

Extensive Expression of the Virulome Related to Antibiotic Genotyping in Nosocomial Strains of Klebsiella pneumoniae

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Multidrug-Resistance Phenotype
2.2. Origin of the Strains and In Vitro Infection of Human Epithelial Cell Lines to Determine Viruloma Expression
2.3. Detection of Antibiotic Resistance Genes by Conventional PCR
2.4. Virulence Genes Expression According to Antibiotic Resistance Genes
2.5. PFGE and Expression Patterns of Virulence Genes
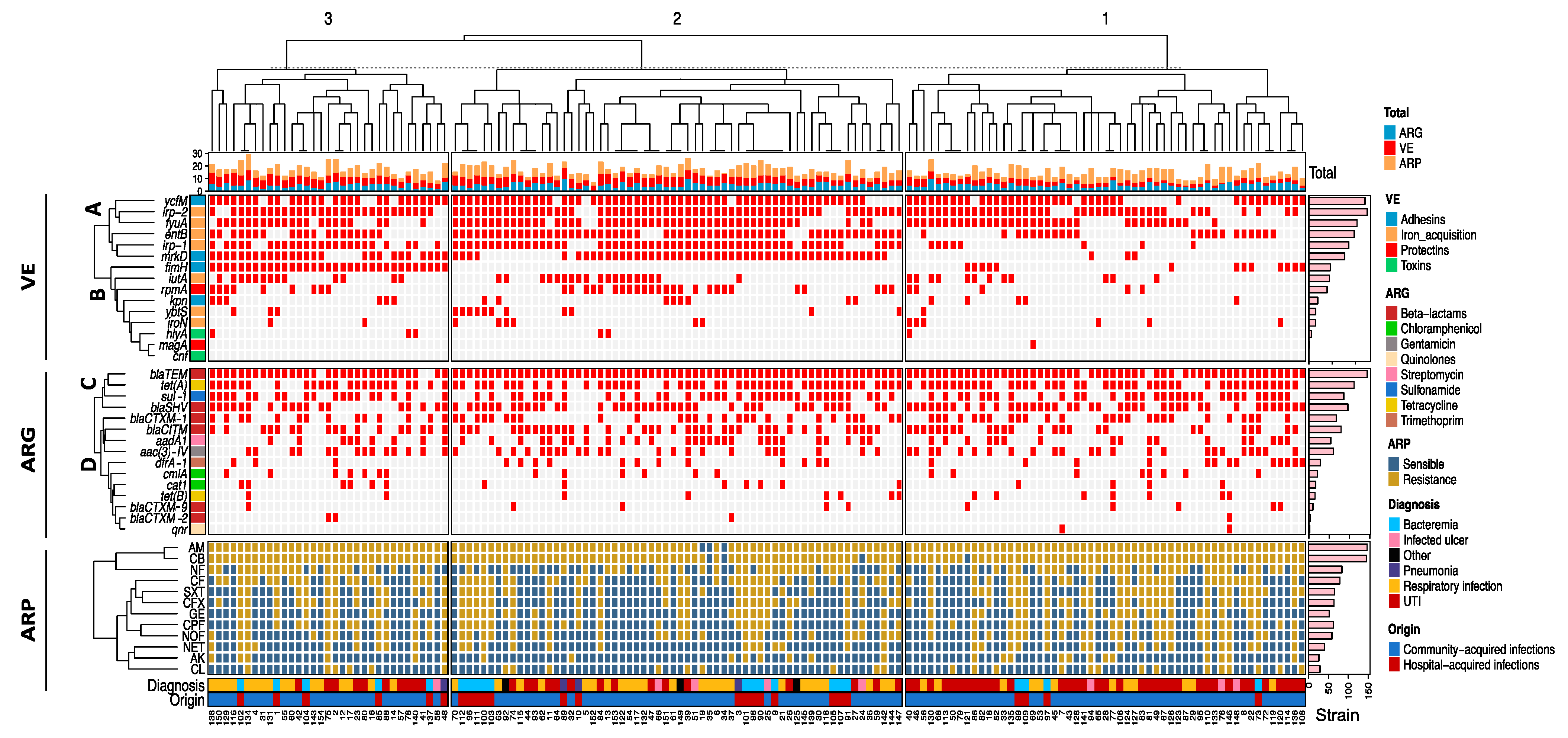
2.6. Unsupervised Hierarchical Clustering
3. Discussion
4. Materials and Methods
4.1. Origin of the Strains
4.2. Bacterial DNA Extraction
4.3. Identification of K. pneumoniae through Polymerase Chain Reaction (PCR)
4.4. Multidrug-Resistance Phenotype
4.5. Detection of Antibiotic-Resistance Genes Using Conventional PCR
4.6. Bacterial Dilution of Strains for Infection of Human Epithelial Cell Lines
4.7. Preparation of Human Epithelial Cell Lines and In Vitro Infection Model
4.8. K. pneumoniae RNA Extraction and Reverse-Transcription to cDNA
4.9. Determination of K. pneumoniae Virulome Expression Using Real-Time PCR
4.10. Pulse Field Gel Electrophoresis (PFGE)
4.11. Unsupervised Hierarchical Clustering
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patil, S.; Dong, S.; Francisco, N.M.; Liu, S.; Wen, F. Emergence of multidrug-resistant Klebsiella pneumoniae in hospitalised young children. Lancet Microbe 2023, 4, 390. [Google Scholar] [CrossRef]
- Davoudabadi, S.; Goudarzi, M.; Hashemi, A. Detection of Virulence Factors and Antibiotic Resistance among Klebsiella pneumoniae Isolates from Iran. Biomed. Res. Int. 2023, 3624497. [Google Scholar] [CrossRef] [PubMed]
- Kochan, T.J.; Nozick, S.H.; Medernach, R.L.; Cheung, B.H.; Gatesy, S.W.M.; Lebrun-Corbin, M.; Mitra, S.D.; Khalatyan, N.; Krapp, F.; Qi, C.; et al. Genomic surveillance for multidrug-resistant or hypervirulent Klebsiella pneumoniae among United States bloodstream isolates. BMC Infect. Dis. 2022, 22, 603. [Google Scholar] [CrossRef]
- Gu, D.; Dong, N.; Zheng, Z.; Lin, D.; Huang, M.; Wang, L.; Chan, E.W.; Shu, L.; Yu, J.; Zhang, R.; et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: A molecular epidemiological study. Lancet Infect. Dis. 2018, 18, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Toledano-Tableros, J.E.; Gayosso-Vázquez, C.; Jarillo-Quijada, M.D.; Fernández-Vázquez, J.L.; Morfin-Otero, R.; Rodríguez-Noriega, E.; Giono-Cerezo, S.; Gutkind, G.; Di Conza, J.; Santos-Preciado, J.I.; et al. Dissemination of bla NDM- 1 Gene Among Several Klebsiella pneumoniae Sequence Types in Mexico Associated With Horizontal Transfer Mediated by IncF-Like Plasmids. Front. Microbiol. 2021, 12, 611274. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Ortíz, I.A.; Juárez-Gómez, J.C.; Cu-Quijano, C.; Flores-Paz, R.; Durán-Manuel, E.M.; Cruz-Cruz, C.; Gutiérrez-Muñoz, V.H.; Sosa-Hernández, O.; Escobar-Escamilla, N.; Bravata-Alcántara, J.C.; et al. Klebsiella pneumoniae blaNDM-1 carrying a class 1 integron causing a hospital outbreak in a Mexican attention center. J. Infect. Dev. Ctries 2021, 15, 657–664. [Google Scholar] [CrossRef]
