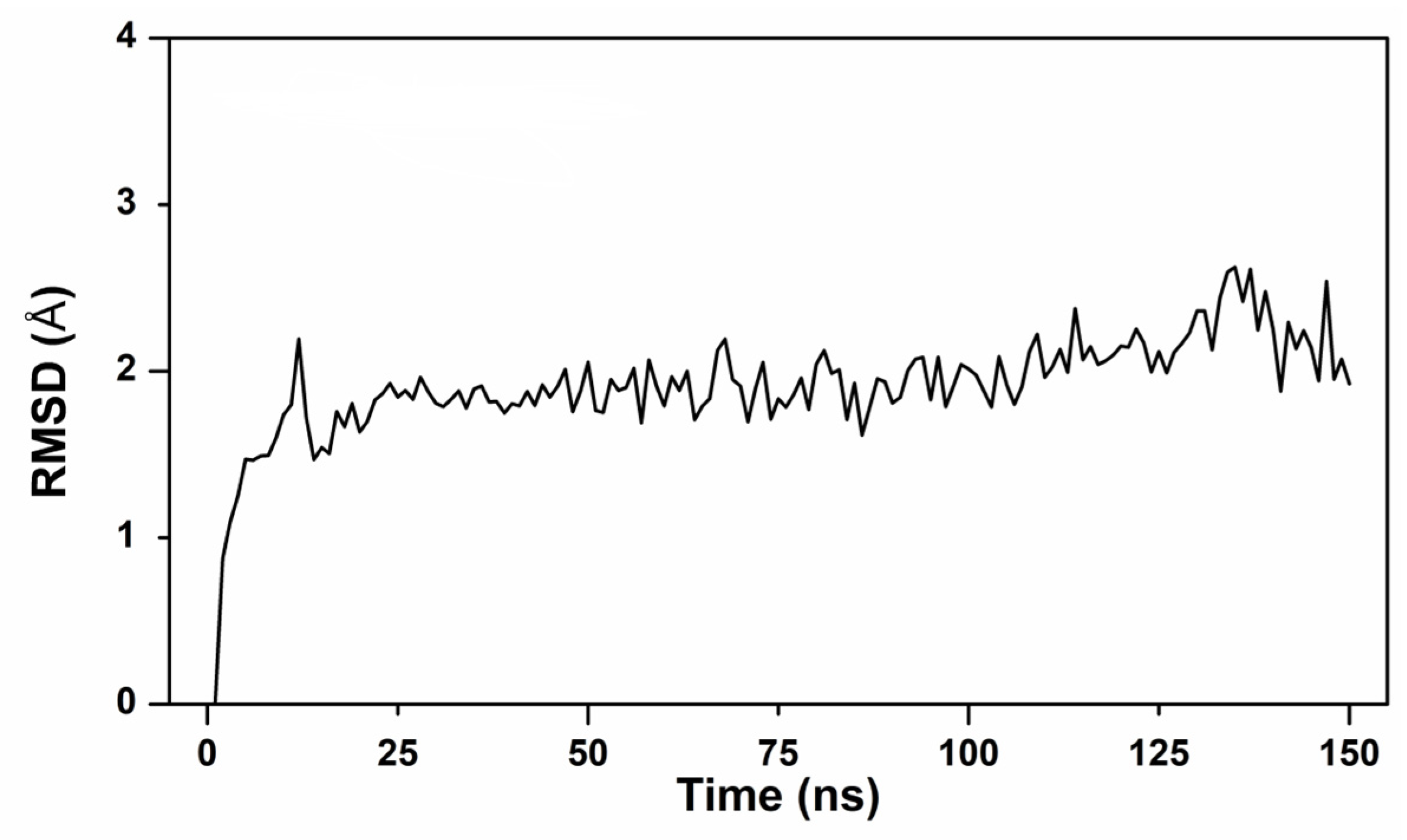
2.7. Molecular Docking and Kinetic Simulation Analysis
2.8. MM/GBSA Combined Free-Energy Calculations and Decompositions
3. Discussion
4. Materials and Methods
4.1. Strains, Cells, and Culture Conditions
4.2. Chemicals
4.3. Nitrocefin Assay and IC50 Assay
4.4. In Vitro Bacteriostatic Activity Determination
4.5. Bacterial Growth and Sterilization Curves
4.6. CCK-8 Assay
4.7. LDH Assay and Bacterial Count
4.8. Animal Research
4.9. Molecular Docking and MD Simulation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neu, H.C. beta-Lactam antibiotics: Structural relationships affecting in vitro activity and pharmacologic properties. Rev. Infect. Dis. 1986, 8 (Suppl. S3), S237–S259. [Google Scholar] [CrossRef]
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Gandra, S.; Ashok, A.; Caudron, Q.; Grenfell, B.T.; Levin, S.A.; Laxminarayan, R. Global antibiotic consumption 2000 to 2010: An analysis of national pharmaceutical sales data. Lancet Infect Dis. 2014, 14, 742–750. [Google Scholar] [CrossRef]
- Holmes, A.H.; Moore, L.S.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. Understanding the mechanisms and drivers of antimicrobial resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- King, D.T.; Worrall, L.J.; Gruninger, R.; Strynadka, N.C. New Delhi metallo-β-lactamase: Structural insights into β-lactam recognition and inhibition. J. Am. Chem. Soc. 2012, 134, 11362–11365. [Google Scholar] [CrossRef]
- Zavascki, A.P.; Barth, A.L.; Gonçalves, A.L.; Moro, A.L.; Fernandes, J.F.; Martins, A.F.; Ramos, F.; Goldani, L.Z. The influence of metallo-beta-lactamase production on mortality in nosocomial Pseudomonas aeruginosa infections. J. Antimicrob. Chemother. 2006, 58, 387–392. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, Y.H.; Schofield, C.J.; McNally, A.; Zong, Z.; Li, G.B. Metallo-β-lactamase-mediated antimicrobial resistance and progress in inhibitor discovery. Trends Microbiol. 2023, 31, 735–748. [Google Scholar] [CrossRef]
- Warrington, S.J.; Turner, P.; Kilborn, J.R.; Bianchetti, G.; Morselli, P.L. Blood concentrations and pharmacodynamic effects of betaxolol (SL 75212) a new beta-adrenoceptor antagonist after oral and intravenous administration. Br. J. Clin. Pharmacol. 1980, 10, 449–452. [Google Scholar] [CrossRef]
