
Redox Regulation of Salt Tolerance in Eutrema salsugineum by Proteomics

 , and
, and 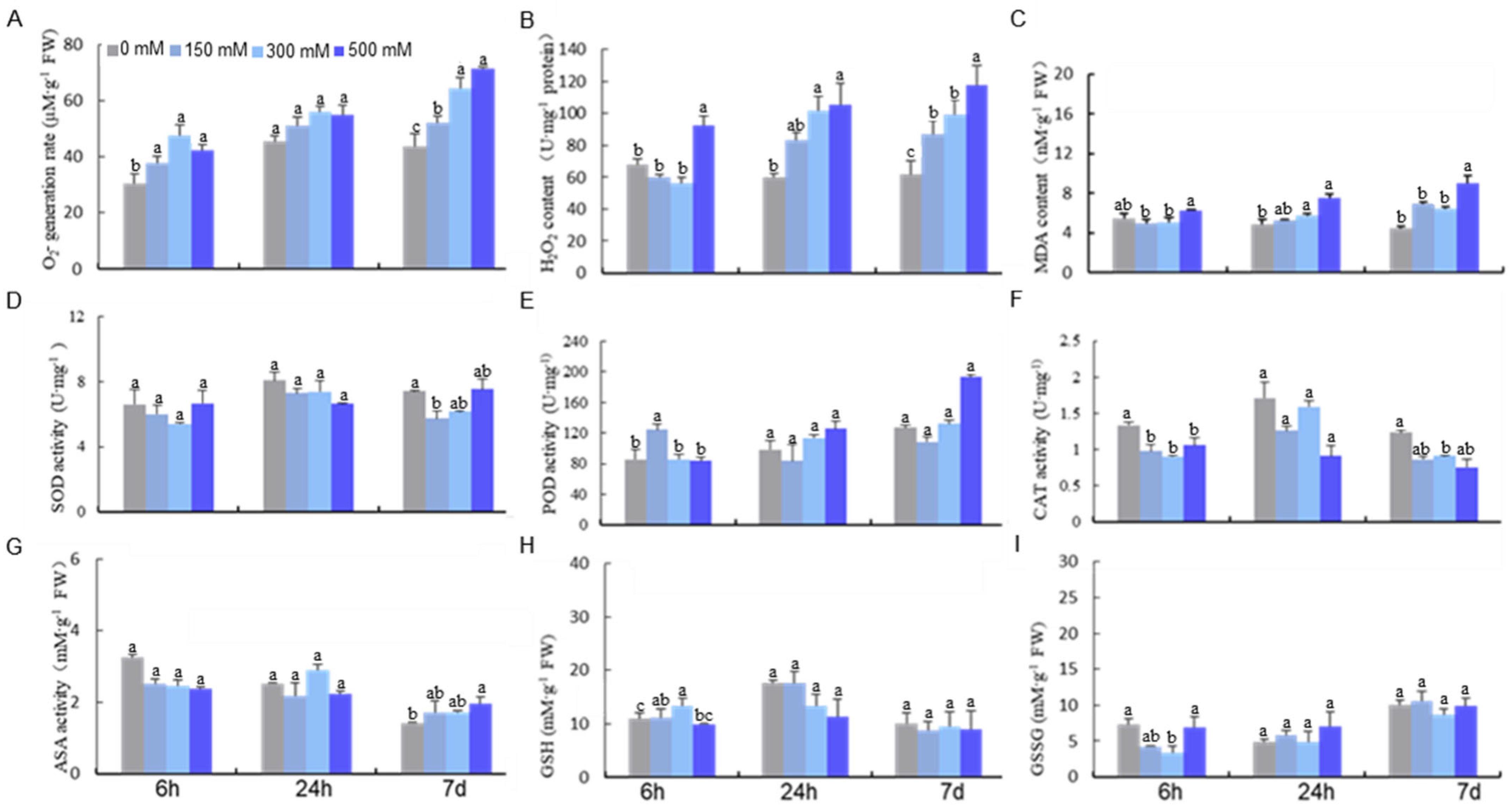{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Impact of Salt on the ROS Scavenging System
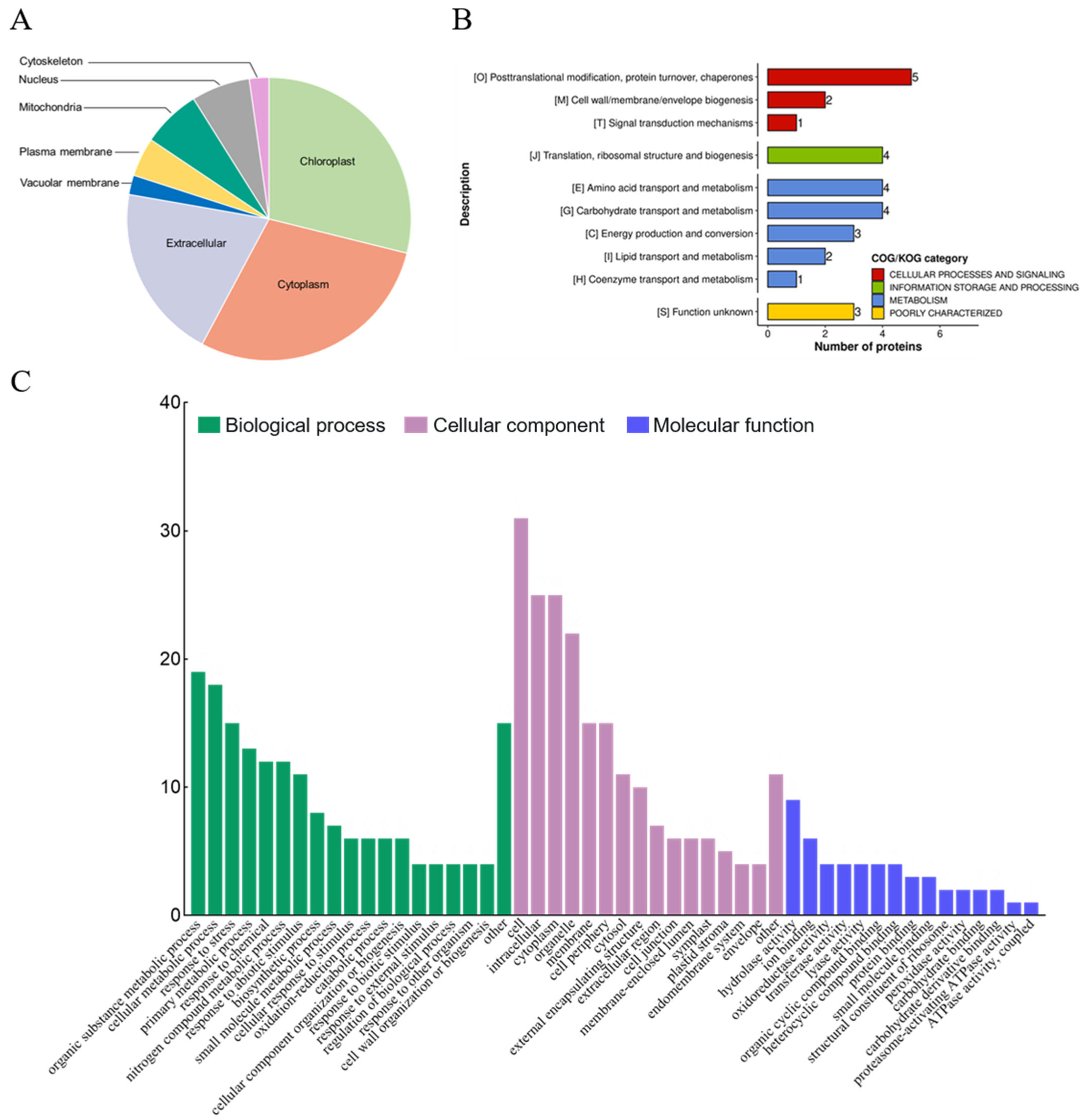
2.2. Redox Proteomics Reveals Proteins Sensitive to Salt Stress in E. salsugineum Leaves
2.3. Ectopic Expression of EsGPX in Arabidopsis Enhances Salt Tolerance
2.4. Overexpressing EsGPX Alleviates Salt-Induced Oxidative Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Vector Constructions and Plant Transformation
4.3. Phenotypical Analysis of Seed Germination, Root Length, and Flowering
4.4. Sample Preparation of Proteomics
4.5. LC-MS/MS
4.6. Database Searching and Data Analysis
4.7. Chlorophylls
4.8. NBT and DAB Staining of Leaves
4.9. Determination of the H2O2 and Malondialdehyde Contents
4.10. Antioxidant Enzyme Activities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, K.; Vitamvas, P.; Prasil, I.T.; Renaut, J. Plant proteome changes under abiotic stress contribution of proteomics studies to understanding plant stress response. J. Proteome 2011, 74, 1301–1322. [Google Scholar]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Xu, Q.T.; Yue, Y.P.; Duan, Y.H.; Liu, H.; Chen, X.L.; Huang, J.B.; Zheng, L. Comparative oxidation proteomics analyses suggest redox regulation of cytosolic translation in rice leaves upon Magnaporthe oryzae infection. Plant Commun. 2023, 4, 100550. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Van Breusegem, F.; Mhamdi, A. Redox dependent control of nuclear transcription in plants. J. Exp. Bot. 2018, 69, 3359–3372. [Google Scholar] [CrossRef]
- Tada, Y.; Spoel, S.H.; Pajerowska Mukhtar, K.; Mou, Z.; Song, J.; Wang, C.; Zuo, J.; Dong, X. Plant immunity requires conformational charges of NPR1 via Snitrosylation and thioredoxins. Science 2008, 321, 952–956. [Google Scholar] [CrossRef]
- Criqui, M.C.; Jamet, E.; Parmentier, Y.; Marbach, J.; Durr, A.; Fleck, J. Isolation and characterization of a plant cDNA showing homology to animal glutathione peroxidases. Plant Mol. Biol. 1992, 18, 623–627. [Google Scholar] [CrossRef]
- Li, W.J.; Feng, H.; Fan, J.H.; Zhang, R.Q.; Zhao, N.M.; Liu, J.Y. Molecular cloning and expression of a phospholipid hydroperoxide glutathione peroxidase homolog in Oryza sativa. Biochim. Biophys. Acta 2000, 1493, 225–230. [Google Scholar] [CrossRef]
