

Genome-Wide Evolutionary Characterization and Expression Analysis of Major Latex Protein (MLP) Family Genes in Tomato


Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of the MLP Family Genes in Tomato
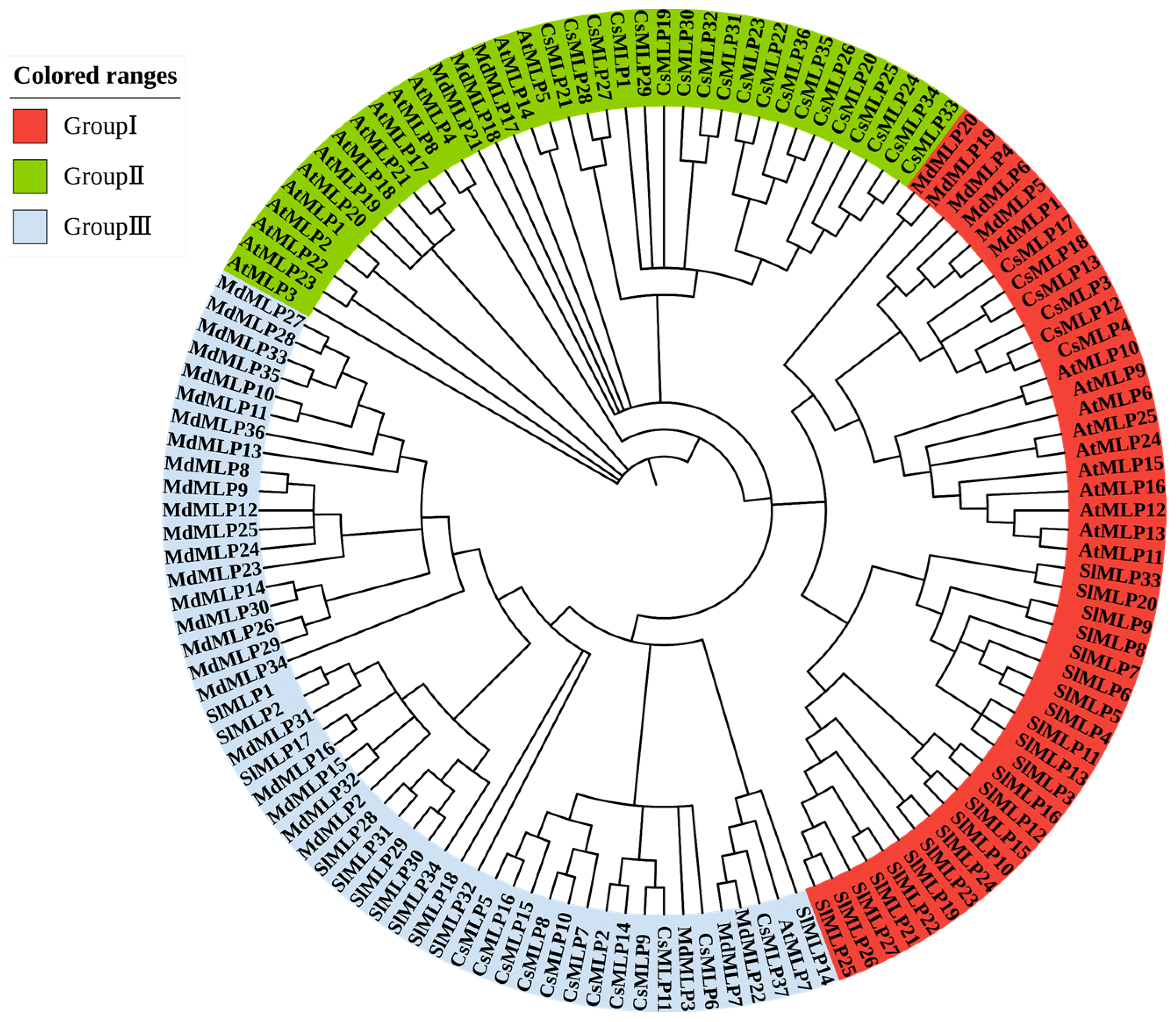
2.2. Phylogenetic Tree Analysis of SlMLP Genes
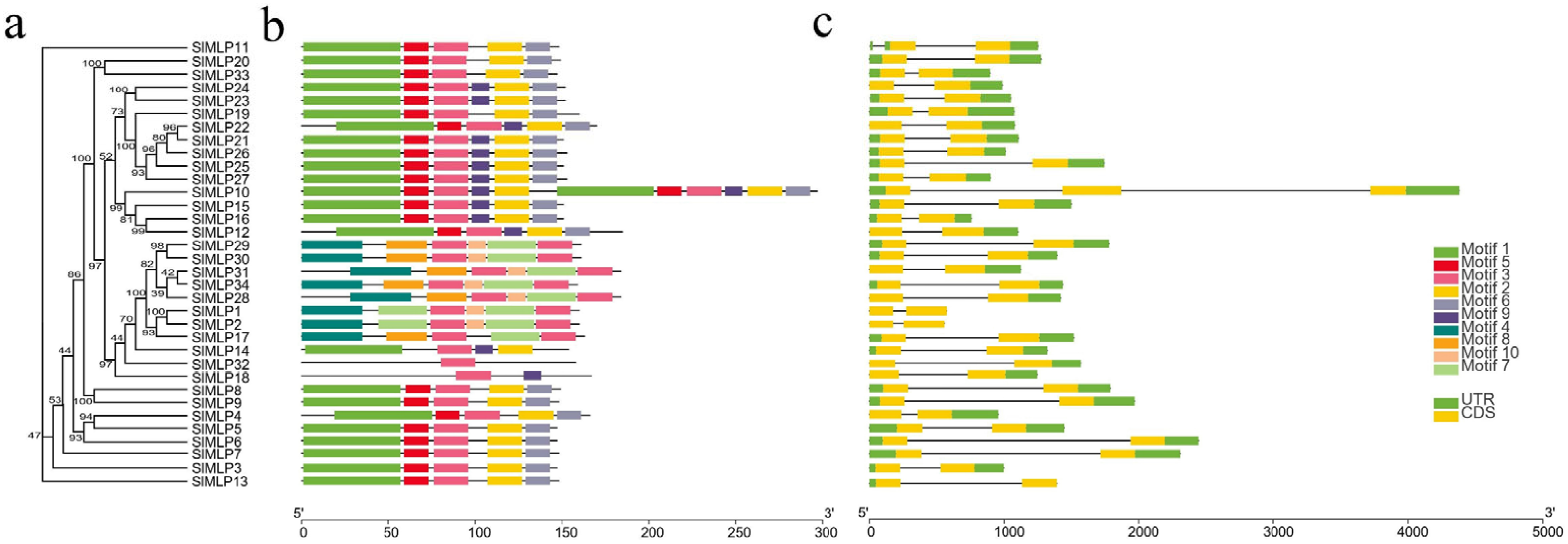
2.3. Gene Structure, Conserved Motif, and Domain Analysis of SlMLPs
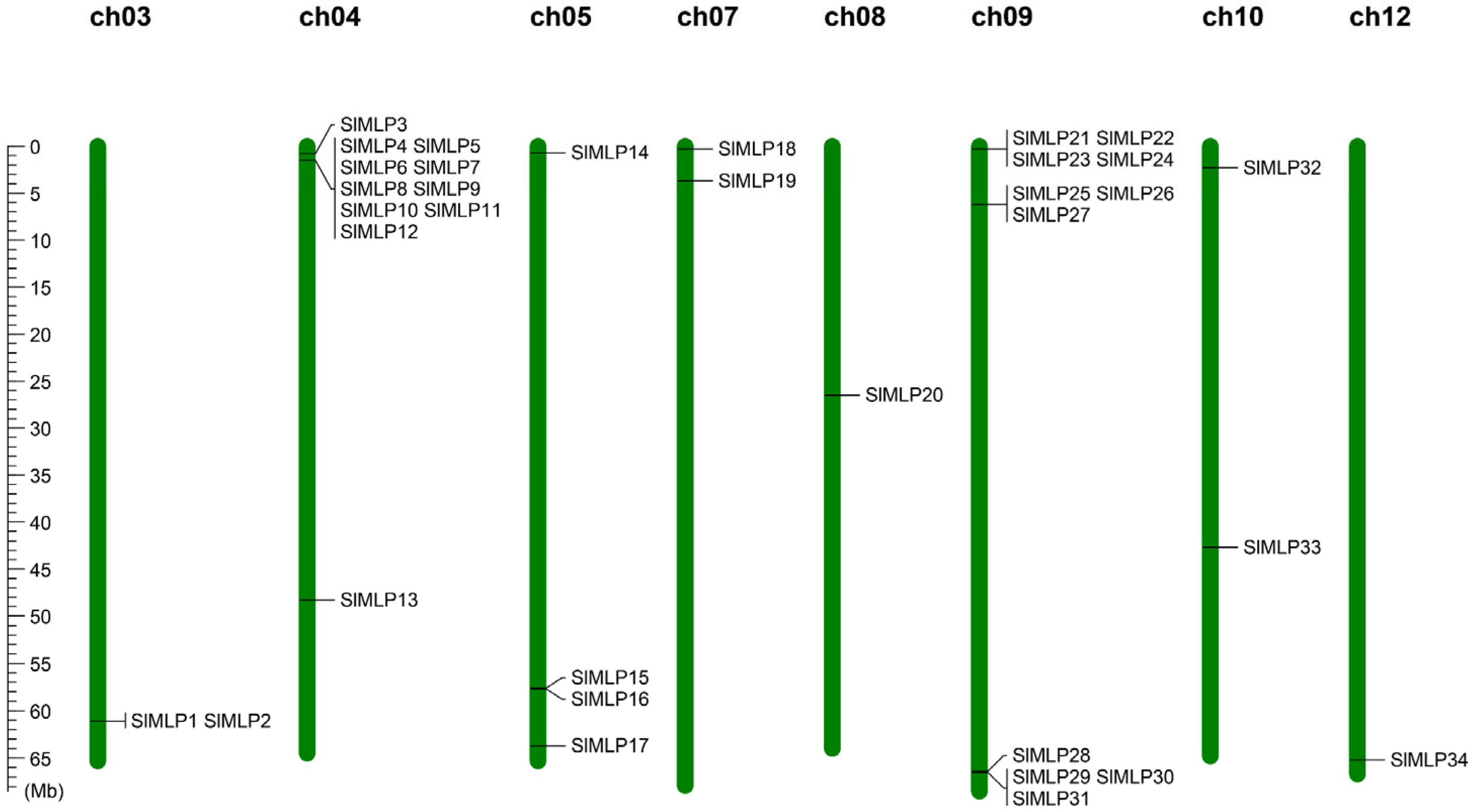
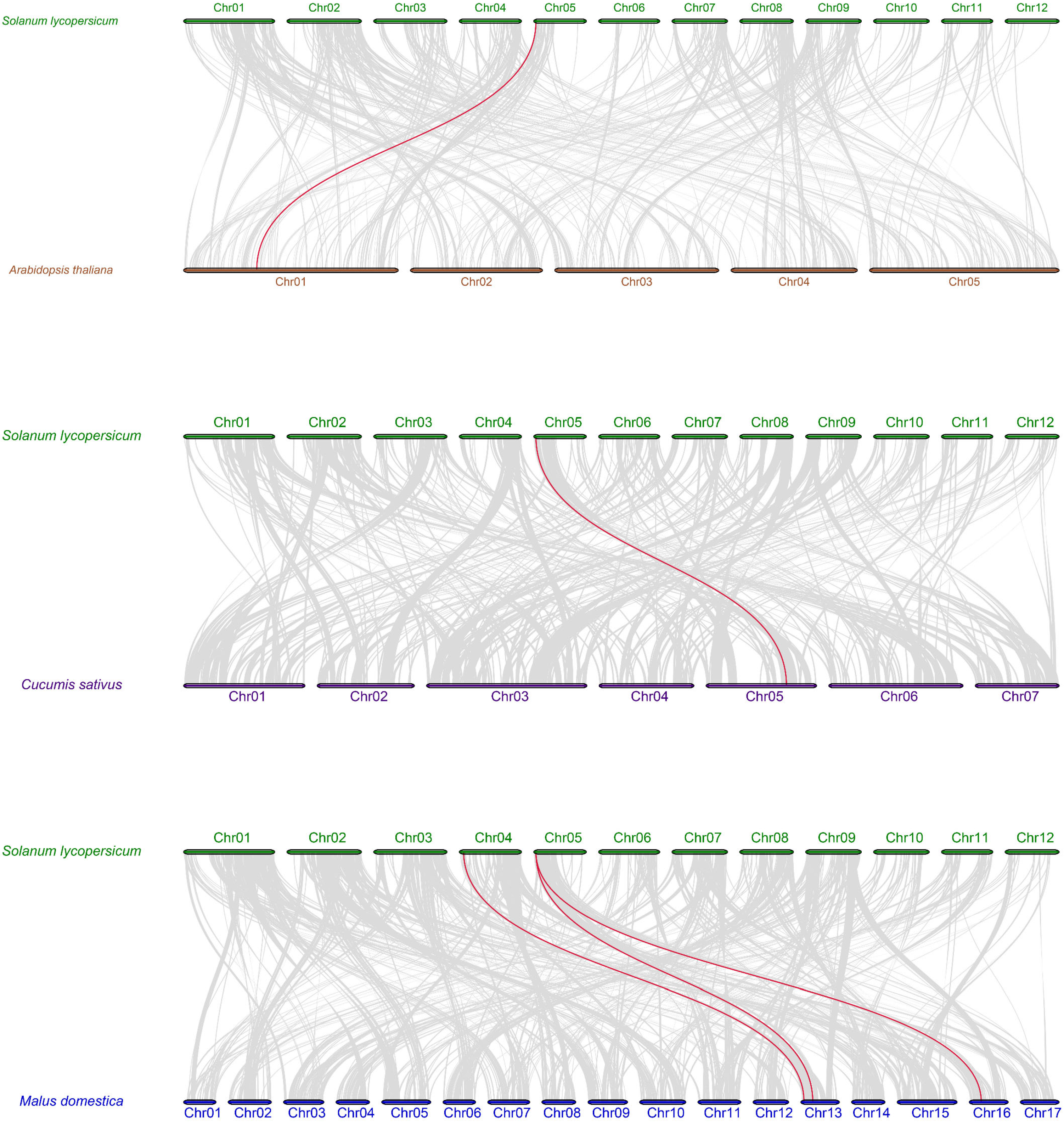
2.4. Chromosomal Location and Synteny Analysis
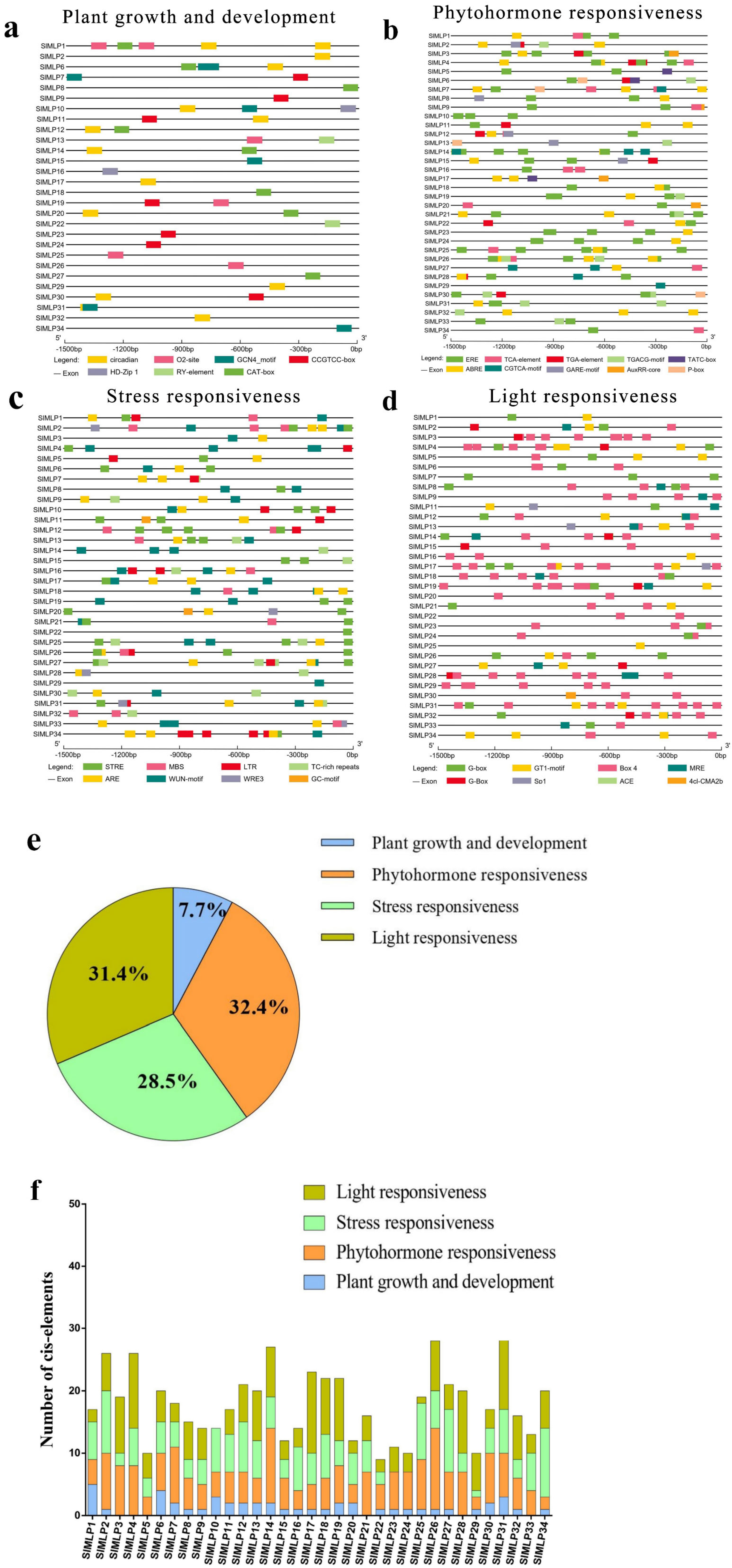
2.5. Cis-Element Analysis of SlMLP Promoters
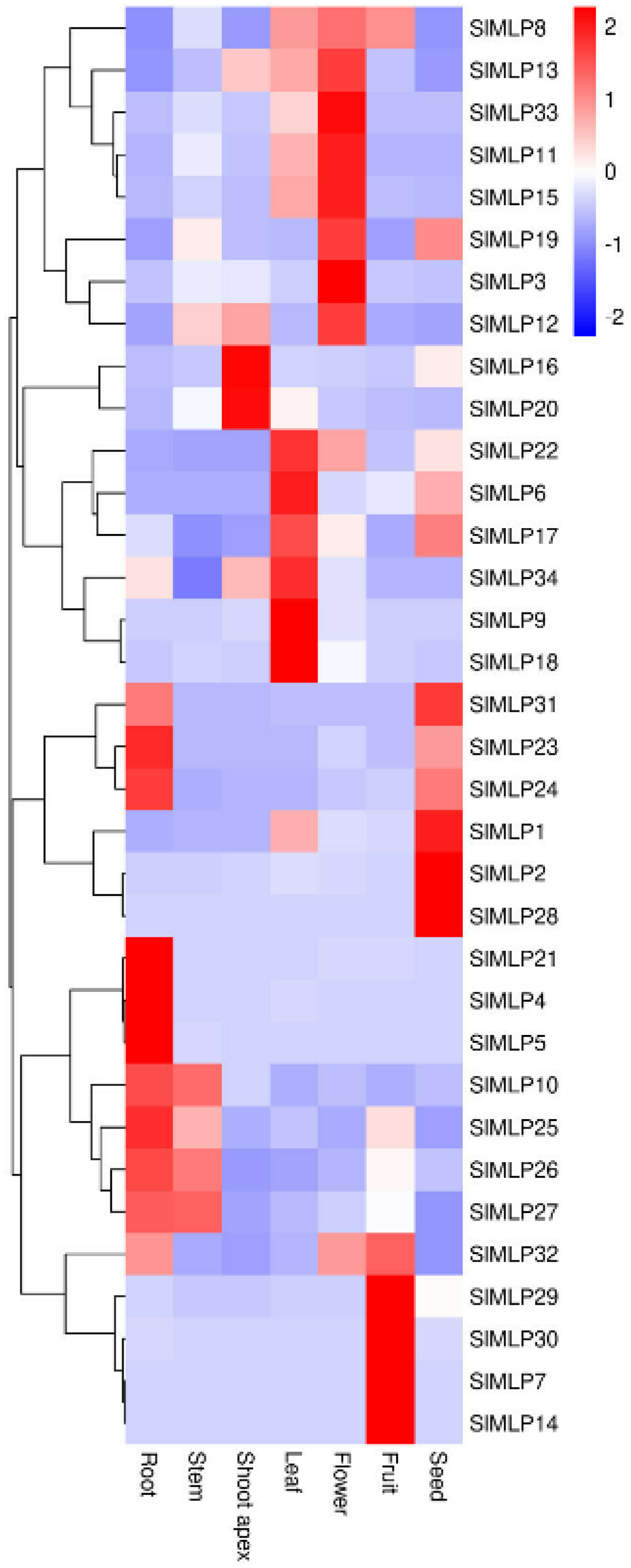
2.6. Tissue-Specific Expression Patterns of SlMLP Genes
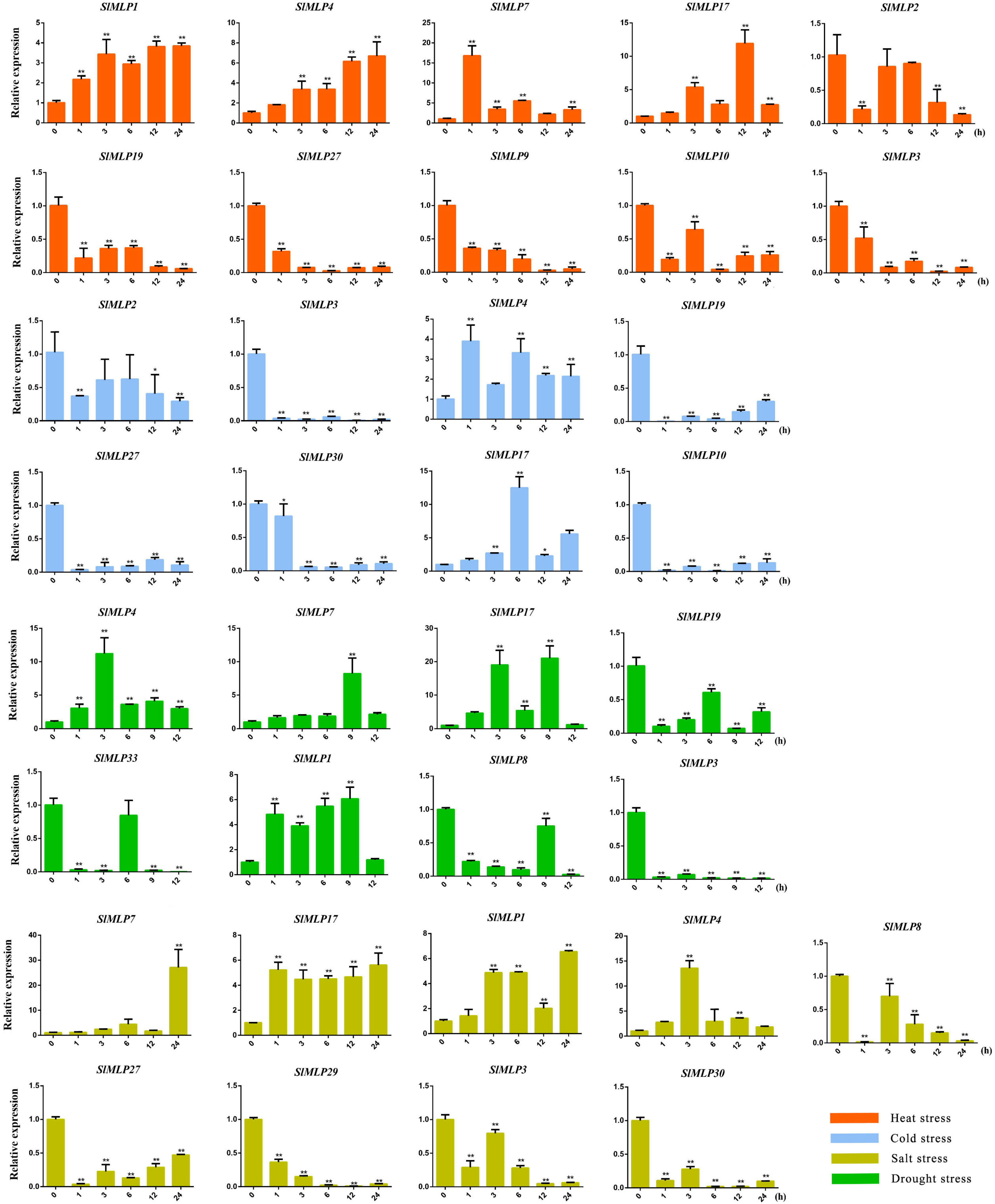
2.7. Expression Profiles of SlMLPs in Response to Abiotic Stresses
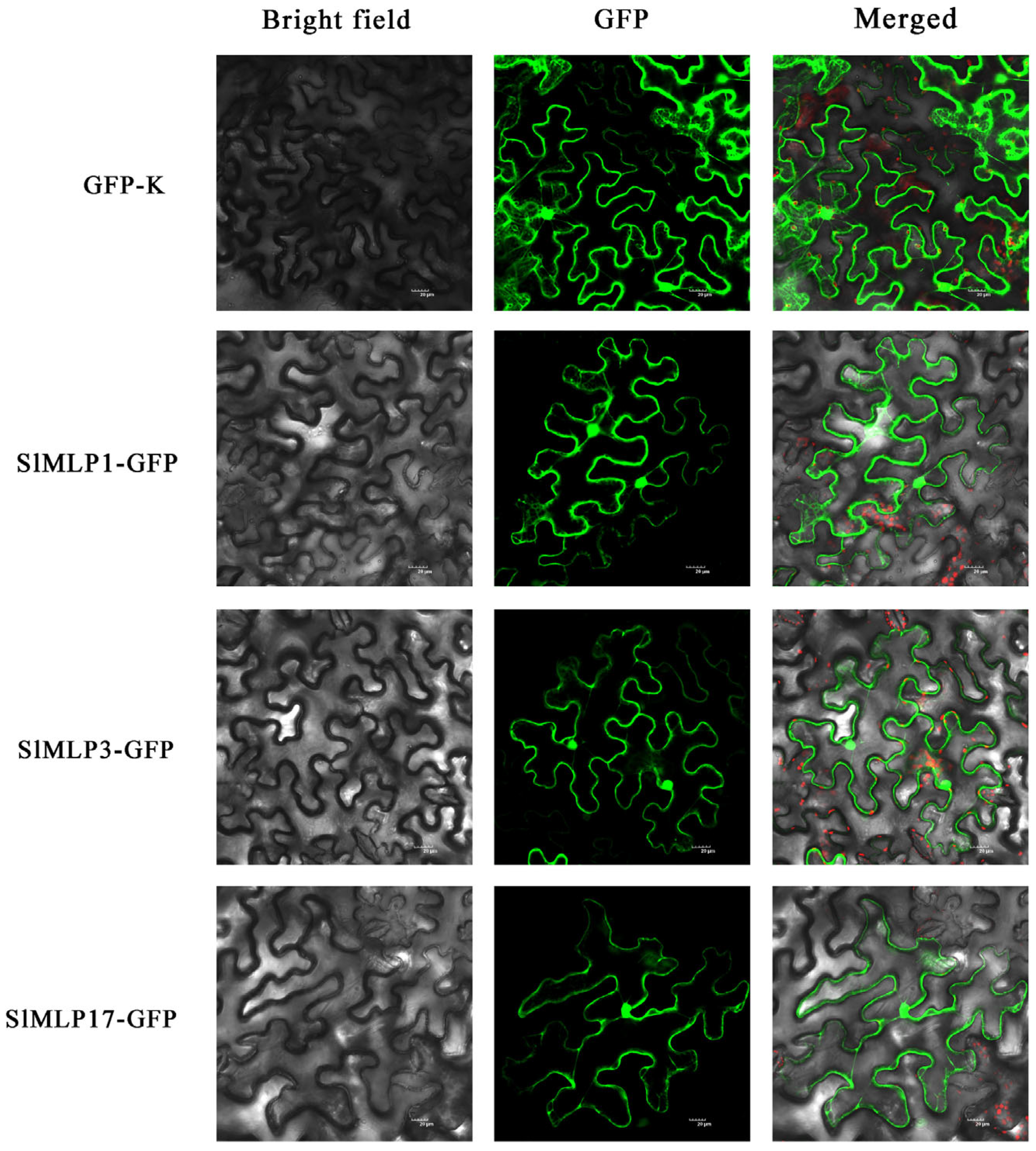
2.8. Subcellular Localization of SlMLP Proteins
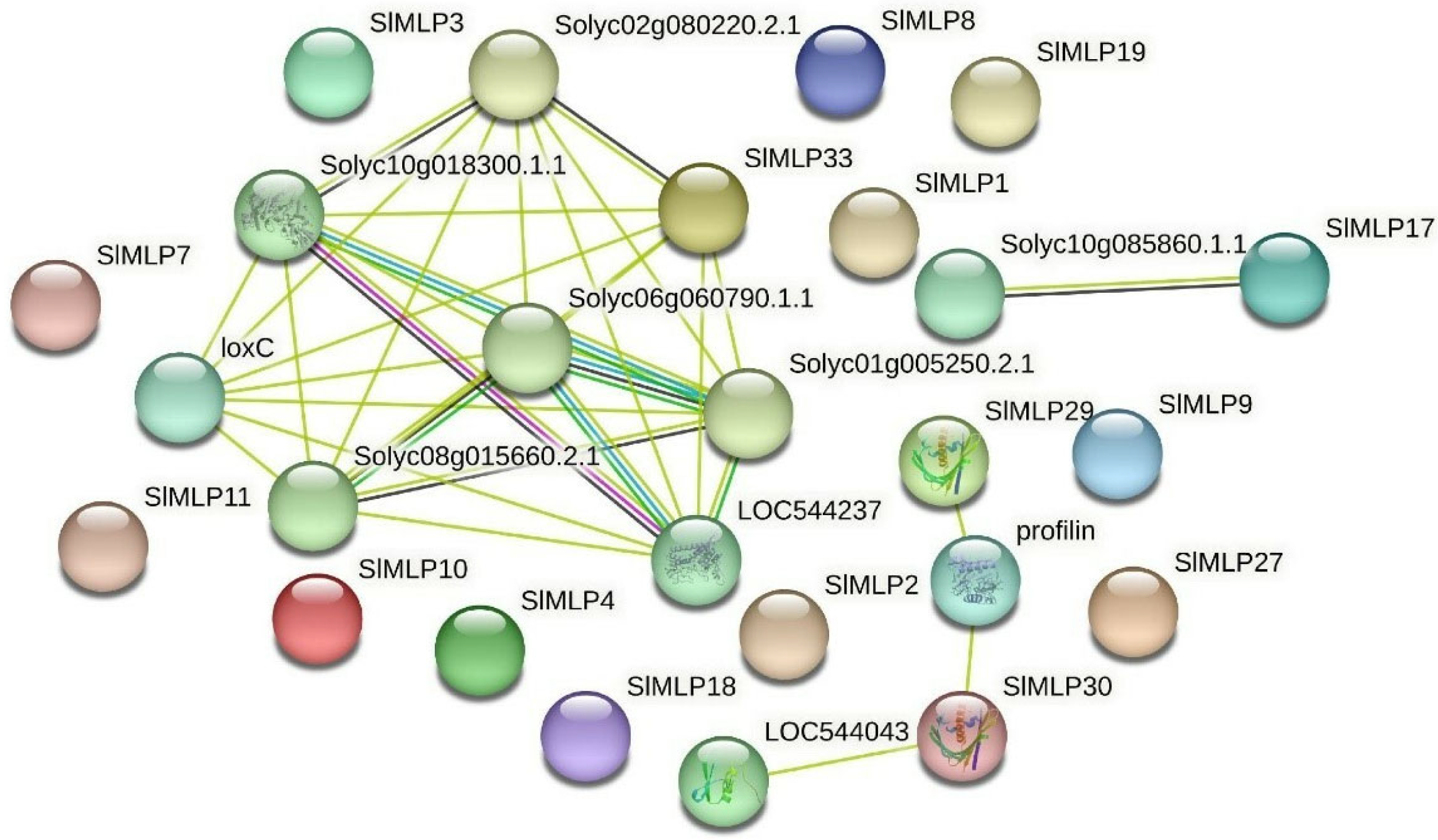
2.9. Analysis of the SlMLP Gene Expression Network
3. Discussion
3.1. Conservation and Evolution of the MLP Family
3.2. Conservation and Evolution of the MLP Family
3.3. Cis-Element Analysis of SlMLP Promoters
3.4. Potential Functions of Tomato SlMLP Genes in the Regulation of Plant Growth and Development
3.5. SlMLP Is Involved in the Response to Abiotic Stresses
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Identification of SlMLP Genes in Tomato
4.3. Phylogenetic Analysis
4.4. Analysis of Motifs, Gene Structures, and Conserved Domains
4.5. Chromosome Localization and Synteny Analysis
4.6. Analysis of the MLP Gene Promoter in Tomato
4.7. RNA Extraction and qRT-PCR Analysis
4.8. Subcellular Localization
4.9. Interaction Network Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, W.; Liu, K.; Chen, D.; Zhang, Z.; Li, B.; El-Mogy, M.M.; Tian, S.; Chen, T. Solanum lycopersicum, a Model Plant for the Studies in Developmental Biology, Stress Biology and Food Science. Foods 2022, 11, 2402. [Google Scholar] [CrossRef] [PubMed]
- Ram, K.; Akhter, A.W.; Akhilesh, Y.; Kumar, J.D.; Sudhir, K.; Kumar, S.A.; Major, S.; Prakash, V.J. Biotechnological Interventions in Tomato (Solanum lycopersicum) for Drought Stress Tolerance: Achievements and Future Prospects. Biotech 2022, 11, 48. [Google Scholar]
- Christian, R.; Peter, L.; Heimo, B. The Bet v 1 fold: An ancient, versatile scaffold for binding of large, hydrophobic ligands. BMC Evol. Biol. 2008, 8, 286. [Google Scholar]
