
Melatonin and Vitamins as Protectors against the Reproductive Toxicity of Bisphenols: Which Is the Most Effective? A Systematic Review and Meta-Analysis

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
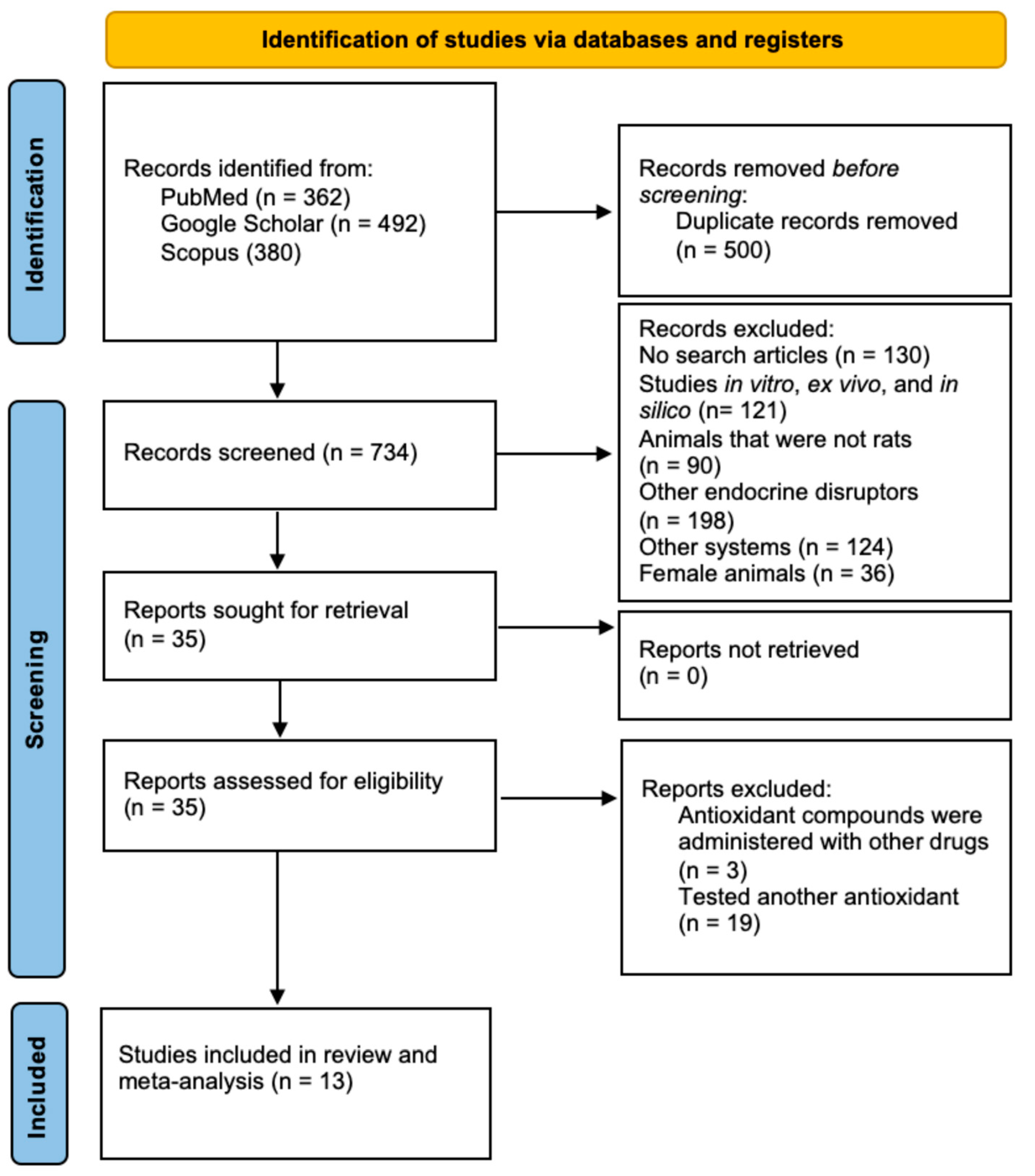
2.1. Selection and Identification of Studies
2.2. Study Characteristics
2.3. Meta-Analysis
2.3.1. Final Body Weight
2.3.2. Testosterone
2.3.3. Sperm Characteristics
2.3.4. Testis Weights
2.3.5. Outliers
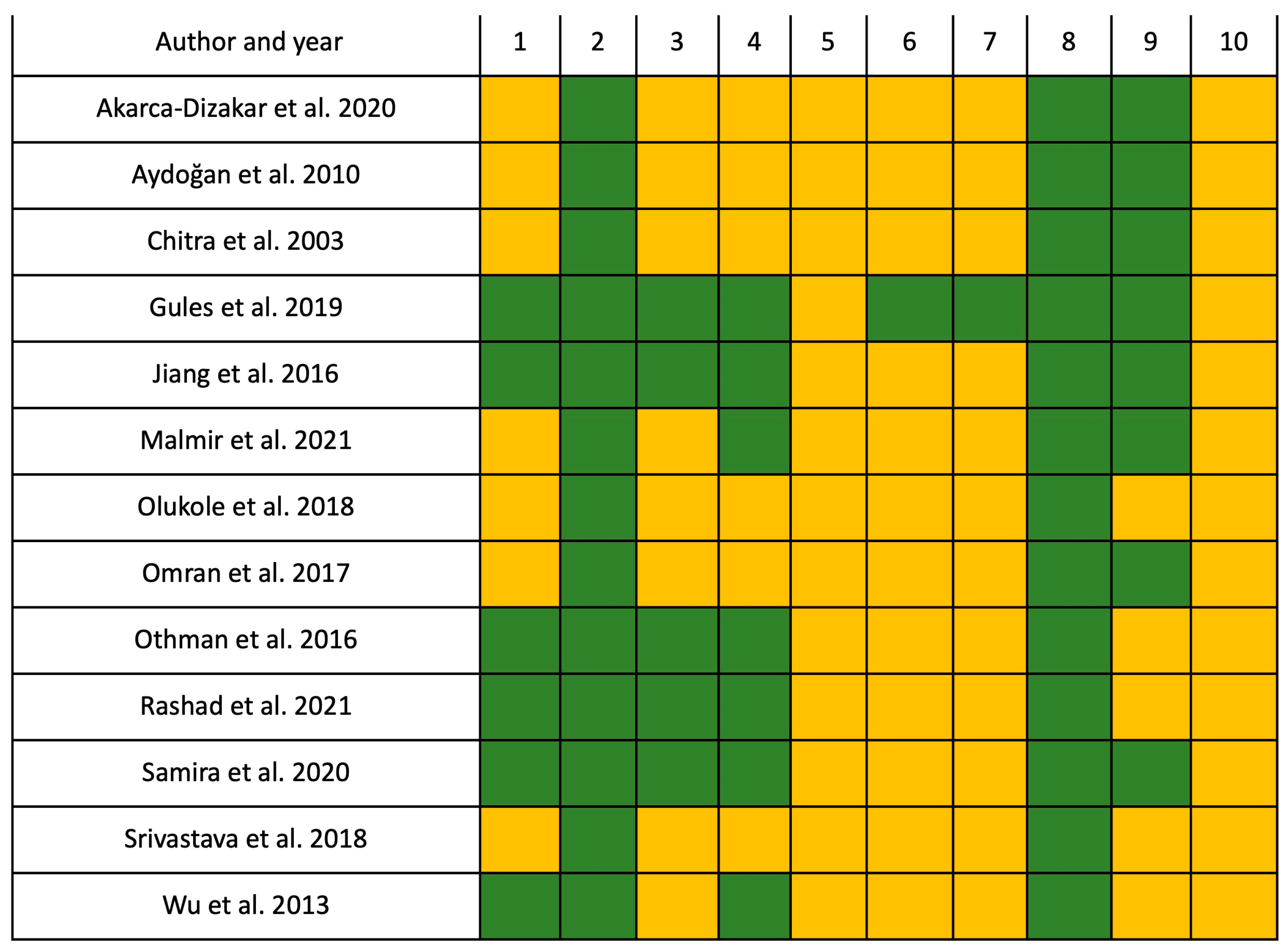
2.4. Risk of Bias Assessment
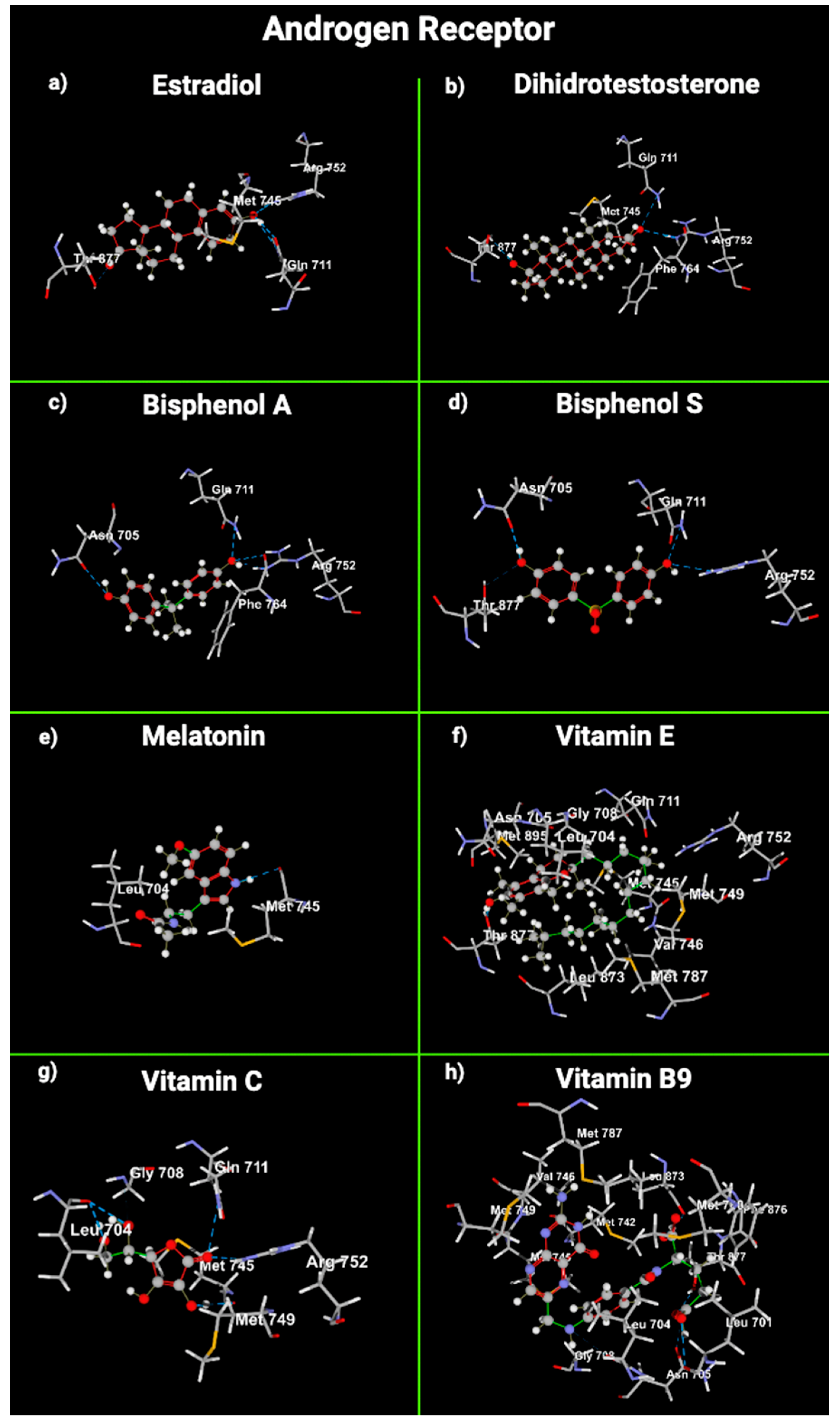
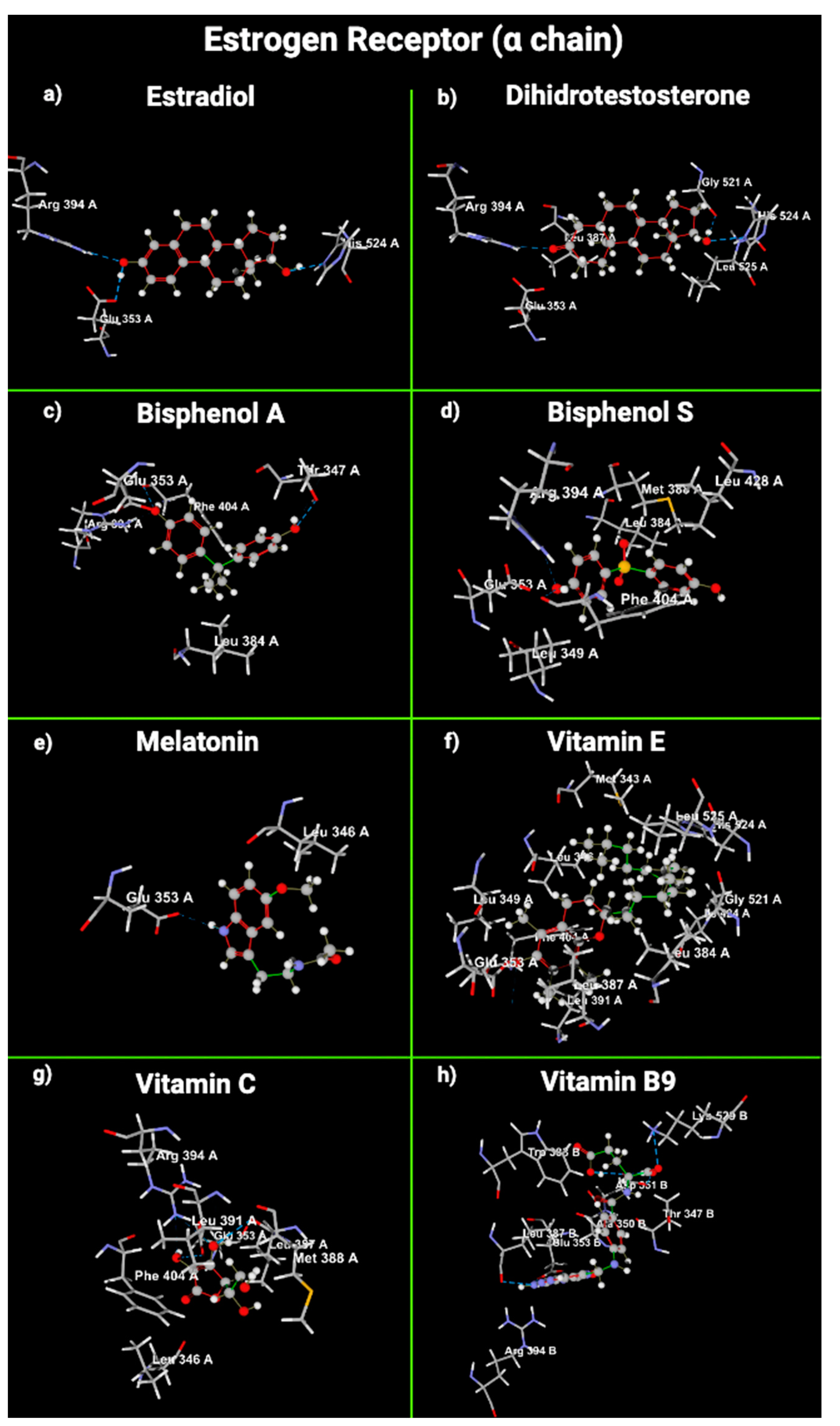
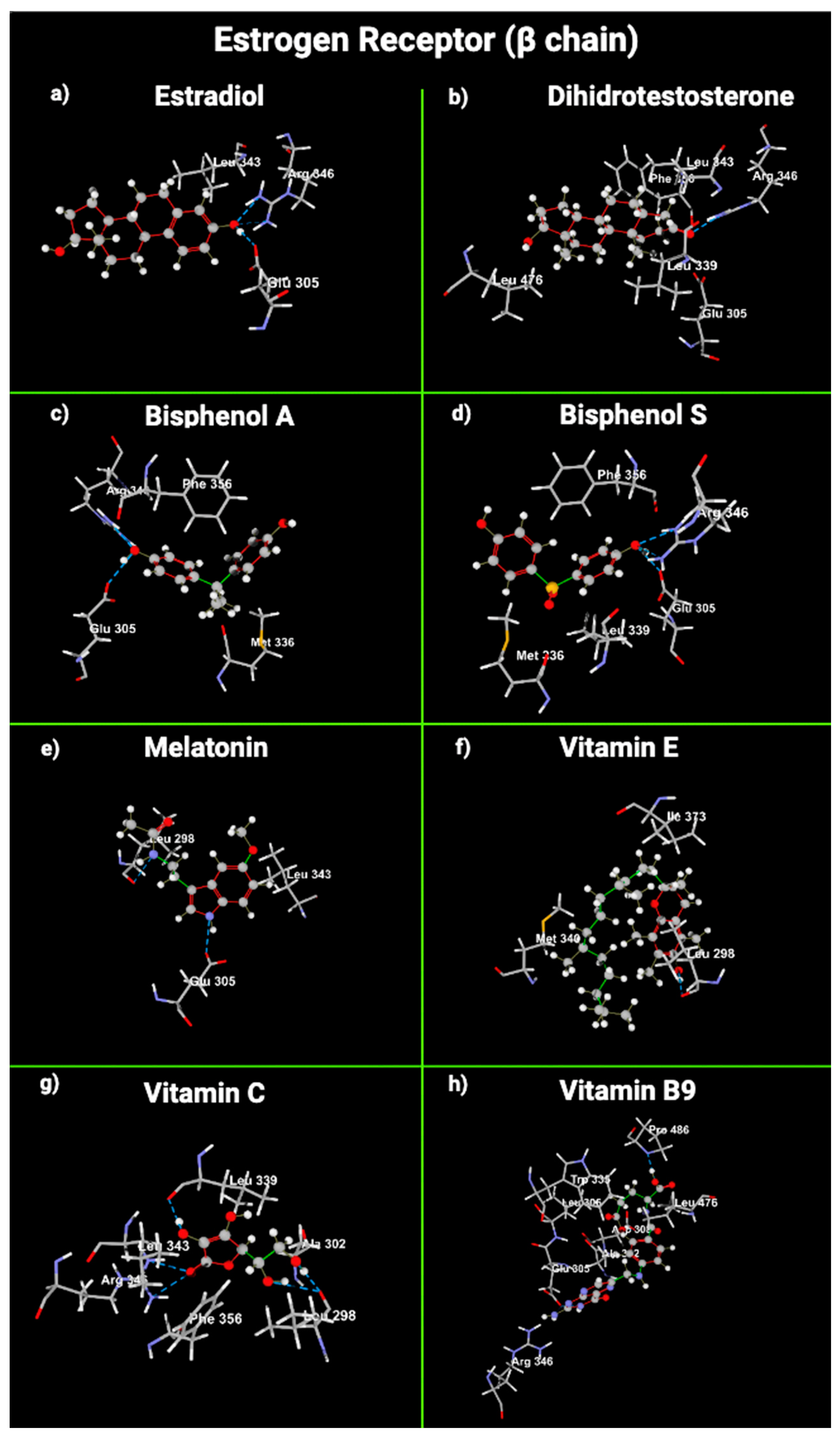
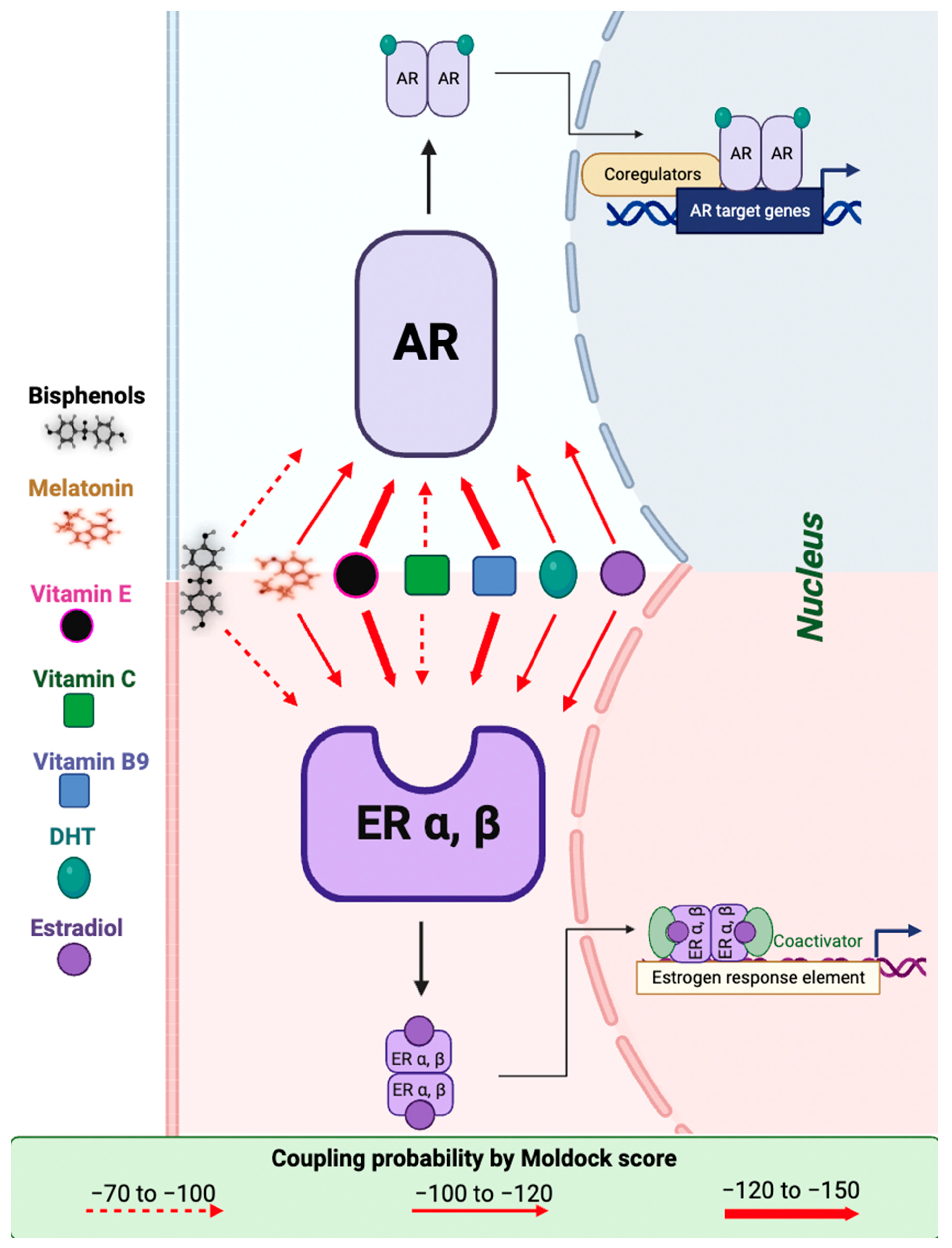
2.5. Molecular Modeling of Melatonin and Vitamin Interactions with Estrogen and Testosterone Receptors
3. Discussion
4. Materials and Methods
4.1. Study Selection and Eligibility Criteria
4.2. Data Extraction
- Article characteristics (first author and publication year).
- Rat population characteristics (rat strain, total number of animals (intervention, control), weight (g), initial age of exposure (weeks), and age of sample collection (weeks).
- Antioxidant compound (melatonin or vitamins, dose (mg/kg bw/d), the span of exposure (days), administration route (intraperitoneally: i.p., orally: p.o.).
- Bisphenol dose (mg/kg bw/d), the span of exposure (days), administration route (i.p. or p.o.)
- Primary outcomes (reproductive parameters).
4.3. Statistical Analysis
- Data from the bisphenol vs. control group to identify the damage produced by the administration of bisphenols to rats without antioxidant treatment.
- Data from the bisphenol plus antioxidant (melatonin or vitamins) vs. bisphenol group to determine if the antioxidants restored the adverse effects produced by bisphenol administration alone.
- Data from the bisphenol plus melatonin or vitamins vs. control group to determine whether administration of antioxidants returns the reproductive variable to control group levels.
4.4. Study Quality
4.5. Molecular Docking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sutunkova, M.P.; Solovyeva, S.N.; Minigalieva, I.A.; Gurvich, V.B.; Valamina, I.E.; Makeyev, O.H.; Shur, V.Y.; Shishkina, E.V.; Zubarev, I.V.; Saatkhudinova, R.R.; et al. Toxic Effects of Low-Level Long-Term Inhalation Exposures of Rats to Nickel Oxide Nanoparticles. Int. J. Mol. Sci. 2019, 20, 1778. [Google Scholar] [CrossRef] [PubMed]
- Minigalieva, I.A.; Katsnelson, B.A.; Privalova, L.I.; Sutunkova, M.P.; Gurvich, V.B.; Shur, V.Y.; Shishkina, E.V.; Valamina, I.E.; Makeyev, O.H.; Panov, V.G. Attenuation of Combined Nickel(II) Oxide and Manganese(II, III) Oxide Nanoparticles’ Adverse Effects with a Complex of Bioprotectors. Int. J. Mol. Sci. 2015, 16, 22555–22583. [Google Scholar] [CrossRef]
- Klinova, S.V.; Minigalieva, I.A.; Privalova, L.I.; Valamina, I.E.; Makeyev, O.H.; Shuman, E.A.; Korotkov, A.A.; Panov, V.G.; Sutunkova, M.P.; Ryabova, J.V. Further verification of some postulates of the combined toxicity theory: New animal experimental data on separate and joint adverse effects of lead and cadmium. Food Chem. Toxicol. 2020, 136, 110971. [Google Scholar] [CrossRef]
- Chitra, K.C.; Rao, K.R.; Mathur, P.P. Effect of bisphenol A and co-administration of bisphenol A and vitamin C on epididymis of adult rats: A histological and biochemical study. Asian J. Androl. 2003, 5, 203–208. [Google Scholar]
- Vandenberg, L.N.; Hunt, P.A.; Myers, J.P.; Vom Saal, F.S. Human exposures to bisphenol A: Mismatches between data and assumptions. Rev. Environ. Health 2013, 28, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Kleywegt, S.; Pileggi, V.; Yang, P.; Hao, C.; Zhao, X.; Rocks, C.; Thach, S.; Cheung, P.; Whitehead, B. Pharmaceuticals, hormones and bisphenol A in untreated source and finished drinking water in Ontario, Canada—Occurrence and treatment efficiency. Sci. Total Environ. 2011, 409, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Bošnjak, I.; Borra, M.; Iamunno, F.; Benvenuto, G.; Ujević, I.; Bušelić, I.; Roje-Busatto, R.; Mladineo, I. Effect of bisphenol A on P-glycoprotein-mediated efflux and ultrastructure of the sea urchin embryo. Aquat. Toxicol. 2014, 156, 21–29. [Google Scholar] [CrossRef]
- Cabaton, N.J.; Canlet, C.; Wadia, P.R.; Tremblay-Franco, M.; Gautier, R.; Molina, J.; Sonnenschein, C.; Cravedi, J.P.; Rubin, B.S.; Soto, A.M.; et al. Effects of low doses of bisphenol A on the metabolome of perinatally exposed CD-1 mice. Environ. Health Perspect. 2013, 121, 586–593. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, H.Q.; Cui, Z.H.; Yin, L.; Zhang, W.L.; Liu, W.B.; Han, F.; Ao, L.; Cao, J.; Liu, J.Y. Low-dose and combined effects of oral exposure to bisphenol A and diethylstilbestrol on the male reproductive system in adult Sprague-Dawley rats. Environ. Toxicol. Pharmacol. 2016, 43, 94–102. [Google Scholar] [CrossRef]
