
Pro-Inflammatory Cytokine Priming and Purification Method Modulate the Impact of Exosomes Derived from Equine Bone Marrow Mesenchymal Stromal Cells on Equine Articular Chondrocytes

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Results
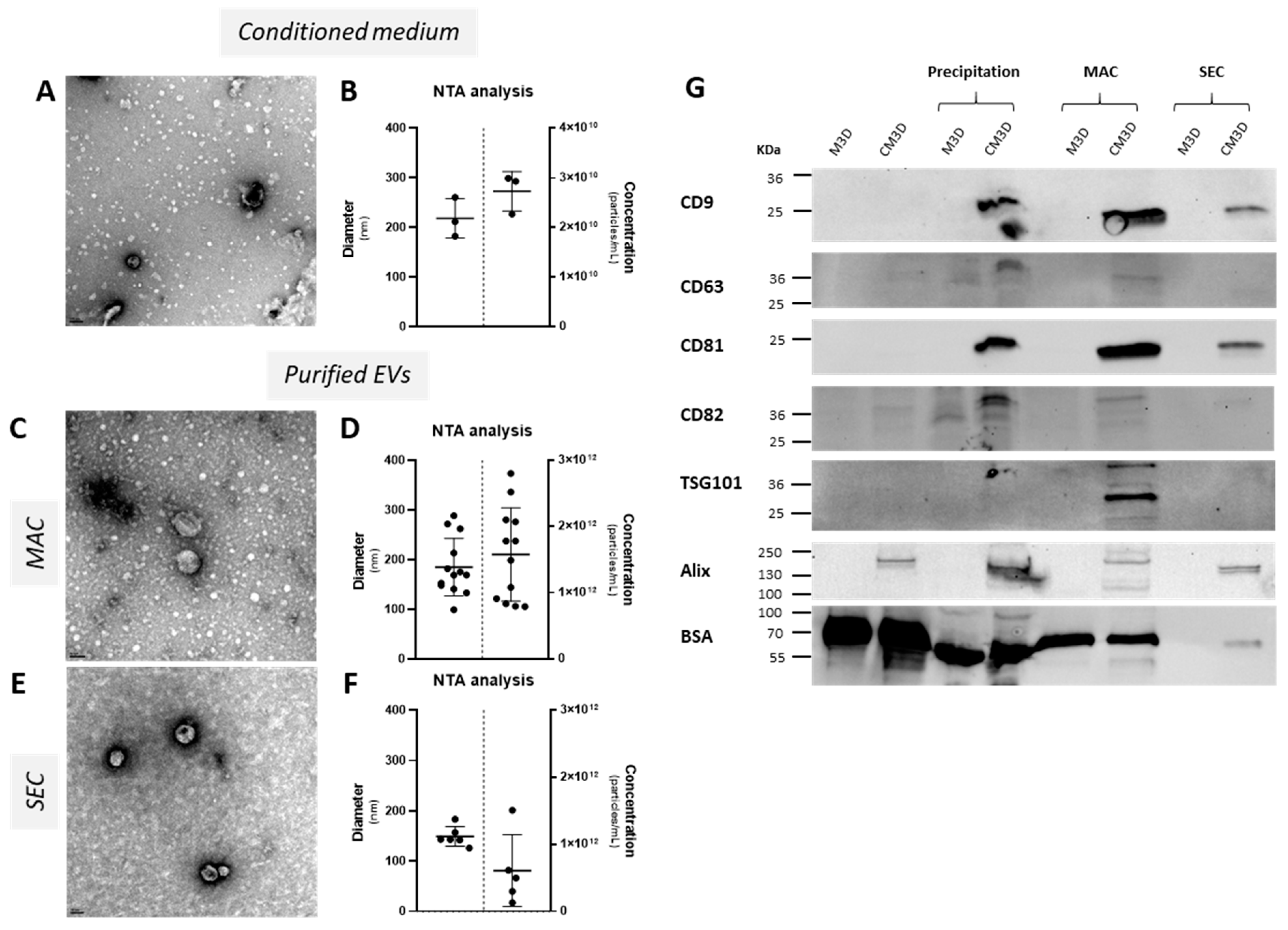
2.1. The Precipitation, MAC, and SEC Isolation Methods Effectively Purify Exosomes
2.2. Purified Exosomes Can Be Internalized in eACs
2.3. eACs Cultured with MAC-Exos Exhibit a Gene Expression Profile Closer to Hyaline Cartilage than eACs Cultured with SEC-Exos
2.4. MAC-Exos Induce a Greater Cartilage-Associated Protein Expression Profile in eACs than SEC-Exos
2.5. UF Does Not Improve MAC-Exos Potential for Enhancing eAC Phenotype
2.6. The Effect of Primed MSC-Derived Exosomes on eAC Gene Expression Is Masked by the Presence of the Elution Buffer
2.7. The Presence of the Elution Buffer Also Conceals the Impact of Primed MSC-Exos on the Synthesis of Healthy and OA Cartilage-Related Markers in eACs
2.8. Additional UF Step after MAC Purification Reveals the Effect of Naive and Primed Exosomes on the eAC Phenotype
3. Discussion
3.1. MSC-Exos Isolation and Characterization
3.2. Comparison between MAC and SEC Purification Strategies
3.3. ExoEasy Elution Buffer Composition
3.4. The Potential of Pro-Inflammatory Priming on the Therapeutic Effect of Exosomes from Equine BM-MSCs
3.5. Toward Mass Production
4. Materials and Methods
4.1. Cell Isolation and Culture
4.2. Priming and Medium Conditioning
4.3. Exosome Isolation with Polyethylene Glycol
4.4. Exosome Isolation by Membrane Affinity
4.5. Exosome Isolation by Size-Exclusion Chromatography
4.6. Exosome Protein Extraction and Assay
4.7. Exosome Characterization
4.8. Exosome Medium Supplementation
4.9. eAC Three-Dimensional Culture
4.10. eAC RNA Extraction and RT-qPCR
4.11. eAC Protein Extraction
4.12. Western Blot
4.13. PKH26 Labeling of Exosomes
4.14. Exosome Internalization by eACs
4.15. F-Actin Staining and Confocal Imaging
4.16. Nanoparticle Tracking Analysis
4.17. Transmission Electron Microscopy
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McIlwraith, C.W.; Frisbie, D.D.; Kawcak, C.E. The Horse as a Model of Naturally Occurring Osteoarthritis. Bone Joint. Res. 2012, 1, 297–309. [Google Scholar] [CrossRef]
- Aigner, T.; Stöve, J. Collagens—Major Component of the Physiological Cartilage Matrix, Major Target of Cartilage Degeneration, Major Tool in Cartilage Repair. Adv. Drug Deliv. Rev. 2003, 55, 1569–1593. [Google Scholar] [CrossRef] [PubMed]