- Beresford, R.; Heel, R.C. Betaxolol. A review of its pharmacodynamic and pharmacokinetic properties, and therapeutic efficacy in hypertension. Drugs 1986, 31, 6–28. [Google Scholar] [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Present and Future of Carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics 2019, 8, 122. [Google Scholar] [CrossRef]
- Emeraud, C.; Escaut, L.; Boucly, A.; Fortineau, N.; Bonnin, R.A.; Naas, T.; Dortet, L. Aztreonam plus Clavulanate, Tazobactam, or Avibactam for Treatment of Infections Caused by Metallo-β-Lactamase-Producing Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2019, 63, e00010-19. [Google Scholar] [CrossRef]
- Akova, M. Sulbactam-containing beta-lactamase inhibitor combinations. Clin. Microbiol. Infect. 2008, 14 (Suppl. S1), 185–188. [Google Scholar] [CrossRef]
- Li, X.; Zhao, D.; Zhao, X.; Cheng, P.; Li, F.; Sun, J.; Li, Y.; Bai, J.; Zhang, X. The activity and mechanism of vidofludimus as a potent enzyme inhibitor against NDM-1-positive E. coli. Eur. J. Med. Chem. 2023, 250, 115225. [Google Scholar] [CrossRef]
- Shi, C.; Bao, J.; Sun, Y.; Kang, X.; Lao, X.; Zheng, H. Discovery of Baicalin as NDM-1 inhibitor: Virtual screening, biological evaluation and molecular simulation. Bioorg. Chem. 2019, 88, 102953. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, X.; Kong, F.; Xia, L.; Deng, X.; Wang, D.; Wang, J. Specific NDM-1 Inhibitor of Isoliquiritin Enhances the Activity of Meropenem against NDM-1-positive Enterobacteriaceae in vitro. Int. J. Environ. Res. Public. Health 2020, 17, 2162. [Google Scholar] [CrossRef]
- Steigerwalt, R.D., Jr.; Laurora, G.; Belcaro, G.V.; Cesarone, M.R.; De Sanctis, M.T.; Incandela, L.; Minicucci, R. Ocular and retrobulbar blood flow in ocular hypertensives treated with topical timolol, betaxolol and carteolol. J. Ocul. Pharmacol. Ther. 2001, 17, 537–544. [Google Scholar] [CrossRef]
- Arend, O.; Harris, A.; Arend, S.; Remky, A.; Martin, B.J. The acute effect of topical beta-adrenoreceptor blocking agents on retinal and optic nerve head circulation. Acta Ophthalmol. Scand. 1998, 76, 43–49. [Google Scholar] [CrossRef]
- Setoguchi, M.; Ohya, Y.; Abe, I.; Fujishima, M. Inhibitory action of betaxolol, a beta 1-selective adrenoceptor antagonist, on voltage-dependent calcium channels in guinea-pig artery and vein. Br. J. Pharmacol. 1995, 115, 198–202. [Google Scholar] [CrossRef]