- Betsy, L.L.; Jikui, S.; de la Cruz Norberto, B.; Peterson Francis, C.; Johnson Kenneth, A.; Bingman Craig, A.; Phillips George, N.; Volkman, B.F. Structures of two Arabidopsis thaliana major latex proteins represent novel helix-grip folds. Proteins 2009, 76, 237. [Google Scholar]
- Kentaro, F.; Hideyuki, I. Review: Biological functions of major latex-like proteins in plants. Plant Sci. 2021, 306, 110856. [Google Scholar]
- Nessler, C.L.; Allen, R.D.; Galewsky, S. Identification and Characterization of Latex-Specific Proteins in Opium Poppy. Plant Physiol. 1985, 79, 499–504. [Google Scholar] [CrossRef]
- Osmark, P.; Boyle, B.; Brisson, N. Sequential and structural homology between intracellular pathogenesis-related proteins and a group of latex proteins. Plant Mol. Biol. 1998, 38, 1243–1246. [Google Scholar] [CrossRef]
- Jie-Yin, C.; Xiao-Feng, D. Cloning and characterization of the Gossypium hirsutum major latex protein gene and functional analysis in Arabidopsis thaliana. Planta 2010, 231, 861–873. [Google Scholar]
- Suyama, T.; Yamada, K.; Mori, H.; Takeno, K.; Yamaki, S. Cloning cDNAs for Genes Preferentially Expressed during Fruit Growth in Cucumber. J. Am. Soc. Hortic. Sci. 1999, 124, 136–139. [Google Scholar] [CrossRef]
- Hye, C.S.; Ki, H.M.; Joong, K.H.; Nayeon, R.; Hyewhon, R.; Yeol, N.S.; Woo, K.L. Structure of ginseng major latex-like protein 151 and its proposed lysophosphatidic acid-binding mechanism. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71 Pt 5, 1039–1050. [Google Scholar]
- Humberto, F.; Karolina, M.; Michal, S.; Mariusz, J. Structural and functional aspects of PR-10 proteins. FEBS J. 2013, 280, 1169–1199. [Google Scholar]
- Ningbo, Z.; Ruimin, L.; Wei, S.; Shuzhen, J.; Junxiang, Z.; Weirong, X. Genome-wide evolutionary characterization and expression analyses of major latex protein (MLP) family genes in Vitis vinifera. Mol. Genet. Genom. MGG 2018, 293, 1061–1075. [Google Scholar]
- He, S.; Yuan, G.; Bian, S.; Han, X.; Liu, K.; Cong, P.; Zhang, C. Major Latex Protein MdMLP423 Negatively Regulates Defense against Fungal Infections in Apple. Int. J. Mol. Sci. 2020, 21, 1879. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; He, S.; Bian, S.; Han, X.; Liu, K.; Cong, P.; Zhang, C. Genome-wide identification and expression analysis of major latex protein (MLP) family genes in the apple (Malus domestica Borkh.) genome. Gene 2020, 733, 144275. [Google Scholar] [CrossRef] [PubMed]
