
Single-Cell Analysis of Contractile Forces in iPSC-Derived Cardiomyocytes: Paving the Way for Precision Medicine in Cardiovascular Disease



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
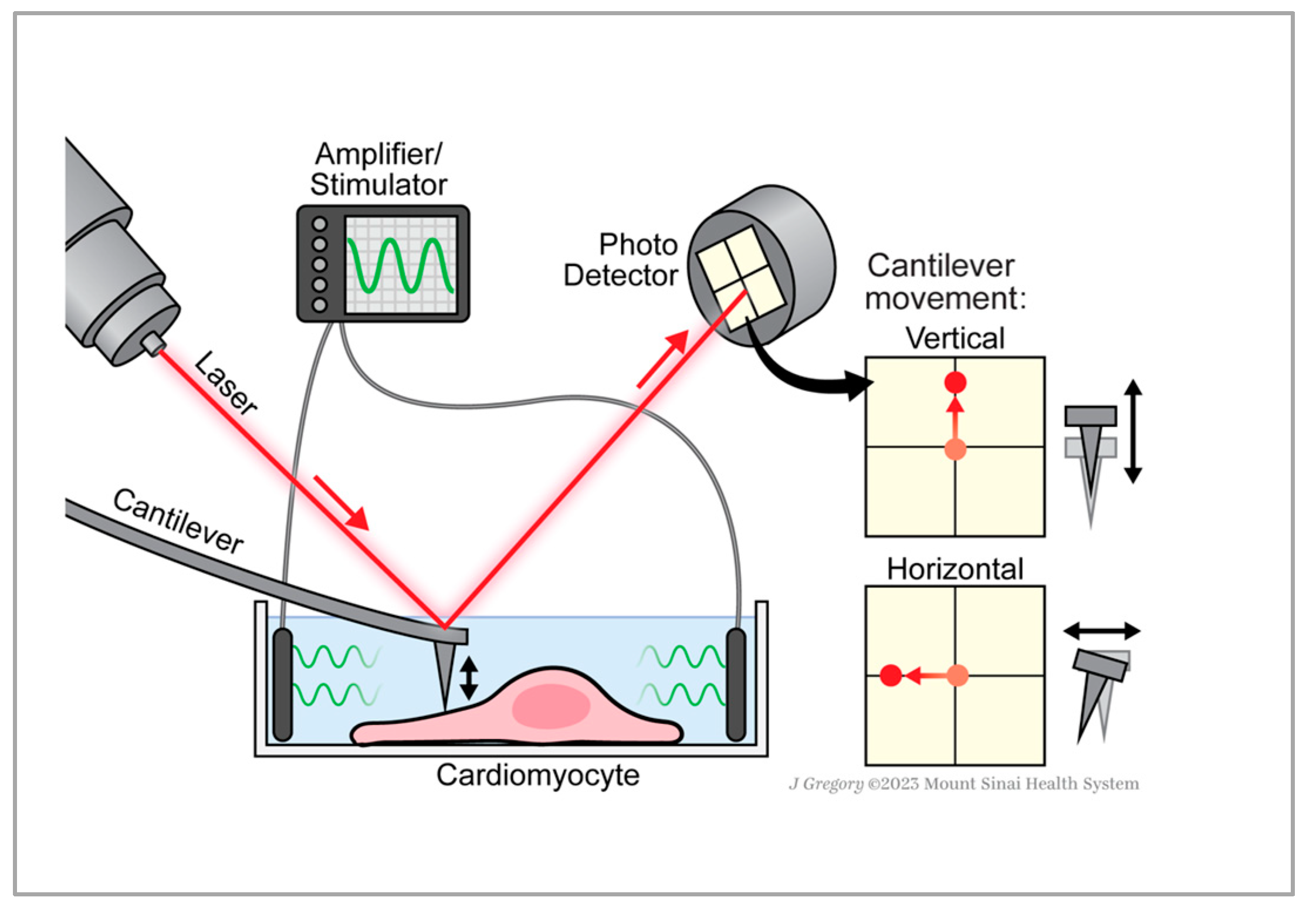
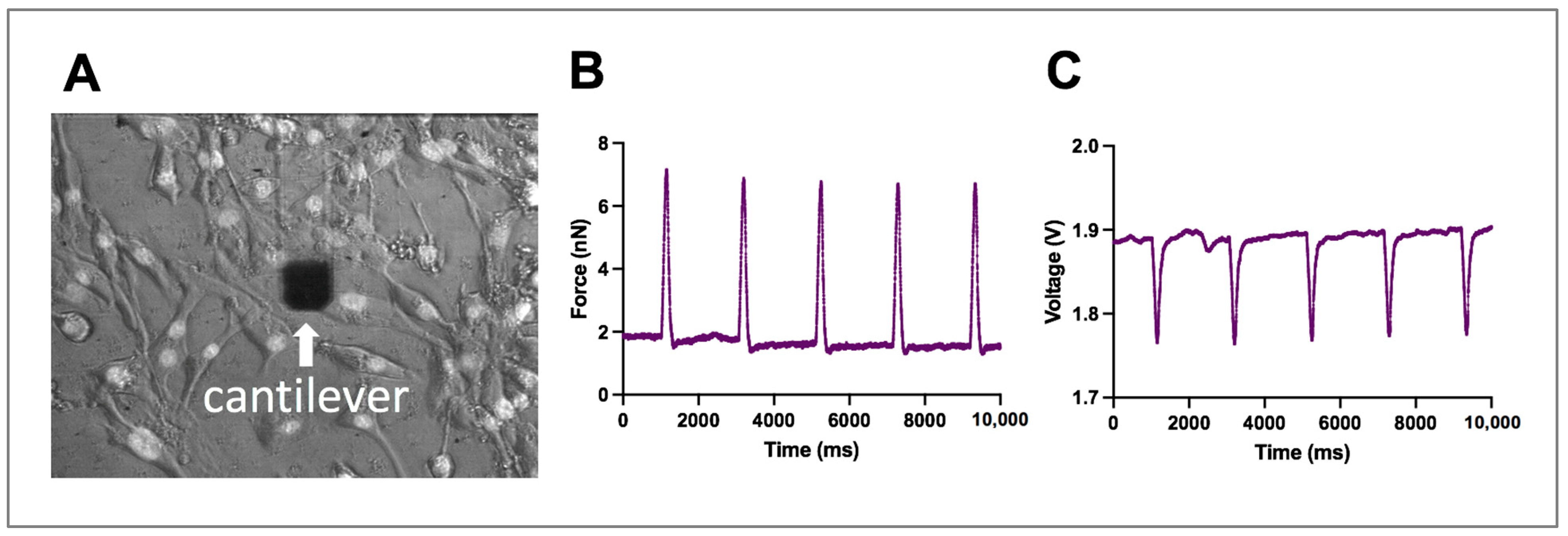
2.1. AFM Measurements Distinguish Normal and Lateral Force from Single-Cells
2.2. Normal and Lateral Force Correlation and Frequency Dependence
3. Discussion
4. Materials and Methods
4.1. Cell Source and Differentiation
4.2. Cell Preparation for AFM Experiments
4.3. AFM Measurements
4.4. Immunofluorescence of iPSC-CMs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Febbraro, F.; Chen, M.; Denham, M. Generation of human iPSCs by episomal reprogramming of skin fibroblasts and peripheral blood mononuclear cells. Methods Mol. Biol. 2021, 2239, 135–151. [Google Scholar] [PubMed]
- Mummery, C.L.; Zhang, J.; Ng, E.S.; Elliott, D.A.; Elefanty, A.G.; Kamp, T.J. Differentiation of human embryonic stem cells and induced pluripotent stem cells to cardiomyocytes. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Robinton, D.A.; Daley, G.Q. The promise of induced pluripotent stem cells in research and therapy. Nature 2012, 481, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Yamato, M.; Isoi, Y.; Akutsu, T.; Setomaru, T.; Abe, K.; Kikuchi, A.; Umezu, M.; Okano, T. Fabrication of pulsatile cardiac tissue grafts using a novel 3-dimensional cell sheet manipulation technique and temperature-responsive cell culture surfaces. Circ. Res. 2002, 90, e40. [Google Scholar] [CrossRef]
- Park, I.-H.; Arora, N.; Huo, H.; Maherali, N.; Ahfeldt, T.; Shimamura, A.; Lensch, M.W.; Cowan, C.; Hochedlinger, K.; Daley, G.Q. Disease-specific induced pluripotent stem cells. Cell 2008, 134, 877–886. [Google Scholar] [CrossRef]
- Bellin, M.; Casini, S.; Davis, R.P.; D’aniello, C.; Haas, J.; van Oostwaard, D.W.; Tertoolen, L.G.; Jung, C.B.; Elliott, D.A.; Welling, A. Isogenic human pluripotent stem cell pairs reveal the role of a KCNH2 mutation in long-QT syndrome. EMBO J. 2013, 32, 3161–3175. [Google Scholar] [CrossRef]
- Zhao, Q.; Wang, X.; Wang, S.; Song, Z.; Wang, J.; Ma, J. Cardiotoxicity evaluation using human embryonic stem cells and induced pluripotent stem cell-derived cardiomyocytes. Stem Cell Res. Ther. 2017, 8, 54. [Google Scholar] [CrossRef]
- van Meer, B.J.; Tertoolen, L.G.; Mummery, C.L. Concise review: Measuring physiological responses of human pluripotent stem cell derived cardiomyocytes to drugs and disease. Stem Cells 2016, 34, 2008–2015. [Google Scholar] [CrossRef]