- Bela, K.; Riyazuddin, R.; Horváth, E.; Hurton, Á.; Gallé, Á.; Takács, Z.; Zsigmond, L.; Szabados, L.; Tari, I.; Csiszár, J. Comprehensive analysis of antioxidant mechanisms in Arabidopsis glutathione peroxidase-like mutants under salt- and osmotic stress reveals organ-specific significance of the AtGPXL’s activities. Environ. Exp. Bot. 2018, 150, 127–140. [Google Scholar]
- Ma, T.T. Cloning and Expression Analysis of Salt-Mustard Glutathione Peroxidase Gene; Minzu University of China: Beijing, China, 2012. [Google Scholar]
- Koch, M.A.; German, D.A. Taxonomy and systematics are key to biological information: Arabidopsis, Eutrema (Thellungiella), Noccaea and Schrenkiella (Brassicaceae) as examples. Front. Plant Sci. 2013, 4, 267. [Google Scholar] [CrossRef]
- Bressan, R.A.; Zhang, C.; Zhang, H.; Hasegawa, P.M.; Bohnert, H.J.; Zhu, J.K. Learning from the Arabidopsis experience. The next gene search paradigm. Plant Physiol. 2001, 127, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Jarvis, D.E.; Chen, H.; Beilstein, M.A.; Grimwood, J.; Jenkins, J.; Shu, S.Q.; Prochnik, S.; Xin, M.M.; Ma, C.; et al. The reference genome of the halophytic plant Eutrema salsugineum. Front. Plant Sci. 2013, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.; Timonin, M.; Wong, A.C. Thellungiella: An Arabidopsis related model plant adapted to cold temperatures. Plant Cell Environ. 2007, 30, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.J.; Zhang, Z.; Wang, J.Y.; Oh, D.H.; Dassanayake, M.; Liu, B.; Huang, Q.F.; Sun, H.X.; Xia, R.; Wu, Y.R.; et al. Insights into salt tolerance from the genome of Thellungiella salsuginea. Proc. Natl. Acad. Sci. USA 2012, 109, 12219–12224. [Google Scholar] [CrossRef]
- Yin, Z.P.; Kelly, B.; Geng, S.; Zhu, N.; Zhang, T.; Craig, D.; Dai, S.J.; Chen, S.X. Bicarbonate Induced Redox Proteome Changes in Arabidopsis Suspension Cells. Front. Plant Sci. 2017, 8, 58. [Google Scholar] [CrossRef]
- Jue, D.; Sang, X.; Liu, L.; Shu, B.; Wang, Y.; Liu, C.; Wang, Y.; Xie, J.; Shi, S. Comprehensive analysis of the longan transcriptome reveals distinct regulatory programs during the foral transition. BMC Genom. 2019, 20, 126. [Google Scholar] [CrossRef]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Xanthopoulou, A.; Moysiadis, T.; Bazakos, C.; Karagiannis, E.; Karamichali, I.; Stamatakis, G.; Samiotaki, M.; Manioudaki, M.; Michailidis, M.; Madesis, P.; et al. The perennial fruit tree proteogenomics atlas: A spatial map of the sweet cherry proteome and transcriptome. Plant J. 2022, 109, 1319–1336. [Google Scholar] [CrossRef]