- Tian, J.; Ding, Y.; She, R.; Ma, L.; Du, F.; Xia, K.; Chen, L. Histologic study of testis injury after bisphenol A exposure in mice. Toxicol. Ind. Health 2017, 33, 36–45. [Google Scholar] [CrossRef]
- Sartain, C.V.; Hunt, P.A. An old culprit but a new story: Bisphenol A and “NextGen” bisphenols. Fertil. Steril. 2016, 106, 820–826. [Google Scholar] [CrossRef]
- Reif, D.M.; Martin, M.T.; Tan, S.W.; Houck, K.A.; Judson, R.S.; Richard, A.M.; Knudsen, T.B.; Dix, D.J.; Kavlock, R.J. Endocrine profiling and prioritization of environmental chemicals using ToxCast data. Environ. Health Perspect. 2010, 118, 1714–1720. [Google Scholar] [CrossRef] [PubMed]
- Thoene, M.; Thoene, M.; Rytel, L.; Nowicka, N.; Wojtkiewicz, J. The state of bisphenol research in the lesser developed countries of the EU: A mini-review. Toxicol. Res. 2018, 7, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Shankar, A.; Teppala, S. Relationship between urinary bisphenol A levels and diabetes mellitus. J. Clin. Endocrinol. Metab. 2011, 96, 3822–3826. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.M.; Kalkbrenner, A.E.; Calafat, A.M.; Yolton, K.; Ye, X.; Dietrich, K.N.; Lanphear, B.P. Impact of early-life bisphenol A exposure on behavior and executive function in children. Pediatrics 2011, 128, 873–882. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Roychoudhury, S.; Sharma, R.; Gupta, S.; Majzoub, A.; Sabanegh, E. Diagnostic application of oxidation-reduction potential assay for measurement of oxidative stress: Clinical utility in male factor infertility. Reprod. Biomed. Online 2017, 34, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Baker, M.A. Oxidative stress, sperm survival and fertility control. Mol. Cell Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef]
- Kitamura, S.; Suzuki, T.; Sanoh, S.; Kohta, R.; Jinno, N.; Sugihara, K.; Yoshihara, S.; Fujimoto, N.; Watanabe, H.; Ohta, S. Comparative study of the endocrine-disrupting activity of bisphenol A and 19 related compounds. Toxicol. Sci. 2005, 84, 249–259. [Google Scholar] [CrossRef]
- Matsushima, A.; Teramoto, T.; Okada, H.; Liu, X.; Tokunaga, T.; Kakuta, Y.; Shimohigashi, Y. ERRgamma tethers strongly bisphenol A and 4-alpha-cumylphenol in an induced-fit manner. Biochem. Biophys. Res. Commun. 2008, 373, 408–413. [Google Scholar] [CrossRef]
- Krüger, T.; Long, M.; Bonefeld-Jørgensen, E.C. Plastic components affect the activation of the aryl hydrocarbon and the androgen receptor. Toxicology 2008, 246, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Virant-Klun, I.; Imamovic-Kumalic, S.; Pinter, B. From Oxidative Stress to Male Infertility: Review of the Associations of Endocrine-Disrupting Chemicals (Bisphenols, Phthalates, and Parabens) with Human Semen Quality. Antioxidants 2022, 11, 1617. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Akarca-Dizakar, S.Ö.; Erdoğan, D.; Peker, T.; Coşkun Akçay, N.; Türkoğlu, I.; Eşmekaya, M.A.; Ömeroğlu, S. Effects of co-administered melatonin, fructose and bisphenol A (BPA) on rat epididymis and sperm characteristics. Biotech. Histochem. 2020, 95, 18–26. [Google Scholar] [CrossRef]
- Eshak, M.G.; Osman, H.F. Effects of Chitosan against Bisphenol- A Induced EndocrineToxicity and Androgen Receptor Gene Expression Changes in Male Rats. Int. J. Pharm. Clin. Res. 2014, 6, 300–311. [Google Scholar]
- Behmanesh, M.A.; Najafzadehvarzi, H.; Poormoosavi, S.M. Protective Effect of Aloe vera Extract against Bisphenol A Induced Testicular Toxicity in Wistar Rats. Cell J. 2018, 20, 278–283. [Google Scholar]
- Yousaf, B.; Amina; Liu, G.; Wang, R.; Qadir, A.; Ali, M.U.; Kanwal, Q.; Munir, B.; Asmatullah; Abbas, Z. Bisphenol A exposure and healing effects of Adiantum capillus-veneris L. plant extract (APE) in bisphenol A-induced reproductive toxicity in albino rats. Environ. Sci. Pollut. Res. Int. 2016, 23, 11645–11657. [Google Scholar] [CrossRef]
- Zahra, Z.; Khan, M.R.; Majid, M.; Maryam, S.; Sajid, M. Gonadoprotective ability of Vincetoxicum arnottianum extract against bisphenol A-induced testicular toxicity and hormonal imbalance in male Sprague Dawley rats. Andrologia 2020, 52, e13590. [Google Scholar] [CrossRef]
- Sahu, C.; Charaya, A.; Singla, S.; Dwivedi, D.K.; Jena, G. Zinc deficient diet increases the toxicity of bisphenol A in rat testis. J. Biochem. Mol. Toxicol. 2020, 34, e22549. [Google Scholar] [CrossRef]
- Khalaf, A.A.; Ahmed, W.; Moselhy, W.A.; Abdel-Halim, B.R.; Ibrahim, M.A. Protective effects of selenium and nano-selenium on bisphenol-induced reproductive toxicity in male rats. Hum. Exp. Toxicol. 2019, 38, 398–408. [Google Scholar] [CrossRef]