- Tan, S.H.; Ye, L. Maturation of pluripotent stem cell-derived cardiomyocytes: A critical step for drug development and cell therapy. J. Cardiovasc. Transl. Res. 2018, 11, 375–392. [Google Scholar] [CrossRef]
- Jonsson, M.K.B.; Wang, Q.D.; Becker, B. Impedance-Based Detection of Beating Rhythm and Proarrhythmic Effects of Compounds on Stem Cell-Derived Cardiomyocytes. Assay Drug Dev. Technol. 2011, 9, 589–599. [Google Scholar] [CrossRef]
- Laverty, H.; Benson, C.; Cartwright, E.; Cross, M.; Garland, C.; Hammond, T.; Holloway, C.; McMahon, N.; Milligan, J.; Park, B. How can we improve our understanding of cardiovascular safety liabilities to develop safer medicines? Br. J. Pharmacol. 2011, 163, 675–693. [Google Scholar] [CrossRef] [PubMed]
- Redfern, W.; Carlsson, L.; Davis, A.; Lynch, W.; MacKenzie, I.; Palethorpe, S.; Siegl, P.; Strang, I.; Sullivan, A.; Wallis, R.; et al. Relationships between preclinical cardiac electrophysiology, clinical QT interval prolongation and torsade de pointes for a broad range of drugs: Evidence for a provisional safety margin in drug development. Cardiovasc. Res. 2003, 58, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, I.C.; Mayourian, J.; Murphy, J.F.; Stillitano, F.; Ceholski, D.K.; Costa, K.D. Cardiac tissue engineering models of inherited and acquired cardiomyopathies. In Experimental Models of Cardiovascular Diseases: Methods and Protocols; Springer: New York, NY, USA, 2018; pp. 145–159. [Google Scholar]
- Querdel, E.; Reinsch, M.; Castro, L.; Köse, D.; Bähr, A.; Reich, S.; Geertz, B.; Ulmer, B.; Schulze, M.; Lemoine, M.D. Human engineered heart tissue patches remuscularize the injured heart in a dose-dependent manner. Circulation 2021, 143, 1991–2006. [Google Scholar] [CrossRef] [PubMed]
- Goldfracht, I.; Efraim, Y.; Shinnawi, R.; Kovalev, E.; Huber, I.; Gepstein, A.; Arbel, G.; Shaheen, N.; Tiburcy, M.; Zimmermann, W.H. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 2019, 92, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Ronaldson-Bouchard, K.; Teles, D.; Yeager, K.; Tavakol, D.N.; Zhao, Y.; Chramiec, A.; Tagore, S.; Summers, M.; Stylianos, S.; Tamargo, M.; et al. A multi-organ chip with matured tissue niches linked by vascular flow. Nat. Biomed. Eng. 2022, 6, 351–371. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A platform for generation of chamber-specific cardiac tissues and disease modeling. Cell 2019, 176, 913–927.e18. [Google Scholar] [CrossRef]