- Dehshiri, M.; Khoramrooz, S.S.; Zoladl, M.; Khosravani, S.A.; Parhizgari, N.; Motazedian, M.H.; Jahedi, S.; Sharifi, A. The frequency of Klebsiella pneumonia encoding genes for CTX-M, TEM-1 and SHV-1 extended-spectrum beta lactamases enzymes isolated from urinary tract infection. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 4. [Google Scholar] [CrossRef]
- Troyano-Hernáez, P.; Gutiérrez-Arroyo, A.; Gómez-Gil, R.; Mingorance, J.; Lázaro-Perona, F. Emergence of Klebsiella pneumoniae Harboring the aac(6ʹ)-Ian Amikacin Resistance Gene. Antimicrob. Agents Chemother. 2018, 62, e01952-18. [Google Scholar] [CrossRef]
- Ferreira, R.L.; da Silva, B.C.M.; Rezende, G.S.; Nakamura-Silva, R.; Pitondo-Silva, A.; Campanini, E.B.; Brito, M.C.A.; da Silva, E.M.L.; Freire, C.C.M.; da Cunha, A.F.; et al. High Prevalence of Multidrug-Resistant Klebsiella pneumoniae Harboring Several Virulence and β-Lactamase Encoding Genes in a Brazilian Intensive Care Unit. Front. Microbiol. 2019, 9, 3198. [Google Scholar] [CrossRef]
- Yang, X.; Dong, N.; Chan, E.W.; Zhang, R.; Chen, S. Carbapenem Resistance-Encoding and Virulence-Encoding Conjugative Plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021, 29, 65–83. [Google Scholar] [CrossRef]
- Hua, Y.; Wang, J.; Huang, M.; Huang, Y.; Zhang, R.; Bu, F.; Yang, B.; Chen, J.; Lin, X.; Hu, X.; et al. Outer membrane vesicles-transmitted virulence genes mediate the emergence of new antimicrobial-resistant hypervirulent Klebsiella pneumoniae. Emerg. Microbes Infect. 2022, 11, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sun, Q.; Li, J.; Jiang, Y.; Li, Y.; Lin, J.; Chen, K.; Chan, E.W.; Zhang, R.; Chen, S. Molecular epidemiology of carbapenem-resistant hypervirulent Klebsiella pneumoniae in China. Emerg. Microbes Infect. 2022, 11, 841–849. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, T.; Chen, L.; Du, H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 642484. [Google Scholar] [CrossRef] [PubMed]
- Namikawa, H.; Oinuma, K.I.; Yamada, K.; Kaneko, Y.; Kakeya, H.; Shuto, T. Predictors of hypervirulent Klebsiella pneumoniae infections: A systematic review and meta-analysis. J. Hosp. Infect. 2023, 134, 153–160. [Google Scholar] [CrossRef] [PubMed]
- El Fertas, A.R.; Messai, Y.; Alouache, S.; Bakour, R. Virulence profiles and antibiotic susceptibility patterns of Klebsiella pneumoniae strains isolated from different clinical specimens. Pathol. Biol. Paris 2013, 61, 209–216. [Google Scholar] [CrossRef]
- Struve, C.; Bojer, M.; Krogfelt, K.A. Characterization of Klebsiella pneumoniae type 1 fimbriae by detection of phase variation during colonization and infection and impact on virulence. Infect. Immun. 2008, 76, 4055–4065. [Google Scholar] [CrossRef]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense whit a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef]
- Subashchandrabose, S.; Mobley, H.L.T. Virulence and Fitness Determinants of Uropathogenic Escherichia coli. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Han, Y.L.; Wen, X.H.; Zhao, W.; Cao, X.S.; Wen, J.X.; Wang, J.R.; Hu, Z.D.; Zheng, W.Q. Epidemiological characteristics and molecular evolution mechanisms of carbapenem-resistant hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2022, 13, 1003783. [Google Scholar] [CrossRef]