- Ye, Z.; Yu, J.; Yan, W.; Zhang, J.; Yang, D.; Yao, G.; Liu, Z.J.; Wu, Y.G.; Hou, X.L. Integrative iTRAQ-based proteomic and transcriptomic analysis reveals the accumulation patterns of key metabolites associated with oil quality during seed ripening of Camellia oleifera. Hortic. Res. 2021, 8, 157. [Google Scholar] [CrossRef]
- Balsera, M.; Uberegui, E.; Schürmann, P.; Buchanan, B.B. Evolutionary development of redox regulation in chloroplasts. Antioxid. Redox Signal. 2014, 21, 1327–1355. [Google Scholar] [CrossRef]
- Mills, C. Hemoglobin catabolism I. Glutathione peroxidase, an erythrocyte enzyme which protects hemoglobin from oxidative breakdown. J. Biol. Chem. 1957, 229, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Stiti, N.; Missihoun, T.D.; Kotchoni, S.O.; Kirch, H.H.; Bartels, D. Aldehyde dehydrogenases in Arabidopsis thaliana: Biochemical requirements, metabolic pathways, and functional analysis. Front. Plant Sci. 2011, 2, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Bouche, N.; Fait, A.; Bouchez, D.; Møller, S.G.; Fromm, H. Mitochondrial succinic-semialdehyde dehydrogenase of the gamma-aminobutyrate shunt is required to restrict levels of reactive oxygen intermediates in plants. Proc. Natl. Acad. Sci. USA 2003, 100, 6843–6848. [Google Scholar] [CrossRef]
- Hermans, C.; Porco, S.; Vandenbussche, F.; Gille, S.; Pessemier, J.D.; Straeten, D.V.D.; Verbruggen, N.; Bush, D.R.B. Dissecting the role of CHITINASE-LIKE1 in nitrate dependent changes in root architecture. Plant Physiol. 2017, 2, 538–548. [Google Scholar] [CrossRef]
- Hull, P.A.; Pauly, M.; Souza, A.D.; Pauly, M. Identification and functional characterization of the distinct plant pectin esterases PAE8 and PAE9 and their deletion mutants. Planta Int. J. Plant Biol. 2014, 240, 1123–1138. [Google Scholar]
- Zhao, R.; Houry, W.A. Hsp90: A chaperone for protein folding and gene regulation. Biochem. Cell Biol. Biochim. Biol. Cell. 2005, 83, 703–710. [Google Scholar] [CrossRef]