- Abou-Saleh, H.; Zouein, F.A.; El-Yazbi, A.; Sanoudou, D.; Raynaud, C.; Rao, C.; Pintus, G.; Dehaini, H.; Eid, A.H. The march of pluripotent stem cells in cardiovascular regenerative medicine. Stem Cell Res. Ther. 2018, 9, 201. [Google Scholar] [CrossRef]
- Ichimura, H.; Chino, S.; Shiba, Y. Cardiac Regeneration Using Pluripotent Stem Cells and Controlling Immune Responses. Heart Lung Circ. 2023, 32, 836–843. [Google Scholar] [CrossRef]
- Borin, D.; Pecorari, I.; Pena, B.; Sbaizero, O. Novel insights into cardiomyocytes provided by atomic force microscopy. Semin. Cell Dev. Biol. 2018, 73, 4–12. [Google Scholar] [CrossRef]
- Liu, J.; Sun, N.; Bruce, M.A.; Wu, J.C.; Butte, M.J. Atomic force mechanobiology of pluripotent stem cell-derived cardiomyocytes. PLoS ONE 2012, 7, e37559. [Google Scholar] [CrossRef]
- Nagarajan, N.; Vyas, V.; Huey, B.D.; Zorlutuna, P. Modulation of the contractility of micropatterned myocardial cells with nanoscale forces using atomic force microscopy. Nanobiomedicine 2016, 3, 1849543516675348. [Google Scholar] [CrossRef]
- Vučković, S.; Dinani, R.; Nollet, E.E.; Kuster, D.W.; Buikema, J.W.; Houtkooper, R.H.; Nabben, M.; van der Velden, J.; Goversen, B. Characterization of cardiac metabolism in iPSC-derived cardiomyocytes: Lessons from maturation and disease modeling. Stem Cell Res. Ther. 2022, 13, 332. [Google Scholar] [CrossRef]
- Wu, P.; Deng, G.; Sai, X.; Guo, H.; Huang, H.; Zhu, P. Maturation strategies and limitations of induced pluripotent stem cell-derived cardiomyocytes. Biosci. Rep. 2021, 41, BSR20200833. [Google Scholar] [CrossRef] [PubMed]
- Maroli, G.; Braun, T. The long and winding road of cardiomyocyte maturation. Cardiovasc. Res. 2020, 117, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Gaitas, A.; Malhotra, R.; Li, T.; Herron, T.; Jalife, J. A device for rapid and quantitative measurement of cardiac myocyte contractility. Rev. Sci. Instrum. 2015, 86, 034302. [Google Scholar] [CrossRef] [PubMed]
- Gaitas, A.; Malhotra, R.; Pienta, K. A method to measure cellular adhesion utilizing a polymer micro-cantilever. Appl. Phys. Lett. 2013, 103, 123702. [Google Scholar] [CrossRef]
- Costa, K.D. Single-cell elastography: Probing for disease with the atomic force microscope. Dis. Markers 2004, 19, 139–154. [Google Scholar] [CrossRef]
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef]