- Ostria-Hernandez, M.L.; Juárez-de la Rosa, K.C.; Arzate-Barbosa, P.; Lara-Hernández, A.; Sakai, F.; Ibarra, J.A.; Castro-Escarpulli, G.; Vidal, J.E. Nosocomial, Multidrug-Resistant Klebsiella pneumoniae Strains Isolated from Mexico City Produce Robust Biofilms on Abiotic Surfaces but Not on Human Lung Cells. Microb. Drug Resist. 2018, 24, 422–433. [Google Scholar] [CrossRef]
- Catalán-Nájera, J.C.; Barrios-Camacho, H.; Duran-Bedolla, J.; Sagal-Prado, A.; Hernández-Castro, R.; García-Méndez, J.; Morfín-OteroMorfín-Otero, R.; Velázquez-Larios, M.D.R.; Ortíz-Navarrete, V.; Gutierrez-Xicotencatl, L.; et al. Molecular characterization and pathogenicity determination of hypervirulent Klebsiella pneumoniae clinical isolates serotype K2 in Mexico. Diagn. Microbiol. Infect. Dis. 2019, 94, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Remya, P.A.; Shanthi, M.; Sekar, U. Characterisation of virulence genes associated with pathogenicity in Klebsiella pneumoniae. Indian J. Med. Microbiol. 2019, 37, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.M.; Moreno Mochi, M.P.; Nuñez, J.M.; Cáceres, M.; Mochi, S.; Del Campo Moreno, R.; Jure, M.A. Virulence factors and clinical patterns of multiple-clone hypermucoviscous KPC-2 producing K. pneumoniae. Heliyon 2019, 5, e01829. [Google Scholar] [CrossRef] [PubMed]
- Gołębiewska, J.E.; Krawczyk, B.; Wysocka, M.; Ewiak, A.; Komarnicka, J.; Bronk, M.; Rutkowski, B.; Dębska-Ślizień, A. Host and pathogen factors in Klebsiella pneumoniae upper urinary tract infections in renal transplant patients. J. Med. Microbiol. 2019, 68, 382–394. [Google Scholar] [CrossRef] [PubMed]
- Hallal Ferreira Raro, O.; Nordmann, P.; Dominguez Pino, M.; Findlay, J.; Poirel, L. Emergence of Carbapenemase-Producing Hypervirulent Klebsiella pneumoniae in Switzerland. Antimicrob. Agents Chemother. 2023, 67, e0142422. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef]
- Bautista-Cerón, A.; Monroy-Pérez, E.; García-Cortés, L.R.; Rojas-Jiménez, E.A.; Vaca-Paniagua, F.; Paniagua-Contreras, G.L. Hypervirulence and Multiresistance to Antibiotics in Klebsiella pneumoniae Strains Isolated from Patients with Hospital- and Community-Acquired Infections in a Mexican Medical Center. Microorganisms 2022, 10, 2043. [Google Scholar] [CrossRef]
- Bakhtiari, R.; Javadi, A.; Aminzadeh, M.; Molaee-Aghaee, E.; Shaffaghat, Z. Association between Presence of RmpA, MrkA and MrkD Genes and Antibiotic Resistance in Clinical Klebsiella pneumoniae Isolates from Hospitals in Tehran, Iran. Iran. J. Public Health 2021, 50, 1009–1016. [Google Scholar] [CrossRef]
- Braun, V. Iron uptake mechanisms and their regulation in pathogenic bacteria. Int. J. Med. Microbiol. 2001, 291, 67–79. [Google Scholar] [CrossRef]
- Chen, T.; Dong, G.; Zhang, S.; Zhang, X.; Zhao, Y.; Cao, J.; Zhou, T.; Wu, Q. Effects of iron on the growth, biofilm formation and virulence of Klebsiella pneumoniae causing liver abscess. BMC Microbiol. 2020, 20, 36. [Google Scholar] [CrossRef]
- Bachman, M.A.; Oyler, J.E.; Burns, S.H.; Caza, M.; Lepine, F.; Dozois, C.M.; Weiser, J.N. Klebsiella pneumoniae yersiniabactin promotes respiratory tract infection through evasion of lipocalin 2. Infect. Immun. 2011, 79, 3309–3316. [Google Scholar] [CrossRef] [PubMed]