- Armiento, A.R.; Alini, M.; Stoddart, M.J. Articular Fibrocartilage—Why Does Hyaline Cartilage Fail to Repair? Adv. Drug Deliv. Rev. 2019, 146, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, B.; Caldera, F.E. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. N. Am. 2020, 104, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Merimi, M.; El-Majzoub, R.; Lagneaux, L.; Moussa Agha, D.; Bouhtit, F.; Meuleman, N.; Fahmi, H.; Lewalle, P.; Fayyad-Kazan, M.; Najar, M. The Therapeutic Potential of Mesenchymal Stromal Cells for Regenerative Medicine: Current Knowledge and Future Understandings. Front. Cell. Dev. Biol. 2021, 9, 661532. [Google Scholar] [CrossRef]
- Han, Y.; Li, X.; Zhang, Y.; Han, Y.; Chang, F.; Ding, J. Mesenchymal Stem Cells for Regenerative Medicine. Cells 2019, 8, 886. [Google Scholar] [CrossRef]
- Chahal, J.; Gómez-Aristizábal, A.; Shestopaloff, K.; Bhatt, S.; Chaboureau, A.; Fazio, A.; Chisholm, J.; Weston, A.; Chiovitti, J.; Keating, A.; et al. Bone Marrow Mesenchymal Stromal Cell Treatment in Patients with Osteoarthritis Results in Overall Improvement in Pain and Symptoms and Reduces Synovial Inflammation. Stem Cells Transl. Med. 2019, 8, 746–757. [Google Scholar] [CrossRef]
- Harrell, C.R.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicles as New Remedies in the Therapy of Inflammatory Diseases. Cells 2019, 8, 1605. [Google Scholar] [CrossRef]
- Theeuwes, W.F.; van den Bosch, M.H.J.; Thurlings, R.M.; Blom, A.B.; van Lent, P.L.E.M. The Role of Inflammation in Mesenchymal Stromal Cell Therapy in Osteoarthritis, Perspectives for Post-Traumatic Osteoarthritis: A Review. Rheumatology 2021, 60, 1042–1053. [Google Scholar] [CrossRef]
- Song, Y.; Jorgensen, C. Mesenchymal Stromal Cells in Osteoarthritis: Evidence for Structural Benefit and Cartilage Repair. Biomedicines 2022, 10, 1278. [Google Scholar] [CrossRef]
- Fortier, L.A.; Potter, H.G.; Rickey, E.J.; Schnabel, L.V.; Foo, L.F.; Chong, L.R.; Stokol, T.; Cheetham, J.; Nixon, A.J. Concentrated Bone Marrow Aspirate Improves Full-Thickness Cartilage Repair Compared with Microfracture in the Equine Model. JBJS 2010, 92, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- McIlwraith, C.W.; Frisbie, D.D.; Rodkey, W.G.; Kisiday, J.D.; Werpy, N.M.; Kawcak, C.E.; Steadman, J.R. Evaluation of Intra-Articular Mesenchymal Stem Cells to Augment Healing of Microfractured Chondral Defects. Arthroscopy 2011, 27, 1552–1561. [Google Scholar] [CrossRef] [PubMed]
- Ferris, D.J.; Frisbie, D.D.; Kisiday, J.D.; McIlwraith, C.W.; Hague, B.A.; Major, M.D.; Schneider, R.K.; Zubrod, C.J.; Kawcak, C.E.; Goodrich, L.R. Clinical Outcome After Intra-Articular Administration of Bone Marrow Derived Mesenchymal Stem Cells in 33 Horses with Stifle Injury. Vet. Surg. 2014, 43, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Herberts, C.A.; Kwa, M.S.; Hermsen, H.P. Risk Factors in the Development of Stem Cell Therapy. J. Transl. Med. 2011, 9, 29. [Google Scholar] [CrossRef]
