
Transcriptional Dysregulation and Impaired Neuronal Activity in FMR1 Knock-Out and Fragile X Patients’ iPSC-Derived Models
 , , , , , , add
Show full author list
, , , , , , add
Show full author list
Abstract
:1. Introduction
2. Results
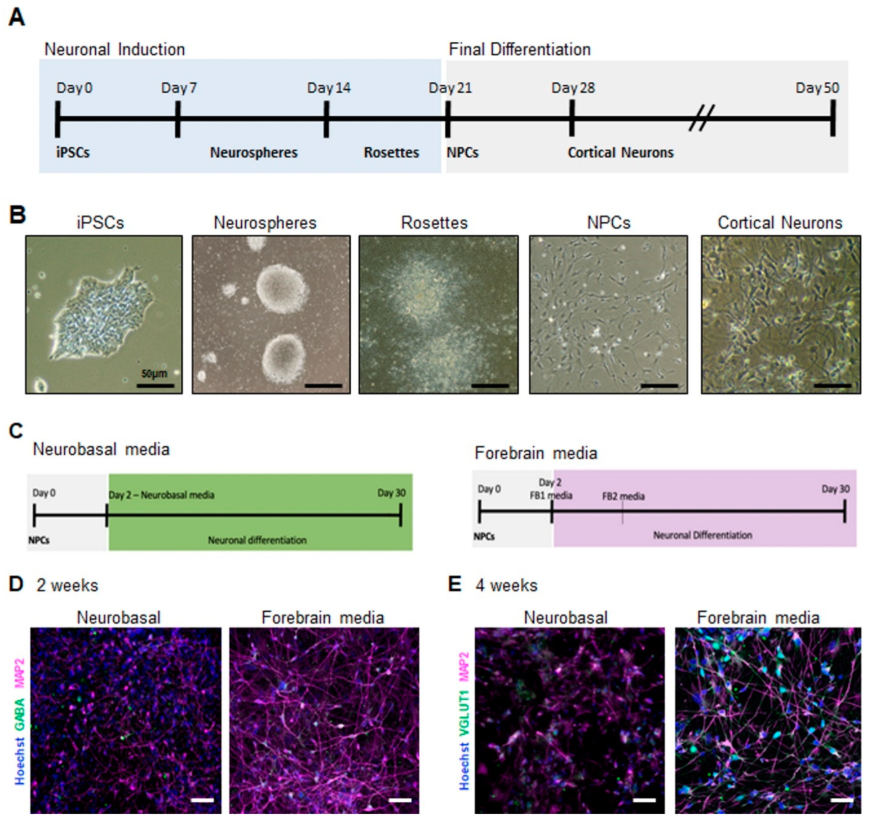
2.1. Generation of Electrically Active iPSC-Derived Cortical Neurons
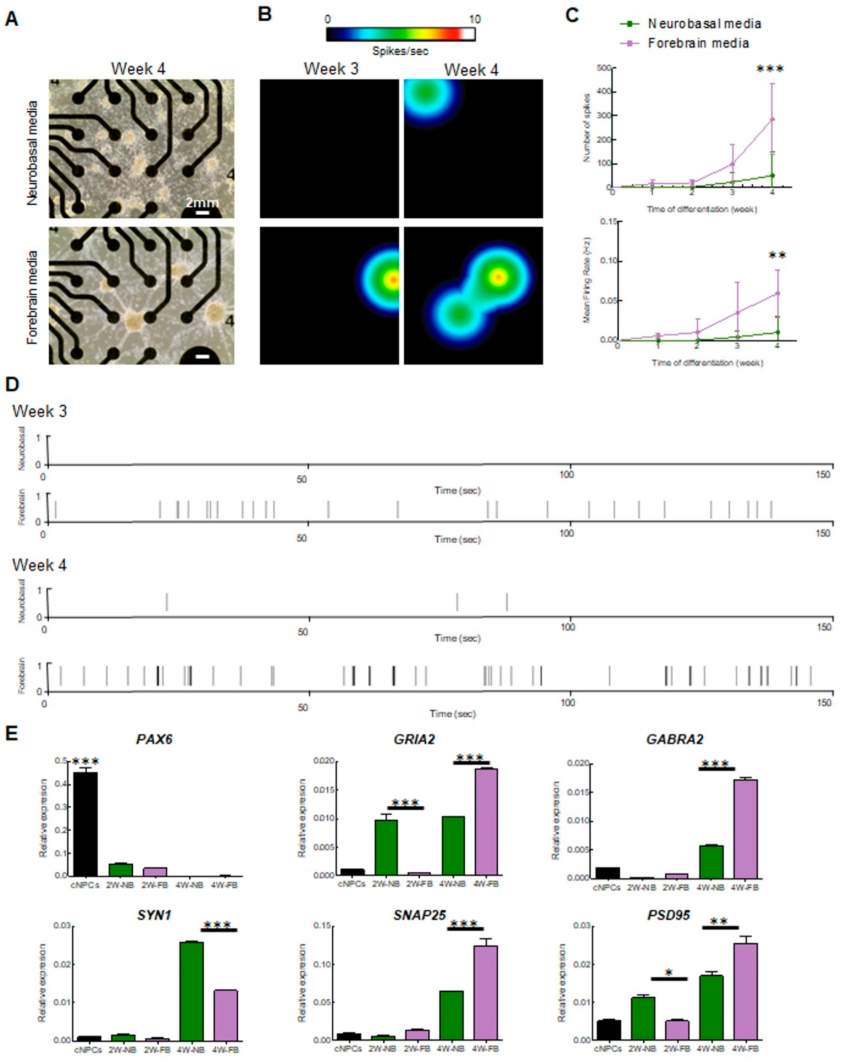
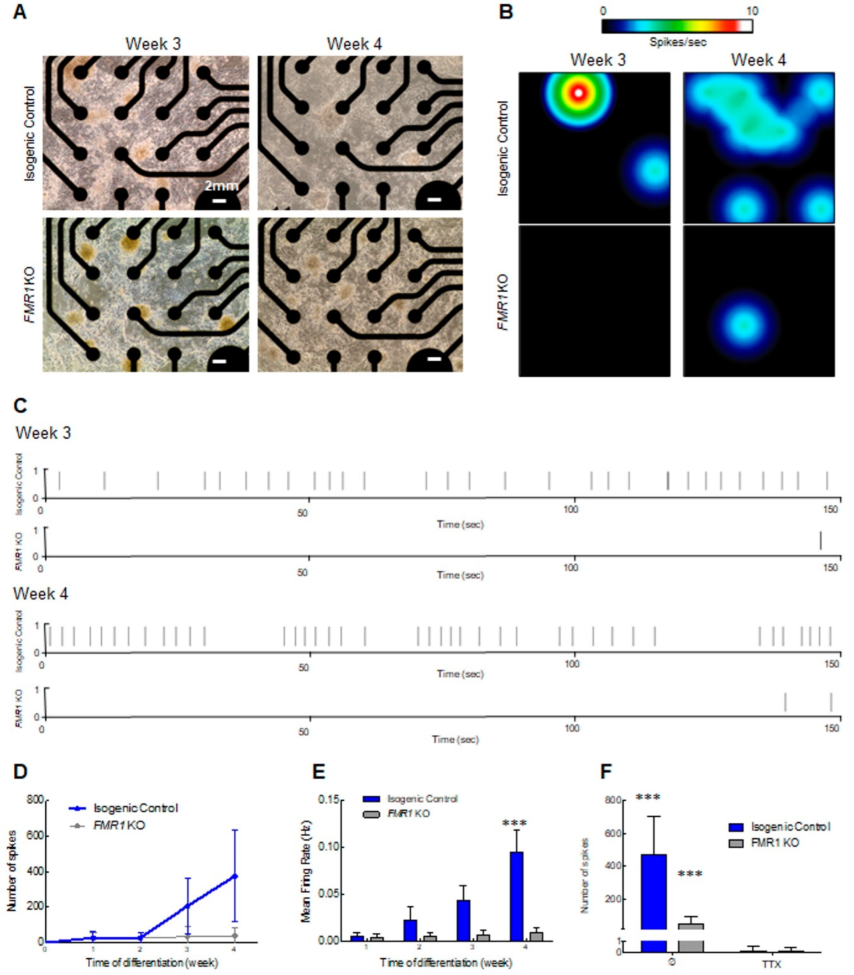
2.2. Multielectrode Array (MEA) Recording and Analysis of Electrical Activity of iPSC-Derived Cortical Neurons
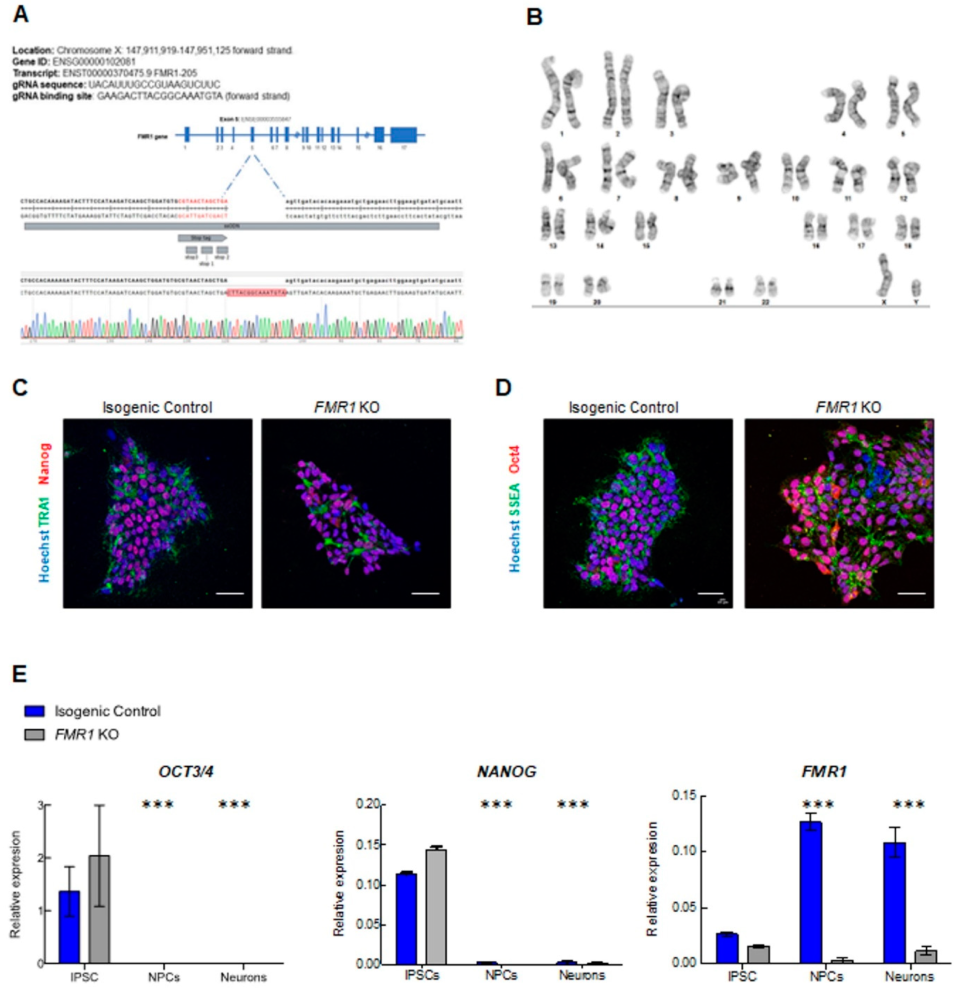
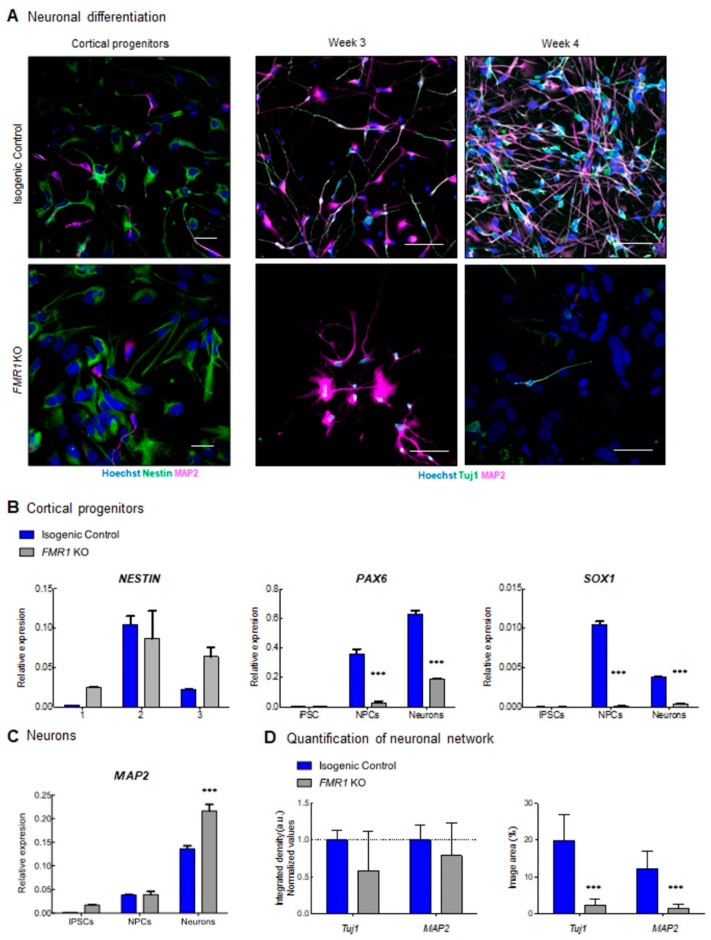
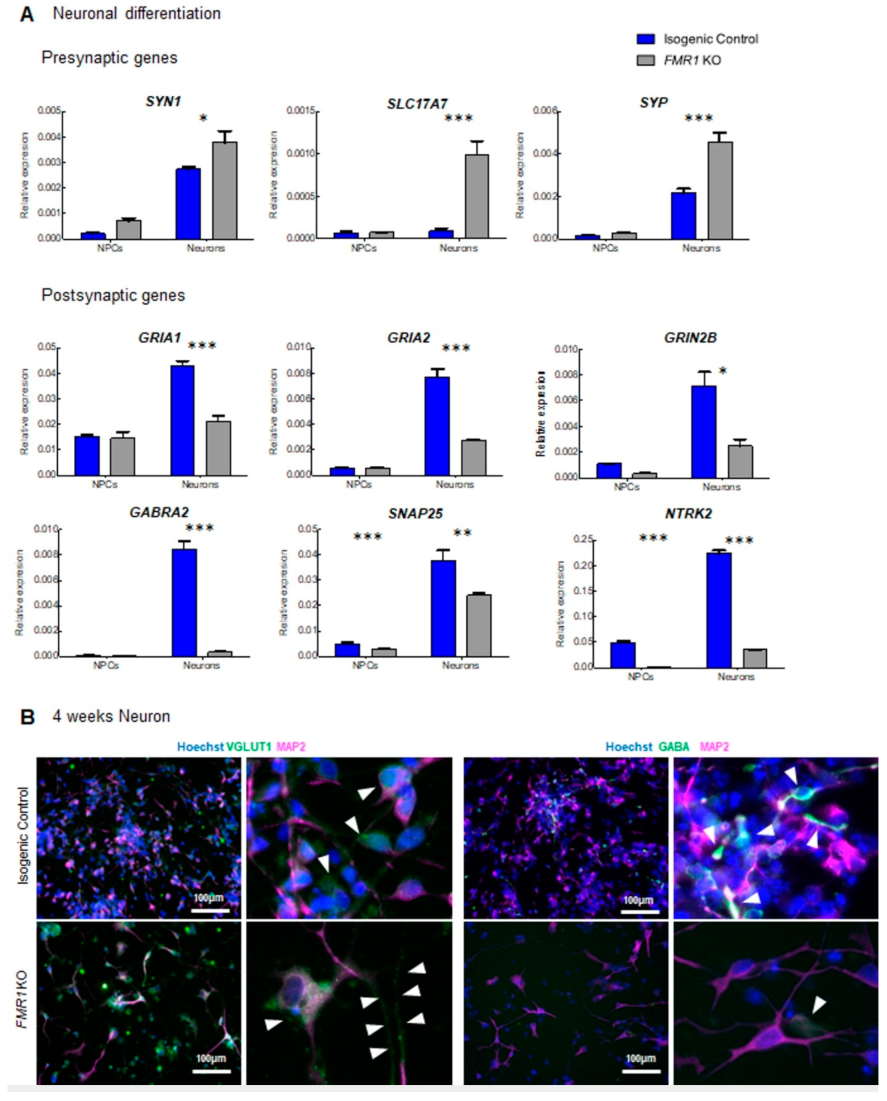
2.3. Characterization of Neuronal Differentiation with FMR1 KO iPSC
2.4. Neuronal Spontaneous Electrical Activity Impairment in the FMR1 KO
2.5. Whole Transcriptome Profiling of FMR1 KO iPSC-Derived NPCs and Neurons
2.6. Differential Expression of Synaptic Markers in the FMR1 KO
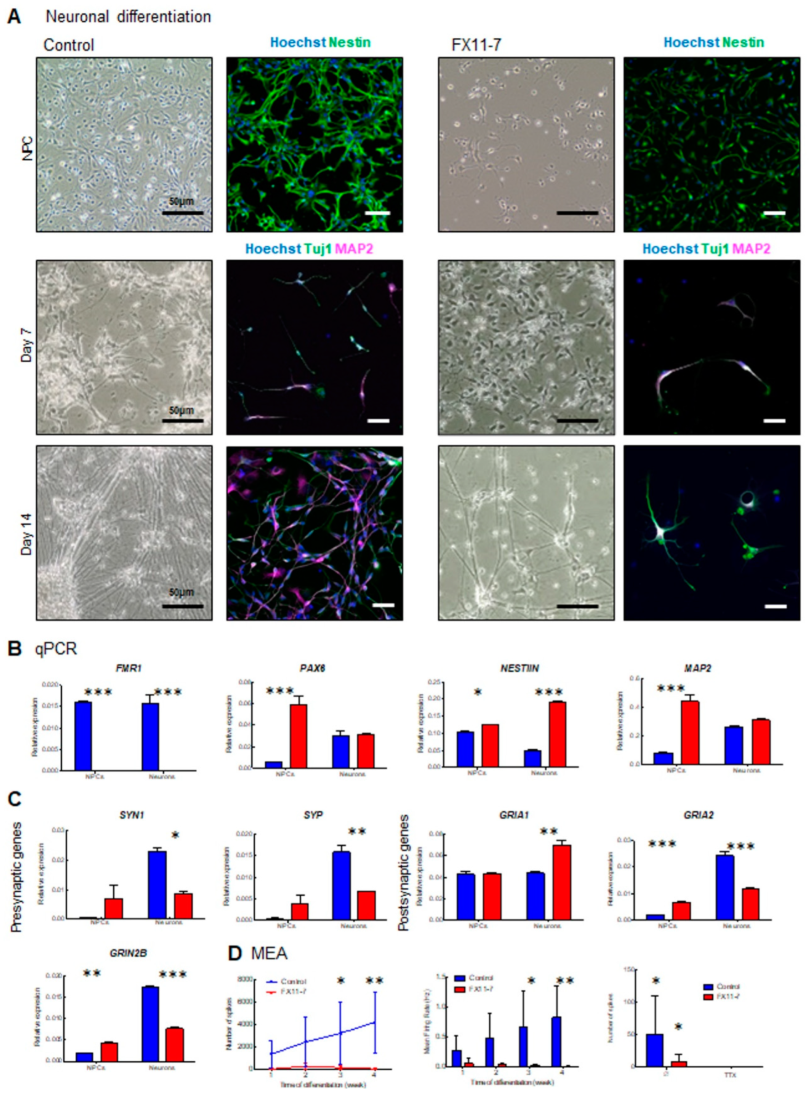
2.7. Fragile X Syndrome Patient Cell Line Displays Impaired Neuronal Development and Activity
3. Materials and Methods
3.1. Cell Line Description
3.2. Cell Culture and Cortical Neuron Differentiation
3.3. CRISPR Genome Editing
3.4. Immunofluorescence Staining and Image Acquisition
3.5. Multielectrode Array (MEA) Analysis
3.6. Gene Expression Analyses
RNA Extraction, cDNA Synthesis, and Quantitative PCR
3.7. RNA Sequencing
3.7.1. Sample Preparation and Sequencing
3.7.2. Data Processing, Alignment, and Differential Analysis
