


Human Astrocyte Spheroids as Suitable In Vitro Screening Model to Evaluate Synthetic Cannabinoid MAM2201-Induced Effects on CNS
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
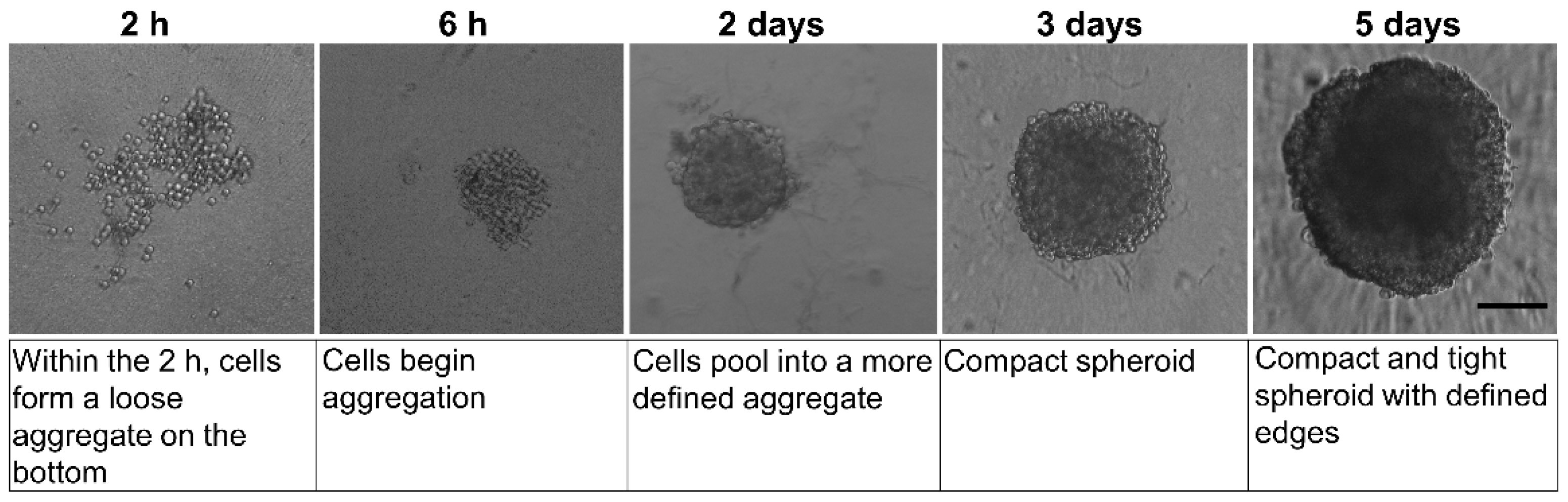
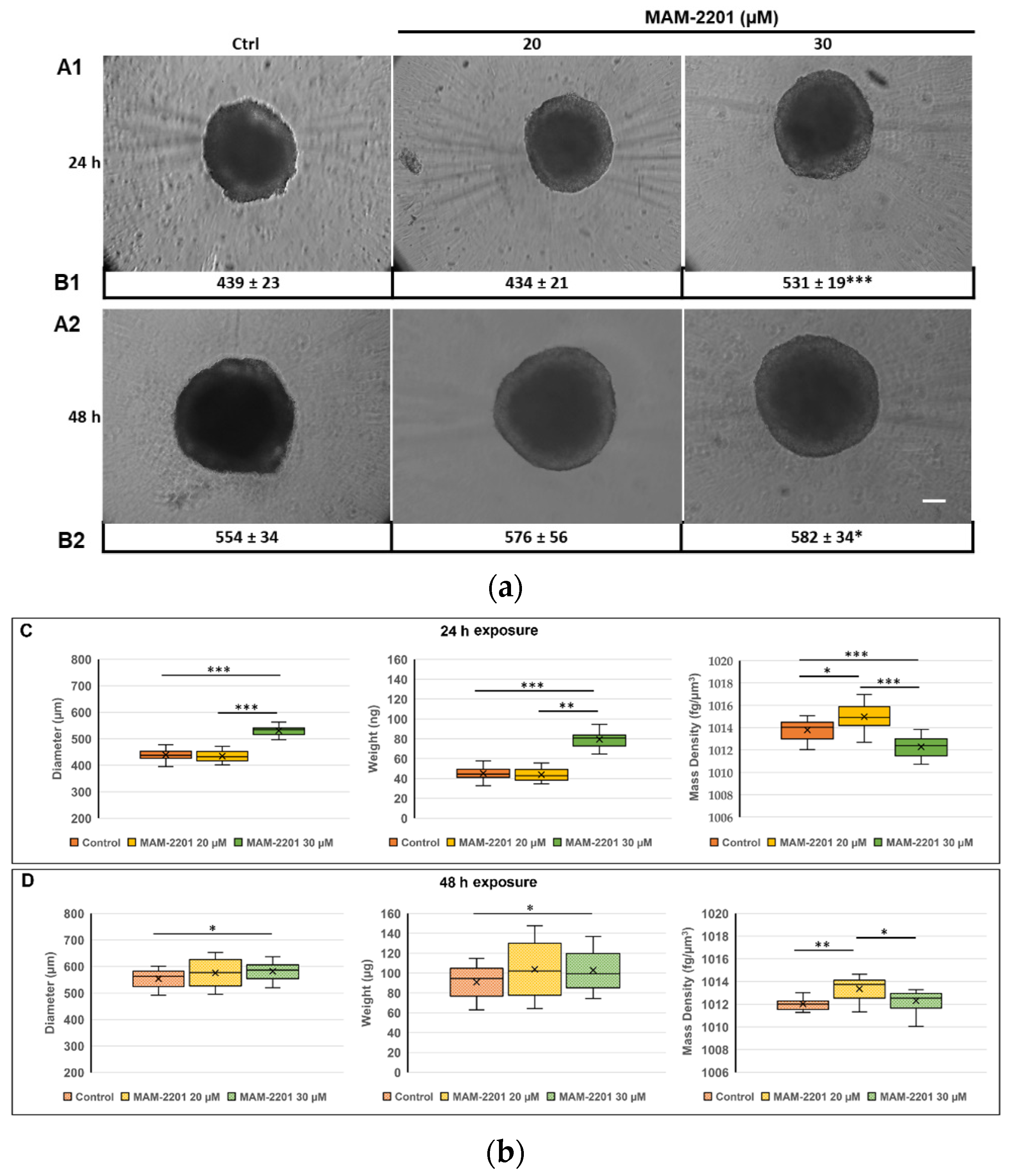
2.1. Effects of MAM-2201 Exposure on Astrocyte Spheroid Morphology
2.1.1. Spheroid Characterization by Phase-Contrast Microscopy Imaging and Physical Cytometry
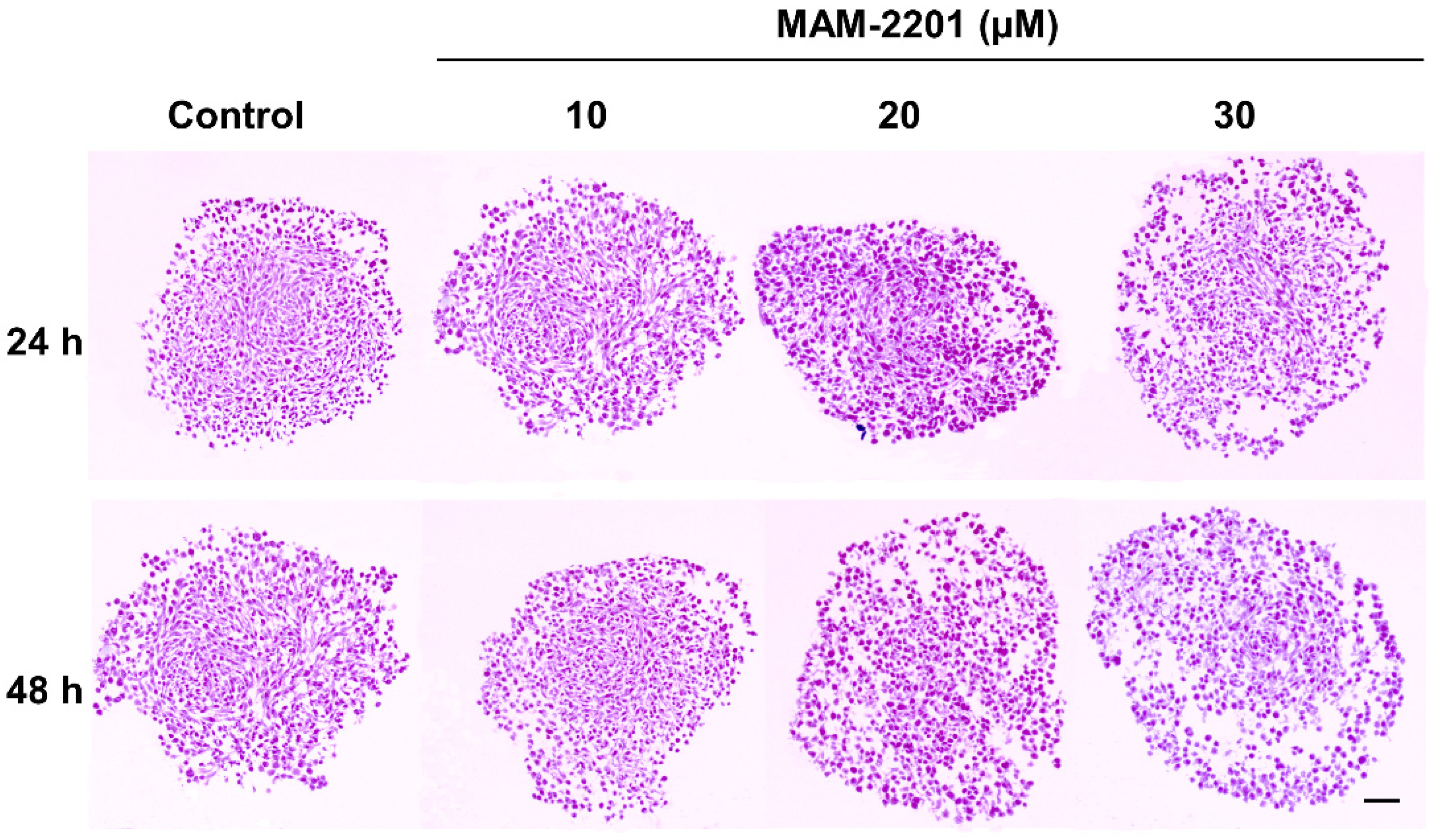
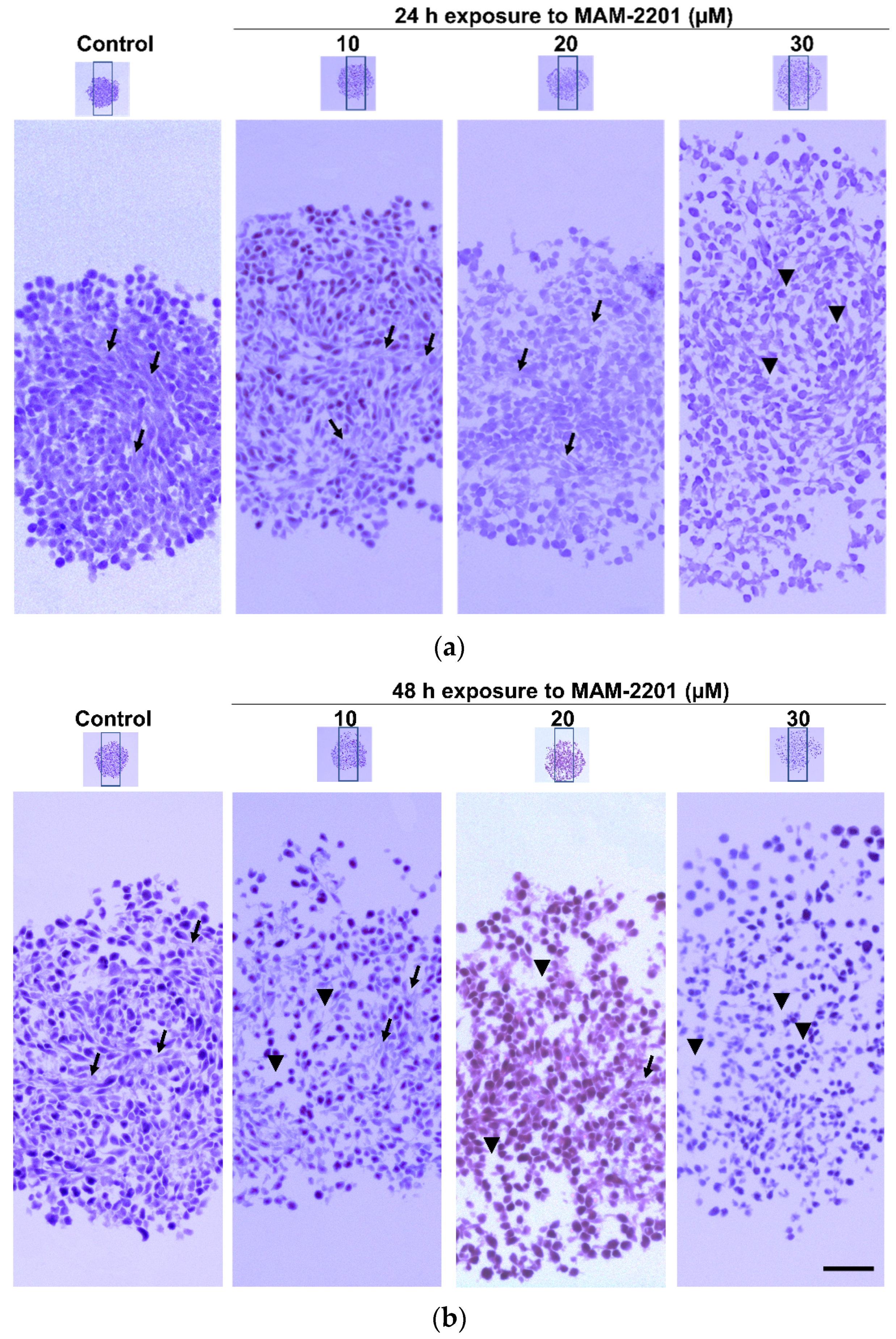
2.1.2. Basic Tissue Structure of the Astrocyte Spheroids by H&E Light Microscopy, ECM, and E-Cadherin Stains
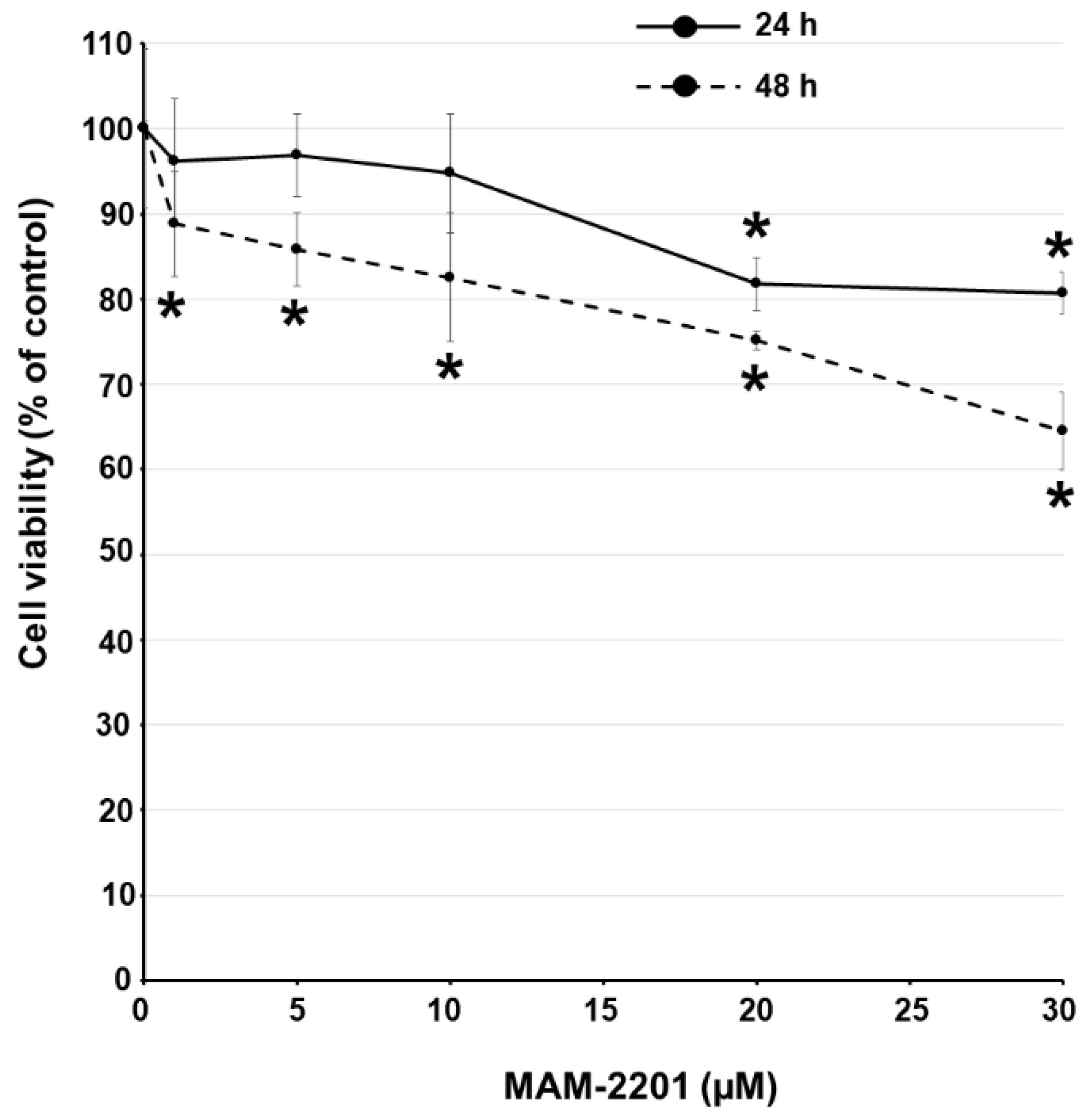
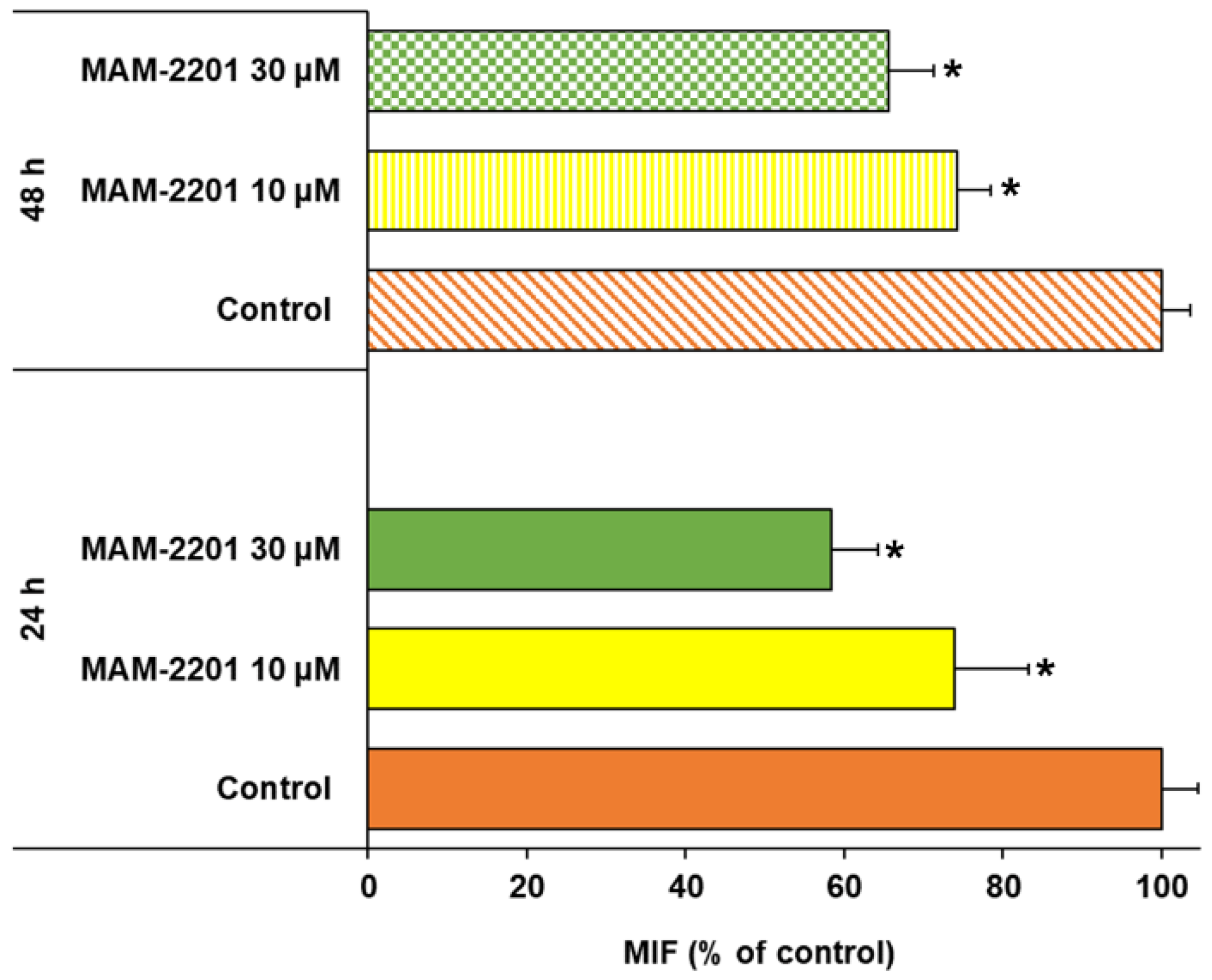
2.2. Cell Viability Evaluation
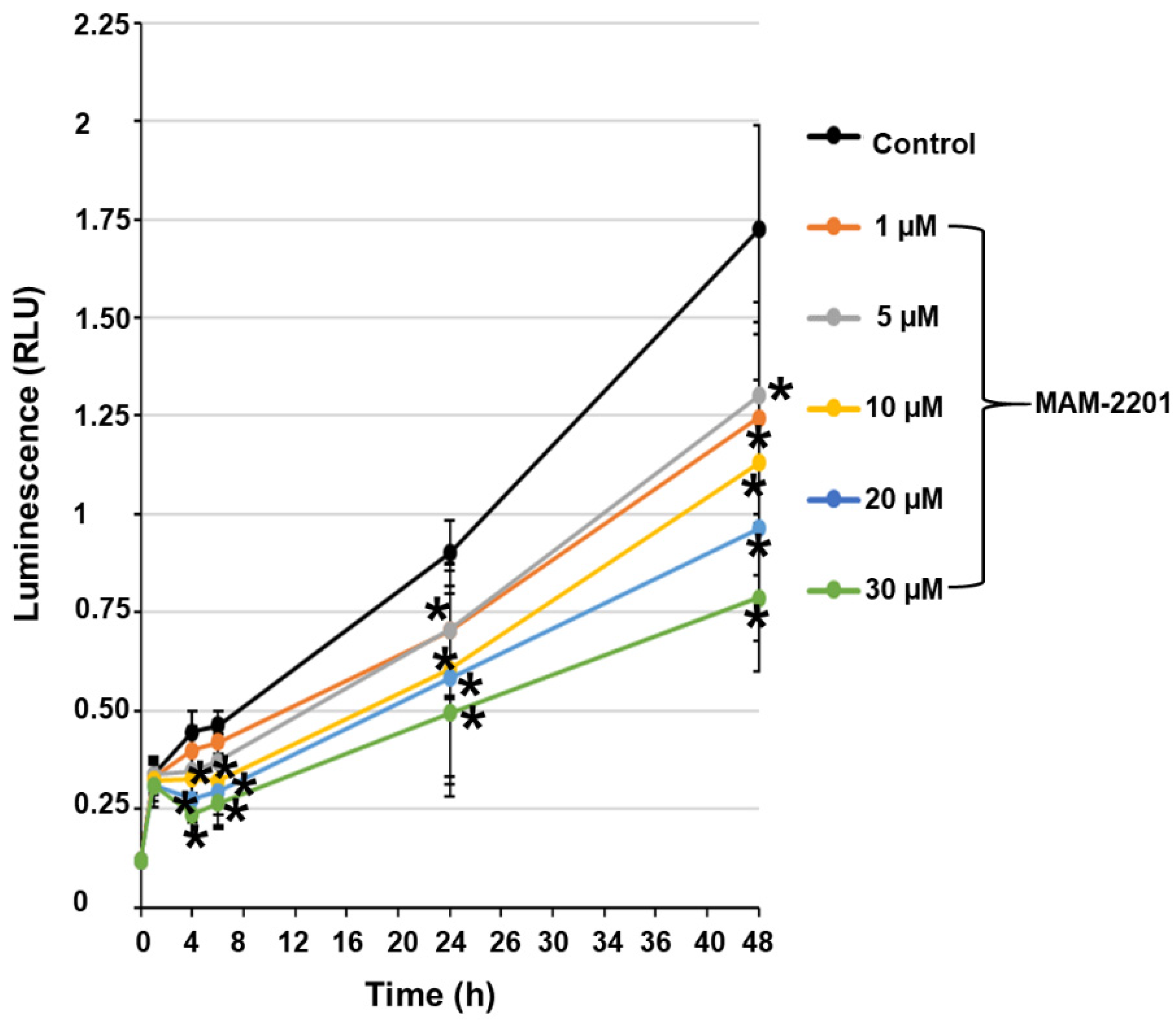
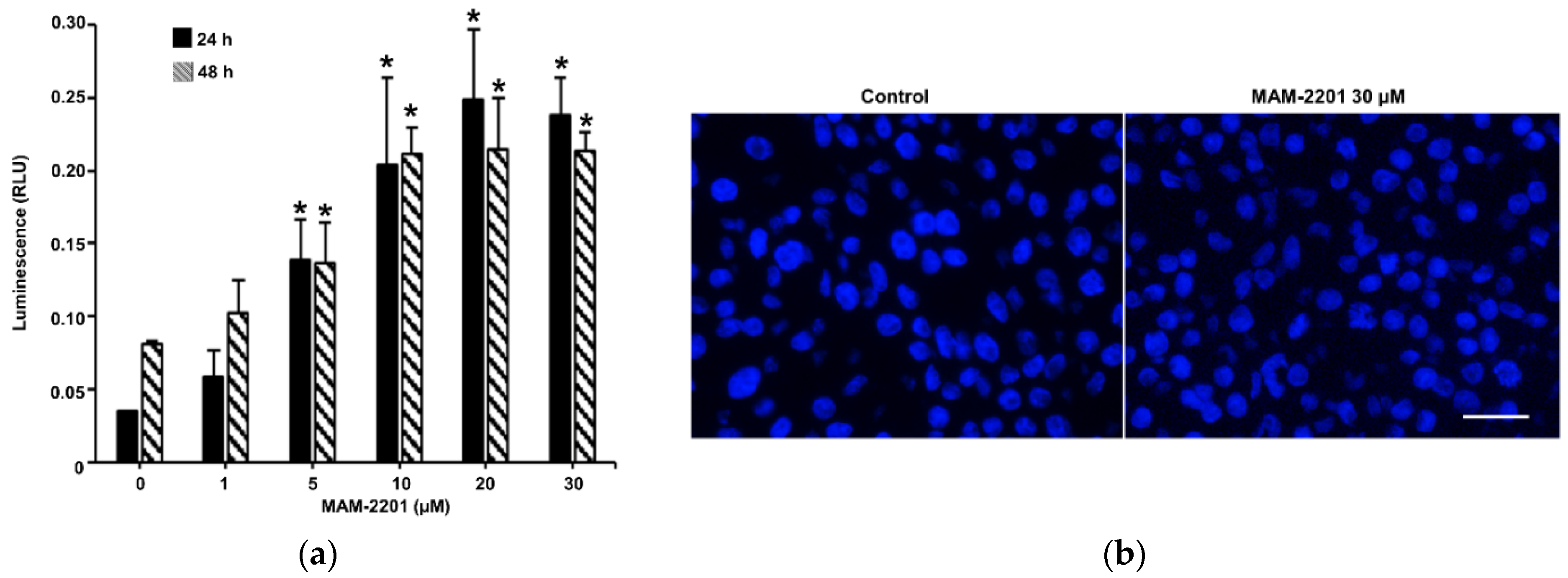
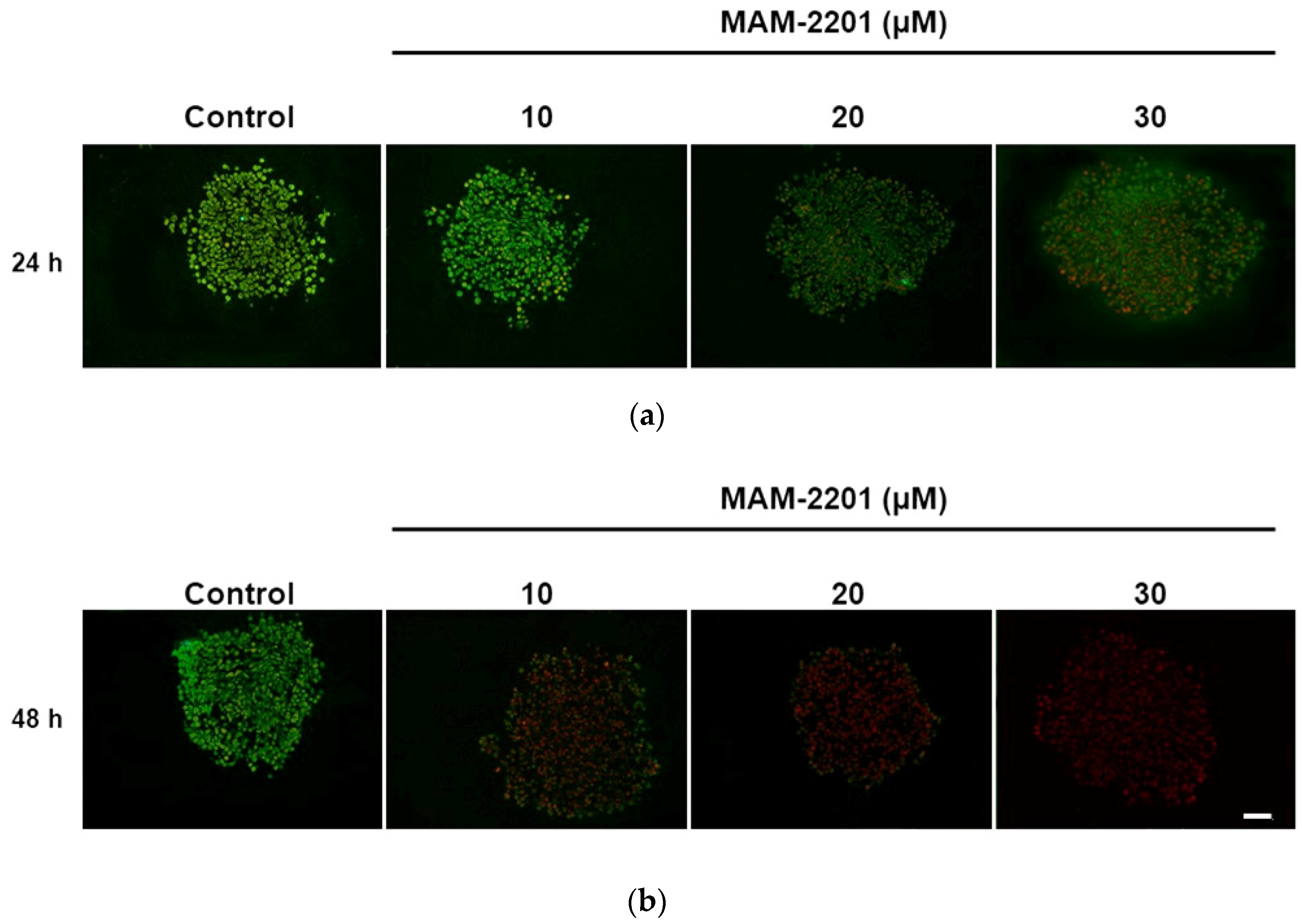
2.3. Evaluation of Caspase-3/7 Activity in Astrocyte Spheroids after MAM-2201 Exposure
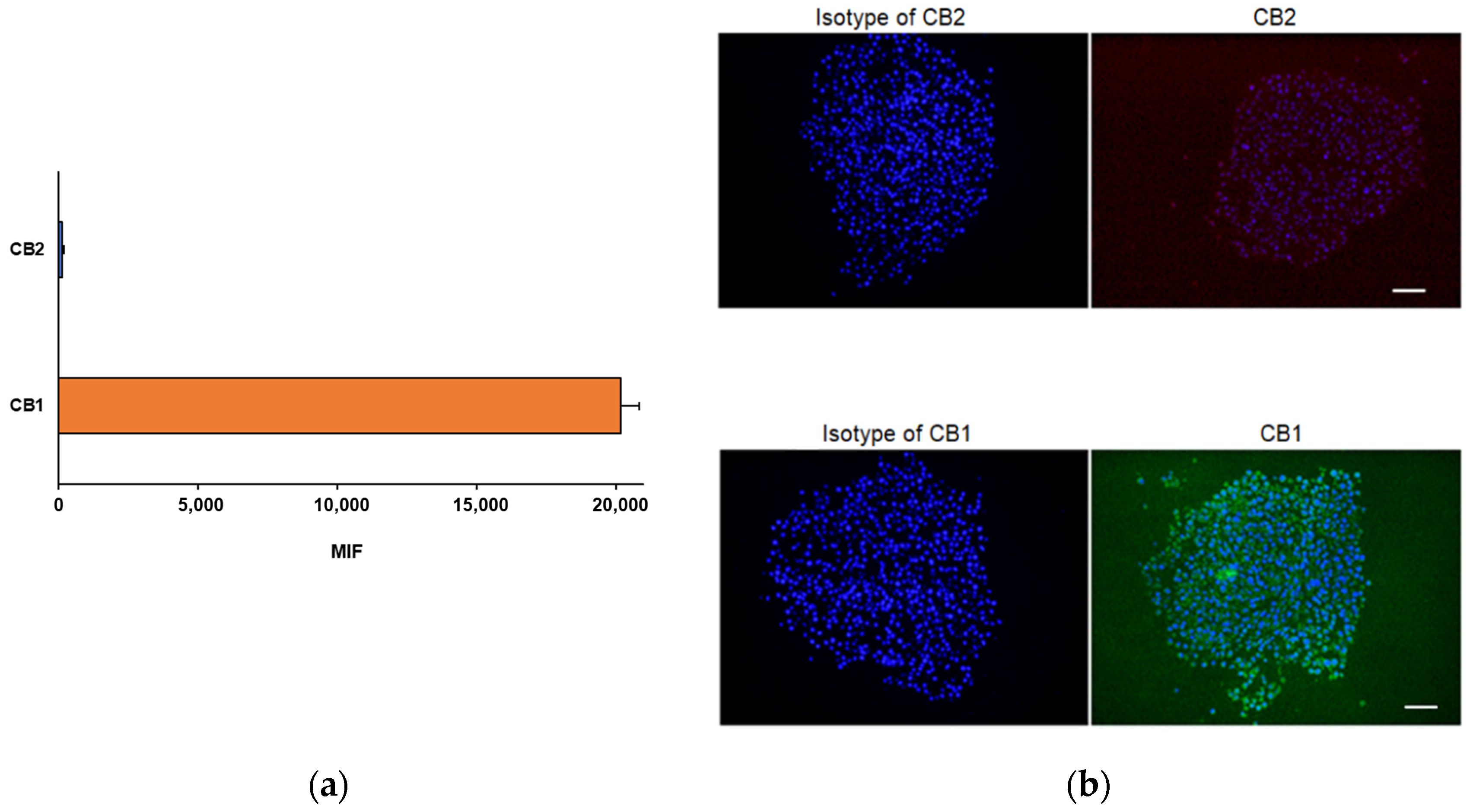
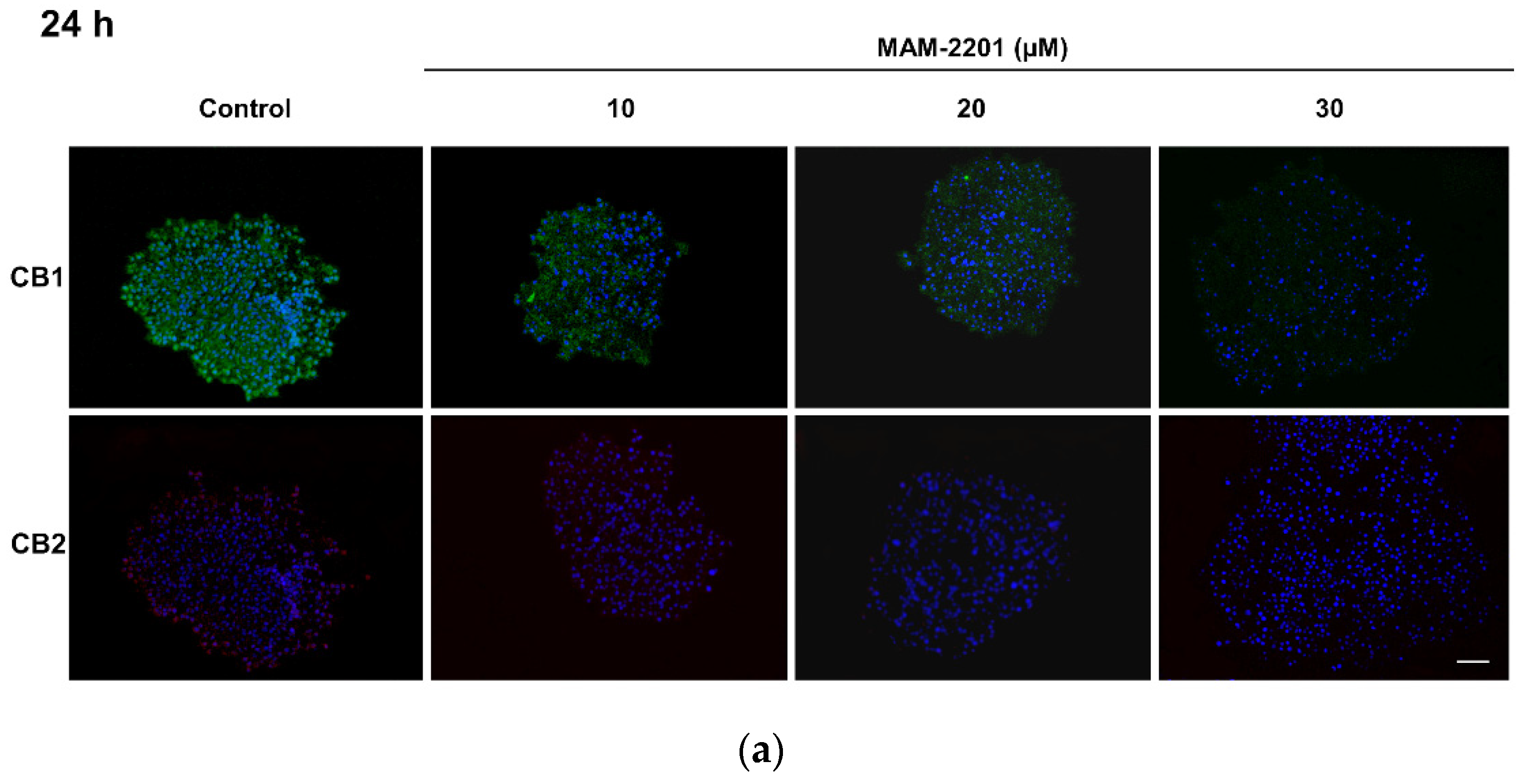
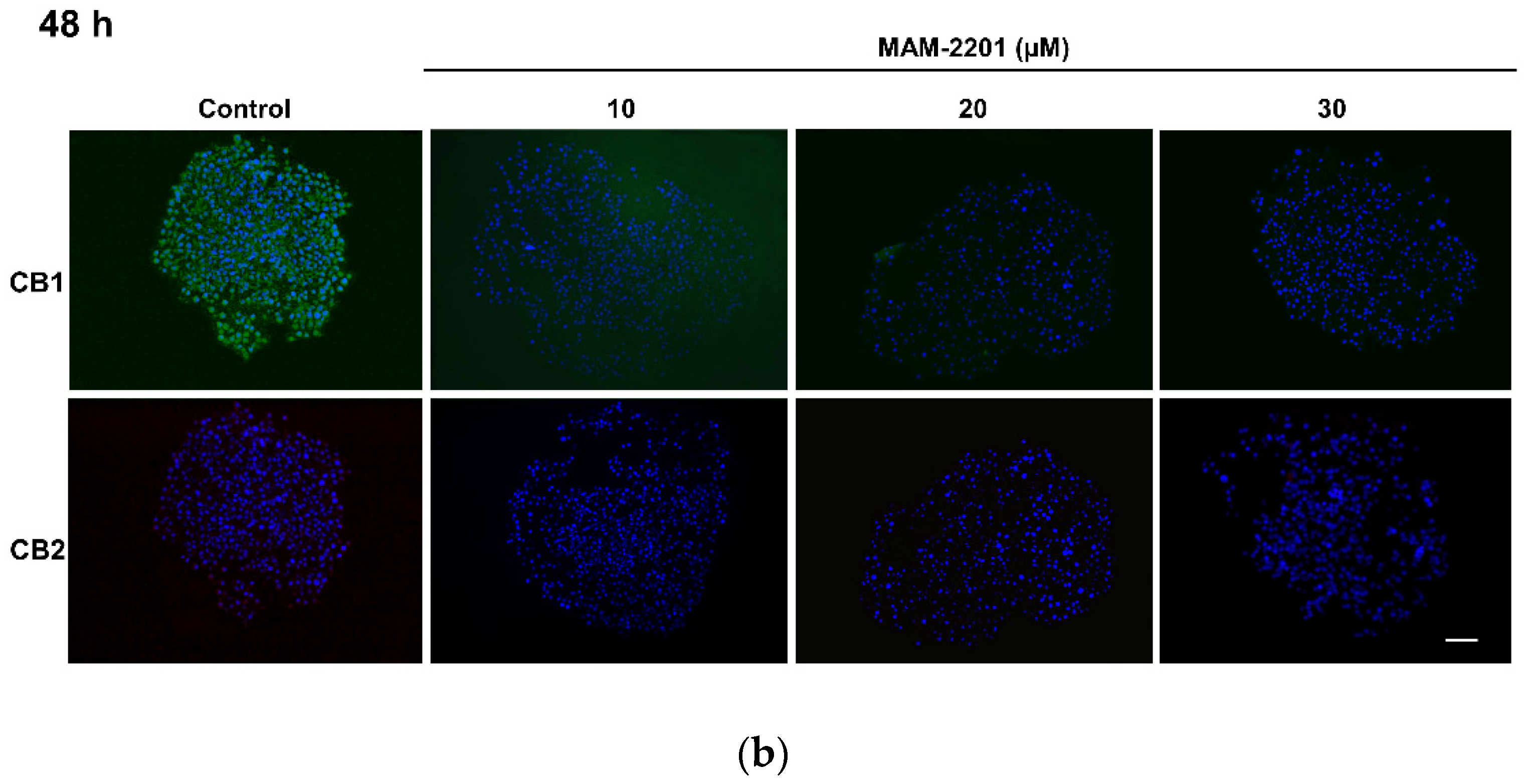
2.4. Evaluation of CB1 and CB2 Receptors
2.5. Effects on GFAP
3. Discussion
4. Materials and Methods
4.1. Culture Reagents
4.2. Drug and Reagents
4.3. Astrocyte Spheroid Model
4.4. Treatment with MAM-2201
4.5. Cell Morphology Analysis by Phase-Contrast Microscopy
4.6. Frozen Sections of Astrocyte Spheroids for Histochemistry and Immunofluorescence Analyses
4.6.1. Hematoxylin and Eosin (H&E) Staining
4.6.2. Masson’s Trichrome Staining
4.7. E-Cadherin Biochemical Evaluation
4.8. Expression of CB Receptors and GFAP by Immunofluorescence Analysis
4.9. Detection of CB Receptor Expression by Flow Cytometry Analysis
4.10. Real-Time-Glo MT Cell Viability Assay
4.11. Cell Viability Assay by Trypan Blue Exclusion Test (TB)
4.12. Caspase-3/7 Activity
4.13. Weight, Diameter, and Mass Density Evaluation by W8 Physical Cytometry
4.14. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EMCDDA. European Drug Report 2019: Trends and Developments. Publications Office of the European Union, Luxembourg. Available online: http://www.emcdda.europa.eu/system/files/publications/11364/20191724_TDAT19001ENNPDF.pdf (accessed on 2 December 2022).
- EMCDDA. European Drug Report 2022: Trends and Developments. Available online: https://www.emcdda.europa.eu/publications/edr/trends-developments/2022_en (accessed on 2 December 2022).
- Auwärter, V.; Dresen, S.; Weinmann, W.; Müller, M.; Pütz, M.; Ferreirós, N. ‘Spice’ and other herbal blends: Harmless incense or cannabinoid designer drugs? J. Mass Spectrom. 2009, 44, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, N.; Kikura-Hanajiri, R.; Ogata, J.; Goda, Y. Chemical analysis of synthetic cannabinoids as designer drugs in herbal products. Forensic Sci. Int. 2010, 198, 31–38. [Google Scholar] [CrossRef]
- Uchiyama, N.; Kawamura, M.; Kikura-Hanajiri, R.; Goda, Y. Identification and quantitation of two cannabimimetic phenylacetylindoles JWH-251 and JWH-250, and four cannabimimetic naphthoylindoles JWH-081, JWH-015, JWH-200, and JWH-073 as designer drugs in illegal products. Forensic Toxicol. 2011, 29, 25–37. [Google Scholar] [CrossRef]
- Ossato, A.; Canazza, I.; Trapella, C.; Vincenzi, F.; De Luca, M.A.; Rimondo, C.; Varani, K.; Borea, P.A.; Serpelloni, G.; Marti, M. Effect of JWH-250, JWH-073 and their interaction on “tetrad”, sensorimotor, neurological and neurochemical responses in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 67, 31–50. [Google Scholar] [CrossRef] [PubMed]
- Canazza, I.; Ossato, A.; Vincenzi, F.; Gregori, A.; Di Rosa, F.; Nigro, F.; Rimessi, A.; Pinton, P.; Varani, K.; Borea, P.A.; et al. Pharmaco-toxicological effects of the novel third-generation fluorinate synthetic cannabinoids, 5F-ADBINACA, AB-FUBINACA, and STS-135 in mice. In vitro and in vivo studies. Hum. Psychopharmacol. 2017, 32, e2601. [Google Scholar] [CrossRef] [Green Version]