- Srivastava, S.; Gupta, P. Alteration in apoptotic rate of testicular cells and sperms following administration of Bisphenol A (BPA) in Wistar albino rats. Environ. Sci. Pollut. Res. Int. 2018, 25, 21635–21643. [Google Scholar] [CrossRef] [PubMed]
- Gules, O.; Yildiz, M.; Naseer, Z.; Tatar, M. Effects of folic acid on testicular toxicity induced by bisphenol-A in male Wistar rats. Biotech. Histochem. 2019, 94, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Aydoğan, M.; Korkmaz, A.; Barlas, N.; Kolankaya, D. Pro-oxidant effect of vitamin C coadministration with bisphenol A, nonylphenol, and octylphenol on the reproductive tract of male rats. Drug Chem. Toxicol. 2010, 33, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Das, U.B.; Mallick, M.; Debnath, J.M.; Ghosh, D. Protective effect of ascorbic acid on cyclophosphamide- induced testicular gametogenic and androgenic disorders in male rats. Asian J. Androl. 2002, 4, 201–207. [Google Scholar]
- Haghi-Aminjan, H.; Asghari, M.H.; Farhood, B.; Rahimifard, M.; Hashemi Goradel, N.; Abdollahi, M. The role of melatonin on chemotherapy-induced reproductive toxicity. J. Pharm. Pharmacol. 2018, 70, 291–306. [Google Scholar] [CrossRef]
- Othman, A.I.; Edrees, G.M.; El-Missiry, M.A.; Ali, D.A.; Aboel-Nour, M.; Dabdoub, B.R. Melatonin controlled apoptosis and protected the testes and sperm quality against bisphenol A-induced oxidative toxicity. Toxicol. Ind. Health 2016, 32, 1537–1549. [Google Scholar] [CrossRef]
- Olukole, S.G.; Ajani, S.O.; Ola-Davies, E.O.; Lanipekun, D.O.; Aina, O.O.; Oyeyemi, M.O.; Oke, B.O. Melatonin ameliorates bisphenol A-induced perturbations of the prostate gland of adult Wistar rats. Biomed. Pharmacother. 2018, 105, 73–82. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviewsDeclaración PRISMA 2020: Una guía actualizada para la publicación de revisiones sistemáticas. Rev. Panam. Salud Publica 2022, 46, e112. [Google Scholar]
- Wu, H.J.; Liu, C.; Duan, W.X.; Xu, S.C.; He, M.D.; Chen, C.H.; Wang, Y.; Zhou, Z.; Yu, Z.P.; Zhang, L. Melatonin ameliorates bisphenol A-induced DNA damage in the germ cells of adult male rats. Mutat. Res. 2013, 752, 57–67. [Google Scholar] [CrossRef]
- Samira, N.; Malmir, M.; Mehranji, M.; Faraji, T.; Khodabandehlo, H. Potential Ameliorative Action of Vitamin E on Spermatogenesis and Sperm Count in Bisphenol A-Treated Rats: Reproductive Health Research. Biomed. J. Sci. Tech. Res. 2020, 29, 22693–22701. [Google Scholar]
- Jiang, Z.; Wang, J.; Li, X.; Zhang, X. Echinacoside and Cistanche tubulosa (Schenk) R. wight ameliorate bisphenol A-induced testicular and sperm damage in rats through gonad axis regulated steroidogenic enzymes. J. Ethnopharmacol. 2016, 193, 321–328. [Google Scholar] [CrossRef]
- Malmir, M.; Mehranjani, M.S.; Faraji, T.; Noreini, S. Antioxidant effect of Vitamin E on the male rat reproductive system by a high oral dose of Bisphenol-A. Toxicol. Res. Appl. 2021, 5, 23978473211005562. [Google Scholar] [CrossRef]
- Omran, B.; Abdallah, E.; Abdelwahab, M. Study of Probable Toxic Effects of Bisphenol A & the Protective Role of Vitamin E on Testes and Prostate of Adult Male Albino Rats. Ain Shams J. Forensic Med. Clin. Toxicol. 2017, 29, 7–18. [Google Scholar]
- Rashad, S.; Ahmed, S.; El-Sayed, M.; Ahmed, D. The Toxic Effect of Bisphenol A on Albino Rat Testicles and the Possible Protective Value of Vitamin E and Melatonin. Egypt. Soc. Clin. Toxicol. J. 2021, 9, 1–12. [Google Scholar] [CrossRef]
- Pena-Corona, S.I.; Vásquez Aguire, W.S.; Vargas, D.; Juárez, I.; Mendoza-Rodríguez, C.A. Effects of bisphenols on Blood-Testis Barrier protein expression in vitro: A systematic review and meta-analysis. Reprod. Toxicol. 2021, 103, 139–148. [Google Scholar] [CrossRef]
- Peña-Corona, S.I.; Vargas-Estrada, D.; Juárez-Rodríguez, I.; Retana-Márquez, S.; Mendoza-Rodríguez, C.A. Bisphenols as promoters of the dysregulation of cellular junction proteins of the blood-testis barrier in experimental animals: A systematic review of the literature. J. Biochem. Mol. Toxicol. 2023, 37, e23416. [Google Scholar] [CrossRef]
- Ahmed, A.E.; Alshehri, A.; Al-Kahtani, M.A.; Elbehairi, S.E.I.; Alshehri, M.A.; Shati, A.A.; Alfaifi, M.Y.; Al-Doais, A.A.; Taha, R.; Morsy, K. Vitamin E and selenium administration synergistically mitigates ivermectin and doramectin-induced testicular dysfunction in male Wistar albino rats. Biomed. Pharmacother. 2020, 124, 109841. [Google Scholar] [CrossRef]
- Khan, D.; Ahmed, S.A. Epigenetic Regulation of Non-Lymphoid Cells by Bisphenol A, a Model Endocrine Disrupter: Potential Implications for Immunoregulation. Front. Endocrinol. 2015, 6, 91. [Google Scholar] [CrossRef]
- Sofikitis, N.; Giotitsas, N.; Tsounapi, P.; Baltogiannis, D.; Giannakis, D.; Pardalidis, N. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol. 2008, 109, 323–330. [Google Scholar] [CrossRef]