- Schubert, S.; Picard, B.; Gouriou, S.; Heesemann, J.; Denamur, E. Yersinia high-pathogenicity island contributes to virulence in Escherichia coli causing extraintestinal infections. Infect. Immun. 2002, 70, 5335–5337. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, S.; Kosai, K.; Ota, K.; Mitsumoto-Kaseida, F.; Sakamoto, K.; Hasegawa, H.; Izumikawa, K.; Mukae, H.; Yanagihara, K. Clinical and microbiological characteristics of bloodstream infection caused by Klebsiella pneumoniae harboring rmpA in Japanese adults. Sci. Rep. 2023, 13, 6571. [Google Scholar] [CrossRef]
- Schulz, E.; Schumann, M.; Schneemann, M.; Dony, V.; Fromm, A.; Nagel, O.; Schulzke, J.D.; Bücker, R. Escherichia coli Alpha-Hemolysin HlyA Induces Host Cell Polarity Changes, Epithelial Barrier Dysfunction and Cell Detachment in Human Colon Carcinoma Caco-2 Cell Model via PTEN-Dependent Dysregulation of Cell Junctions. Toxins 2021, 13, 520. [Google Scholar] [CrossRef] [PubMed]
- Gallegos-Miranda, V.; Garza-Ramos, U.; Bolado-Martínez, E.; Navarro-Navarro, M.; Félix-Murray, K.R.; Candia-Plata, M.D.C.; Sanchez-Martinez, G.; Dúran-Bedolla, J.; Silva-Sánchez, J. ESBL-producing Escherichia coli and Klebsiella pneumoniae from health-care institutions in Mexico. J. Chemother. 2021, 33, 122–127. [Google Scholar] [CrossRef]
- Al-Sheboul, S.A.; Al-Madi, G.S.; Brown, B.; Hayajneh, W.A. Prevalence of Extended-Spectrum β-Lactamases in Multidrug-Resistant Klebsiella pneumoniae Isolates in Jordanian Hospitals. J. Epidemiol. Glob. Health 2023, 13, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Dan, B.; Dai, H.; Zhou, D.; Tong, H.; Zhu, M. Relationship Between Drug Resistance Characteristics and Biofilm Formation in Klebsiella pneumoniae Strains. Infect. Drug Resist. 2023, 16, 985–998. [Google Scholar] [CrossRef]
- Córdova-Espinoza, M.G.; Giono-Cerezo, S.; Sierra-Atanacio, E.G.; Escamilla-Gutiérrez, A.; Carrillo-Tapia, E.; Carrillo-Vázquez, L.I.; Mendoza-Pérez, F.; Leyte-Lugo, M.; González-Vázquez, R.; Mayorga-Reyes, L.; et al. Isolation and Identification of Multidrug-Resistant Klebsiella pneumoniae Clones from the Hospital Environment. Pathogens 2023, 12, 634. [Google Scholar] [CrossRef]
- Hwang, J.H.; Handigund, M.; Hwang, J.H.; Cho, Y.G.; Kim, D.S.; Lee, J. Clinical Features and Risk Factors Associated With 30-Day Mortality in Patients With Pneumonia Caused by Hypervirulent Klebsiella pneumoniae (hvKP). Ann. Lab. Med. 2020, 40, 481–487. [Google Scholar] [CrossRef]
- Xu, L.; Sun, X.; Ma, X. Systematic review and meta-analysis of mortality of patients infected with carbapenem-resistant Klebsiella pneumoniae. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 18. [Google Scholar] [CrossRef]
- Rippere-Lampe, K.E.; O’Brien, A.D.; Conran, R.; Lockman, H.A. Mutation of the gene encoding cytotoxic necrotizing factor type 1 (cnf-1) attenuates the virulence of uropathogenic Escherichia coli. Infect. Immun. 2001, 69, 3954–3964. [Google Scholar] [CrossRef]
- Yeh, K.M.; Lin, J.C.; Yin, F.Y.; Fung, C.P.; Hung, H.C.; Siu, L.K.; Chang, F.Y. Revisiting the importance of virulence determinant magA and its surrounding genes in Klebsiella pneumoniae causing pyogenic liver abscesses: Exact role in serotype K1 capsule formation. J. Infect. Dis. 2010, 201, 1259–1267. [Google Scholar] [CrossRef]