- Seely, K.A.; Brents, L.K.; Radominska-Pandya, A.; Endres, G.W.; Keyes, G.S.; Moran, J.H.; Prather, P.L. A major glucuronidated metabolite of JWH-018 is a neutral antagonist at CB1 receptors. Chem. Res. Toxicol. 2012, 25, 825–827. [Google Scholar] [CrossRef] [Green Version]
- Van Amsterdam, J.; Brunt, T.; Van Den Brink, W. The adverse health effects of synthetic cannabinoids with emphasis on psychosis-like effects. J. Psychopharmacol. 2015, 29, 254–263. [Google Scholar] [CrossRef]
- Castaneto, M.S.; Gorelick, D.A.; Desrosiers, N.A.; Hartman, R.L.; Pirard, S.; Huestis, M.A. Synthetic cannabinoids: Epidemiology, pharmacodynamics, and clinical implications. Drug Alcohol Depend. 2014, 144, 12–41. [Google Scholar] [CrossRef] [Green Version]
- Vigolo, A.; Ossato, A.; Trapella, C.; Vincenzi, F.; Rimondo, C.; Seri, C.; Varani, K.; Serpelloni, G.; Marti, M. Novel halogenated derivates of JWH-018: Behavioral and binding studies in mice. Neuropharmacology 2015, 95, 68–82. [Google Scholar] [CrossRef]
- Miliano, C.; Serpelloni, G.; Rimondo, C.; Mereu, M.; Marti, M.; De Luca, M.A. Neuropharmacology of new psychoactive substances (NPS): Focus on the rewarding and reinforcing properties of cannabimimetics and amphetamine-like stimulants. Front. Neurosci. 2016, 10, 153. [Google Scholar] [CrossRef]
- Sachdev, S.; Vemuri, K.; Banister, S.D.; Longworth, M.; Kassiou, M.; Santiago, M.; Makriyannis, A.; Connor, M. In vitro determination of the efficacy of illicit synthetic cannabinoids at CB1 receptors. Br. J. Pharmacol. 2019, 176, 4653–4665. [Google Scholar] [CrossRef] [PubMed]
- Giorgetti, A.; Busardò, F.P.; Tittarelli, R.; Auwärter, V.; Giorgetti, R. Post-mortem toxicology: A systematic review of death cases involving synthetic cannabinoid receptor agonists. Front. Psychiatry 2020, 11, 1–22. [Google Scholar] [CrossRef]
- Bilel, S.; Tirri, M.; Arfe, R.; Ossato, A.; Trapella, C.; Serpelloni, G.; Neri, M.; Fattore, L.; Marti, M. Novel halogenated synthetic cannabinoids impair sensorimotor functions in mice. Neurotoxicology 2020, 76, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, J.; Deng, H.; Gatley, S.J.; Makriyannis, A.; Akinfeleye, T.; Bruneus, M.; Dimaio, A.A.; Gifford, A.N. Evaluation of the in vivo receptor occupancy for the behavioral effects of cannabinoids using a radiolabeled cannabinoid receptor agonist, R-[125/131I]AM2233. Synapse 2006, 60, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Müller, H.; Sperling, W.; Köhrmann, M.; Huttner, H.B.; Kornhuber, J.; Maler, J.M. The synthetic cannabinoid Spice as a trigger for an acute exacerbation of cannabis induced recurrent psychotic episodes. Schizophr. Res. 2010, 118, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Lonati, D.; Buscaglia, E.; Papa, P.; Valli, A.; Coccini, T.; Giampreti, A.; Petrolini, V.M.; Vecchio, S.; Serpelloni, G.; Locatelli, C.A. MAM-2201 (analytically confirmed) intoxication after “synthacaine” consumption. Ann. Emerg. Med. 2014, 64, 629–632. [Google Scholar] [CrossRef]
- Hess, C.; Stockhausen, S.; Kernbach-Wighton, G.; Madea, B. Death due to diabetic ketoacidosis: Induction by the consumption of synthetic cannabinoids? Case Rep. Forensic Sci. Int. 2015, 257, e6–e11. [Google Scholar] [CrossRef]
- Wiley, J.L.; Marusich, J.A.; Lefever, T.W.; Antonazzo, K.R.; Wallgren, M.T.; Cortes, R.A.; Patel, P.R.; Grabenauer, M.; Moore, K.N.; Thomas, B.F. AB-CHMINACA, AB-PINACA, and FUBIMINA: Affinity and potency of novel synthetic cannabinoids in producing Δ9-Tetrahydrocannabinol-like effects in mice. J. Pharmacol. Exp. Ther. 2015, 354, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Wiley, J.L.; Lefever, T.W.; Marusich, J.A.; Grabenauer, M.; Moore, K.N.; Huffman, J.W.; Thomas, B.F. Evaluation of first generation synthetic cannabinoids on binding at non-cannabinoid receptors and in a battery of in vivo assays in mice. Neuropharmacology 2016, 110 Pt A, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Namera, A.; Miura, N.; Ohta, S.; Miyazaki, S.; Osawa, M.; Inokuchi, S.A. A fatal case of MAM-2201 poisoning. Forensic Toxicol. 2013, 31, 333–337. [Google Scholar] [CrossRef]