- Tavakoli, S.; Adili, A.; Akbari, M.; Tamjidifar, R.; Shotorbani, S.S. Inhibition effect of Hsp90 on TLR2, TLR4, and MAPK signaling pathway in melanoma in vitro. Gene Rep. 2021, 24, 101234. [Google Scholar] [CrossRef]
- Hirasawa, M.; Tripathy, J.N.; Sommer, F.; Somasundaram, R.; Chung, J.S.; Nestander, M.; Kruthiventi, M.; Zabet-Moghaddam, M.; Johnson, M.K.; Merchant, S.S.; et al. Enzymatic properties of the ferredoxin-dependent nitrite reductase from Chlamydomonas reinhardtii. Evidence for hydroxylamine as a late intermediate in ammonia production. Photosynth. Res. 2010, 103, 67–77. [Google Scholar] [CrossRef]
- Gao, F.; Chen, J.; Ma, T.; Li, H.; Wang, N.; Li, Z.; Zhang, Z.; Zhou, Y. The glutathione peroxidase gene family in Thellungiella salsuginea: Genome-wide identification, classification, and gene and protein expression analysis under stress conditions. Int. J. Mol. Sci. 2014, 15, 3319–3335. [Google Scholar] [CrossRef]
- Jiang, K.; Moe-Lange, J.; Hennet, L.; Feldman, L.J. Salt stress affects the redox status of Arabidopsis root meristems. Front. Plant Sci. 2016, 7, 81. [Google Scholar] [CrossRef]
- Locato, V.; Cimini, S.; De Gara, L. Glutathione as a key player in plant abiotic stress responses and tolerance. In Glutathione in Plant Growth, Development, and Stress Tolerance; Springer: Cham, Switzerland, 2017; pp. 127–145. [Google Scholar]
- Foyer, C.H.; Noctor, G. Redox regulation in photosynthetic organisms: Signaling, acclimation, and practical implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Dietz, K.J.; Hell, R. Thiol switches in redox regulation of chloroplasts: Balancing redox state, metabolism and oxidative stress. Biol. Chem. 2015, 396, 483–494. [Google Scholar] [CrossRef]
- Maiorino, M.; Ursini, F.; Bosello, V.; Toppo, S.; Tosatto, S.C.; Mauri, P.; Becker, K.; Roveri, A.; Bulato, C.; Benazzi, L.; et al. The thioredoxin specificity of Drosophila GPX: A paradigm for a peroxiredoxin-like mechanism of many glutathione peroxidases. J. Mol. Biol. 2007, 365, 1033–1046. [Google Scholar] [CrossRef]
- Rigó, G.; Valkai, I.; Faragó, D.; Kiss, E.; Van Houdt, S.; Vande Steene, N.; Hannah, M.A.; Szabados, L. Gene mining in halophytes: Functional identification of stress tolerance genes in Lepidium crassifolium. Plant Cell Environ. 2016, 39, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Woo, N.S.; Badger, M.R.; Pogson, B.J. A rapid, non-invasive procedure for quantitative assessment of drought survival using chlorophyll fluorescence. Plant Methods 2008, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Song, Y. Identification and Analysis the Function of Jerusalem artichoke AQP Genes in Redponse to Abiotic Stress; Northeast Forestry University: Harbin, China, 2022. [Google Scholar]