- Pigott, J.H.; Ishihara, A.; Wellman, M.L.; Russell, D.S.; Bertone, A.L. Investigation of the Immune Response to Autologous, Allogeneic, and Xenogeneic Mesenchymal Stem Cells after Intra-Articular Injection in Horses. Vet. Immunol. Immunopathol. 2013, 156, 99–106. [Google Scholar] [CrossRef]
- Paterson, Y.Z.; Rash, N.; Garvican, E.R.; Paillot, R.; Guest, D.J. Equine Mesenchymal Stromal Cells and Embryo-Derived Stem Cells Are Immune Privileged in Vitro. Stem Cell Res. Ther. 2014, 5, 90. [Google Scholar] [CrossRef]
- Schnabel, L.V.; Pezzanite, L.M.; Antczak, D.F.; Felippe, M.J.B.; Fortier, L.A. Equine Bone Marrow-Derived Mesenchymal Stromal Cells Are Heterogeneous in MHC Class II Expression and Capable of Inciting an Immune Response in Vitro. Stem Cell Res. Ther. 2014, 5, 13. [Google Scholar] [CrossRef]
- Bertoni, L.; Branly, T.; Jacquet, S.; Desancé, M.; Desquilbet, L.; Rivory, P.; Hartmann, D.-J.; Denoix, J.-M.; Audigié, F.; Galéra, P.; et al. Intra-Articular Injection of 2 Different Dosages of Autologous and Allogeneic Bone Marrow- and Umbilical Cord-Derived Mesenchymal Stem Cells Triggers a Variable Inflammatory Response of the Fetlock Joint on 12 Sound Experimental Horses. Stem Cells Int. 2019, 17. [Google Scholar] [CrossRef]
- Wang, Y.; Huso, D.L.; Harrington, J.; Kellner, J.; Jeong, D.K.; Turney, J.; McNiece, I.K. Outgrowth of a Transformed Cell Population Derived from Normal Human BM Mesenchymal Stem Cell Culture. Cytotherapy 2005, 7, 509–519. [Google Scholar] [CrossRef]
- Pan, Q.; Fouraschen, S.M.G.; de Ruiter, P.E.; Dinjens, W.N.M.; Kwekkeboom, J.; Tilanus, H.W.; van der Laan, L.J.W. Detection of Spontaneous Tumorigenic Transformation during Culture Expansion of Human Mesenchymal Stromal Cells. Exp. Biol. Med. 2014, 239, 105–115. [Google Scholar] [CrossRef]
- Contentin, R.; Jammes, M.; Bourdon, B.; Cassé, F.; Bianchi, A.; Audigié, F.; Branly, T.; Velot, É.; Galéra, P. Bone Marrow MSC Secretome Increases Equine Articular Chondrocyte Collagen Accumulation and Their Migratory Capacities. Int. J. Mol. Sci. 2022, 23, 5795. [Google Scholar] [CrossRef] [PubMed]
- Jammes, M.; Contentin, R.; Cassé, F.; Galéra, P. Equine Osteoarthritis: Strategies to Enhance Mesenchymal Stromal Cell-Based Acellular Therapies. Front. Vet. Sci. 2023, 10, 1115774. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Kim, J.; Kim, M.Y.; Bae, Y.-S.; Ryu, S.H.; Lee, T.G.; Kim, J.H. Proteomic Analysis of Tumor Necrosis Factor-Alpha-Induced Secretome of Human Adipose Tissue-Derived Mesenchymal Stem Cells. J. Proteome Res. 2010, 9, 1754–1762. [Google Scholar] [CrossRef] [PubMed]