- Tamilselvan, P.; Langeswaran, K.; Vijayaprakash, S.; Kumar, S.G.; Revathy, R.; Balasubramanian, M.P. Efficiency of lycopene against reproductive and developmental toxicity of Bisphenol A in male Sprague Dawley rats. Biomed. Prev. Nutr. 2014, 4, 491–498. [Google Scholar] [CrossRef]
- Muthuvel, R.; Venkataraman, P.; Krishnamoorthy, G.; Gunadharini, D.N.; Kanagaraj, P.; Jone Stanley, A.; Srinivasan, N.; Balasubramanian, K.; Aruldhas, M.M.; Arunakaran, J. Antioxidant effect of ascorbic acid on PCB (Aroclor 1254) induced oxidative stress in hypothalamus of albino rats. Clin. Chim. Acta 2006, 365, 297–303. [Google Scholar] [CrossRef]
- Sikka, S.C. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem. 2001, 8, 851–862. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Singh, G.; Sadwal, S.; Aniqa, A. Alleviating impact of hydroethanolic Murraya koenigii leaves extract on bisphenol A instigated testicular lethality and apoptosis in mice. Andrologia 2020, 52, e13504. [Google Scholar] [CrossRef] [PubMed]
- Yoganathan, T.; Eskild, W.; Hansson, V. Investigation of detoxification capacity of rat testicular germ cells and Sertoli cells. Free Radic. Biol. Med. 1989, 7, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Ayinde, O.C.; Ogunnowo, S.; Ogedegbe, R.A. Influence of Vitamin C and Vitamin E on testicular zinc content and testicular toxicity in lead exposed albino rats. BMC Pharmacol. Toxicol. 2012, 13, 17. [Google Scholar] [CrossRef]
- Gavazza, M.; Catalá, A. The effect of alpha-tocopherol on the lipid peroxidation of mitochondria and microsomes obtained from rat liver and testis. Mol. Cell Biochem. 2001, 225, 121–128. [Google Scholar] [CrossRef]
- Ibrahim, W.; Tousson, E.; Ali, E.M.; Mansour, M.A. Folic acid alleviates oxidative stress and hyperhomocysteinemia involved in testicular dysfunction of hypothyroid rats. Gen. Comp. Endocrinol. 2011, 174, 143–149. [Google Scholar] [CrossRef]
- Shalaby, M.A.; El Zorba, H.Y.; Ziada, R.M. Reproductive toxicity of methomyl insecticide in male rats and protective effect of folic acid. Food Chem. Toxicol. 2010, 48, 3221–3226. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- Fang, Y.; Zhou, Y.; Zhong, Y.; Gao, X.; Tan, T. Effect of vitamin E on reproductive functions and anti-oxidant activity of adolescent male mice exposed to bisphenol A. Wei Sheng Yan Jiu 2013, 42, 18–22. [Google Scholar]
- Peña-Corona, S.I.; Vargas-Estrada, D.; Chávez-Corona, J.I.; Mendoza-Rodríguez, C.A.; Caballero-Chacón, S.; Pedraza-Chaverri, J.; Gracia-Mora, M.I.; Galván-Vela, D.P.; García-Rodríguez, H.; Sánchez-Bartez, F. Vitamin E (α-Tocopherol) Does Not Ameliorate the Toxic Effect of Bisphenol S on the Metabolic Analytes and Pancreas Histoarchitecture of Diabetic Rats. Toxics 2023, 11, 626. [Google Scholar] [CrossRef] [PubMed]
- Preethi, S.; Sandhya, K.; Sreedevi, B.; Chandrasekhar, K.B.; Kumari, J.P. Toxicity of Bisphenol a on Humans: A Review. Int. Lett. Nat. Sci. 2014, 22, 32–46. [Google Scholar] [CrossRef]
- Sikka, S.C. Oxidative stress and role of antioxidants in normal and abnormal sperm function. Front. Biosci. 1996, 1, e78–e86. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Rhee, S.K. Inhibitory effect of bisphenol A on gap junctional intercellular communication in an epithelial cell line of rat mammary tissue. Arch. Pharm. Res. 2007, 30, 337–343. [Google Scholar] [CrossRef]
- Helal, E.G.E.; Soliman, M.G.; Badawi, M.M.M.; Abdel-Kawi, N.A.; Fadel, H.A.E.; Abozaid, N.M.G. Physiological and Histopathological studies on Bisphenol-A compound as xenoestrogen in male albino rats. Egypt. J. Hosp. Med. 2018, 50, 127–136. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Khallouki, F.; de Medina, P.; Caze-Subra, S.; Bystricky, K.; Balaguer, P.; Poirot, M.; Silvente-Poirot, S. Molecular and Biochemical Analysis of the Estrogenic and Proliferative Properties of Vitamin E Compounds. Front. Oncol. 2015, 5, 287. [Google Scholar] [CrossRef]
- Wang, Y.; Song, J.; Li, Y.; Lin, C.; Chen, Y.; Zhang, X.; Yu, H. Melatonin inhibited the progression of gastric cancer induced by Bisphenol S via regulating the estrogen receptor 1. Ecotoxicol. Environ. Saf. 2023, 259, 115054. [Google Scholar] [CrossRef]
- Rocha, C.S.; Rato, L.; Martins, A.D.; Alves, M.G.; Oliveira, P.F. Melatonin and male reproductive health: Relevance of darkness and antioxidant properties. Curr. Mol. Med. 2015, 15, 299–311. [Google Scholar] [CrossRef]
- Yu, K.; Deng, S.L.; Sun, T.C.; Li, Y.Y.; Liu, Y.X. Melatonin Regulates the Synthesis of Steroid Hormones on Male Reproduction: A Review. Molecules 2018, 23, 447. [Google Scholar] [CrossRef]