- Schaefer, N.; Peters, B.; Bregel, D.; Kneisel, S.; Auwärter, V.; Schmidt, P.H.; Ewald, A.H. A fatal case involving several synthetic cannabinoids. Toxichem. Krimtech 2013, 80, 248–251. [Google Scholar]
- Tomiyama, K.; Funada, M. Cytotoxicity of synthetic cannabinoids found in “Spice” products: The role of cannabinoid receptors and the caspase cascade in the NG 108-15 cell line. Toxicol. Lett. 2011, 207, 12–17. [Google Scholar] [CrossRef]
- Tomiyama, K.; Funada, M. Cytotoxicity of synthetic cannabinoids on primary neuronal cells of the forebrain: The involvement of cannabinoid CB1 receptors and apoptotic cell death. Toxicol. Appl. Pharmacol. 2014, 274, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Koller, V.J.; Auwärter, V.; Grummt, T.; Moosmann, B.; Mišík, M.; Knasmüller, S. Investigation of the in vitro toxicological properties of the synthetic cannabimimetic drug CP-47,497-C8. Toxicol. Appl. Pharmacol. 2014, 277, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Costain, W.J.; Tauskela, J.S.; Rasquinha, I.; Comas, T.; Hewitt, M.; Marleau, V.; Soo, E.C. Pharmacological characterization of emerging synthetic cannabinoids in HEK293T cells and hippocampal neurons. Eur. J. Pharmacol. 2016, 786, 234–245. [Google Scholar] [CrossRef]
- Irie, T.; Kikura-Hanajiri, R.; Usami, M.; Uchiyama, N.; Goda, Y.; Sekino, Y. MAM-2201, a synthetic cannabinoid drug of abuse, suppresses the synaptic input to cerebellar Purkinje cells via activation of presynaptic CB1 receptors. Neuropharmacology 2015, 95, 479–491. [Google Scholar] [CrossRef]
- Tauskela, J.S.; Comas, T.; Hewitt, M.; Aylsworth, A.; Zhao, X.; Martina, M.; Costain, W.J. Effect of synthetic cannabinoids on spontaneous neuronal activity: Evaluation using Ca2+ spiking and multi-electrode arrays. Eur. J. Pharmacol. 2016, 786, 148–160. [Google Scholar] [CrossRef] [Green Version]
- Zaitsu, K.; Hayashi, Y.; Suzuki, K.; Nakayama, H.; Hattori, N.; Takahara, R.; Kusano, M.; Tsuchihashi, H.; Ishii, A. Metabolome disruption of the rat cerebrum induced by the acute toxic effects of the synthetic cannabinoid MAM-2201. Life Sci. 2015, 137, 49–55. [Google Scholar] [CrossRef]
- Coccini, T.; De Simone, U.; Lonati, D.; Scaravaggi, G.; Marti, M.; Locatelli, C.A. MAM-2201, one of the most potent-naphthoyl indole derivative-synthetic cannabinoids, exerts toxic effects on human cell-based models of neurons and astrocytes. Neurotox. Res. 2021, 39, 1251–1273. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacology 2010, 35, 217–238. [Google Scholar] [CrossRef] [Green Version]
- Coller, J.K.; Hutchinson, M.R. Implications of central immune signaling caused by drugs of abuse: Mechanisms, mediators and new therapeutic approaches for prediction and treatment of drug dependence. Pharmacol. Ther. 2012, 134, 219–245. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Hidalgo, J.J. The role of glial cells in drug abuse. Curr. Drug Abuse Rev. 2009, 2, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matias, I.; Morgado, J.; Gomes, F.C.A. Astrocyte heterogeneity: Impact to brain aging and disease. Front. Aging Neurosci. 2019, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Molecular dissection of reactive astrogliosis and glial scar formation. Trends Neurosci. 2009, 32, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacagnina, M.J.; Rivera, P.D.; Bilbo, S.D. Glial and neuroimmune mechanisms as critical modulators of drug use and abuse. Neuropsychopharmacology 2017, 42, 156–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozela, E.; Juknat, A.; Vogel, Z. Modulation of astrocyte activity by cannabidiol, a nonpsychoactive cannabinoid. Int. J. Mol. Sci. 2017, 18, 1669. [Google Scholar] [CrossRef] [Green Version]