- Turnbull, I.C.; Zhu, W.; Stillitano, F.; Chien, C.-C.; Gaitas, A. A micromachined force sensing apparatus and method for human engineered cardiac tissue and induced pluripotent stem cell characterization. Sens. Actuators A Phys. 2021, 331, 112874. [Google Scholar] [CrossRef]
- Tian, J.; Tu, C.; Huang, B.; Liang, Y.; Zhou, J.; Ye, X. Study of the union method of microelectrode array and AFM for the recording of electromechanical activities in living cardiomyocytes. Eur. Biophys. J. 2016, 46, 495–507. [Google Scholar] [CrossRef]
- Pesl, M.; Pribyl, J.; Acimovic, I.; Vilotic, A.; Jelinkova, S.; Salykin, A.; Lacampagne, A.; Dvorak, P.; Meli, A.C.; Skladal, P.; et al. Atomic force microscopy combined with human pluripotent stem cell derived cardiomyocytes for biomechanical sensing. Biosens. Bioelectron. 2016, 85, 751–757. [Google Scholar] [CrossRef]
- Pelling, A.E.; Nicholls, B.M.; Silberberg, Y.R.; Horton, M.A. Approaches for investigating mechanobiological dynamics in living cells with fluorescence and atomic force microscopies. Mod. Res. Educ. Top. Microsc. 2007, 3, 3–10. [Google Scholar]
- Smolyakov, G.; Cauquil, M.; Severac, C.; Lachaize, V.; Guilbeau-Frugier, C.; Sénard, J.-M.; Galés, C.; Dague, E. Biophysical properties of cardiomyocyte surface explored by multiparametric AFM. J. Struct. Biol. 2017, 198, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, E.; Hoes, M.; Ustyantsev, K.; Bomer, N.; de Jong, T.V.; van der Mei, H.; Berezikov, E.; van der Meer, P. Modeling human cardiac hypertrophy in stem cell-derived cardiomyocytes. Stem Cell Rep. 2018, 10, 794–807. [Google Scholar] [CrossRef] [PubMed]
- Germanguz, I.; Sedan, O.; Zeevi-Levin, N.; Shtrichman, R.; Barak, E.; Ziskind, A.; Eliyahu, S.; Meiry, G.; Amit, M.; Itskovitz-Eldor, J.; et al. Molecular characterization and functional properties of cardiomyocytes derived from human inducible pluripotent stem cells. J. Cell. Mol. Med. 2011, 15, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Del Álamo, J.C.; Lemons, D.; Serrano, R.; Savchenko, A.; Cerignoli, F.; Bodmer, R.; Mercola, M. High throughput physiological screening of iPSC-derived cardiomyocytes for drug development. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 1717–1727. [Google Scholar] [CrossRef]
- Shroff, S.G.; Saner, D.R.; Lal, R. Dynamic micromechanical properties of cultured rat atrial myocytes measured by atomic force microscopy. Am. J. Physiol.-Cell Physiol. 1995, 269, C286–C292. [Google Scholar] [CrossRef]