- del Río, B.; del Río, B.; García Pedrero, J.M.; Martínez-Campa, C.; Zuazua, P.; Lazo, P.S.; Ramos, S. Melatonin, an endogenous-specific inhibitor of estrogen receptor alpha via calmodulin. J. Biol. Chem. 2004, 279, 38294–38302. [Google Scholar] [CrossRef] [PubMed]
- Jayashree, B.S.; Thomas, S.; Nayak, Y. Design and synthesis of 2-quinolones as antioxidants and antimicrobials: A rational approach. Med. Chem. Res. 2010, 19, 193–209. [Google Scholar] [CrossRef]
- Thomsen, R.; Christensen, M.H. MolDock: A New Technique for High-Accuracy Molecular Docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Naamneh Elzenaty, R.; du Toit, T.; Flück, C.E. Basics of androgen synthesis and action. Best. Pract. Res. Clin. Endocrinol. Metab. 2022, 36, 101665. [Google Scholar] [CrossRef]
- O’Hara, L.; Smith, L.B. Androgen receptor roles in spermatogenesis and infertility. Best. Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 595–605. [Google Scholar] [CrossRef]
- Cooke, P.S.; Young, P.; Hess, R.A.; Cunha, G.R. Estrogen receptor expression in developing epididymis, efferent ductules, and other male reproductive organs. Endocrinology 1991, 128, 2874–2879. [Google Scholar] [CrossRef]
- Holderegger, C.; Keefer, D. The ontogeny of the mouse estrogen receptor: The pelvic region. Am. J. Anat. 1986, 177, 285–297. [Google Scholar] [CrossRef]
- Leung, H.W.; Ballantyne, B. Developmental toxicity evaluation of rats dosed orally or cutaneously with octoxynol-9. J. Appl. Toxicol. 1999, 19, 267–273. [Google Scholar] [CrossRef]
- Bosch, R.J.; Quiroga, B.; Muñoz-Moreno, C.; Olea-Herrero, N.; Arenas, M.I.; González-Santander, M.; Reventún, P.; Zaragoza, C.; de Arriba, G.; Saura, M. Bisphenol A: An environmental factor implicated in renal vascular damage. Nefrologia 2016, 36, 5–9. [Google Scholar] [CrossRef]
- Liao, C.; Liu, F.; Alomirah, H.; Loi, V.D.; Mohd, M.A.; Moon, H.B.; Nakata, H.; Kannan, K. Bisphenol S in urine from the United States and seven Asian countries: Occurrence and human exposures. Environ. Sci. Technol. 2012, 46, 6860–6866. [Google Scholar] [CrossRef]
- Zhang, B.; He, Y.; Zhu, H.; Huang, X.; Bai, X.; Kannan, K.; Zhang, T. Concentrations of bisphenol A and its alternatives in paired maternal-fetal urine, serum and amniotic fluid from an e-waste dismantling area in China. Environ. Int. 2020, 136, 105407. [Google Scholar] [CrossRef] [PubMed]
- Iliadi, A.; Koletsi, D.; Papageorgiou, S.N.; Eliades, T. Safety Considerations for Thermoplastic-Type Appliances Used as Orthodontic Aligners or Retainers. A Systematic Review and Meta-Analysis of Clinical and In-Vitro Research. Materials 2020, 13, 1843. [Google Scholar] [CrossRef] [PubMed]
- The Jamovi Project. Jamovi, version 2.3. 2023. Available online: https://www.jamovi.org (accessed on 30 June 2023).
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rat Strain | Total Animals (Control, Bisphenols, Bisphenol Plus Antioxidants) | Weight (g) | Initial Age of Exposure (Weeks) | Antioxidant Compound | Age at Sample Collection (Weeks) | Reference |
|---|---|---|---|---|---|---|
| Sprague Dawley | 42 (6, 6, 6) | 150–200 | 8 | mel | 16–17 | Akarca-Dizakar (2020) [24] |
| Wistar | 42 (6, 6, 6) | 150–170 | 6 | VitC | 13 | Aydoğan (2010) [33] |
| Wistar | 12 (4, 4, 4) | ~150–200 | 7–8 | VitC | 15–16 | Chitra (2003) [4] |
| Wistar | 28 (7, 7, 7) | 350–400 | ~11–12 | VitB9 | ~14–15 | Gules (2019) [32] |
| Sprague Dawley | 42 (6, 6, 6) | 190–200 | ~8 | VitE | ~14 | Jiang (2016) [41] |
| Wistar | 24 (6, 6, 6) | 231 ± 10 | 8 | VitE | 16 | Malmir (2021) [42] |
| Wistar | 40 (10, 10, 10) | 240 ± 10 | 16 | mel | 18 | Olukole (2018) [37] |
| Albino | 30 (6, 6, 6) | 150–200 | 7–8 | VitE | 15–16 | Omran (2017) [43] |
| Sprague Dawley | 56 (6, 16, 16) | 200–220 | 8 | mel | 11–14 | Othman (2016) [36] |
| Sprague Dawley | 42 (7, 7, 7) | 200–220 | 8 | mel VitE | 11 | Rashad (2021) [44] |
| Wistar | 24 (6, 6, 6) | 231 ± 10 | 8 | VitE | 16 | Samira (2020) [40] |
| Wistar | 70 (10, 10, 10) | 150–200 | 12 | VitE | 24 | Srivastava (2018) [31] |
| Sprague Dawley | 40 (10, 10, 10) | 200–220 | 8 | mel | 9 | Wu (2013) [39] |
| Bisphenol | Antioxidant Compound | Main Outcomes | Reference | |||||
|---|---|---|---|---|---|---|---|---|
| BP | Via | Dose * | SE | Via | Dose * | SE | ||
| Studies that used melatonin as antioxidant compound | ||||||||
| A | p.o. | 25 | 60 | p.o. | 20 | 60 | BPA: Sperm motility and viability vs. control. Sperm motility and viability vs. control.BPA + mel:  Sperm motility and viability vs. BPA group. Sperm motility and viability vs. BPA group. | Akarca-Dizakar (2020) [24] |
| A | p.o. | 10 | 14 | i.p. | 10 | 14 | BPA: E2 T vs. control, pathological lesions in the prostate. BPA + mel: E2 T vs. BPA group. Protector against BPA-induced toxicity in the prostate. | Olukole (2018) [37] |