- Müller, C.M.; Aberg, A.; Straseviçiene, J.; Emody, L.; Uhlin, B.E.; Balsalobre, C. Type 1 fimbriae, a colonization factor of uropathogenic Escherichia coli, are controlled by the metabolic sensor CRP- cAMP. PLoS Pathog. 2009, 5, e1000303. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Canovas, L.; Martinez Benitez, M.B.; Herrera Isidron, J.A.; Flores Soto, E. Pulsed Field Gel Electrophoresis: Past, present, and future. Anal. Biochem. 2019, 573, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Ooka, T.; Terajima, J.; Kusumoto, M.; Iguchi, A.; Kurokawa, K.; Ogura, Y.; Asadulghani, M.; Nakayama, K.; Murase, K.; Ohnishi, M.; et al. Development of a multiplex PCR-based rapid typing method for enterohemorrhagic Escherichia coli O157 strains. J. Clin. Microbiol. 2009, 47, 2888–2894. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, C.; Zheng, W.; Zhang, X.; Yu, J.; Gao, Q.; Hou, Y.; Huang, X. PCR detection of Klebsiella pneumoniae in infant formula based on 16S-23S internal transcribed spacer. Int. J. Food Microbiol. 2008, 125, 230–235. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance standards for antimicrobial susceptibility testing. In CLSI, Twenty-Third Informational Supplement M100-S23; Carpenter, D.E., Ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013; p. 74. [Google Scholar]
- Dallenne, C.; Da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Momtaz, H.; Karimian, A.; Madani, M.; Safarpoor Dehkordi, F.; Ranjbar, R.; Sarshar, M.; Souod, N. Uropathogenic Escherichia coli in Iran: Serogroup distributions, virulence factors and antimicrobial resistance properties. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 8. [Google Scholar] [CrossRef]
- Cheng, G.; Wozniak, K.; Wallig, M.A.; Fidel, P.L., Jr.; Trupin, S.R.; Hoyer, L.L. Comparison between Candida albicans Agglutinin-Like Sequence Gene Expression Patterns in Human Clinical Specimens and Models of Vaginal Candidiasis. Infect. Immun. 2005, 73, 1656–1663. [Google Scholar] [CrossRef]
- Naglik, J.R.; Moyes, D.; Makwana, J.; Kanzaria, P.; Tsichlaki, E.; Weindl, G.; Tappuni, A.R.; Rodgers, C.A.; Woodman, A.J.; Challacombe, S.J.; et al. Quantitative expression of the Candida albicans secreted aspartyl proteinase gene family in human oral and vaginal candidiasis. Microbiology 2008, 154, 3266–3280. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed- field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antibiotics | Antibiotic Group | Number of Resistant Strains (%) |
|---|---|---|
| Carbenicillin (CB) | beta-lactams | 148 (98.6) |
| Ampicillin (AM) | 148 (98.6) | |
| Cefotaxime (CFX) | 66 (44) | |
| Cephalothin (CF) | 81 (54) | |
| Gentamicin (GE) | Aminoglycosides | 63 (42) |
| Amikacin (AK) | 28 (18.6) | |
| Netilmycin (NET) | 42 (28) | |
| Ciprofloxacin (CPF) | Quinolones | 64 (42.6) |
| Norfloxacin (NOF) | 61 (40.6) | |
| Nitrofurantoin (NF) | Nitrofurans | 86 (57.3) |
| Chloramphenicol (CL) | Phenicols | 31 (20.6) |
| Sulfamethoxazole/trimethoprim (SXT) | Sulfonamide/Trimethoprim | 67 (44.6) |
| Origin of the Strains | Number of Strains (%) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adhesins | Iron Acquisition Systems | Protectins | Toxins | |||||||||||||
| fimH | mrkD | kpn | ycfM | entB | irp1 | irp2 | ybtS | fyuA | iutA | iroN | magA | rmpA | hlyA | cnf-1 | ||
| Hospital-acquired (n = 25) | Bacteremia (n = 21) | 5 (23.8) | 15 (71.4) | 5 (23.8) | 21 (100) | 19 (90.5) | 14 (66.7) | 19 (90.5) | 6 (28.6) | 17 (81) | 2 (9.5) | 0 | 0 | 0 | 0 | 0 |