- D’Avino, R.; Bernardi, M.L.; Wallner, M.; Palazzo, P.; Camardella, L.; Tuppo, L.; Alessandri, C.; Breiteneder, H.; Ferreira, F.; Ciardiello, M.A.; et al. Kiwifruit Act d 11 is the first member of the ripening-related protein family identified as an allergen. Allergy 2011, 66, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Aggelis, A.; John, I.; Karvouni, Z.; Grierson, D. Characterization of two cDNA clones for mRNAs expressed during ripening of melon (Cucumis melo L.) fruits. Plant Mol. Biol. 1997, 33, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Strömvik, M.V.; Sundararaman, V.P.; Vodkin, L.O. A novel promoter from soybean that is active in a complex developmental pattern with and without its proximal 650 base pairs. Plant Mol. Biol. 1999, 41, 217–231. [Google Scholar] [CrossRef]
- Yang, C.-L.; Liang, S.; Wang, H.-Y.; Han, L.-B.; Wang, F.-X.; Cheng, H.-Q.; Wu, X.-M.; Qu, Z.-L.; Wu, J.-H.; Xia, G.-X. Cotton Major Latex Protein 28 Functions as a Positive Regulator of the Ethylene Responsive Factor 6 in Defense against Verticillium dahliae. Mol. Plant 2015, 8, 399–411. [Google Scholar] [CrossRef]
- Jie, L.; Ruier, Z.; Zijun, H.; Hengkuan, G.; Shiyuan, L.; Yu, G.; Suzhe, Y.; Ying, W.; Hui, Z.; Lei, Z.; et al. Genome-wide charac-terization of major latex protein gene family in peanut and expression analyses under drought and waterlogging stress. Front. Plant Sci. 2023, 14, 1152824. [Google Scholar]
- Hideyuki, I.; Mami, S.; Junya, G.; Kiyoshi, Y.; Noriko, K.; Hiroki, T.; Heesoo, E. A major latex-like protein is a key factor in crop contamination by persistent organic pollutants. Plant Physiol. 2013, 161, 2128–2135. [Google Scholar]
- Li, P.; Chen, L.; Zhou, Y.; Xia, X.; Shi, K.; Chen, Z.; Yu, J. Brassinosteroids-Induced Systemic Stress Tol-erance was Associated with Increased Transcripts of Several Defence-Related Genes in the Phloem in Cucumis sativus. PLoS ONE 2013, 8, e66582. [Google Scholar]
- Philip, C.; Juliane, M.-P.; Daniel, C.W.; Sanjukta, D.; Stefanie, M.H.; Corina, V.A.; Cameron, R.K. Comparative Proteomics Analysis of Phloem Exudates Collected during the Induction of Systemic Acquired Resistance. Plant Physiol. 2016, 171, 1495–1510. [Google Scholar]
- Goto, J.; Iwabuchi, A.; Yoshihara, R.; Kodama, N.; Matsui, T.; Hirota, M.; Eun, H.; Inui, H. Uptake mechanisms of poly-chlorinated biphenyls in Cucurbita pepo via xylem sap containing major latex-like proteins. Environ. Exp. Bot. 2019, 162, 399–405. [Google Scholar] [CrossRef]