- di Giacomo, V.; Chiavaroli, A.; Recinella, L.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Ronci, M.; Leone, S.; Brunetti, L.; et al. Antioxidant and neuroprotective effects induced by cannabidiol and cannabigerol in rat CTX-TNA2 astrocytes and isolated cortexes. Int. J. Mol. Sci. 2020, 21, 3575. [Google Scholar] [CrossRef]
- Huestis, M.A.; Gorelick, D.A.; Heishman, S.J.; Preston, K.L.; Nelson, R.A.; Moolchan, E.T.; Frank, R.A. Blockade of effects of smoked marijuana by the CBl-selective cannabinoid receptor antagonist SR141716. Arch. Gen. Psychiatry 2001, 58, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Stella, N. Cannabinoid and cannabinoid-like receptors in microglia, astrocytes and astrocytomas. Glia 2010, 58, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Landucci, E.; Mazzantini, C.; Lana, D.; Giovannini, M.G.; Pellegrini-Giampietro, D.E. Neuronal and astrocytic morphological alterations driven by prolonged exposure with δ9-tetrahydrocannabinol but not cannabidiol. Toxics 2022, 10, 48. [Google Scholar] [CrossRef]
- Andrade-Talavera, Y.; Duque-Feria, P.; Paulsen, O.; Rodríguez-Moreno, A. Presynaptic spike timing-dependent long-term depression in the mouse hippocampus. Cereb. Cortex 2016, 26, 3637–3654. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Kesner, P.; Metna-Laurent, M.; Duan, T.; Xu, L.; Georges, F.; Koehl, M.; Abrous, D.N.; Mendizabal-Zubiaga, J.; Grandes, P.; et al. Acute cannabinoids impair working memory through astroglial CB1 receptor modulation of hippocampal LTD. Cell 2012, 148, 1039–1050. [Google Scholar] [CrossRef] [Green Version]
- Min, R.; Nevian, T. Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nat. Neurosci. 2012, 15, 746–753. [Google Scholar] [CrossRef]
- Navarrete, M.; Araque, A. Endocannabinoids mediate neuron-astrocyte communication. Neuron 2008, 57, 883–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasooli-Nejad, S.; Palygin, O.; Lalo, U.; Pankratov, Y. Cannabinoid receptors contribute to astroglial Ca²⁺-signalling and control of synaptic plasticity in the neocortex. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20140077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Rodríguez, A.; Bonilla-Del Río, I.; Puente, N.; Gómez-Urquijo, S.M.; Fontaine, C.J.; Egaña-Huguet, J.; Elezgarai, I.; Ruehle, S.; Lutz, B.; Robin, L.M.; et al. Localization of the cannabinoid type-1 receptor in subcellular astrocyte compartments of mutant mouse hippocampus. Glia 2018, 66, 1417–1431. [Google Scholar] [CrossRef] [Green Version]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.R.; Robitaille, R.; Volterra, A. Gliotransmitters travel in time and space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite synapses: Astrocytes process and control synaptic information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Robin, L.M.; Oliveira da Cruz, J.F.; Langlais, V.C.; Martin-Fernandez, M.; Metna-Laurent, M.; Busquets-Garcia, A.; Bellocchio, L.; Soria-Gomez, E.; Papouin, T.; Varilh, M.; et al. Astroglial CB1 receptors determine synaptic d-serine availability to enable recognition memory. Neuron 2018, 98, 935–944.e5. [Google Scholar] [CrossRef] [Green Version]
- Bender, C.L.; Calfa, G.D.; Molina, V.A. Astrocyte plasticity induced by emotional stress: A new partner in psychiatric physiopathology? Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 65, 68–77. [Google Scholar] [CrossRef]
- Bender, C.L.; Sun, X.; Farooq, M.; Yang, Q.; Davison, C.; Maroteaux, M.; Huang, Y.S.; Ishikawa, Y. Emotional stress induces structural plasticity in bergmann glial cells via an AC5-CPEB3-GluA1 pathway. J. Neurosci. 2020, 40, 3374–3384. [Google Scholar] [CrossRef] [PubMed]