| A | p.o. | 50 | 21 and 42 | p.o. | 10 | 21 and 42 | BPA: Sperm concentration and motility, T, SOD, and CAT vs. control group. Sperm abnormalities vs. control. BPA + mel: Sperm concentration and motility, T, vs. BPA group. | Othman (2016) [36] |
| A | p.o. | 200 | 10 | p.o. | 10 | 10 | BPA: SOD, TBARS, sperm DNA damage vs. control group. BPA + mel: SOD, TBARS, and sperm DNA damage vs. BPA group. | Wu (2013) [39] |
| Studies that used vitamins as antioxidant compound | ||||||||
| A | p.o. | 25 | 50 1 | p.o. | 60 | 50 1 | BPA, NP, OP: No alterations in testicles and epididymis absolute weight vs. control group. pathological lesions in testicles vs. the control group. BPA + VitC, NP + VitC, OP + VitC: Histopathological lesions in testicles vs. control group. Relative epididymis weight | Aydoğan (2009) [33] |
| NP | p.o. | 25 | p.o. | 60 | ||||
| OP | p.o. | 25 | p.o. | 60 | ||||
| A | p.o. | 0.0002, 0.002, 0.02 | 60 | p.o. | 40 | 60 | BPA: Epididymal sperm motility and concentration, antioxidant enzymes, equal sperm viability compared to control group. BPA + VitC: Epididymal sperm motility and concentration, antioxidant enzymes vs. BPA group. | Chitra (2003) [4] |
| A | p.o. | 50 | 14 | p.o. | 20 | 14 | BPA: Seminiferous tubule height and T. TUNEL-positive cells per seminiferous tubule. BPA + VitB9: Seminiferous tubule height, T and TUNEL-positive cells equal to control. | Gules (2019) [32] |
| A | p.o. | 200 | 42 | p.o. | 300 | 42 | BPA: Sperm concentration, semen production, T, LH vs control group. BPA + VitE: No differences in sperm variables, abnormal tail, and FSH vs. control and BPA groups. | Jiang (2016) [41] |
| A | p.o. | 250 | 56 | p.o. | 150 | 56 | BPA: TUNEL-positive spermatocytes, spermatids, and Sertoli cells vs. control group. T, motility, concentration, viability vs. control group. Body weight is equal to the control group. BPA + VitE: TUNEL-positive spermatic cells, body weight, and T similar to control group. | Malmir (2021) [42] |
| A | p.o. | 325 | 56 | p.o. | 200 | 56 | BPA: T, sloughing of the germinal epithelium vs. control. BPA + VitE: T, Spermatogonia number, and mild sloughing in germinal epithelia vs. BPA group. Disruption of germinal epithelium. | Omran (2017) [43] |
| A | i.p. | 50 | 21 1 | p.o. | VitE (100) | 21 1 | BPA: Abnormal sperm morphology vs control. Sperm concentration and motility, T vs. control group. BPA + VitE o BPA + mel: Abnormal spermatozoa vs. BPA group. Sperm concentration and motility, T vs. BPA group. | Rashad (2021) [44] |
| i.p. | mel (10) | |||||||
| A | p.o. | 250 | 56 | p.o. | 150 | 56 | BPA: Seminiferous tubule diameter, germinal epithelium thickness, number of spermatocytes, spermatids, Sertoli cells, sperm concentration, and T vs. control group. BPA + VitE: equal T vs. control group. seminiferous tubule diameter, germinal epithelium thickness, spermatocyte, sperm concentration vs. BPA group. | Samira (2020) [40] |
| A | p.o. | 0.05, 0.5, 1 | 90 | p.o. | 40 | 90 | BPA (high dose): Epididymis apoptosis, T vs. control group. BPA + VitE: Testis weight vs. BPA group. | Srivastava (2018) [31] |
decrease; increase; 1 three times each week.| Analysis | SMD | 95% CI | Heterogeneity | p | Begg Test | Egger Test | No. Studies (No. Comparisons) | ||
|---|---|---|---|---|---|---|---|---|---|
| I2 (%) | p Het | Q | |||||||
| Final Body Weight | |||||||||
| BP vs. Ctrl 2 | −0.65 | −2.38, 1.07 | 89.35 | <0.001 | 22.5 | 0.380 | 0.817 | <0.001 | 3(5) |
| BP + vit vs. BP | −0.63 | −2.68, 1.42 | 91.1 | <0.001 | 37.4 | 0.547 | 0.233 | 0.430 | 3(5) |
| BP + vit vs. Ctrl | −1.07 | −1.61, −0.53 | 0.00 | 0.436 | 3.8 | <0.001 * | 0.083 | 0.105 | 3(5) |
| Testosterone serum concentration | |||||||||
| BP vs. Ctrl 1 | −9.37 | −12.66, −6.08 | 77.56 | 0.002 | 12.5 | <0.001 * | 0.083 | <0.001 | 3(4) |
| BP + mel vs. BP | 6.34 | 3.92, 8.77 | 82.87 | <0.001 | 25.1 | < 0.001 * | 1.000 | 0.007 | 3(4) |
| BP + mel vs. Ctrl | −2.53 | −5.65, 0.60 | 95.78 | < 0.001 | 39.8 | 0.113 | 0.083 | <0.001 | 3(4) |
| BP vs. Ctrl 2 | −3.68 | −6.10, −1.26 | 95.73 | <0.001 | 56.5 | 0.003 * | 0.006 | < 0.001 | 6(8) |
| BP + vit vs. BP | 1.61 | 0.32, 2.91 | 89.72 | <0.001 | 31.5 | 0.015 * | 0.002 | <0 .001 | 6(8) |
| BP + vit vs. Ctrl | −2.08 | −3.83, −0.32 | 93.74 | <0.001 | 46.9 | 0.021 * | 0.075 | < 0.001 | 6(8) |
| Sperm concentration | |||||||||
| BP vs. Ctrl 1 | −3.82 | −7.16, −0.47 | 95.32 | < 0.001 | 46.2 | 0.025 * | 0.017 | <0.001 | 4(5) |
| BP + mel vs. BP | 2.17 | 0.05, 4.30, | 91.68 | < 0.001 | 32.1 | 0.045 * | 0.017 | <0.001 | 4(5) |
| BP + mel vs. Ctrl | −0.77 | −2.71, 1.16 | 92.04 | <0.001 | 27.5 | 0.434 | 0.483 | <0.200 | 4(5) |
| BP vs. Ctrl 2 | −2.10 | −3.01, −1.19 | 83.18 | <0.001 | 54.3 | 0.001 * | 0.006 | <0.001 | 6(14) |