- Jammes, M.; Contentin, R.; Audigié, F.; Cassé, F.; Galéra, P. Effect of Pro-Inflammatory Cytokine Priming and Storage Temperature of the Mesenchymal Stromal Cell (MSC) Secretome on Equine Articular Chondrocytes. Front. Bioeng. Biotechnol. 2023, 11, 1204737. [Google Scholar] [CrossRef]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [PubMed]
- Maumus, M.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cells in Regenerative Medicine Applied to Rheumatic Diseases: Role of Secretome and Exosomes. Biochimie 2013, 95, 2229–2234. [Google Scholar] [CrossRef]
- Toh, W.S.; Lai, R.C.; Hui, J.H.P.; Lim, S.K. MSC Exosome as a Cell-Free MSC Therapy for Cartilage Regeneration: Implications for Osteoarthritis Treatment. Semin. Cell. Dev. Biol. 2017, 67, 56–64. [Google Scholar] [CrossRef]
- Boulestreau, J.; Maumus, M.; Jorgensen, C.; Noël, D. Extracellular Vesicles from Mesenchymal Stromal Cells: Therapeutic Perspectives for Targeting Senescence in Osteoarthritis. Adv. Drug Deliv. Rev. 2021, 175, 113836. [Google Scholar] [CrossRef]
- Stranska, R.; Gysbrechts, L.; Wouters, J.; Vermeersch, P.; Bloch, K.; Dierickx, D.; Andrei, G.; Snoeck, R. Comparison of Membrane Affinity-Based Method with Size-Exclusion Chromatography for Isolation of Exosome-like Vesicles from Human Plasma. J. Transl. Med. 2018, 16, 1. [Google Scholar] [CrossRef]
- Patel, G.K.; Khan, M.A.; Zubair, H.; Srivastava, S.K.; Khushman, M.; Singh, S.; Singh, A.P. Comparative Analysis of Exosome Isolation Methods Using Culture Supernatant for Optimum Yield, Purity and Downstream Applications. Sci. Rep. 2019, 9, 5335. [Google Scholar] [CrossRef]
- Liu, D.S.K.; Upton, F.M.; Rees, E.; Limb, C.; Jiao, L.R.; Krell, J.; Frampton, A.E. Size-Exclusion Chromatography as a Technique for the Investigation of Novel Extracellular Vesicles in Cancer. Cancers 2020, 12, 3156. [Google Scholar] [CrossRef] [PubMed]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of Exosomes by Differential Centrifugation: Theoretical Analysis of a Commonly Used Protocol. Sci. Rep. 2015, 5, 17319. [Google Scholar] [CrossRef] [PubMed]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. Biomed. Res. Int. 2018, e8545347. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Haylett, A.K.; Moore, J.V. Comparative Analysis of Foetal Calf and Human Low Density Lipoprotein: Relevance for Pharmacodynamics of Photosensitizers. J. Photochem. Photobiol. B 2002, 66, 171–178. [Google Scholar] [CrossRef]
- ExoEasy Maxi Kit. Available online: https://www.qiagen.com/us/products/discovery-and-translational-research/exosomes-ctcs/exosomes/exoeasy-maxi-kit (accessed on 3 July 2023).