- Codeluppi, S.A.; Chatterjee, D.; Prevot, T.D.; Bansal, Y.; Misquitta, K.A.; Sibille, E.; Banasr, M. Chronic stress alters astrocyte morphology in mouse prefrontal cortex. Int. J. Neuropsychopharmacol. 2021, 24, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Jorfi, M.; D’Avanzo, C.; Kim, D.Y.; Irimia, D. Three-dimensional models of the human brain development and diseases. Adv. Healthc. Mater. 2018, 7, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, S.N.; Xu, T.Y.; Miao, Z.W.; Su, D.F.; Miao, C.Y. Organoid technology for brain and therapeutics research. CNS Neurosci. Ther. 2017, 23, 771–778. [Google Scholar] [CrossRef]
- Kosheleva, N.V.; Efremov, Y.M.; Shavkuta, B.S.; Zurina, I.M.; Zhang, D.; Minaev, N.V.; Gorkun, A.A.; Wei, S.; Shpichka, A.I.; Saburina, I.N.; et al. Cell spheroid fusion: Beyond liquid drops model. Sci. Rep. 2020, 10, 12614. [Google Scholar] [CrossRef]
- Leite, P.E.C.; Pereira, M.R.; Harris, G.; Pamies, D.; Gobbo Dos Santos, L.M.; Granjeiro, J.M.; Hogberg, H.T.; Hartung, T.; Smirnova, L. Suitability of 3D human brain spheroid models to distinguish toxic effects of gold and poly-lactic acid nanoparticles to assess biocompatibility for brain drug delivery. Part. Fibre Toxicol. 2019, 16, 22. [Google Scholar] [CrossRef] [Green Version]
- Goodman, T.T.; Ng, C.P.; Pun, S.H. 3-D tissue culture systems for the evaluation and optimization of nanoparticle-based drug carriers. Bioconjug. Chem. 2008, 19, 1951–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoelting, L.; Scheinhardt, B.; Bondarenko, O.; Schildknecht, S.; Kapitza, M.; Tanavde, V.; Tan, B.; Lee, Q.Y.; Mecking, S.; Leist, M.; et al. A 3-dimensional human embryonic stem cell (hESC)-derived model to detect developmental neurotoxicity of nanoparticles. Arch. Toxicol. 2013, 87, 721–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langhans, S.A. Three-Dimensional in vitro cell culture models in drug discovery and drug repositioning. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef]
- Lee, J.; Lilly, G.D.; Doty, R.C.; Podsiadlo, P.; Kotov, N.A. In vitro toxicity testing of nanoparticles in 3D cell culture. Small 2009, 5, 1213–1221. Available online: http://www.ncbi.nlm.nih.gov/pubmed/19263430 (accessed on 2 December 2022). [CrossRef] [PubMed]
- Onuwaje, I.; Phillips, J.B. Chapter 16: Three-dimensional culture systems in central nervous system research. In Handbook of Innovations in Central Nervous System Regenerative Medicine; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 571–601. [Google Scholar] [CrossRef]
- Sreekanthreddy, P.; Gromnicova, R.; Davies, H.; Phillips, J.; Romero, I.A.; Male, D. A three-dimensional model of the human blood-brain barrier to analyse the transport of nanoparticles and astrocyte/endothelial interactions. F1000Research 2015, 4, 1279. Available online: http://www.ncbi.nlm.nih.gov/pubmed/26870320 (accessed on 2 December 2022). [CrossRef] [PubMed]
- Zeng, Y.; Kurokawa, Y.; Zeng, Q.; Win-Shwe, T.T.; Nansai, H.; Zhang, Z.; Sone, H. Effects of Polyamidoamine dendrimers on a 3-D Neurosphere system using human neural progenitor cells. Toxicol. Sci. 2016, 152, 128–144. Available online: http://www.ncbi.nlm.nih.gov/pubmed/27125967 (accessed on 2 December 2022). [CrossRef] [Green Version]
- Smyrek, I.; Mathew, B.; Fischer, S.C.; Lissek, S.M.; Becker, S.; Stelzer, E.H.K. E-cadherin, actin, microtubules and FAK dominate different spheroid formation phases and important elements of tissue integrity. Biol. Open 2019, 8, bio037051. [Google Scholar] [CrossRef] [Green Version]
- De Simone, U.; Roccio, M.; Gribaldo, L.; Spinillo, A.; Caloni, F.; Coccini, T. Human 3D cultures as models for evaluating magnetic nanoparticle CNS cytotoxicity after short- and repeated long-term exposure. Int. J. Mol. Sci. 2018, 19, 1993. [Google Scholar] [CrossRef] [Green Version]