- Hodges, D.M.; Delong, J.M.; Forney, C.F. Improving the thiobarbituric acid reactive substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Benyó, D.; Horváth, E.; Németh, E.; Leviczky, T.; Takács, K.; Lehotai, N.; Feigl, G.; Kolbert, Z.; Ördög, A.; Gallé, R.; et al. Physiological and molecular responses to heavy metal stresses suggest different detoxification mechanism of Populus deltoides and P. X canadensis. J. Plant Physiol. 2016, 201, 62–70. [Google Scholar] [CrossRef]
- Qian, W.G. Genome Wide Characterization of AQP Gene Family and Functional Analysis on Stress Redidtance of EsPIP1;2 and EsPIP2;1 in Eutrema salsugineum; Northeast Forestry University: Harbin, China, 2020. [Google Scholar]
- Tari, I.; Csiszár, J.; Horváth, E.; Poór, P.; Takács, Z.; Szepesi, Á. The alleviation of the adverse effects of salt stress in the tomato plant by salicylic acid shows a time-and organ specific antioxidant response. Acta Biol. Cracov. Bot. 2015, 57, 21–30. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Yang, X.; Liu, F.; Liu, X.; Zhao, T.; Yan, X.; Pang, Q. Redox Regulation of Salt Tolerance in Eutrema salsugineum by Proteomics. Int. J. Mol. Sci. 2023, 24, 14518. https://doi.org/10.3390/ijms241914518
Li J, Yang X, Liu F, Liu X, Zhao T, Yan X, Pang Q. Redox Regulation of Salt Tolerance in Eutrema salsugineum by Proteomics. International Journal of Molecular Sciences. 2023; 24(19):14518. https://doi.org/10.3390/ijms241914518
Chicago/Turabian StyleLi, Jiawen, Xiaomin Yang, Fuqing Liu, Xinxin Liu, Tong Zhao, Xiufeng Yan, and Qiuying Pang. 2023. "Redox Regulation of Salt Tolerance in Eutrema salsugineum by Proteomics" International Journal of Molecular Sciences 24, no. 19: 14518. https://doi.org/10.3390/ijms241914518
APA StyleLi, J., Yang, X., Liu, F., Liu, X., Zhao, T., Yan, X., & Pang, Q. (2023). Redox Regulation of Salt Tolerance in Eutrema salsugineum by Proteomics. International Journal of Molecular Sciences, 24(19), 14518. https://doi.org/10.3390/ijms241914518