- Almiñana, C.; Rudolf Vegas, A.; Tekin, M.; Hassan, M.; Uzbekov, R.; Fröhlich, T.; Bollwein, H.; Bauersachs, S. Isolation and Characterization of Equine Uterine Extracellular Vesicles: A Comparative Methodological Study. Int. J. Mol. Sci. 2021, 22, 979. [Google Scholar] [CrossRef]
- Mann, M.; Jensen, O.N. Proteomic Analysis of Post-Translational Modifications. Nat. Biotechnol. 2003, 21, 255–261. [Google Scholar] [CrossRef]
- Vilette, D.; Laulagnier, K.; Huor, A.; Alais, S.; Simoes, S.; Maryse, R.; Provansal, M.; Lehmann, S.; Andreoletti, O.; Schaeffer, L.; et al. Efficient Inhibition of Infectious Prions Multiplication and Release by Targeting the Exosomal Pathway. Cell. Mol. Life Sci. 2015, 72, 4409–4427. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Y.; Wang, H.; Zhu, Z.; Xiao, Z. Visualizing of the Cellular Uptake and Intracellular Trafficking of Exosomes by Live-Cell Microscopy. J. Cell. Biochem. 2010, 111, 488–496. [Google Scholar] [CrossRef]
- Pham, C.V.; Midge, S.; Barua, H.; Zhang, Y.; Ngoc-Gia Nguyen, T.; Barrero, R.A.; Duan, A.; Yin, W.; Jiang, G.; Hou, Y.; et al. Bovine Extracellular Vesicles Contaminate Human Extracellular Vesicles Produced in Cell Culture Conditioned Medium When “exosome-Depleted Serum” Is Utilised. Arch. Biochem. Biophys. 2021, 708, 108963. [Google Scholar] [CrossRef]
- Heidarzadeh, M.; Zarebkohan, A.; Rahbarghazi, R.; Sokullu, E. Protein Corona and Exosomes: New Challenges and Prospects. Cell Commun. Signal. 2023, 21, 64. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.; Poupardin, R.W.; Ebner-Peking, P.; Andrade, A.C.; Blöchl, C.; Obermayer, A.; Gomes, F.G.; Vari, B.; Maeding, N.; Eminger, E.; et al. A Functional Corona around Extracellular Vesicles Enhances Angiogenesis, Skin Regeneration and Immunomodulation. J. Extracell. Vesicles 2022, 11, e12207. [Google Scholar] [CrossRef] [PubMed]
- Giannasi, C.; Niada, S.; Magagnotti, C.; Ragni, E.; Andolfo, A.; Brini, A.T. Comparison of Two ASC-Derived Therapeutics in an in Vitro OA Model: Secretome versus Extracellular Vesicles. Stem Cell Res. Ther. 2020, 11, 521. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Yoo, S.M.; Park, H.H.; Lim, H.J.; Kim, Y.-L.; Lee, S.; Seo, K.-W.; Kang, K.-S. Exosomes Derived from Human Umbilical Cord Blood Mesenchymal Stem Cells Stimulates Rejuvenation of Human Skin. Biochem. Biophys. Res. Commun. 2017, 493, 1102–1108. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Ma, C.; Wang, G.; Zhang, Y.; Sun, S. Exosomes Derived from Platelet-Rich Plasma Present a Novel Potential in Alleviating Knee Osteoarthritis by Promoting Proliferation and Inhibiting Apoptosis of Chondrocyte via Wnt/β-Catenin Signaling Pathway. J. Orthop. Surg. Res. 2019, 14, 470. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cong, M.; Huang, W.; Chen, J.; Zhang, M.; Gu, X.; Sun, C.; Yang, H. The Effect of Human Bone Marrow Mesenchymal Stem Cell-Derived Exosomes on Cartilage Repair in Rabbits. Stem Cells Int. 2022, 2022, 5760107. [Google Scholar] [CrossRef] [PubMed]