| Pneumonia (n = 4) | 1 (25) | 4 (100) | 0 | 4 (100) | 3 (75) | 2 (50) | 2 (50) | 1 (25) | 3 (75) | 3 (75) | 0 | 0 | 1 (25) | 0 | 0 | |
| Community-acquired (n = 125) | UTI (n = 61) | 22 (36.1) | 13 (21.3) | 6 (9.8) | 45 (73.8) | 27 (44.3) | 27 (44.3) | 52 (85.2) | 2 (3.3) | 38 (62.3) | 17 (27.9) | 7 (11.5) | 0 | 11 (18) | 4 (6.6) | 0 |
| Respiratory infection (n = 53) | 16 (30.2) | 37 (69.8) | 6 (11.3) | 40 (75.5) | 41 (77.4) | 36 (67.9) | 45 (84.9) | 5 (9.4) | 38 (71.7) | 20 (37.7) | 5 (9.4) | 1 (1.9) | 23 (43.4) | 2 (3.8) | 0 | |
| Infected ulcer (n = 8) | 2 (25) | 6 (75) | 1 (12.5) | 7 (87.5) | 5 (62.5) | 4 (50) | 5 (62.5) | 0 | 4 (50) | 1 (12.5) | 1 (12.5) | 0 | 2 (25) | 0 | 0 | |
| * Other (n = 3) | 0 | 2 (66.7) | 1 (33.3) | 3 (100) | 2 (66.7) | 2 (66.7) | 3 (100) | 1 (33.3) | 3 (100) | 1 (33.3) | 1 (33.3) | 0 | 2 (66.7) | 0 | 0 | |
| p-value | 0.732 | 1.8 × 10−8 | 0.264 | 0.054 | 0.0001 | 0.103 | 0.422 | 0.009 | 0.265 | 0.058 | 0.210 | 0.593 | 0.0001 | 0.763 | - | |
| Total (n = 150) | 46 (30.7) | 77 (51.3) | 19 (12.7) | 120 (80) | 97 (64.7) | 85 (56.7) | 126 (84) | 15 (10) | 103 (68.7) | 44 (29.3) | 14 (9.3) | 1 (0.7) | 39 (26) | 6 (4) | 0 | |
| Origin of the Strains | Number of Strains (%) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Beta–Lactams | Streptomycin | Gentamicin | Sulfonamide | Chloramphenicol | Tetracycline | Trimethoprim | Quinolones | |||||||||
| blaSHV | blaCITM | blaTEM | blaCTXM-1 | blaCTXM-2 | blaCTXM-9 | aadA1 | aac(3)-IV | sul1 | cat1 | cmlA | tet(A) | tet(B) | dfrA1 | qnr | ||
| Hospital-acquired (n = 25) | Bacteremia (n = 21) | 16 (76.2) | 10 (47.6) | 20 (95.2) | 17 (80.9) | 0 | 1 (4.7) | 5 (23.8) | 2 (9.5) | 12 (57.1) | 5 (23.8) | 2 (9.52) | 20 (95.2) | 1 (4.7) | 1 (4.7) | 0 |
| Pneumonia (n = 4) | 2 (50) | 3 (75) | 4 (100) | 2 (50) | 0 | 0 | 2 (50) | 3 (75) | 3 (75) | 1 (25) | 1 (25) | 3 (75) | 1 (25) | 0 | 0 | |
| Community-acquired (n = 125) | UTI (n = 61) | 39 (63.9) | 36 (59) | 56 (91.8) | 30 (49) | 3 (4.9) | 4 (6.5) | 20 (32.7) | 32 (52.4) | 32 (52.4) | 3 (4.9) | 10 (16.3) | 46 (75.4) | 8 (13.1) | 14 (22.9) | 2 (3.2) |
| Respiratory infection (n = 53) | 32 (60.3) | 26 (49) | 53 (100) | 13 (24.5) | 1 (1.8) | 5 (9.4) | 24 (45.2) | 19 (35.8) | 34 (64.1) | 7 (13.2) | 7 (13.2) | 35 (66) | 4 (7.5) | 10 (18.8) | 0 | |
| Infected ulcer (n = 8) | 5 (62.5) | 4 (50) | 8 (100) | 3 (37.5) | 0 | 0 | 3 (37.5) | 5 (62.5) | 4 (50) | 1 (12.5) | 1 (12.5) | 5 (62.5) | 1 (12.5) | 2 (25) | 0 | |
| Other (n = 3) | 2 (66.6) | 0 | 3 (100) | 2 (66.6) | 0 | 0 | 0 | 0 | 1 (33.3) | 0 | 0 | 2 (66.6) | 0 | 1 (33.3) | 0 | |
| p-value | 0.840 | 0.389 | 0.363 | 0.0001 | 0.765 | 0.941 | 0.363 | 0.001 | 0.709 | 0.125 | 0.906 | 0.094 | 0.565 | 0.334 | 0.710 | |
| Total (n = 150) | 96 (64) | 79 (52.6) | 144 (96) | 67 (44.7) | 4 (2.7) | 10 (6.7) | 54 (36) | 61 (40.7) | 86 (57.3) | 17 (11.3) | 21 (14) | 111 (74) | 15 (10) | 28 (18.7) | 2 (1.3) | |
| Hospital-Acquired Infections (n = 25) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotic Resistance Gene | Number of Strains (%) | |||||||||||||||
| Adhesins | Iron Acquisition Systems | Protectins | Toxins | |||||||||||||
| fimH | mrkD | kpn | ycfM | entB | irp1 | irp2 | ybtS | fyuA | iutA | iroN | magA | rmpA | hlyA | cnf-1 | ||