- Bessho, H.; Suzuki, J.; Tobe, A. Vascular effects of betaxolol, a cardioselective beta-adrenoceptor antagonist, in isolated rat arteries. Jpn. J. Pharmacol. 1991, 55, 351–358. [Google Scholar] [CrossRef]
- Al-Wadei, M.J.; Bakheit, A.H.; Abdel-Aziz, A.A.; Wani, T.A. Betaxolol: A comprehensive profile. Profiles Drug Subst. Excip. Relat. Methodol. 2021, 46, 91–136. [Google Scholar] [CrossRef]
- Cheng, P.; Li, F.; Liu, R.; Yang, Y.; Xiao, T.; Ishfaq, M.; Xu, G.; Zhang, X. Prevalence and molecular epidemiology characteristics of carbapenem-resistant Escherichia coli in Heilongjiang Province, China. Infect. Drug Resist. 2019, 12, 2505–2518. [Google Scholar] [CrossRef]
- Shannon, K.; Phillips, I. Beta-Lactamase detection by three simple methods: Intralactam, nitrocefin and acidimetric. J. Antimicrob. Chemother. 1980, 6, 617–621. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Dieases (ESCMID). EUCAST Definitive Document E.Def 1.2, May 2000: Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect 2000, 6, 503–508. [Google Scholar] [CrossRef]
- Rizvi, M.; Ahmed, J.; Khan, F.; Shukla, I.; Malik, A. Assessment of combination therapy by time kill curve analysis and chequerboard assay for treatment of multi-drug resistant Pseudomonas aeruginosa isolates. J. Glob. Antimicrob. Resist. 2013, 1, 103–108. [Google Scholar] [CrossRef]
- Xu, T.; Ma, Q.; Li, Y.; Yu, Q.; Pan, P.; Zheng, Y.; Li, Z.; Xiong, X.; Hou, T.; Yu, B.; et al. A small molecule inhibitor of the UBE2F-CRL5 axis induces apoptosis and radiosensitization in lung cancer. Signal Transduct. Target. Ther. 2022, 7, 354. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 2018, 465–468. [Google Scholar] [CrossRef]
- Zhao, B.; Zhang, X.; Yu, T.; Liu, Y.; Zhang, X.; Yao, Y.; Feng, X.; Liu, H.; Yu, D.; Ma, L.; et al. Discovery of thiosemicarbazone derivatives as effective New Delhi metallo-β-lactamase-1 (NDM-1) inhibitors against NDM-1 producing clinical isolates. Acta Pharm. Sin. B 2021, 11, 203–221. [Google Scholar] [CrossRef]
- Yarlagadda, V.; Sarkar, P.; Samaddar, S.; Manjunath, G.B.; Mitra, S.D.; Paramanandham, K.; Shome, B.R.; Haldar, J. Vancomycin Analogue Restores Meropenem Activity against NDM-1 Gram-Negative Pathogens. ACS Infect. Dis. 2018, 4, 1093–1101. [Google Scholar] [CrossRef]