- Tsaousis, K.T.; Kopsachilis, N.; Tsinopoulos, I.T.; Dimitrakos, S.A.; Kruse, F.E.; Welge-Luessen, U. Time-dependent morphological alterations and viability of cultured human trabecular cells after exposure to trypan blue. Clin. Exp. Ophthalmol. 2013, 41, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Vu, B.; Souza, G.R.; Dengjel, J. Scaffold-free 3D cell culture of primary skin fibroblasts induces profound changes of the matrisome. Matrix Biol. Plus 2021, 11, 100066. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, W.; Wang, J.; Zhao, M.; Shen, K.; Li, Y.; Zhang, J.; Cai, W.; Xiao, D.; Bai, X.; et al. A novel 3D culture model of human ASCS reduces cell death in spheroid cores and maintains inner cell proliferation compared with a nonadherent 3D culture. Front. Cell Dev. Biol. 2021, 9, 737275. [Google Scholar] [CrossRef]
- Han, S.J.; Kwon, S.; Kim, K.S. Challenges of applying multicellular tumor spheroids in preclinical phase. Cancer Cell Int. 2021, 21, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Stenzel, M.H. Multicellular tumor spheroids (MCTS) as a 3D in vitro evaluation tool of nanoparticles. Small 2018, 14, e1702858. [Google Scholar] [CrossRef]
- Mendonsa, A.M.; Na, T.Y.; Gumbiner, B.M. E-cadherin in contact inhibition and cancer. Oncogene 2018, 37, 4769–4780. [Google Scholar] [CrossRef]
- Li, L.T.; Zhao, F.F.; Jia, Z.M.; Qi, L.Q.; Zhang, X.Z.; Zhang, L.; Li, Y.Y.; Yang, J.J.; Wang, S.J.; Lin, H.; et al. Cannabinoid receptors promote chronic intermittent hypoxia-induced breast cancer metastasis via IGF-1R/AKT/GSK-3β. Mol. Ther. Oncolytics 2021, 23, 220–230. [Google Scholar] [CrossRef]
- Chen, A.; Beetham, H.; Black, M.A.; Priya, R.; Telford, B.J.; Guest, J.; Wiggins, G.A.R.; Godwin, T.D.; Yap, A.S.; Guilford, P.J. E-cadherin loss alters cytoskeletal organization and adhesion in non-malignant breast cells but is insufficient to induce an epithelial-mesenchymal transition. BMC Cancer 2014, 14, 552. [Google Scholar] [CrossRef] [Green Version]
- Lei, B.; Zhou, X.; Lv, D.; Wan, B.; Wu, H.; Zhong, L.; Shu, F.; Mao, X. Apoptotic and nonapoptotic function of caspase 7 in spermatogenesis. Asian J. Androl. 2017, 19, 47–51. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Festjens, N.; Declercq, W.; Vanden Berghe, T.; Vandenabeele, P. Caspases in cell survival, proliferation and differentiation. Cell Death Differ. 2007, 14, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Svandova, E.; Lesot, H.; Vanden Berghe, T.; Tucker, A.S.; Sharpe, P.T.; Vandenabeele, P.; Matalova, E. Non-apoptotic functions of caspase-7 during osteogenesis. Cell Death Dis. 2014, 5, e1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baena-Lopez, L.A.; Arthurton, L.; Xu, D.C.; Galasso, A. Non-apoptotic caspase regulation of stem cell properties. Semin. Cell Dev. Biol. 2018, 82, 118–126. [Google Scholar] [CrossRef]
- Hollville, E.; Deshmukh, M. Physiological functions of non-apoptotic caspase activity in the nervous system. Semin. Cell Dev. Biol. 2018, 82, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Acarin, L.; Villapol, S.; Faiz, M.; Rohn, T.T.; Castellano, B.; Gonzalez, B. Caspase-3 activation in astrocytes following postnatal excitotoxic damage correlates with cytoskeletal remodeling but not with cell death or proliferation. Glia 2007, 55, 954–965. [Google Scholar] [CrossRef]
- Li, Z.; Jo, J.; Jia, J.M.; Lo, S.C.; Whitcomb, D.J.; Jiao, S.; Cho, K.; Sheng, M. Caspase-3 activation via mitochondria is required for long-term depression and AMPA receptor internalization. Cell 2010, 141, 859–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burguillos, M.A.; Deierborg, T.; Kavanagh, E.; Persson, A.; Hajji, N.; Garcia-Quintanilla, A.; Cano, J.; Brundin, P.; Englund, E.; Venero, J.L.; et al. Caspase signalling controls microglia activation and neurotoxicity. Nature 2011, 472, 319–324. [Google Scholar] [CrossRef]