| Beta–lactams | blaSHV | 6 (24) | 18 (72) | 4 (16) | 24 (96) | 18 (72) | 15 (60) | 20 (80) | 7 (28) | 18 (72) | 5 (20) | 0 | 0 | 1 (4) | 0 | 0 |
| blaCITM | 4 (16) | 12 (48) | 2 (8) | 13 (52) | 10 (40) | 8 (32) | 12 (48) | 5 (20) | 10 (20) | 3 (12) | 0 | 0 | 1 (4) | 0 | 0 | |
| blaTEM | 5 (20) | 22 (88) | 5 (20) | 24 (96) | 22 (88) | 16 (64) | 20 (80) | 7 (28) | 20 (80) | 5 (20) | 0 | 0 | 1 (4) | 0 | 0 | |
| blaCTXM-1 | 6 (24) | 13 (52) | 5 (20) | 19 (76) | 16 (64) | 11 (44) | 17 (68) | 5 (20) | 15 (60) | 3 (12) | 0 | 0 | 0 | 0 | 0 | |
| blaCTXM-2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| blaCTXM-9 | 0 | 1 (4) | 1 (4) | 0 | 1 (4) | 0 | 1 (4) | 0 | 1 (4) | 0 | 0 | 0 | 0 | 0 | 0 | |
| Streptomycin | aadA1 | 1 (4) | 6 (24) | 1 (4) | 7 (28) | 6 (24) | 5 (20) | 6 (24) | 3 (12) | 6 (24) | 1 (4) | 0 | 0 | 1 (4) | 0 | 0 |
| Gentamicin | aac(3)-IV | 1 (4) | 4 (16) | 0 | 5 (20) | 4 (16) | 2 (8) | 3 (12) | 1 (4) | 3 (12) | 2 (8) | 0 | 0 | 1 (4) | 0 | 0 |
| Sulfonamide | sul1 | 4 (16) | 13 (52) | 2 (8) | 15 (60) | 13 (52) | 11 (44) | 12 (48) | 3 (12) | 11 (44) | 3 (12) | 0 | 0 | 1 (4) | 0 | 0 |
| Chloramphenicol | cat1 | 1 (4) | 4 (16) | 1 (4) | 5 (20) | 5 (20) | 5 (20) | 5 (20) | 2 (8) | 5 (20) | 2 (8) | 0 | 0 | 1 (4) | 0 | 0 |
| cmlA | 1 (4) | 2 (8) | 1 (4) | 3 (12) | 3 (12) | 1 (4) | 3 (12) | 1 (4) | 2 (8) | 1 (4) | 0 | 0 | 1 (4) | 0 | 0 | |
| Tetracycline | tet(A) | 6 (24) | 14 (56) | 5 (20) | 21 (84) | 15 (60) | 14 (56) | 20 (80) | 5 (20) | 18 (72) | 4 (16) | 0 | 0 | 3 (12) | 0 | 0 |
| tet(B) | 0 | 2 (8) | 0 | 2 (8) | 2 (8) | 1 (4) | 1 (4) | 1 (4) | 1 (4) | 0 | 0 | 0 | 1 (4) | 0 | 0 | |
| Trimethoprim | dfrA1 | 0 | 0 | 1 (4) | 1 (4) | 1 (4) | 0 | 1 (4) | 0 | 1 (4) | 0 | 0 | 0 | 0 | 0 | 0 |
| Quinolones | qnr | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| p-value | 7.1 × 10−11 | 0.0004 | 1.2 × 10−7 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | - | - | 1 | - | - | |
| Community-Acquired Infections (n = 125) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Antibiotic Resistance Gene | Number of Strains (%) | |||||||||||||||
| Adhesins | Iron-Acquisition Systems | Protectins | Toxins | |||||||||||||
| fimH | mrkD | kpn | ycfM | entB | irp1 | irp2 | ybtS | fyuA | iutA | iroN | magA | rmpA | hlyA | cnf-1 | ||
| Beta-lactams | blaSHV | 31 (24.8) | 40 (32) | 8 (6.4) | 72 (57.6) | 56 (44.8) | 51 (40.8) | 76 (60.8) | 5 (4) | 58 (46.4) | 28 (22.4) | 12 (9.6) | 0 | 26 (20.8) | 3 (2.4) | 0 |
| blaCITM | 18 (14.4) | 30 (24) | 6 (4.8) | 56 (44.8) | 41 (32.8) | 38 (30.4) | 59 (47.2) | 3 (2.4) | 45 (36) | 23 (18.4) | 8 (6.4) | 0 | 18 (14.4) | 3 (2.4) | 0 | |
| blaTEM | 38 (30.4) | 57 (45.6) | 14 (11.2) | 91 (72.8) | 73 (58.4) | 65 (52) | 98 (78.4) | 8 (6.4) | 75 (60) | 39 (31.2) | 12 (9.6) | 1 (0.8) | 35 (28) | 5 (4) | 0 | |
| blaCTXM-1 | 14 (11.2) | 20 (16) | 6 (4.8) | 36 (28.8) | 26 (20.8) | 27 (21.6) | 42 (33.6) | 3 (2.4) | 30 (24) | 12 (9.6) | 6 (4.8) | 0 | 9 (7.2) | 2 (1.6) | 0 | |
| blaCTXM-2 | 1 (0.8) | 1 (0.8) | 0 | 1 (0.8) | 2 (1.6) | 2 (1.6) | 2 (1.6) | 0 | 2 (1.6) | 1 (0.8) | 0 | 0 | 1 (0.8) | 0 | 0 | |
| blaCTXM-9 | 3 (2.4) | 3 (2.4) | 1 (0.8) | 10 (8) | 5 (4) | 4 (3.2) | 5 (4) | 0 | 5 (4) | 4 (3.2) | 2 (1.6) | 0 | 1 (0.8) | 0 | 0 | |