3.7.3. Functional Annotation
3.7.4. Enrichment Analyses
4. Discussion
4.1. FMRP, a Multifunction Protein
4.2. Delayed Transition from Progenitor-like State towards a Neuron and Impaired Differentiation
4.3. Neuronal Differentiation and Electrical Activity Impairment
4.4. Impaired Transcriptome and Cellular Activity: Common Traits between FXS and Other Neurodevelopmental Disorders
4.5. Advantages of the iPSC Model and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abrahams, B.S.; Geschwind, D.H. Advances in autism genetics: On the threshold of a new neurobiology. Nat. Rev. Genet. 2008, 9, 341–355. [Google Scholar] [CrossRef]
- Verkerk, A.J.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.H.; Kuhl, D.P.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.P.; et al. Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell 1991, 65, 905–914. [Google Scholar] [CrossRef]
- Davis, J.K.; Broadie, K. Multifarious Functions of the Fragile X Mental Retardation Protein. Trends Genet. 2017, 33, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Khandjian, E.W.; Robert, C.; Davidovic, L. FMRP, a multifunctional RNA-binding protein in quest of a new identity. Front. Genet. 2022, 13, 976480. [Google Scholar] [CrossRef]
- Richter, J.D.; Zhao, X. The molecular biology of FMRP: New insights into fragile X syndrome. Nat. Rev. Neurosci. 2021, 22, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Penagarikano, O.; Mulle, J.G.; Warren, S.T. The pathophysiology of fragile x syndrome. Annu. Rev. Genom. Hum. Genet. 2007, 8, 109–129. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, G.B.; Mendelsohn, N.J. Genetics evaluation for the etiologic diagnosis of autism spectrum disorders. Genet. Med. 2008, 10, 4–12. [Google Scholar] [CrossRef]
- Wang, L.W.; Berry-Kravis, E.; Hagerman, R.J. Fragile X: Leading the way for targeted treatments in autism. Neurotherapeutics 2010, 7, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Maussion, G.; Rocha, C.; Bernard, G.; Beitel, L.K.; Durcan, T.M. Patient-Derived Stem Cells, Another in vitro Model, or the Missing Link toward Novel Therapies for Autism Spectrum Disorders? Front. Pediatr. 2019, 7, 225. [Google Scholar] [CrossRef]
- Maussion, G.; Rocha, C.; Pimentel, L.; Beitel, L.K.; Durcan, T.M. Chapter 3—Human induced pluripotent stem cell-based studies; a new route toward modeling autism spectrum disorders. In iPSCs for Modeling Central Nervous System Disorders; Birbrair, A., Ed.; Academic Press: Cambridge, MA, USA, 2021; Volume 6, pp. 37–81. [Google Scholar]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Ardhanareeswaran, K.; Mariani, J.; Coppola, G.; Abyzov, A.; Vaccarino, F.M. Human induced pluripotent stem cells for modelling neurodevelopmental disorders. Nat. Rev. Neurol. 2017, 13, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.S.; Gigek, C.O.; Rosenfeld, J.A.; Diallo, A.B.; Maussion, G.; Chen, G.G.; Vaillancourt, K.; Lopez, J.P.; Crapper, L.; Poujol, R.; et al. Molecular convergence of neurodevelopmental disorders. Am. J. Hum. Genet. 2014, 95, 490–508. [Google Scholar] [CrossRef] [PubMed]