- Carpick, R.W.; Salmeron, M. Scratching the surface: Fundamental investigations of tribology with atomic force microscopy. Chem. Rev. 1997, 97, 1163–1194. [Google Scholar] [CrossRef]
- Peyronnet, R.; Desai, A.; Edelmann, J.-C.; Cameron, B.A.; Emig, R.; Kohl, P.; Dean, D. Simultaneous assessment of radial and axial myocyte mechanics by combining atomic force microscopy and carbon fibre techniques. Philos. Trans. R. Soc. B 2022, 377, 20210326. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, J.; Shi, L.; Niu, R.; Sun, Q.; Liu, H.; Li, J.; Guo, J.; Zhu, J.; Han, D. In situ mechanical analysis of cardiomyocytes at nano scales. Nanoscale 2012, 4, 99–102. [Google Scholar] [CrossRef]
- Crocini, C.; Gotthardt, M. Cardiac sarcomere mechanics in health and disease. Biophys. Rev. 2021, 13, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Bers, D. Excitation-Contraction Coupling and Cardiac Contractile Force; Springer: Dordrecht, The Netherlands, 2001; Volume 237. [Google Scholar]
- Peter, A.K.; Bjerke, M.A.; Leinwand, L.A. Biology of the cardiac myocyte in heart disease. Mol. Biol. Cell 2016, 27, 2149–2160. [Google Scholar] [CrossRef] [PubMed]
- Russell, B.; Motlagh, D.; Ashley, W.W.; Chopra, A.; Tabdanov, E.; Patel, H.; Janmey, P.A.; Kresh, J.Y.; Hartman, T.J.; Martin, J.L.; et al. Form follows function: How muscle shape is regulated by work. J. Appl. Physiol. 2000, 88, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Bartman, T.; Hove, J. Mechanics and function in heart morphogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2005, 233, 373–381. [Google Scholar] [CrossRef]
- Czubryt, M.P.; Olson, E.N. Balancing contractility and energy production: The role of myocyte enhancer factor 2 (MEF2) in cardiac hypertrophy. Rec. Prog. Horm. Res. 2004, 59, 105–124. [Google Scholar] [CrossRef]
- Silver, F.H.; Siperko, L.M.; Seehra, G.P. Mechanobiology of force transduction in dermal tissue. Skin Res. Technol. 2003, 9, 3–23. [Google Scholar] [CrossRef]
- Borau, C.; Kamm, R.D.; García-Aznar, J.M. Mechano-sensing and cell migration: A 3D model approach. Phys. Biol. 2011, 8, 066008. [Google Scholar] [CrossRef]
- Periasamy, M.; Kalyanasundaram, A. SERCA pump isoforms: Their role in calcium transport and disease. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 2007, 35, 430–442. [Google Scholar] [CrossRef]
- Schaub, M.; Hefti, M.A.; Harder, B.A.; Eppenberger, H.M. Various hypertrophic stimuli induce distinct phenotypes in cardiomyocytes. J. Mol. Med. 1997, 75, 901–920. [Google Scholar] [CrossRef]