| Streptomycin | aadA1 | 15 (12) | 23 (18.4) | 4 (3.2) | 39 (31.2) | 31 (24.8) | 26 (20.8) | 40 (32) | 2 (1.6) | 32 (25.6) | 17 (13.6) | 5 (4) | 0 | 10 (8) | 1 (0.8) | 0 |
| Gentamicin | aac(3)-IV | 17 (13.6) | 23 (18.4) | 6 (4.8) | 41 (32.8) | 35 (28) | 29 (23.2) | 48 (38.4) | 1 (0.8) | 36 (28.8) | 18 (14.4) | 8 (6.4) | 1 (0.8) | 8 (6.4) | 3 (2.4) | 0 |
| Sulfonamide | sul1 | 24 (19.2) | 32 (25.6) | 9 (7.2) | 52 (41.6) | 42 (33.6) | 41 (32.8) | 58 (46.4) | 2 (1.6) | 46 (36.8) | 18 (14.4) | 3 (2.4) | 0 | 15 (12) | 3 (2.4) | 0 |
| Chloramphenicol | cat1 | 4 (3.2) | 4 (3.2) | 1 (0.8) | 6 (4.8) | 6 (4.8) | 6 (4.8) | 8 (6.4) | 1 (0.8) | 6 (4.8) | 1 (0.8) | 1 (0.8) | 0 | 1 (0.8) | 0 | 0 |
| cmlA | 6 (4.8) | 7 (5.6) | 3 (2.4) | 12 (9.6) | 8 (6.4) | 14 (11.2) | 16 (12.8) | 1 (0.8) | 17 (13.6) | 5 (4) | 4 (3.2) | 0 | 4 (3.2) | 0 | 0 | |
| Tetracycline | tet(A) | 26 (20.8) | 35 (28) | 12 (9.6) | 64 (51.2) | 48 (38.4) | 48 (38.4) | 71 (56.8) | 5 (4) | 52 (41.6) | 24 (19.2) | 9 (7.2) | 1 (0.8) | 21 (16.8) | 5 | 0 |
| tet(B) | 4 (3.2) | 2 (1.6) | 2 (1.6) | 7 (5.6) | 4 (3.2) | 7 (5.6) | 10 (8) | 1 (0.8) | 5 (4) | 3 (2.4) | 0 | 0 | 0 | 0 | 0 | |
| Trimethoprim | dfrA1 | 7 (5.6) | 10 (8) | 1 (0.8) | 20 (16) | 18 (14.4) | 15 (12) | 19 (15.2) | 2 (1.6) | 14 (11.2) | 9 (7.2) | 3 (2.4) | 0 | 5 (4) | 1 (0.8) | 0 |
| Quinolones | qnr | 0 | 0 | 0 | 1 (0.8) | 1 (0.8) | 0 | 1 (0.8) | 0 | 1 (0.8) | 0 | 0 | 0 | 0 | 0 | 0 |
| p-value | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 0.0004 | 1 | 0.0004 | 0.0004 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paniagua-Contreras, G.L.; Bautista-Cerón, A.; Morales-Espinosa, R.; Delgado, G.; Vaca-Paniagua, F.; Díaz-Velásquez, C.E.; de la Cruz-Montoya, A.H.; García-Cortés, L.R.; Sánchez-Yáñez, M.P.; Monroy-Pérez, E. Extensive Expression of the Virulome Related to Antibiotic Genotyping in Nosocomial Strains of Klebsiella pneumoniae. Int. J. Mol. Sci. 2023, 24, 14754. https://doi.org/10.3390/ijms241914754
Paniagua-Contreras GL, Bautista-Cerón A, Morales-Espinosa R, Delgado G, Vaca-Paniagua F, Díaz-Velásquez CE, de la Cruz-Montoya AH, García-Cortés LR, Sánchez-Yáñez MP, Monroy-Pérez E. Extensive Expression of the Virulome Related to Antibiotic Genotyping in Nosocomial Strains of Klebsiella pneumoniae. International Journal of Molecular Sciences. 2023; 24(19):14754. https://doi.org/10.3390/ijms241914754
Chicago/Turabian StylePaniagua-Contreras, Gloria Luz, Areli Bautista-Cerón, Rosario Morales-Espinosa, Gabriela Delgado, Felipe Vaca-Paniagua, Clara Estela Díaz-Velásquez, Aldo Hugo de la Cruz-Montoya, Luis Rey García-Cortés, María Patricia Sánchez-Yáñez, and Eric Monroy-Pérez. 2023. "Extensive Expression of the Virulome Related to Antibiotic Genotyping in Nosocomial Strains of Klebsiella pneumoniae" International Journal of Molecular Sciences 24, no. 19: 14754. https://doi.org/10.3390/ijms241914754
APA StylePaniagua-Contreras, G. L., Bautista-Cerón, A., Morales-Espinosa, R., Delgado, G., Vaca-Paniagua, F., Díaz-Velásquez, C. E., de la Cruz-Montoya, A. H., García-Cortés, L. R., Sánchez-Yáñez, M. P., & Monroy-Pérez, E. (2023). Extensive Expression of the Virulome Related to Antibiotic Genotyping in Nosocomial Strains of Klebsiella pneumoniae. International Journal of Molecular Sciences, 24(19), 14754. https://doi.org/10.3390/ijms241914754