- Gigek, C.O.; Chen, E.S.; Ota, V.K.; Maussion, G.; Peng, H.; Vaillancourt, K.; Diallo, A.B.; Lopez, J.P.; Crapper, L.; Vasuta, C.; et al. A molecular model for neurodevelopmental disorders. Transl. Psychiatry 2015, 5, e565. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, R.; Qiao, N.; Peng, G.; Zhang, K.; Tang, K.; Han, J.J.; Jing, N. Transcriptome analysis reveals determinant stages controlling human embryonic stem cell commitment to neuronal cells. J. Biol. Chem. 2017, 292, 19590–19604. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.; Maussion, G.; Jefri, M.; Peng, H.; Theroux, J.F.; Silveira, H.; Soubannier, V.; Wu, H.; Hu, P.; Galat, E.; et al. Disruption of GRIN2B Impairs Differentiation in Human Neurons. Stem Cell Rep. 2018, 11, 183–196. [Google Scholar] [CrossRef]
- Mor-Shaked, H.; Eiges, R. Modeling Fragile X Syndrome Using Human Pluripotent Stem Cells. Genes 2016, 7, 77. [Google Scholar] [CrossRef]
- Volpato, V.; Webber, C. Addressing variability in iPSC-derived models of human disease: Guidelines to promote reproducibility. Dis. Models Mech. 2020, 13, dmm042317. [Google Scholar] [CrossRef]