- Nikfarjam, S.; Rezaie, J.; Zolbanin, N.M.; Jafari, R. Mesenchymal Stem Cell Derived-Exosomes: A Modern Approach in Translational Medicine. J. Transl. Med. 2020, 18, 449. [Google Scholar] [CrossRef]
- Kim, Y.G.; Choi, J.; Kim, K. Mesenchymal Stem Cell-Derived Exosomes for Effective Cartilage Tissue Repair and Treatment of Osteoarthritis. Biotechnol. J. 2020, 15, e2000082. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Dao, H.H.; Duong, C.M.; Nguyen, X.-H.; Hoang, D.H.; Do, X.-H.; Truong, T.Q.; Nguyen, T.D.; Nguyen, L.T.; Than, U.T.T. Cytokine-Primed Umbilical Cord Mesenchymal Stem Cells Enhanced Therapeutic Effects of Extracellular Vesicles on Osteoarthritic Chondrocytes. Front. Immunol. 2022, 13, 1041592. [Google Scholar] [CrossRef]
- Andrews, S.H.; Klinker, M.W.; Bauer, S.R.; Marklein, R.A. Morphological Landscapes from High Content Imaging Reveal Cytokine Priming Strategies That Enhance Mesenchymal Stromal Cell Immunosuppression. Biotechnol. Bioeng. 2022, 119, 361–375. [Google Scholar] [CrossRef]
- De Angelis, E.; Cacchioli, A.; Ravanetti, F.; Bileti, R.; Cavalli, V.; Martelli, P.; Borghetti, P. Gene Expression Markers in Horse Articular Chondrocytes: Chondrogenic Differentiaton IN VITRO Depends on the Proliferative Potential and Ageing. Implication for Tissue Engineering of Cartilage. Res. Vet. Sci. 2020, 128, 107–117. [Google Scholar] [CrossRef]
- Costa, L.A.; Eiro, N.; Fraile, M.; Gonzalez, L.O.; Saá, J.; Garcia-Portabella, P.; Vega, B.; Schneider, J.; Vizoso, F.J. Functional Heterogeneity of Mesenchymal Stem Cells from Natural Niches to Culture Conditions: Implications for Further Clinical Uses. Cell. Mol. Life Sci. 2021, 78, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Szabó, E.; Fajka-Boja, R.; Kriston-Pál, É.; Hornung, Á.; Makra, I.; Kudlik, G.; Uher, F.; Katona, R.L.; Monostori, É.; Czibula, Á. Licensing by Inflammatory Cytokines Abolishes Heterogeneity of Immunosuppressive Function of Mesenchymal Stem Cell Population. Stem Cells Dev. 2015, 24, 2171–2180. [Google Scholar] [CrossRef]
- Chen, T.S.; Arslan, F.; Yin, Y.; Tan, S.S.; Lai, R.C.; Choo, A.B.H.; Padmanabhan, J.; Lee, C.N.; de Kleijn, D.P.; Lim, S.K. Enabling a Robust Scalable Manufacturing Process for Therapeutic Exosomes through Oncogenic Immortalization of Human ESC-Derived MSCs. J. Transl. Med. 2011, 9, 47. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Mo, Y.; Chen, Z.; Chen, T.; Li, Y.; Zheng, Y.; Deng, S.; Xu, X.; Chen, H.; et al. Immortalized Mesenchymal Stem Cells: A Safe Cell Source for Cellular or Cell Membrane-Based Treatment of Glioma. Stem Cells Int. 2022, 6430565. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Miller, R.; Stoppato, M.; Sere, Y.Y.; Coles, A.; Didiot, M.-C.; Wollacott, R.; Sapp, E.; Dubuke, M.L.; Li, X.; et al. Exosomes Produced from 3D Cultures of MSCs by Tangential Flow Filtration Show Higher Yield and Improved Activity. Mol. Ther. 2018, 26, 2838–2847. [Google Scholar] [CrossRef] [PubMed]