| BP + vit vs. BP | 1.76 | 0.98, 2.53 | 79.15 | <0.001 | 50.0 | <0.001 * | < 0.001 | <0.001 | 6(14) |
| BP + vit vs. Ctrl | −1.23 | −1.86, −0.60 | 71.38 | <0.001 | 42.2 | <0.001 * | 0.101 | 0.045 | 6(14) |
| Sperm motility | |||||||||
| BP vs. Ctrl 1 | −10.85 | −18.78, −2.93 | 95.47 | <0.001 | 82.1 | 0.007 * | 0.083 | <0.001 | 3(4) |
| BP + mel vs. BP | 6.11 | −0.72, 12.94 | 98.69 | <0.001 | 50.1 | 0.079 | 0.750 | <0.001 | 3(4) |
| BP + mel vs. Ctrl | 0.46 | −3.71, 4.63 | 97.10 | <0.001 | 53.1 | 0.827 | 1.000 | 0.751 | 3(4) |
| BP vs. Ctrl 2 | −11.10 | −13.71, −8.49 | 0.00 | 0.583 | 1.95 | <0.001 * | 0.083 | 0.170 | 2(4) |
| BP + vit vs. BP | 13.42 | 5.32, 21.52 | 84.51 | <0.001 | 22.5 | 0.001 | 0.083 | <0.001 | 2(4) |
| BP + vit vs. Ctrl | −1.23 | −3.19, 0.73 | 85.08 | 0.001 | 15.6 | 0.218 | 0.083 | <0.001 | 2(4) |
| Sperm viability | |||||||||
| BP vs. Ctrl 1 | −2.58 | −4.29, −0.88 | 79.95 | 0.005 | 10.5 | 0.003 * | 0.333 | 0.001 | 2(3) |
| BP + mel vs. BP | 5.27 | 1.03, 9.50 | 94.73 | <0.001 | 46.0 | <0.001 * | 0.333 | <0.001 | 2(3) |
| BP + mel vs. Ctrl | −2.58 | −4.29, −0.88 | 79.95 | 0.005 | 10.5 | 0.003 * | 0.333 | 0.001 | 2(3) |
| BP vs. Ctrl 2 | −2.79 | −6.23, 0.66 | 93.49 | <0.001 | 19.3 | 0.113 | 0.083 | <0.001 | 2(4) |
| BP + vit vs. BP | 1.83 | −0.42, 4.09 | 87.24 | 0.001 | 16.1 | 0.111 | 0.083 | <0.001 | 2(4) |
| BP + vit vs. Ctrl | −0.93 | −1.75, −0.11 | 29.08 | 0.254 | 4.1 | 0.026 * | 1.000 | 0.201 | 2(4) |
| Abnormal sperm morphology | |||||||||
| BP vs. Ctrl 2 | 4.99 | 1.92, 8.06 | 91.21 | <0.001 | 30.1 | 0.001 * | 0.017 | <0.001 | 3(5) |
| BP + vit vs. BP | −1.43 | −4.26, 1.40 | 94.43 | <0.001 | 47.4 | 0.321 | 0.483 | 0.166 | 3(5) |
| BP + vit vs. Ctrl | 5.06 | 2.75, 7.36 | 81.86 | <0.001 | 21.3 | <0.001 * | 0.017 | <0.001 | 3(5) |
| Abnormal sperm head morphology | |||||||||
| BP vs. Ctrl 2 | 1.55 | −0.73, 3.83 | 90.7 | <0.001 | 26.2 | 0.184 | 0.333 | <0.001 | 2(4) |
| BP + vit vs. BP | 0.63 | −3.68, 4.93 | 95.9 | <0.001 | 50.8 | 0.775 | 1.000 | 0.736 | 2(4) |
| BP + vit vs. Ctrl | 2.17 | 0.17, 4.18 | 85.58 | <0.001 | 26.6 | 0.034 * | 0.083 | <0.001 | 2(4) |
| Abnormal sperm neck morphology | |||||||||
| BP vs. Ctrl 2 | 3.50 | −0.30, 7.30 | 95.64 | <0.001 | 17.9 | 0.071 | 0.083 | <0.001 | 2(4) |
| BP + vit vs. BP | −1.31 | −3.89, 1.28 | 92.61 | <0.001 | 29.4 | 0.321 | 0.750 | 0.066 | 2(4) |
| BP + vit vs. Ctrl | 2.68 | 0.40, 4.96 | 88.35 | <0.001 | 19.8 | 0.021 * | 0.083 | <0.001 | 2(4) |
| Abnormal sperm tail morphology | |||||||||
| BP vs. Ctrl 2 | 2.49 | −0.08, 5.06 | 91.73 | <0.001 | 18.5 | 0.058 | 0.083 | <0.001 | 2(4) |
| BP + vit vs. BP | −0.15 | −1.09, 0.78 | 61.37 | 0.054 | 7.6 | 0.746 | 0.750 | 0.171 | 2(4) |
| BP + vit vs. Ctrl | 3.64 | −0.11, 7.39 | 94.35 | <0.001 | 36.1 | 0.057 | 0.083 | <0.001 | 2(4) |
| Total testis weight | |||||||||
| BP vs. Ctrl 2 | −1.73 | −2.87, −0.59 | 79.34 | <0.001 | 18.9 | 0.003 * | 0.233 | 0.104 | 3(5) |
| BP + vit vs. BP | 1.42 | 0.17, 2.67 | 83.46 | <0.001 | 23.8 | 0.026 * | 0.817 | 0.428 | 3(5) |
| BP + vit vs. Ctrl | −0.70 | −1.14, −0.26 | 0.00 | 0.803 | 1.6 | 0.002 * | 0.483 | 0.284 | 3(5) |
| Right testis weight | |||||||||
| BP vs. Ctrl 2 | −0.81 | −1.84, 0.21 | 65.24 | 0.039 | 8.39 | 0.119 | 0.083 | 0.005 | 2(4) |
| BP + vit vs. BP | −0.13 | −1.01, 0.76 | 57.43 | 0.071 | 7.03 | 0.778 | 0.333 | 0.555 | 2(4) |
| BP + vit vs. Ctrl | −1.57 | −2.50, −0.63 | 50.80 | 0.106 | 6.13 | <0.001 * | 0.083 | 0.013 | 2(4) |
| Left testis weight | |||||||||
| BP vs. Ctrl 2 | −0.49 | −1.44, 0.45 | 62.22 | 0.054 | 7.64 | 0.304 | 0.333 | 0.012 | 2(4) |
| BP + vit vs. BP | −0.19 | −0.97, 0.59 | 46.31 | 0.133 | 5.60 | 0.634 | 0.750 | 0.609 | 2(4) |
| BP + vit vs. Ctrl | −0.79 | −1.37, −0.22 | 0.00 | 0.938 | 0.41 | 0.007 * | 0.083 | 0.524 | 2(4) |
| Receptor | Ligand | Amino Acids with Hydrogen Bonds | Amino Acids with Steric Interaction | Functional Groups and Attached Amino Acids | MolDock Score |
|---|---|---|---|---|---|
| Androgen | E2 | Arg 752, Thr 877, Gln 711 | Met 745 | OH (phenol) -> Gln 711, Arg 752, Met 745 OH (cyclopentane) -> Thr 877 | −117.344 |
| DHT | Arg752, Thr 877, Gln711 | Met 745, Phe 764 | OH (carbonyl) -> Gln 711, Arg 752 OH (cyclopentane) -> Thr 877 | −109.764 | |
| BPA | Arg 752, Gln 711, Asn 705, Phe 764 | - | OH (phenol) -> Arg 752, Gln 711, Phe 764, Asn 705 | −90.4377 | |
| BPS | Arg 752, Gln 711, Asn 705 | Thr 877 | OH (phenol) -> Gln 711, Arg 752, Asn 705 | −91.184 | |
| mel | Met 745 | Leu 704 | Pyrrole -> Met 745 | −114.947 | |
| VitE | Thr 877 | Met 787, Asn 705, Leu 880, Thr 877, Phe 876, Leu 873, Met 745, Phe 764, Leu 704, Met 895, Gly 708, Arg 752, Met 749, Gln 711, Val 746 | OH (phenol) -> Thr 877 Phenol -> Asn 705, Met 895, Leu 704 | −139.586 | |