- Autar, K.; Guo, X.; Rumsey, J.W.; Long, C.J.; Akanda, N.; Jackson, M.; Narasimhan, N.S.; Caneus, J.; Morgan, D.; Hickman, J.J. A functional hiPSC-cortical neuron differentiation and maturation model and its application to neurological disorders. Stem Cell Rep. 2022, 17, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.X.; Abdian, N.; Maussion, G.; Thomas, R.A.; Demirova, I.; Cai, E.; Tabatabaei, M.; Beitel, L.K.; Karamchandani, J.; Fon, E.A.; et al. A Multistep Workflow to Evaluate Newly Generated iPSCs and Their Ability to Generate Different Cell Types. Methods Protoc. 2021, 4, 50. [Google Scholar] [CrossRef]
- Bell, S.; Peng, H.; Crapper, L.; Kolobova, I.; Maussion, G.; Vasuta, C.; Yerko, V.; Wong, T.P.; Ernst, C. A Rapid Pipeline to Model Rare Neurodevelopmental Disorders with Simultaneous CRISPR/Cas9 Gene Editing. Stem Cells Transl. Med. 2017, 6, 886–896. [Google Scholar] [CrossRef]
- Bardy, C.; van den Hurk, M.; Eames, T.; Marchand, C.; Hernandez, R.V.; Kellogg, M.; Gorris, M.; Galet, B.; Palomares, V.; Brown, J.; et al. Neuronal medium that supports basic synaptic functions and activity of human neurons in vitro. Proc. Natl. Acad. Sci. USA 2015, 112, E2725–E2734. [Google Scholar] [CrossRef] [PubMed]
- Hyvarinen, T.; Hyysalo, A.; Kapucu, F.E.; Aarnos, L.; Vinogradov, A.; Eglen, S.J.; Yla-Outinen, L.; Narkilahti, S. Functional characterization of human pluripotent stem cell-derived cortical networks differentiated on laminin-521 substrate: Comparison to rat cortical cultures. Sci. Rep. 2019, 9, 17125. [Google Scholar] [CrossRef] [PubMed]
- Harrell, E.R.; Pimentel, D.; Miesenbock, G. Changes in Presynaptic Gene Expression during Homeostatic Compensation at a Central Synapse. J. Neurosci. 2021, 41, 3054–3067. [Google Scholar] [CrossRef] [PubMed]