- Branly, T.; Bertoni, L.; Contentin, R.; Rakic, R.; Gomez-Leduc, T.; Desancé, M.; Hervieu, M.; Legendre, F.; Jacquet, S.; Audigié, F.; et al. Characterization and Use of Equine Bone Marrow Mesenchymal Stem Cells in Equine Cartilage Engineering. Study of Their Hyaline Cartilage Forming Potential When Cultured under Hypoxia within a Biomaterial in the Presence of BMP-2 and TGF-SS1. Stem Cell. Rev. Rep. 2017, 13, 611–630. [Google Scholar] [CrossRef]
- Branly, T.; Contentin, R.; Desancé, M.; Jacquel, T.; Bertoni, L.; Jacquet, S.; Mallein-Gerin, F.; Denoix, J.-M.; Audigié, F.; Demoor, M.; et al. Improvement of the Chondrocyte-Specific Phenotype upon Equine Bone Marrow Mesenchymal Stem Cell Differentiation: Influence of Culture Time, Transforming Growth Factors and Type I Collagen SiRNAs on the Differentiation Index. Int. J. Mol. Sci. 2018, 19, 435. [Google Scholar] [CrossRef] [PubMed]
- Desancé, M.; Contentin, R.; Bertoni, L.; Gomez-Leduc, T.; Branly, T.; Jacquet, S.; Betsch, J.-M.; Batho, A.; Legendre, F.; Audigié, F.; et al. Chondrogenic Differentiation of Defined Equine Mesenchymal Stem Cells Derived from Umbilical Cord Blood for Use in Cartilage Repair Therapy. Int. J. Mol. Sci. 2018, 19, 537. [Google Scholar] [CrossRef]
- Contentin, R.; Demoor, M.; Concari, M.; Desancé, M.; Audigié, F.; Branly, T.; Galéra, P. Comparison of the Chondrogenic Potential of Mesenchymal Stem Cells Derived from Bone Marrow and Umbilical Cord Blood Intended for Cartilage Tissue Engineering. Stem Cell. Rev. Rep. 2020, 16, 126–143. [Google Scholar] [CrossRef]
- Rakic, R.; Bourdon, B.; Hervieu, M.; Branly, T.; Legendre, F.; Saulnier, N.; Audigié, F.; Maddens, S.; Demoor, M.; Galera, P. RNA Interference and BMP-2 Stimulation Allows Equine Chondrocytes Redifferentiation in 3D-Hypoxia Cell Culture Model: Application for Matrix-Induced Autologous Chondrocyte Implantation. Int. J. Mol. Sci. 2017, 18, 1842. [Google Scholar] [CrossRef] [PubMed]
- Xi, X.-M.; Xia, S.-J.; Lu, R. Drug Loading Techniques for Exosome-Based Drug Delivery Systems. Pharmazie 2021, 76, 61–67. [Google Scholar] [PubMed]
- Yin, Y.; Han, X.; Li, C.; Sun, T.; Li, K.; Liu, X.; Liu, M. The Status of Industrialization and Development of Exosomes as a Drug Delivery System: A Review. Front. Pharmacol. 2022, 13, 961127. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| ACAN | TGT CAA CAA CAA TGC CCA AGA C | CTT CTT CCG CCC AAA GGT CC |
| ADAMTS5 | AAG GGA CAC CAT GTG GCAA A | CCC ACA TGA GCG AGA ACA CT |
| β-ACTIN | GAT GAT GAT ATC GCC GCG CTC | TGC CCC ACG TAT GAG TCC TT |
| COL10A1 | GCA CCC CAG TAA TGT ACA CCT ATG | GAG CCA CAC CTG GTC ATT TTC |
| COL1A1 | TGC CGT GAC CTC AAG ATG TG | CGT CTC CAT GTT GCA GAA GA |
| COL1A2 | CCA GAG TGG AGC AGC GGT TA | GGG ATG TTT TCA GGT TGA GCC |
| COL2A1 | GGC AAT AGC AGG TTC ACG TAC A | CGA TAA CAG TCT TGC CCC ACT T |
| HTRA1 | GGA CTT CAT GTT TCC CTC AA | GTT CTG CTG AAC AAG CAA CA |
| MMP1 | CGA AGG GAA CCC TCG GTG GGA | TGG CCT GGT CCA CAT CTG CTC |
| MMP13 | TGA AGA CCC GAA CCC TAA ACA T | GAA GAC TGG TGA TGG CAT CAA G |
| P53 | CAC CTG AGG TTG GCT CTG AC | GCA CAA ACA CGC ACC TCA AA |