- Saucerman, J.J.; Tan, P.M.; Buchholz, K.S.; McCulloch, A.D.; Omens, J.H. Mechanical regulation of gene expression in cardiac myocytes and fibroblasts. Nat. Rev. Cardiol. 2019, 16, 361–378. [Google Scholar] [CrossRef]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering adolescence: Maturation of human pluripotent stem cell–derived cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Galende, E.; Karakikes, I.; Edelmann, L.; Desnick, R.J.; Kerenyi, T.; Khoueiry, G.; Lafferty, J.; McGinn, J.T.; Brodman, M.; Fuster, V. Amniotic Fluid Cells Are More Efficiently Reprogrammed to Pluripotency than Adult Cells; Mary Ann Liebert, Inc.: New Rochelle, NY, USA, 2010. [Google Scholar]
- Oh, J.G.; Dave, J.; Kho, C.; Stillitano, F. Generation of Ventricular-Like HiPSC-Derived Cardiomyocytes and High-Quality Cell Preparations for Calcium Handling Characterization. J. Vis. Exp. 2020, 155, e60135. [Google Scholar]
- Murphy, J.F.; Mayourian, J.; Stillitano, F.; Munawar, S.; Broughton, K.M.; Agullo-Pascual, E.; Sussman, M.A.; Hajjar, R.J.; Costa, K.D.; Turnbull, I.C. Adult human cardiac stem cell supplementation effectively increases contractile function and maturation in human engineered cardiac tissues. Stem Cell Res. Ther. 2019, 10, 373. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turnbull, I.C.; Bajpai, A.; Jankowski, K.B.; Gaitas, A. Single-Cell Analysis of Contractile Forces in iPSC-Derived Cardiomyocytes: Paving the Way for Precision Medicine in Cardiovascular Disease. Int. J. Mol. Sci. 2023, 24, 13416. https://doi.org/10.3390/ijms241713416
Turnbull IC, Bajpai A, Jankowski KB, Gaitas A. Single-Cell Analysis of Contractile Forces in iPSC-Derived Cardiomyocytes: Paving the Way for Precision Medicine in Cardiovascular Disease. International Journal of Molecular Sciences. 2023; 24(17):13416. https://doi.org/10.3390/ijms241713416
Chicago/Turabian StyleTurnbull, Irene C., Apratim Bajpai, Katherine B. Jankowski, and Angelo Gaitas. 2023. "Single-Cell Analysis of Contractile Forces in iPSC-Derived Cardiomyocytes: Paving the Way for Precision Medicine in Cardiovascular Disease" International Journal of Molecular Sciences 24, no. 17: 13416. https://doi.org/10.3390/ijms241713416
APA StyleTurnbull, I. C., Bajpai, A., Jankowski, K. B., & Gaitas, A. (2023). Single-Cell Analysis of Contractile Forces in iPSC-Derived Cardiomyocytes: Paving the Way for Precision Medicine in Cardiovascular Disease. International Journal of Molecular Sciences, 24(17), 13416. https://doi.org/10.3390/ijms241713416