- Mirza, O.; Henriksen, A.; Ipsen, H.; Larsen, J.N.; Wissenbach, M.; Spangfort, M.D.; Gajhede, M. Dominant Epitopes and Allergic Cross-Reactivity: Complex Formation Between a Fab Fragment of a Monoclonal Murine IgG Antibody and the Major Allergen from Birch Pollen Bet v 1. J. Immunol. 2000, 165, 331–338. [Google Scholar] [CrossRef]
- Michael, D.S.; Osman, M.; Henrik, I.; Van Neerven, R.J.J.; Gajhede, M.; Larsen, J.N. Dominating IgE-binding epitope of Bet v 1, the major allergen of birch pollen, characterized by X-ray crystallography and site-directed mutagenesis. J. Immunol. 2003, 171, 3084–3090. [Google Scholar]
- Nessler, C.L.; Kurz, W.G.W.; Pelcher, L.E. Isolation and analysis of the major latex protein genes of opium poppy. Plant Mol. Biol. 1990, 15, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Ruperti, B.; Bonghi, C.; Ziliotto, F.; Pagni, S.; Rasori, A.; Varotto, S.; Tonutti, P.; Giovannoni, J.J.; Ramina, A. Characterization of a major latex protein (MLP) gene down-regulated by ethylene during peach fruitlet abscission. Plant Sci. 2002, 163, 265–272. [Google Scholar] [CrossRef]
- Yanping, W.; Li, Y.; Xi, C.; Tiantian, Y.; Bao, Z.; Ruijie, L.; Yan, W.; Zhulong, C. Major latex protein-like protein 43 (MLP43) functions as a positive regulator during abscisic acid responses and confers drought tolerance in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 421–434. [Google Scholar]
- Facchini, P.J.; Park, S.-U. Developmental and inducible accumulation of gene transcripts involved in alkaloid biosynthesis in opium poppy. Phytochemistry 2003, 64, 177–186. [Google Scholar] [CrossRef]
- Marion, W.; Jörg, Z.; Toni, M.K. The roles of latex and the vascular bundle in morphine biosynthesis in the opium poppy, Papaver somniferum. Proc. Natl. Acad. Sci. USA 2004, 101, 13957–13962. [Google Scholar]
- Toni, M.; Kutchan. A role for intra- and intercellular translocation in natural product biosynthesis. Curr. Opin. Plant Biol. 2005, 8, 292–300. [Google Scholar]
- Di, G.; Shing, W.W.; Zhao, X.W.; Fei, S.F.; Jin, Q.D.; Ning, L. Cis-cinnamic acid-enhanced 1 gene plays a role in regulation of Arabidopsis bolting. Plant Mol. Biol. 2011, 75, 481–495. [Google Scholar]
- Celso, G.L.; Benjamin, L.P.; Andrew, L.E.; Martin, R.L.; Stuart, J.C.; Peter, M.W. Proteomic Identification of Putative Mi-croRNA394 Target Genes in Arabidopsis thaliana Identifies Major Latex Protein Family Members Critical for Normal Devel-opment. Mol. Cell. Proteom. 2016, 15, 2033–2047. [Google Scholar]