| P65 | CAC GGA TAC CAC CAA GAC CC | GTC TGG ATG CGC TGA CTG AT |
| PPIA | CCC TAC CGT GTT CTT CGA CA | GTG AAG TCA CCA CCC TGA CA |
| PRG4 | CTA CCA CCC AAC GCA ACA AA | ACT GTT GTC TCC TTA TTG GGT |
| Antibody | Dilution | Supplier |
|---|---|---|
| Mouse anti-bovine serum albumin (BSA) | 0.1 µg/mL | Santa Cruz Biotechnology, Dallas, TX, USA |
| Mouse anti-human CD9 | 2 µg/mL | |
| Mouse anti-human CD63 | 1 µg/mL | |
| Mouse anti-human CD81 | 0.1 µg/mL | |
| Mouse anti-human Alix | 1 µg/mL | Covalab, Villeurbanne, France |
| Mouse anti-human Tsg101 | 1 µg/mL | |
| Mouse anti-equine CD82 | 10 µg/mL | |
| HRP-conjugated goat anti-mouse IgG antibody | 16 ng/mL | Jackson Immunoresearch, West Grove, PA, USA |
| Antibody | Dilution | Supplier |
|---|---|---|
| Rabbit anti-bovine type I collagen | 1:3000 | Novotec, Bron, France |
| Rabbit anti-human type II collagen | 1:750 | |
| Mouse anti-human type X collagen | 1:1000 | Sigma-Aldrich, Saint Louis, MO, USA |
| Rabbit anti-human GAPDH | 1:3000 | Santa Cruz Biotechnology, Dallas, TX, USA |
| Mouse anti-human PCNA | 1:1000 | |
| Rabbit anti-human Htra1 | 1:3000 | Merck Millipore, Billerica, MA, USA |
| Rabbit anti-human type IIB collagen | 1:750 | Covalab, Villeurbanne, France |
| Rabbit anti-human MMP1 | 1:1000 | Affinity Biosciences, Melbourne, VIC, Australia |
| HRP-conjugated goat anti-rabbit antibody | 1:5000 | Jackson Immunoresearch, West Grove, PA, USA |
| HRP-conjugated goat anti-mouse antibody | 1:5000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jammes, M.; Cassé, F.; Velot, E.; Bianchi, A.; Audigié, F.; Contentin, R.; Galéra, P. Pro-Inflammatory Cytokine Priming and Purification Method Modulate the Impact of Exosomes Derived from Equine Bone Marrow Mesenchymal Stromal Cells on Equine Articular Chondrocytes. Int. J. Mol. Sci. 2023, 24, 14169. https://doi.org/10.3390/ijms241814169
Jammes M, Cassé F, Velot E, Bianchi A, Audigié F, Contentin R, Galéra P. Pro-Inflammatory Cytokine Priming and Purification Method Modulate the Impact of Exosomes Derived from Equine Bone Marrow Mesenchymal Stromal Cells on Equine Articular Chondrocytes. International Journal of Molecular Sciences. 2023; 24(18):14169. https://doi.org/10.3390/ijms241814169
Chicago/Turabian StyleJammes, Manon, Frédéric Cassé, Emilie Velot, Arnaud Bianchi, Fabrice Audigié, Romain Contentin, and Philippe Galéra. 2023. "Pro-Inflammatory Cytokine Priming and Purification Method Modulate the Impact of Exosomes Derived from Equine Bone Marrow Mesenchymal Stromal Cells on Equine Articular Chondrocytes" International Journal of Molecular Sciences 24, no. 18: 14169. https://doi.org/10.3390/ijms241814169
APA StyleJammes, M., Cassé, F., Velot, E., Bianchi, A., Audigié, F., Contentin, R., & Galéra, P. (2023). Pro-Inflammatory Cytokine Priming and Purification Method Modulate the Impact of Exosomes Derived from Equine Bone Marrow Mesenchymal Stromal Cells on Equine Articular Chondrocytes. International Journal of Molecular Sciences, 24(18), 14169. https://doi.org/10.3390/ijms241814169