- Wagner, D.C.; Riegelsberger, U.M.; Michalk, S.; Hartig, W.; Kranz, A.; Boltze, J. Cleaved caspase-3 expression after experimental stroke exhibits different phenotypes and is predominantly non-apoptotic. Brain Res. 2011, 1381, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Guyenet, S.J.; Nguyen, H.T.; Hwang, B.H.; Schwartz, M.W.; Baskin, D.G.; Thaler, J.P. High-fat diet feeding causes rapid, non-apoptotic cleavage of caspase-3 in astrocytes. Brain Res. 2013, 1512, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Oliva, A.M.; García-Revilla, J.; Alonso-Bellido, I.M.; Burguillos, M.A. Brainiac caspases: Beyond the wall of apoptosis. Front. Cell. Neurosci. 2019, 13, 500. [Google Scholar] [CrossRef] [Green Version]
- Rohn, T.T.; Cusack, S.M.; Kessinger, S.R.; Oxford, J.T. Caspase activation independent of cell death is required for proper cell dispersal and correct morphology in PC12 cells. Exp. Cell Res. 2004, 295, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Oomman, S.; Strahlendorf, H.; Dertien, J.; Strahlendorf, J. Bergmann glia utilize active caspase-3 for differentiation. Brain Res. 2006, 1078, 19–34. [Google Scholar] [CrossRef]
- Aras, R.; Barron, A.M.; Pike, C.J. Caspase activation contributes to astrogliosis. Brain Res. 2012, 1450, 102–115. [Google Scholar] [CrossRef] [Green Version]
- Hulse, R.E.; Winterfield, J.; Kunkler, P.E.; Kraig, R.P. Astrocytic clasmatodendrosis in hippocampal organ culture. Glia 2001, 33, 169–179. [Google Scholar] [CrossRef]
- Burrows, B.T.; Watterson, L.R.; Egnatios, J.; Olive, M.F. Chapter 71—Synthetic cannabinoids: A summary of selected phenomena with respect to behavioral pharmacology and abuse liability. In Handbook of Cannabis and Related Pathologies: Biology, Pharmacology, Diagnosis, and Treatment; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 691–699. [Google Scholar] [CrossRef]
- Jouroukhin, Y.; Zhu, X.; Shevelkin, A.V.; Hasegawa, Y.; Abazyan, B.; Saito, A.; Pevsner, J.; Kamiya, A.; Pletnikov, M.V. Adolescent ∆9-Tetrahydrocannabinol exposure and astrocyte-specific genetic vulnerability converge on nuclear factor-κB-cyclooxygenase-2 signaling to impair memory in adulthood. Biol. Psychiatry 2019, 85, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Cong, J.; Lu, K.; Zou, W.; Li, Z.; Guo, Z.; Tong, X.; Zheng, J.; Zhu, J.; Li, S.; Zhang, W.; et al. Astroglial CB1 cannabinoid receptors mediate cp 55,940-induced conditioned place aversion through cyclooxygenase-2 signaling in mice. Front. Cell. Neurosci. 2021, 15, 772549. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Blasco, D.; Busquets-Garcia, A.; Hebert-Chatelain, E.; Serrat, R.; Vicente-Gutierrez, C.; Ioannidou, C.; Gómez-Sotres, P.; Lopez-Fabuel, I.; Resch-Beusher, M.; Resel, E. Glucose metabolism links astroglial mitochondria to cannabinoid effects. Nature 2020, 583, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhang, J.; Fan, N.; Teng, Z.Q.; Wu, Y.; Yang, H.; Tang, Y.P.; Sun, H.; Song, Y.; Chen, C. Δ9-THC-caused synaptic and memory impairments are mediated through COX-2 signaling. Cell 2013, 155, 1154–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadwick, B.; Miller, M.L.; Hurd, Y.L. Cannabis use during adolescent development: Susceptibility to psychiatric illness. Front. Psychiatry 2013, 4, 129. [Google Scholar] [CrossRef] [Green Version]
- Szutorisz, H.; Hurd, Y.L. High times for cannabis: Epigenetic imprint and its legacy on brain and behavior. Neurosci. Biobehav. Rev. 2018, 85, 93–101. [Google Scholar] [CrossRef]
- Broyd, S.J.; van Hell, H.H.; Beale, C.; Yucel, M.; Solowij, N. Acute and chronic effects of cannabinoids on human cognition-a systematic review. Biol. Psychiatry. 2016, 79, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Stella, N. Cannabinoid signaling in glial cells. Glia 2004, 48, 267–277. [Google Scholar] [CrossRef]
- Rubino, T.; Realini, N.; Braida, D.; Alberio, T.; Capurro, V.; Viganò, D.; Guidali, C.; Sala, M.; Fasano, M.; Parolaro, D. The depressive phenotype induced in adult female rats by adolescent exposure to THC is associated with cognitive impairment and altered neuroplasticity in the prefrontal cortex. Neurotox. Res. 2009, 15, 291–302. [Google Scholar] [CrossRef]