- Nawrot, R.; Lippmann, R.; Matros, A.; Musidlak, O.; Nowicki, G.; Mock, H.-P. Proteomic comparison of Chelidonium majus L. latex in different phases of plant development. Plant Physiol. Biochem. 2017, 112, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Yan, Y.; Erik, C.S.; Linda, P.M.; Lara, D.L.; Brian, J.H.; William, C.N.; John, Q. Transcriptional divergence of the du-plicated oxidative stress-responsive genes in the Arabidopsis genome. Plant J. Cell Mol. Biol. 2005, 41, 212–220. [Google Scholar]
- Nam, Y.-W.; Tichit, L.; Leperlier, M.; Cuerq, B.; Marty, I.; Lelièvre, J.-M. Isolation and characterization of mRNAs differentially expressed during ripening of wild strawberry (Fragaria vesca L.) fruits. Plant Mol. Biol. 1999, 39, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Ciardiello, M.A.; Osinski, T.; Majorek, K.A.; Giangrieco, I.; Font, J.; Breiteneder, H.; Thalassinos, K.; Minor, W. Structural and bioinformatic analysis of the kiwifruit allergen Act d 11, a member of the family of ripening-related proteins. Mol. Immunol. 2013, 56, 794–803. [Google Scholar] [CrossRef]
- Liang, Q.Z.; Yun, W.H.; Xian, X.G. GhHb1: A nonsymbiotic hemoglobin gene of cotton responsive to infection by Verticillium dahliae. BBA Gene Struct. Expr. 2005, 1730, 103–113. [Google Scholar]
- Bong-Kwan, P.; Sebyul, P.; Jin-Hwan, C.; Jong-Seong, J.; Hee, B.S.; Tae-Ryong, H. Comparative proteomic analysis of blue light signaling components in the Arabidopsis cryptochrome 1 mutant. Mol. Cells 2007, 23, 154–160. [Google Scholar]
- Hua, S.; Myung-Kyum, K.; Krishna, P.R.; Yu-Jin, K.; Deok-Chun, Y. Isolation and expression analysis of a novel major latex-like protein (MLP151) gene from Panax ginseng. Mol. Biol. Rep. 2010, 37, 2215–2222. [Google Scholar]
- Jeffery, M.K.; Raul, S.-M.; Wendy, F.B.; Christopher, S.B.; Winter, S.H. The fast and transient transcriptional network of gravity and mechanical stimulation in the Arabidopsis root apex. Plant Physiol. 2004, 136, 2790–2805. [Google Scholar]
- Robert, D.F.; Jody, C.; Sean, R.E. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar]
- Ivica, L.; Supriya, K.; Smart, B.P. New developments and status in 2020. Nucleic Acids Res. 2021, 49, D1. [Google Scholar]