- Kasteel, E.E.; Westerink, R.H. Comparison of the acute inhibitory effects of Tetrodotoxin (TTX) in rat and human neuronal networks for risk assessment purposes. Toxicol. Lett. 2017, 270, 12–16. [Google Scholar] [CrossRef]
- Basu, S.N.; Kollu, R.; Banerjee-Basu, S. AutDB: A gene reference resource for autism research. Nucleic Acids Res. 2009, 37, D832–D836. [Google Scholar] [CrossRef]
- Tran, S.S.; Jun, H.I.; Bahn, J.H.; Azghadi, A.; Ramaswami, G.; Van Nostrand, E.L.; Nguyen, T.B.; Hsiao, Y.E.; Lee, C.; Pratt, G.A.; et al. Widespread RNA editing dysregulation in brains from autistic individuals. Nat. Neurosci. 2019, 22, 25–36. [Google Scholar] [CrossRef]
- Pellegrini, R. Edit Single Bases with Benchling! 2016. Available online: https://www.benchling.com/blog/base-editor (accessed on 30 August 2023).
- Deneault, E.; Chaineau, M.; Nicouleau, M.; Castellanos Montiel, M.J.; Franco Flores, A.K.; Haghi, G.; Chen, C.X.; Abdian, N.; Shlaifer, I.; Beitel, L.K.; et al. A streamlined CRISPR workflow to introduce mutations and generate isogenic iPSCs for modeling amyotrophic lateral sclerosis. Methods 2022, 203, 297–310. [Google Scholar] [CrossRef]
- Nicouleau, M.; Pimentel, L.; Shlaifer, I.; Durcan, T.M. Generation of Knockout Cell Lines Using CRISPR-Cas9 and ddPCR Technology. 2020. Available online: https://doi.org/10.5281/zenodo.3875777 (accessed on 30 August 2023).