- Ma, B.; Fang, C.; Lu, L.; Wang, M.; Xue, X.; Zhou, Y.; Li, M.; Hu, Y.; Luo, X.; Hou, Z. The antimicrobial peptide thanatin disrupts the bacterial outer membrane and inactivates the NDM-1 metallo-β-lactamase. Nat. Commun. 2019, 10, 3517. [Google Scholar] [CrossRef]
- Kar, B.; Kundu, C.N.; Pati, S.; Bhattacharya, D. Discovery of phyto-compounds as novel inhibitors against NDM-1 and VIM-1 protein through virtual screening and molecular modelling. J. Biomol. Struct. Dyn. 2023, 41, 1267–1280. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.C.; Pinter, T.B.J.; Talley, D.C.; Baker-Agha, A.; Patel, D.; Smith, P.J.; Franz, K.J. Benzimidazole and Benzoxazole Zinc Chelators as Inhibitors of Metallo-β-Lactamase NDM-1. ChemMedChem 2021, 16, 654–661. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medicine | MIC (μg·mL−1) | FICI | Interaction Mode | |
|---|---|---|---|---|
| Applied Separately | Applied Combined | |||
| MEM | 64 | 8 | 0.25 | Synergistic |
| BET | 1024 | 128 | ||
| System | ΔEvdw | ΔEele | ΔGGB | ΔGSA | ΔGbind |
|---|---|---|---|---|---|
| BET-NDM-1 | −12.88 ± 3.63 | −107.33 ± 23.04 | 110.64 ± 14.21 | −1.74 ± 0.69 | −11.32 ± 4.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Ren, S.; Yang, Y.; Li, X.; Zhang, X. Betaxolol as a Potent Inhibitor of NDM-1-Positive E. coli That Synergistically Enhances the Anti-Inflammatory Effect in Combination with Meropenem. Int. J. Mol. Sci. 2023, 24, 13399. https://doi.org/10.3390/ijms241713399
Sun J, Ren S, Yang Y, Li X, Zhang X. Betaxolol as a Potent Inhibitor of NDM-1-Positive E. coli That Synergistically Enhances the Anti-Inflammatory Effect in Combination with Meropenem. International Journal of Molecular Sciences. 2023; 24(17):13399. https://doi.org/10.3390/ijms241713399
Chicago/Turabian StyleSun, Jichao, Shangjie Ren, Yaozu Yang, Xiaoting Li, and Xiuying Zhang. 2023. "Betaxolol as a Potent Inhibitor of NDM-1-Positive E. coli That Synergistically Enhances the Anti-Inflammatory Effect in Combination with Meropenem" International Journal of Molecular Sciences 24, no. 17: 13399. https://doi.org/10.3390/ijms241713399
APA StyleSun, J., Ren, S., Yang, Y., Li, X., & Zhang, X. (2023). Betaxolol as a Potent Inhibitor of NDM-1-Positive E. coli That Synergistically Enhances the Anti-Inflammatory Effect in Combination with Meropenem. International Journal of Molecular Sciences, 24(17), 13399. https://doi.org/10.3390/ijms241713399