- Timothy, L.B.; James, J.; Charles, E.G. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for In-teractive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shen, Y.; Wei, W.; Zhou, D.-X. Histone Acetylation Enzymes Coordinate Metabolism and Gene Expression. Trends Plant Sci. 2015, 20, 614–621. [Google Scholar] [CrossRef]
- Toyota, M.; Spencer, D.; Sawai-Toyota, S.; Jiaqi, W.; Zhang, T.; Koo, A.J.; Howe, G.A. Glutamate triggers long-distance, calci-um-based plant defense signaling. Science 2018, 361, 1112–1115. [Google Scholar] [CrossRef]
- Tatjana, M.H. Synthesis versus degradation: Directions of amino acid metabolism during Arabidopsis abiotic stress response. Plant Mol. Biol. 2018, 98, 121–135. [Google Scholar]
- Lawrence, R.G.; Craig, L.N. Immunolocalization of the Major Latex Proteins in Developing Laticifers of Opium Poppy (Papaver somniferum). J. Plant Physiol. 1989, 134, 357–363. [Google Scholar]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell 1995, 7, 173–182. [Google Scholar] [PubMed]
- Wu, Q.; Bai, J.; Tao, X.; Mou, W.; Luo, Z.; Mao, L.; Ban, Z.; Ying, T.; Li, L. Synergistic effect of abscisic acid and ethylene on color development in tomato (Solanum lycopersicum L.) fruit. Sci. Hortic. 2018, 235, 169–180. [Google Scholar] [CrossRef]
- Kazuo, N.; Yusuke, I.; Kazuko, Y.-S. Transcriptional regulatory networks in response to abiotic stresses in Arabidopsis and grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Zheng, P.; Zhang, C.; Wang, W.; Li, Y.; Fan, T.; Liu, Y.; Cao, S. The effect of ABRE BINDING FACTOR 4-mediated FYVE1 on salt stress tolerance in Arabidopsis. Plant Sci. 2020, 296, 110489. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Tong, J.; Liu, W.; Jiang, Z.; Pan, G.; Ning, X.; Yang, X.; Zhong, M. Comprehensive Analysis of Major La-tex-Like Protein Family Genes in Cucumber (Cucumis sativus L.) and Their Potential Roles in Phytophthora Blight Resistance. Int. J. Mol. Sci. 2023, 24, 784. [Google Scholar] [CrossRef]
- Jun, L. Tomato pan-genome. Nat. Plants 2019, 5, 6. [Google Scholar]
- Hitomi, T.; Kenta, S.; Kosuke, K.; Atsushi, T.; Yuma, M.; Shinji, I.; Tohru, A. De novo genome assembly of two tomato ancestors, Solanum pimpinellifolium and Solanum lycopersicum var. cerasiforme, by long-read sequencing. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2021, 28, dsaa029. [Google Scholar]
- Zeng, J.; Ruan, Y.; Liu, B.; Ruan, Y.; Huang, Y. Genome-wide identification and abiotic stress-responsive expression of MLP family genes in Brassica rapa. Gene Rep. 2020, 21, 100919. [Google Scholar] [CrossRef]