- Nicouleau, M.; Durcan, T.M. DNA sequencing with the SeqStudio. 2020. Available online: https://doi.org/10.5281/zenodo.3738330 (accessed on 30 August 2023).
- Maussion, G.; Thomas, R.A.; Demirova, I.; Gu, G.; Cai, E.; Chen, C.X.; Abdian, N.; Strauss, T.J.P.; Kelai, S.; Nauleau-Javaudin, A.; et al. Auto-qPCR; a python-based web app for automated and reproducible analysis of qPCR data. Sci. Rep. 2021, 11, 21293. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Grossmann, S.; Bauer, S.; Robinson, P.N.; Vingron, M. Improved detection of overrepresentation of Gene-Ontology annotations with parent child analysis. Bioinformatics 2007, 23, 3024–3031. [Google Scholar] [CrossRef]
- Weisz, E.D.; Monyak, R.E.; Jongens, T.A. Deciphering discord: How Drosophila research has enhanced our understanding of the importance of FMRP in different spatial and temporal contexts. Exp. Neurol. 2015, 274 Pt A, 14–24. [Google Scholar] [CrossRef]
- Raj, N.; McEachin, Z.T.; Harousseau, W.; Zhou, Y.; Zhang, F.; Merritt-Garza, M.E.; Taliaferro, J.M.; Kalinowska, M.; Marro, S.G.; Hales, C.M.; et al. Cell-type-specific profiling of human cellular models of fragile X syndrome reveal PI3K-dependent defects in translation and neurogenesis. Cell Rep. 2021, 35, 108991. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.; McCarty, V.; Peng, H.; Jefri, M.; Hettige, N.; Antonyan, L.; Crapper, L.; O’Leary, L.A.; Zhang, X.; Zhang, Y.; et al. Lesch-Nyhan disease causes impaired energy metabolism and reduced developmental potential in midbrain dopaminergic cells. Stem Cell Rep. 2021, 16, 1749–1762. [Google Scholar] [CrossRef]
- Sharma, A.; Hoeffer, C.A.; Takayasu, Y.; Miyawaki, T.; McBride, S.M.; Klann, E.; Zukin, R.S. Dysregulation of mTOR signaling in fragile X syndrome. J. Neurosci. 2010, 30, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Chao, O.Y.; Pathak, S.S.; Zhang, H.; Dunaway, N.; Li, J.S.; Mattern, C.; Nikolaus, S.; Huston, J.P.; Yang, Y.M. Altered dopaminergic pathways and therapeutic effects of intranasal dopamine in two distinct mouse models of autism. Mol. Brain 2020, 13, 111. [Google Scholar] [CrossRef]
- Kosillo, P.; Ahmed, K.M.; Aisenberg, E.E.; Karalis, V.; Roberts, B.M.; Cragg, S.J.; Bateup, H.S. Dopamine neuron morphology and output are differentially controlled by mTORC1 and mTORC2. eLife 2022, 11, e75398. [Google Scholar] [CrossRef] [PubMed]
- Doers, M.E.; Musser, M.T.; Nichol, R.; Berndt, E.R.; Baker, M.; Gomez, T.M.; Zhang, S.C.; Abbeduto, L.; Bhattacharyya, A. iPSC-derived forebrain neurons from FXS individuals show defects in initial neurite outgrowth. Stem Cells Dev. 2014, 23, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Marro, S.G.; Zhang, Y.; Arendt, K.L.; Patzke, C.; Zhou, B.; Fair, T.; Yang, N.; Sudhof, T.C.; Wernig, M.; et al. The fragile X mutation impairs homeostatic plasticity in human neurons by blocking synaptic retinoic acid signaling. Sci. Transl. Med. 2018, 10, eaar4338. [Google Scholar] [CrossRef]
- Gildin, L.; Rauti, R.; Vardi, O.; Kuznitsov-Yanovsky, L.; Maoz, B.M.; Segal, M.; Ben-Yosef, D. Impaired Functional Connectivity Underlies Fragile X Syndrome. Int. J. Mol. Sci. 2022, 23, 2048. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Wu, H.; Coronado, A.A.; de Laittre, E.; Osterweil, E.K.; Zhang, Y.; Bear, M.F. Negative Allosteric Modulation of mGluR5 Partially Corrects Pathophysiology in a Mouse Model of Rett Syndrome. J. Neurosci. 2016, 36, 11946–11958. [Google Scholar] [CrossRef]
- Brighi, C.; Salaris, F.; Soloperto, A.; Cordella, F.; Ghirga, S.; de Turris, V.; Rosito, M.; Porceddu, P.F.; D’Antoni, C.; Reggiani, A.; et al. Novel fragile X syndrome 2D and 3D brain models based on human isogenic FMRP-KO iPSCs. Cell Death Dis. 2021, 12, 498. [Google Scholar] [CrossRef]