| VitC | Arg 752, Gln 711, Leu 704, Met 745 | Gly 708, Met 749 | Carbonyl -> Gln 711, Arg 752, Phe 764 OH-3 -> Met 745, Met 749 OH-6/7-> Leu 704 | −97.1359 | |
| VitB9 | Leu 701, Thr 877, Asn 705, Leu 873 | Leu 873, Phe 876, Met 780, Leu 704, Gly 708, Met 742, Val 746, Met 749, Met 787, Met 745 | NH2 (cyclohexene): Met 787, Met 749 Carbonyl -> Met 742, Met 745 NH (cyclohexene): Val 746 Phenol: Leu 704 | −136.277 | |
| Estrogens (α chain) | E2 | Arg 394, His 524, Glu 353 | - | OH (phenol) -> Glu 353, Arg 394 OH (cyclopentane) -> His 524 | −102.505 |
| DHT | Arg 394, His 524, Gly 521 | Leu 387, Glu 353, Leu 525 | OH (cyclopentane) -> His 524, Gly 521 OH (carbonyle) -> Glu 353, Arg 394 | −101.016 | |
| BPA | Thr 347, Phe 404 | Arg 394, Glu 353, Leu 384 | OH (phenol) -> Phe 404, Glu 353, Thr 347 | −83.136 | |
| BPS | Phe 404 | Arg 394, Glu 353, Phe 404, Met 343, Thr 347, Leu 349, Leu 384, Met 384, Met 388 | OH (phenol) -> Leu 349, Phe 404 Sulfoxide -> Met 388, Leu 384 | −82.0316 | |
| mel | - | Leu 391, Glu 353 | Phenol-> Leu 346 | −100.315 | |
| VitE | Glu 353 | Asp 351, Leu 428, Met 388, Leu 387, Leu 391, Glu 353, Leu 346, Phe 404, Met 343, Leu 349, Leu 384, Leu 525, Gly 521, His 524, Ile 424 | OH (phenol) -> Leu 391, Glu 349 | −128.716 | |
| VitC | Arg 394, Glu 353, Leu 387 | Leu 346, Glu 353, Leu 391, Met 388, Phe 404 | Carbonyl -> Leu 346 OH-2: Glu 353, Phe 404 OH-3: Leu 387, Leu 391, Arg 394 OH-5: Met 388 | −72.1521 | |
| VitB9 | Leu 387, Glu 353, Asp 351 | Leu 387, Arg 394, Lys 529, Thr 347, Ala 350, Trp 383 | NH2 (cyclohexene): Leu 387, Glu 353, Arg 394, Carbonyl -> Lys 529, Thr 347, Trp 383 | −136,387 | |
| Estrogens (β chain) | E2 | Arg 346, Glu 260 | Leu 343 | OH (phenol) -> Glu 305, Arg 346 | −100.014 |
| DHT | Arg 346 | Leu 343, Leu339, Gly472, Glu 305, Leu 476 | Carbonyl -> Arg 346, Glu 305 | −111.186 | |
| BPA | Arg 346, Glu 305 | Phe 356, Met 336 | OH (phenol) -> Glu 305, Arg 346 | −92.5984 | |
| BPS | Arg 346, Glu 305 | Phe 356, Met 336, Leu 339 | OH (phenol) -> Glu 305, Arg 346 O (sulfoxide) -> Leu 339, Met 336 | −80.4489 | |
| mel | Glu 305, Leu 298 | Leu 343 | N (secondary amine) -> Leu 298 O (Ether) -> Leu 343 N (pyrrole) -> Glu 305 | −105.784 | |
| VitE | Leu 398 | Ile 373, Met 340, Leu 298 | OH (phenol) -> Leu 398 | −148.873 | |
| VitC | Arg 346, Leu 298, Leu 339 | Leu 343, Phe 356, Ala 302 | Carbonyl -> Arg 346, Phe 356 OH-3: Leu 339, Leu 343 OH -> 6/7: Leu 298 | −98.4997 | |
| VitB9 | Asp 303, Pro 486, Glu 305 | Arg 346, Ala 302, Trp 335, Leu 306, Leu 476, Pro 486 | NH2 -> Glu 305 OH -> Pro 486, Asp 303 | −150.355 |
| Receptor | Coordinates | Radio Å | PDB Code |
|---|---|---|---|
| Androgen | X: 0.41, Y: 31.66, Z: 4.44 | 10 | 1E3G |
| Estrogens (α chain) | X:90.42, Y: 13.12, Z: 72.44, | 10 | 1NDE |
| Estrogens (β chain) | X: 109.00, Y: 8.82, Z: −108.68 | 10 | 1A52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña-Corona, S.I.; Chávez-Corona, J.I.; Pérez-Caltzontzin, L.E.; Vargas-Estrada, D.; Mendoza-Rodríguez, C.A.; Ramos-Martínez, E.; Cerbón-Gutiérrez, J.L.; Herrera-Barragán, J.A.; Quintanar-Guerrero, D.; Leyva-Gómez, G. Melatonin and Vitamins as Protectors against the Reproductive Toxicity of Bisphenols: Which Is the Most Effective? A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2023, 24, 14930. https://doi.org/10.3390/ijms241914930
Peña-Corona SI, Chávez-Corona JI, Pérez-Caltzontzin LE, Vargas-Estrada D, Mendoza-Rodríguez CA, Ramos-Martínez E, Cerbón-Gutiérrez JL, Herrera-Barragán JA, Quintanar-Guerrero D, Leyva-Gómez G. Melatonin and Vitamins as Protectors against the Reproductive Toxicity of Bisphenols: Which Is the Most Effective? A Systematic Review and Meta-Analysis. International Journal of Molecular Sciences. 2023; 24(19):14930. https://doi.org/10.3390/ijms241914930
Chicago/Turabian StylePeña-Corona, Sheila I., Juan I. Chávez-Corona, Luis E. Pérez-Caltzontzin, Dinorah Vargas-Estrada, C. Adriana Mendoza-Rodríguez, Edgar Ramos-Martínez, Jose L. Cerbón-Gutiérrez, José A. Herrera-Barragán, David Quintanar-Guerrero, and Gerardo Leyva-Gómez. 2023. "Melatonin and Vitamins as Protectors against the Reproductive Toxicity of Bisphenols: Which Is the Most Effective? A Systematic Review and Meta-Analysis" International Journal of Molecular Sciences 24, no. 19: 14930. https://doi.org/10.3390/ijms241914930
APA StylePeña-Corona, S. I., Chávez-Corona, J. I., Pérez-Caltzontzin, L. E., Vargas-Estrada, D., Mendoza-Rodríguez, C. A., Ramos-Martínez, E., Cerbón-Gutiérrez, J. L., Herrera-Barragán, J. A., Quintanar-Guerrero, D., & Leyva-Gómez, G. (2023). Melatonin and Vitamins as Protectors against the Reproductive Toxicity of Bisphenols: Which Is the Most Effective? A Systematic Review and Meta-Analysis. International Journal of Molecular Sciences, 24(19), 14930. https://doi.org/10.3390/ijms241914930