- Zamberletti, E.; Gabaglio, M.; Grilli, M.; Prini, P.; Catanese, A.; Pittaluga, A.; Marchi, M.; Rubino, T.; Parolaro, D. Long-term hippocampal glutamate synapse and astrocyte dysfunctions underlying the altered phenotype induced by adolescent THC treatment in male rats. Pharmacol. Res. 2016, 111, 459–470. [Google Scholar] [CrossRef]
- Zhou, Y.; Shao, A.; Yao, Y.; Tu, S.; Deng, Y.; Zhang, J. Dual roles of astrocytes in plasticity and reconstruction after traumatic brain injury. Cell. Commun. Signal. 2020, 18, 62. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, S.; Gharagozloo, M.; Simard, C.; Gris, D. Astrocytes maintain glutamate homeostasis in the CNS by controlling the balance between glutamate uptake and release. Cells 2019, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Middeldorp, J.; Hol, E. GFAP in health and disease. Prog. Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef] [PubMed]
- Jacobsson, S.O.P.; Wallin, T.; Fowler, C.J. Inhibition of rat C6 glioma cell proliferation by endogenous and synthetic cannabinoids. Relative involvement of cannabinoid and vanilloid receptors. J. Pharmacol. Exp. Ther. 2001, 299, 951–959. [Google Scholar] [PubMed]
- Balmforth, A.J.; Ball, S.G.; Freshney, R.I.; Graham, D.I.; McNamee, H.B.; Vaughan, P.F. D-1 dopaminergic and β-adrenergic stimulation of adenylate cyclase in a clone derived from the human astrocytoma cell line G-CCM. J. Neurochem. 1986. [Google Scholar] [CrossRef]
- Ulusoy, M.; Lavrentieva, A.; Walter, J.G.; Sambale, F.; Green, M.; Stahl, F.; Schepera, T. Evaluation of CdTe/CdS/ZnS core/shell/shell quantum dot toxicity on three-dimensional spheroid cultures. Toxicol. Res. 2016, 5, 126–135. [Google Scholar] [CrossRef]
- Cristaldi, D.A.; Sargenti, A.; Bonetti, S.; Musmeci, F.; Delprete, C.; Bacchi, F.; Pasqua, S.; Cavallo, C.; Bonsi, L.; Alviano, F.; et al. A reliable flow-based method for the accurate measure of mass density, size and weight of live 3D tumor spheroids. Micromachines 2020, 11, 465. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E-Cadherin (% of Control) in Cell Lysate | ||
|---|---|---|
| 24 h | 48 h | |
| MAM-2201 (µM) | ||
| 0 | 100.00 ± 2.72 | 100.00 ± 5.26 |
| 10 | 94.63 ± 8.82 | 100.70 ± 10.57 |
| 20 | 105.46 ± 2.16 | 72.47 ± 6.30 * |
| 30 | 102.64 ± 2.94 | 68.99 ± 5.53 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Simone, U.; Pignatti, P.; Villani, L.; Russo, L.A.; Sargenti, A.; Bonetti, S.; Buscaglia, E.; Coccini, T. Human Astrocyte Spheroids as Suitable In Vitro Screening Model to Evaluate Synthetic Cannabinoid MAM2201-Induced Effects on CNS. Int. J. Mol. Sci. 2023, 24, 1421. https://doi.org/10.3390/ijms24021421
De Simone U, Pignatti P, Villani L, Russo LA, Sargenti A, Bonetti S, Buscaglia E, Coccini T. Human Astrocyte Spheroids as Suitable In Vitro Screening Model to Evaluate Synthetic Cannabinoid MAM2201-Induced Effects on CNS. International Journal of Molecular Sciences. 2023; 24(2):1421. https://doi.org/10.3390/ijms24021421
Chicago/Turabian StyleDe Simone, Uliana, Patrizia Pignatti, Laura Villani, Luciana Alessandra Russo, Azzurra Sargenti, Simone Bonetti, Eleonora Buscaglia, and Teresa Coccini. 2023. "Human Astrocyte Spheroids as Suitable In Vitro Screening Model to Evaluate Synthetic Cannabinoid MAM2201-Induced Effects on CNS" International Journal of Molecular Sciences 24, no. 2: 1421. https://doi.org/10.3390/ijms24021421
APA StyleDe Simone, U., Pignatti, P., Villani, L., Russo, L. A., Sargenti, A., Bonetti, S., Buscaglia, E., & Coccini, T. (2023). Human Astrocyte Spheroids as Suitable In Vitro Screening Model to Evaluate Synthetic Cannabinoid MAM2201-Induced Effects on CNS. International Journal of Molecular Sciences, 24(2), 1421. https://doi.org/10.3390/ijms24021421