- O’Roak, B.J.; Vives, L.; Fu, W.; Egertson, J.D.; Stanaway, I.B.; Phelps, I.G.; Carvill, G.; Kumar, A.; Lee, C.; Ankenman, K.; et al. Multiplex targeted sequencing identifies recurrently mutated genes in autism spectrum disorders. Science 2012, 338, 1619–1622. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, J.; Guo, H.; Ou, J.; Peng, Y.; Liu, Q.; Shen, Y.; Shi, L.; Liu, Y.; Xiong, Z.; et al. Association of genetic variants of GRIN2B with autism. Sci. Rep. 2015, 5, 8296. [Google Scholar] [CrossRef]
- Kang, Y.; Zhou, Y.; Li, Y.; Han, Y.; Xu, J.; Niu, W.; Li, Z.; Liu, S.; Feng, H.; Huang, W.; et al. A human forebrain organoid model of fragile X syndrome exhibits altered neurogenesis and highlights new treatment strategies. Nat. Neurosci. 2021, 24, 1377–1391. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.A.; Renner, M.; Martin, C.A.; Wenzel, D.; Bicknell, L.S.; Hurles, M.E.; Homfray, T.; Penninger, J.M.; Jackson, A.P.; Knoblich, J.A. Cerebral organoids model human brain development and microcephaly. Nature 2013, 501, 373–379. [Google Scholar] [CrossRef]
- Pasca, A.M.; Sloan, S.A.; Clarke, L.E.; Tian, Y.; Makinson, C.D.; Huber, N.; Kim, C.H.; Park, J.Y.; O’Rourke, N.A.; Nguyen, K.D.; et al. Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat. Methods 2015, 12, 671–678. [Google Scholar] [CrossRef]
- Hehr, U.; Pineda-Alvarez, D.E.; Uyanik, G.; Hu, P.; Zhou, N.; Hehr, A.; Schell-Apacik, C.; Altus, C.; Daumer-Haas, C.; Meiner, A.; et al. Heterozygous mutations in SIX3 and SHH are associated with schizencephaly and further expand the clinical spectrum of holoprosencephaly. Hum. Genet. 2010, 127, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.D.; Ramaswamy, V.; Andersen, J.; Bolduc, F.V. Atypical Rett syndrome with selective FOXG1 deletion detected by comparative genomic hybridization: Case report and review of literature. Eur. J. Hum. Genet. 2009, 17, 1577–15781. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Diagnostic | Cell of Origin | Sex | Ethnicity | Reprogramming | Cell Source | References |
|---|---|---|---|---|---|---|---|
| AIW002-02 | Healthy | PBMC | M | Caucasian | Episomal | The Neuro | PMID: 34287353 |
| FMR1-KO | N/A | AiW002-2 IPSC | M | Caucasian | N/A | The Neuro | |
| FX-11-7 | Fragile X Syndrome | Fibroblast | M | N/A | Lentivirus | WiCell | PMID: 24654675 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maussion, G.; Rocha, C.; Abdian, N.; Yang, D.; Turk, J.; Carrillo Valenzuela, D.; Pimentel, L.; You, Z.; Morquette, B.; Nicouleau, M.; et al. Transcriptional Dysregulation and Impaired Neuronal Activity in FMR1 Knock-Out and Fragile X Patients’ iPSC-Derived Models. Int. J. Mol. Sci. 2023, 24, 14926. https://doi.org/10.3390/ijms241914926
Maussion G, Rocha C, Abdian N, Yang D, Turk J, Carrillo Valenzuela D, Pimentel L, You Z, Morquette B, Nicouleau M, et al. Transcriptional Dysregulation and Impaired Neuronal Activity in FMR1 Knock-Out and Fragile X Patients’ iPSC-Derived Models. International Journal of Molecular Sciences. 2023; 24(19):14926. https://doi.org/10.3390/ijms241914926
Chicago/Turabian StyleMaussion, Gilles, Cecilia Rocha, Narges Abdian, Dimitri Yang, Julien Turk, Dulce Carrillo Valenzuela, Luisa Pimentel, Zhipeng You, Barbara Morquette, Michael Nicouleau, and et al. 2023. "Transcriptional Dysregulation and Impaired Neuronal Activity in FMR1 Knock-Out and Fragile X Patients’ iPSC-Derived Models" International Journal of Molecular Sciences 24, no. 19: 14926. https://doi.org/10.3390/ijms241914926
APA StyleMaussion, G., Rocha, C., Abdian, N., Yang, D., Turk, J., Carrillo Valenzuela, D., Pimentel, L., You, Z., Morquette, B., Nicouleau, M., Deneault, E., Higgins, S., Chen, C. X. -Q., Reintsch, W. E., Ho, S., Soubannier, V., Lépine, S., Modrusan, Z., Lund, J., ... Durcan, T. M. (2023). Transcriptional Dysregulation and Impaired Neuronal Activity in FMR1 Knock-Out and Fragile X Patients’ iPSC-Derived Models. International Journal of Molecular Sciences, 24(19), 14926. https://doi.org/10.3390/ijms241914926