- Kentaro, F.; Natsumi, C.; Maho, C.; Shoya, K.; Chihiro, S.; Minami, Y.; Hideyuki, I. Genome-wide identification and character-ization of major latex-like protein genes responsible for crop contamination in Cucurbita pepo. Mol. Biol. Rep. 2022, 49, 7773–7782. [Google Scholar]
- Yang, N.; Wang, C.-L.; He, W.-P.; Qu, Y.-Z.; Li, Y.-S. Photosynthetic characteristics and effects of exogenous glycine of Chorispora bungeana under drought stres. Photosynthetica 2016, 54, 459–467. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | CDS Size | Protein | GRAVY | Instability Index | Subcellular Localization Prediction | ||
|---|---|---|---|---|---|---|---|---|
| (bp) | Length (aa) | MW (kDa) | pI | |||||
| SlMLP1 | Solyc03g117450.1 | 480 | 159 | 17.61 | 5.02 | −0.350 | 32.81 | Cytoplasm |
| SlMLP2 | Solyc03g117460.1 | 480 | 159 | 17.64 | 4.90 | −0.401 | 34.90 | Cytoplasm |
| SlMLP3 | Solyc04g007010.3 | 441 | 146 | 16.61 | 5.96 | −0.168 | 33.78 | Cytoplasm |
| SlMLP4 | Solyc04g007750.4 | 498 | 165 | 18.93 | 5.85 | −0.256 | 34.38 | Cytoplasm |
| SlMLP5 | Solyc04g007760.3 | 441 | 146 | 16.58 | 6.03 | −0.200 | 25.97 | Cytoplasm |
| SlMLP6 | Solyc04g150102.1 | 441 | 146 | 16.57 | 5.07 | −0.147 | 41.68 | Cytoplasm |
| SlMLP7 | Solyc04g007770.3 | 444 | 147 | 16.60 | 5.96 | −0.162 | 42.21 | Cytoplasm |
| SlMLP8 | Solyc04g007780.3 | 447 | 148 | 17.02 | 5.62 | −0.403 | 19.96 | Cytoplasm |
| SlMLP9 | Solyc04g007790.3 | 444 | 147 | 16.82 | 5.63 | −0.407 | 17.31 | Cytoplasm |
| SlMLP10 | Solyc04g150104.1 | 891 | 296 | 34.40 | 5.10 | −0.429 | 39.14 | Cytoplasm, Cytoskeleton |
| SlMLP11 | Solyc04g007820.3 | 444 | 147 | 16.77 | 5.72 | −0.365 | 35.06 | Nucleus |
| SlMLP12 | Solyc04g007825.2 | 555 | 179 | 20.96 | 5.79 | −0.411 | 35.98 | Cytoplasm |
| SlMLP13 | Solyc04g050950.3 | 444 | 147 | 17.15 | 5.90 | −0.351 | 36.51 | Cytoplasm |
| SlMLP14 | Solyc05g005865.1 | 462 | 153 | 17.09 | 5.17 | −0.231 | 24.73 | Extracellular |
| SlMLP15 | Solyc05g046140.3 | 453 | 150 | 17.60 | 5.22 | −0.467 | 50.34 | Cytoplasm |
| SlMLP16 | Solyc05g046150.3 | 453 | 150 | 17.70 | 5.47 | −0.456 | 51.29 | Cytoplasm |
| SlMLP17 | Solyc05g054380.2 | 489 | 162 | 18.18 | 4.78 | −0.188 | 37.17 | Cytoplasm |
| SlMLP18 | Solyc07g005370.4 | 501 | 166 | 18.82 | 5.48 | −0.150 | 25.62 | Cytoplasm |
| SlMLP19 | Solyc07g008710.3 | 480 | 159 | 18.42 | 5.31 | −0.613 | 32.72 | Cytoplasm |
| SlMLP20 | Solyc08g023660.3 | 447 | 148 | 17.07 | 6.50 | −0.420 | 16.91 | Cytoplasm |
| SlMLP21 | Solyc09g005400.3 | 453 | 150 | 17.07 | 5.17 | −0.187 | 33.43 | Cytoplasm |
| SlMLP22 | Solyc09g005420.4 | 510 | 169 | 19.39 | 5.29 | −0.166 | 28.23 | Extracellular |
| SlMLP23 | Solyc09g005425.1 | 456 | 151 | 17.43 | 5.10 | −0.438 | 40.68 | Extracellular |
| SlMLP24 | Solyc09g005500.3 | 456 | 151 | 17.43 | 5.10 | −0.438 | 40.68 | Extracellular |
| SlMLP25 | Solyc09g014525.1 | 453 | 150 | 17.10 | 5.57 | −0.223 | 25.66 | Cytoplasm |
| SlMLP26 | Solyc09g014550.3 | 459 | 152 | 17.41 | 5.47 | −0.266 | 24.29 | Cytoplasm |
| SlMLP27 | Solyc09g014580.3 | 459 | 152 | 17.42 | 5.62 | −0.248 | 31.78 | Cytoplasm |
| SlMLP28 | Solyc09g090970.4 | 552 | 183 | 20.55 | 6.41 | −0.322 | 25.35 | Cytoplasm |
| SlMLP29 | Solyc09g090980.3 | 483 | 160 | 17.37 | 5.44 | −0.128 | 30.62 | Cytoplasm |
| SlMLP30 | Solyc09g090990.2 | 483 | 160 | 17.91 | 5.34 | −0.426 | 32.81 | Cytoplasm |
| SlMLP31 | Solyc09g091000.4 | 552 | 183 | 20.76 | 6.38 | −0.383 | 35.34 | Cytoplasm |
| SlMLP32 | Solyc10g008330.4 | 474 | 157 | 18.29 | 8.73 | −0.718 | 25.59 | Cytoplasm |
| SlMLP33 | Solyc10g048030.2 | 441 | 146 | 16.69 | 6.37 | −0.305 | 13.79 | Cytoplasm |
| SlMLP34 | Solyc12g096960.2 | 477 | 158 | 18.20 | 5.03 | −0.218 | 38.70 | Cytoplasm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Meng, L.; Yao, Y.; Zhang, Y.; Cheng, B.; Liang, Y. Genome-Wide Evolutionary Characterization and Expression Analysis of Major Latex Protein (MLP) Family Genes in Tomato. Int. J. Mol. Sci. 2023, 24, 15005. https://doi.org/10.3390/ijms241915005
Sun Z, Meng L, Yao Y, Zhang Y, Cheng B, Liang Y. Genome-Wide Evolutionary Characterization and Expression Analysis of Major Latex Protein (MLP) Family Genes in Tomato. International Journal of Molecular Sciences. 2023; 24(19):15005. https://doi.org/10.3390/ijms241915005
Chicago/Turabian StyleSun, Zhengliang, Liangzhe Meng, Yuhe Yao, Yanhong Zhang, Baohui Cheng, and Yan Liang. 2023. "Genome-Wide Evolutionary Characterization and Expression Analysis of Major Latex Protein (MLP) Family Genes in Tomato" International Journal of Molecular Sciences 24, no. 19: 15005. https://doi.org/10.3390/ijms241915005
APA StyleSun, Z., Meng, L., Yao, Y., Zhang, Y., Cheng, B., & Liang, Y. (2023). Genome-Wide Evolutionary Characterization and Expression Analysis of Major Latex Protein (MLP) Family Genes in Tomato. International Journal of Molecular Sciences, 24(19), 15005. https://doi.org/10.3390/ijms241915005